Polyamidoamine Dendrimer Conjugates with Cyclodextrins as Novel Carriers for DNA, shRNA and siRNA

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction

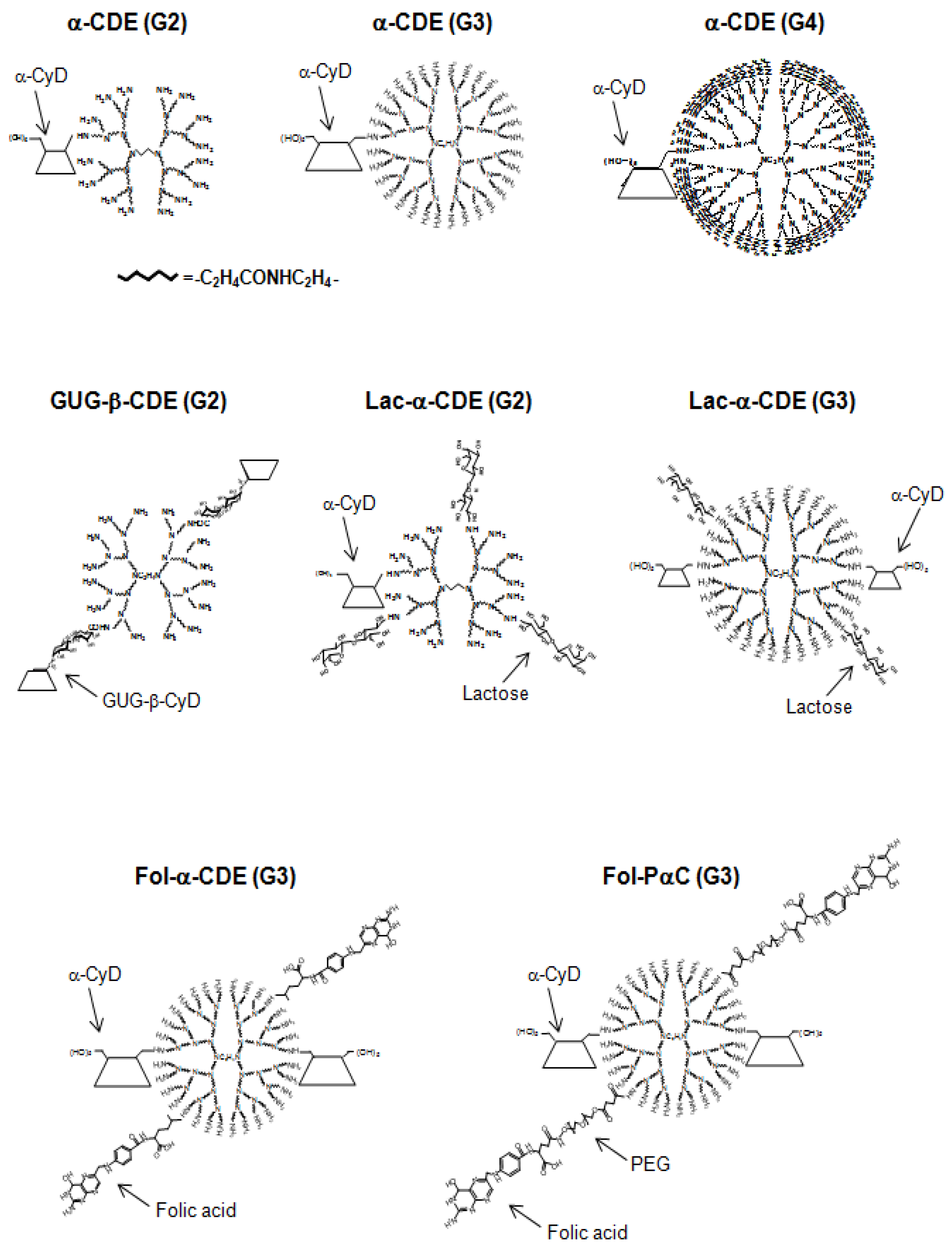
2. α-CyD Conjugates with Dendrimer (α-CDE) as pDNA Carriers

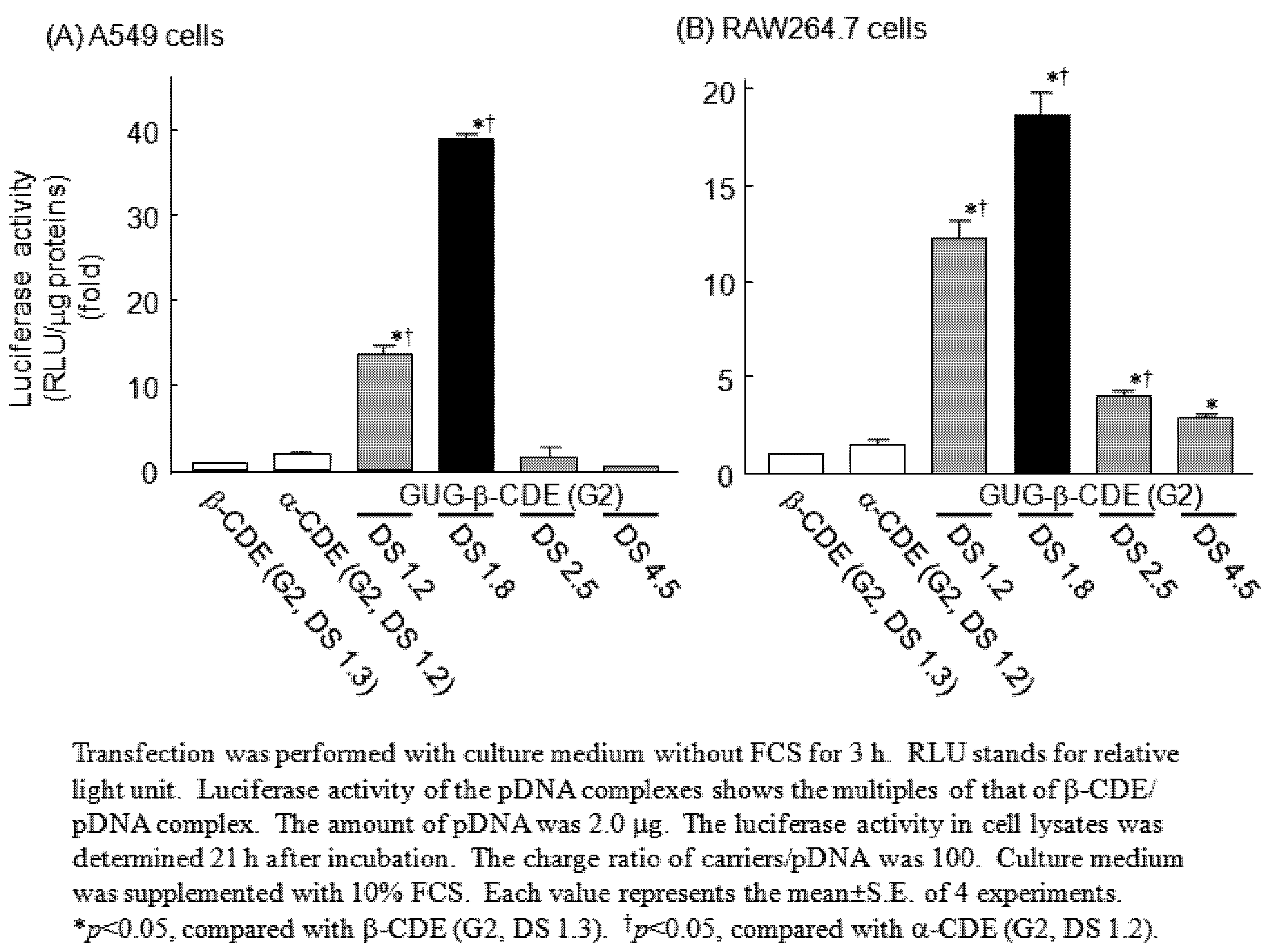
3. GUG-β-CDE (G2) as DNA Carriers

4. α-CDE (G3) as shRNA Carriers

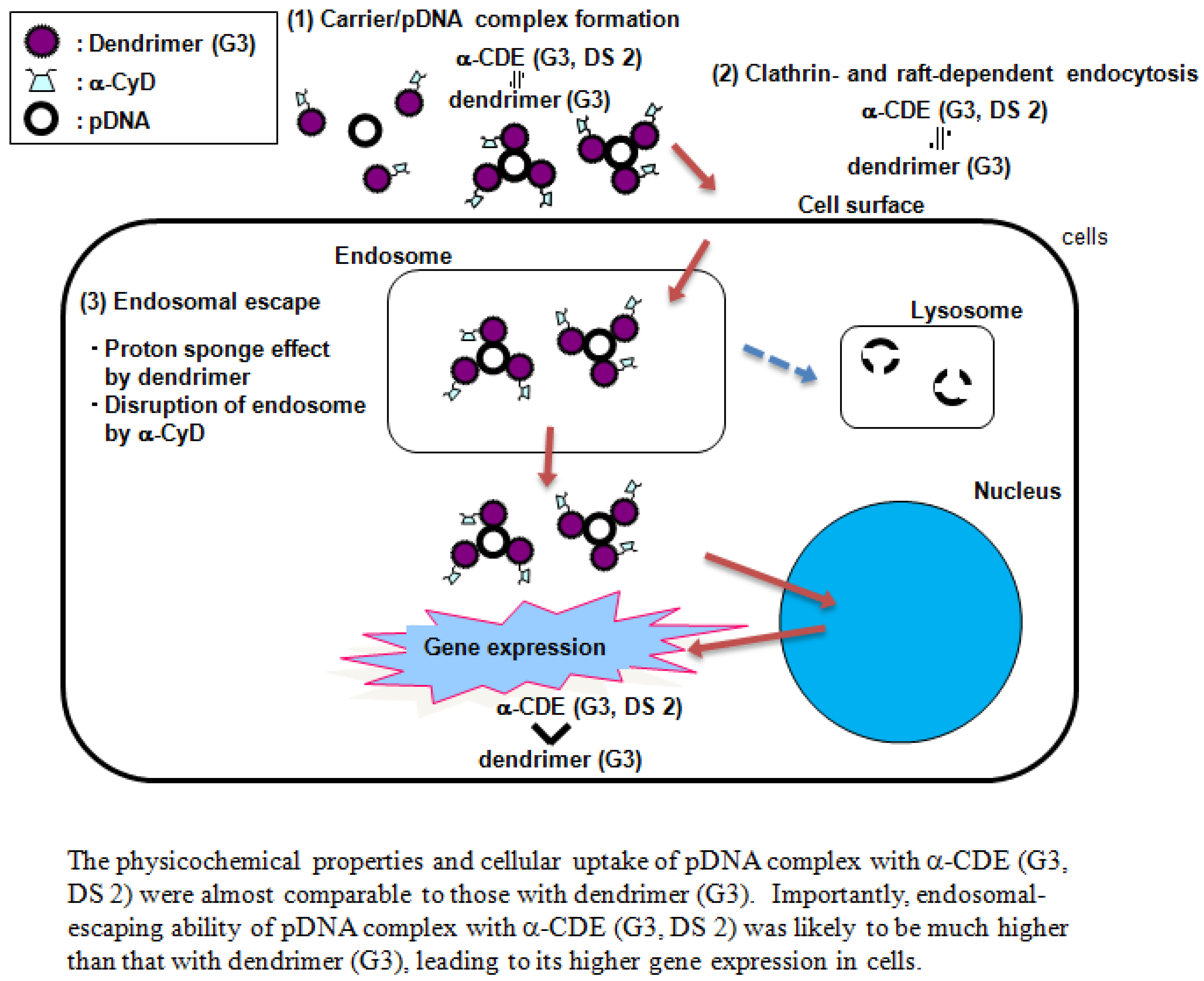
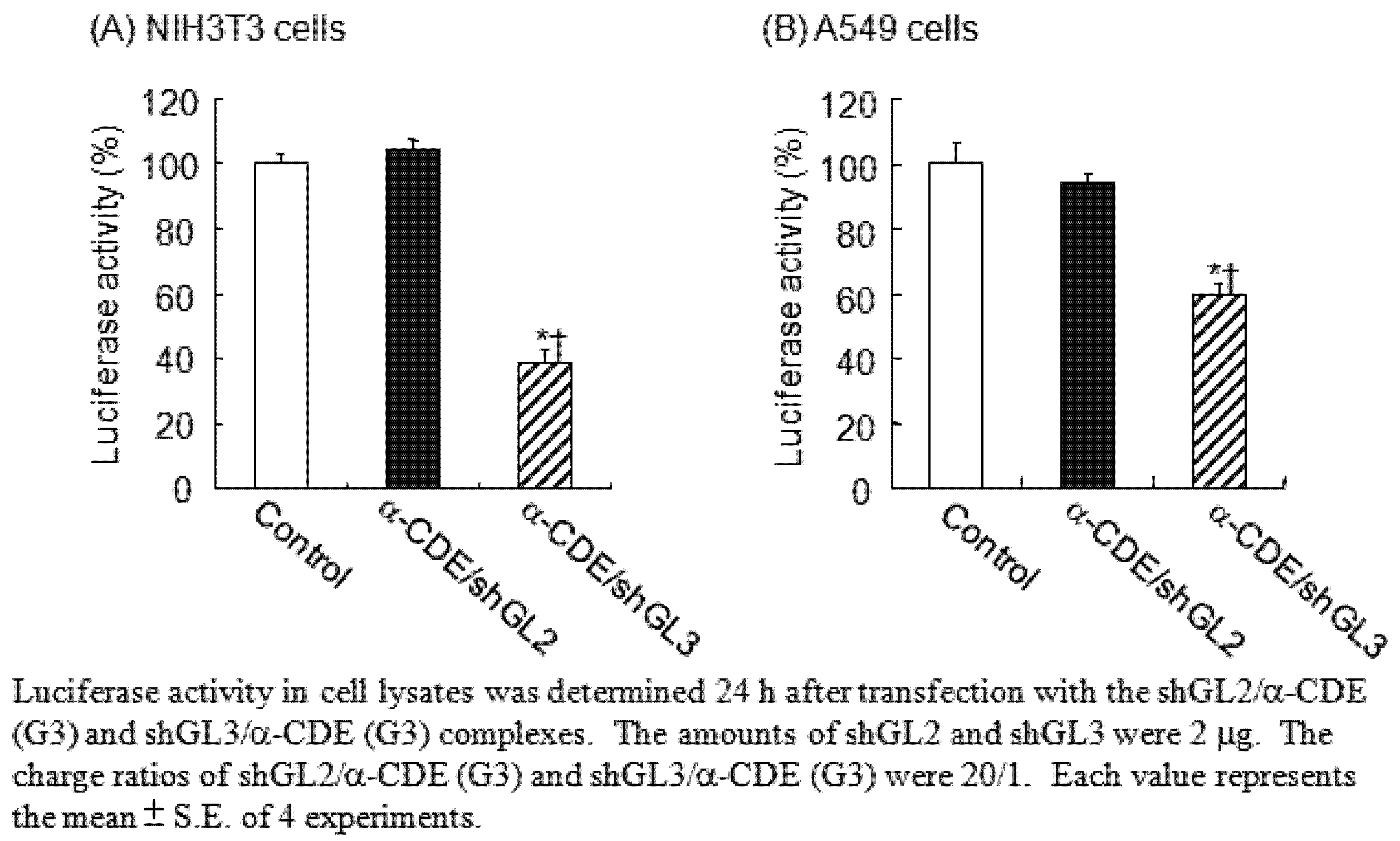
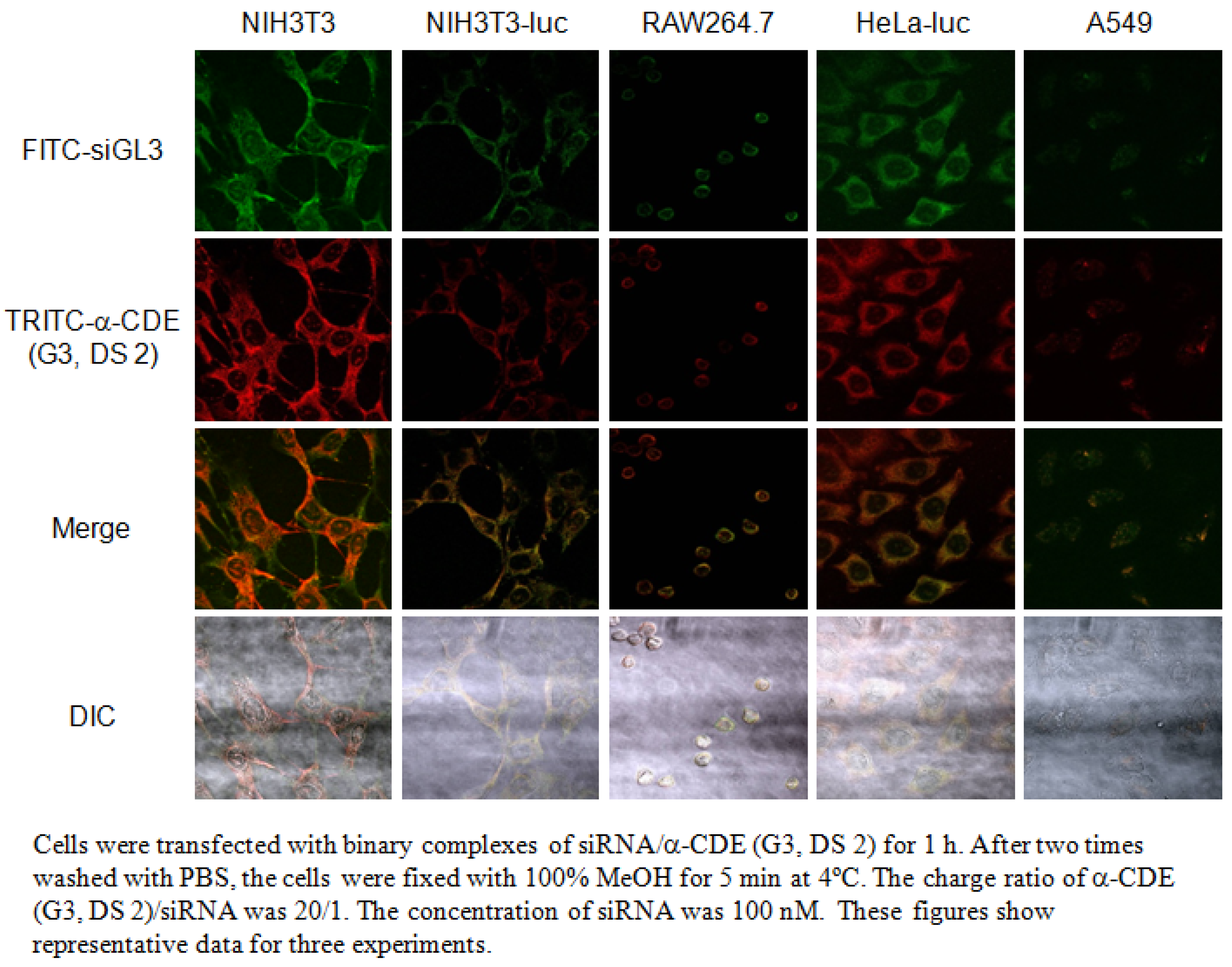
5. α-CDE (G3) as siRNA Carriers


6. Potential Use of Polypseudorotaxanes (PPRXs) of Pegylated Dendrimers (PEG-Dendrimers) with CyDs as the Novel Sustained Release Systems for pDNA
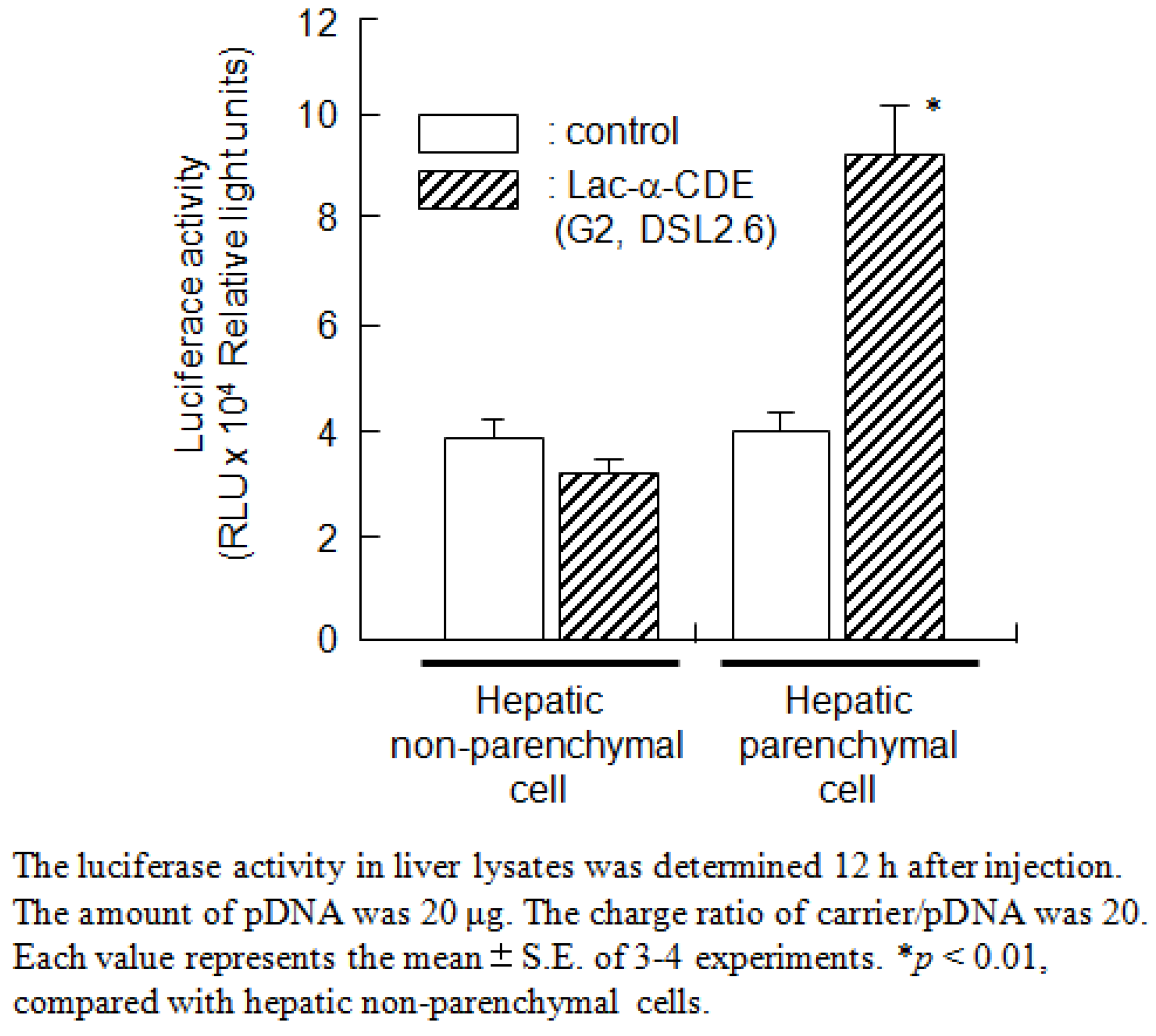
7. Functionalized α-CDEs (G2, G3) for Cell-Specific DNA Carriers



8. Conclusion
Acknowledgments
References
- Pfeifer, A.; Verma, I.M. Gene therapy: Promises and problems. Annu. Rev. Genomics Hum. Genet. 2001, 2, 177–211. [Google Scholar] [CrossRef]
- Elsabahy, M.; Nazarali, A.; Foldvari, M. Non-viral nucleic acid delivery: Key challenges and future directions. Curr. Drug Deliv. 2011, 8, 235–244. [Google Scholar]
- Lowenstein, P.R. Clinical trials in gene therapy: Ethics of informed consent and the future of experimental medicine. Curr. Opin. Mol. Ther. 2008, 10, 428–430. [Google Scholar]
- Fischer, A.; Cavazzana-Calvo, M. Gene therapy of inherited diseases. Lancet 2008, 371, 2044–2047. [Google Scholar] [CrossRef]
- Sliva, K.; Schnierle, B.S. Selective gene silencing by viral delivery of short hairpin RNA. Virol. J. 2010, 7. [Google Scholar]
- Raty, J.K.; Lesch, H.P.; Wirth, T.; Yla-Herttuala, S. Improving safety of gene therapy. Curr. Drug Saf. 2008, 3, 46–53. [Google Scholar] [CrossRef]
- Shen, Y. Advances in the development of siRNA-based therapeutics for cancer. IDrugs 2008, 11, 572–578. [Google Scholar]
- Tiera, M.J.; Shi, Q.; Winnik, F.M.; Fernandes, J.C. Polycation-based gene therapy: Current knowledge and new perspectives. Curr. Gene Ther. 2011, 11, 288–306. [Google Scholar] [CrossRef]
- Tomalia, D.A.; Baker, H.; Dewald, J.; Hall, M.; Kallos, G.; Martin, S.; Roek, J.; Ryder, J.; Smith, P. A new class of polymers: Starburt-dendritic macromolecules. Polym. J. 1985, 17, 117–132. [Google Scholar] [CrossRef]
- Challa, T.; Agaiah Goud, B.; Baskar, S.; Chandra Mouli, G.; Jukuri, R. Dendrimers: A novel polymer for drug delivery. Int. J. Pharm. Sci. Rev. Res. 2011, 9, 88–99. [Google Scholar]
- Dufes, C.; Uchegbu, I.F.; Schatzlein, A.G. Dendrimers in gene delivery. Adv. Drug Deliv. Rev. 2005, 57, 2177–2202. [Google Scholar] [CrossRef] [Green Version]
- Dutta, T.; Jain, N.K.; McMillan, N.A.; Parekh, H.S. Dendrimer nanocarriers as versatile vectors in gene delivery. Nanomedicine 2010, 6, 25–34. [Google Scholar] [CrossRef]
- Kim, J.B.; Choi, J.S.; Nam, K.; Lee, M.; Park, J.S.; Lee, J.K. Enhanced transfection of primary cortical cultures using arginine-grafted PAMAM dendrimer, PAMAM-Arg. J. Control. Release 2006, 114, 110–117. [Google Scholar] [CrossRef]
- Yuan, Q.; Lee, E.; Yeudall, W.A.; Yang, H. Dendrimer-triglycine-EGF nanoparticles for tumor imaging and targeted nucleic acid and drug delivery. Oral Oncol. 2010, 46, 698–704. [Google Scholar] [CrossRef]
- Liu, X.; Huang, H.; Wang, J.; Wang, C.; Wang, M.; Zhang, B.; Pan, C. Dendrimers-delivered short hairpin RNA targeting hTERT inhibits oral cancer cell growth in vitro and in vivo. Biochem. Pharmacol. 2011, 82, 17–23. [Google Scholar]
- Wang, J.; Lu, Z.; Wientjes, M.G.; Au, J.L. Delivery of siRNA therapeutics: Barriers and carriers. AAPS J. 2010, 12, 492–503. [Google Scholar] [CrossRef]
- Yuan, X.; Naguib, S.; Wu, Z. Recent advances of siRNA delivery by nanoparticles. Expert Opin. Drug Deliv. 2011, 8, 521–536. [Google Scholar] [CrossRef]
- Troiber, C.; Wagner, E. Nucleic acid carriers based on precise polymer conjugates. Bioconjug Chem. 2011, 22, 1737–1752. [Google Scholar]
- Ruponen, M.; Honkakoski, P.; Ronkko, S.; Pelkonen, J.; Tammi, M.; Urtti, A. Extracellular and intracellular barriers in non-viral gene delivery. J. Control. Release 2003, 93, 213–217. [Google Scholar] [CrossRef]
- Klajnert, B.; Bryszewska, M. Dendrimers: Properties and applications. Acta Biochim. Pol. 2001, 48, 199–208. [Google Scholar]
- Braun, C.S.; Vetro, J.A.; Tomalia, D.A.; Koe, G.S.; Koe, J.G.; Middaugh, C.R. Structure/function relationships of polyamidoamine/DNA dendrimers as gene delivery vehicles. J. Pharm. Sci. 2005, 94, 423–436. [Google Scholar] [CrossRef]
- Kukowska-Latallo, J.F.; Bielinska, A.U.; Johnson, J.; Spindler, R.; Tomalia, D.A.; Baker, J.R., Jr. Efficient transfer of genetic material into mammalian cells using Starburst polyamidoamine dendrimers. Proc. Natl. Acad. Sci. USA 1996, 93, 4897–4902. [Google Scholar]
- Morgan, D.M.; Larvin, V.L.; Pearson, J.D. Biochemical characterisation of polycation-induced cytotoxicity to human vascular endothelial cells. J. Cell Sci. 1989, 94, 553–559. [Google Scholar]
- Szejtli, J. Medicinal applications of cyclodextrins. Med. Res. Rev. 1994, 14, 353–386. [Google Scholar] [CrossRef]
- Davis, M.E.; Brewster, M.E. Cyclodextrin-based pharmaceutics: Past, present and future. Nat. Rev. Drug Discov. 2004, 3, 1023–1035. [Google Scholar]
- Uekama, K.; Hirayama, F.; Irie, T. Cyclodextrin drug carrier systems. Chem. Rev. 1998, 98, 2045–2076. [Google Scholar] [CrossRef]
- Uekama, K. Design and evaluation of cyclodextrin-based drug formulation. Chem. Pharm. Bull. 2004, 52, 900–915. [Google Scholar] [CrossRef]
- Szente, L.; Szejtli, J. Highly soluble cyclodextrin derivatives: Chemistry, properties, and trends in development. Adv. Drug Deliv. Rev. 1999, 36, 17–28. [Google Scholar] [CrossRef]
- Irie, T.; Otagiri, M.; Sunada, M.; Uekama, K.; Ohtani, Y.; Yamada, Y.; Sugiyama, Y. Cyclodextrin-induced hemolysis and shape changes of human erythrocytes in vitro. J. Pharmacobiodyn. 1982, 5, 741–744. [Google Scholar] [CrossRef]
- Ohtani, Y.; Irie, T.; Uekama, K.; Fukunaga, K.; Pitha, J. Differential effects of α-, β- and γ-cyclodextrins on human erythrocytes. Eur. J. Biochem. 1989, 186, 17–22. [Google Scholar] [CrossRef]
- Fauvelle, F.; Debouzy, J.C.; Crouzy, S.; Goschl, M.; Chapron, Y. Mechanism of α-cyclodextrin-induced hemolysis. 1. The two-step extraction of phosphatidylinositol from the membrane. J. Pharm. Sci. 1997, 86, 935–943. [Google Scholar] [CrossRef]
- Arima, H.; Motoyama, K. Recent findings concerning PAMAM dendrimer conjugates with cyclodextrins as carriers of DNA and RNA. Sensors 2009, 9, 6364–6361. [Google Scholar]
- Pack, D.W.; Hoffman, A.S.; Pun, S.; Stayton, P.S. Design and development of polymers for gene delivery. Nat. Rev. Drug Discov. 2005, 4, 581–593. [Google Scholar] [CrossRef]
- Ortiz Mellet, C.; Garcia Fernandez, J.M.; Benito, J.M. Cyclodextrin-based gene delivery systems. Chem. Soc. Rev. 2011, 40, 1586–1608. [Google Scholar] [CrossRef]
- Gonzalez, H.; Hwang, S.J.; Davis, M.E. New class of polymers for the delivery of macromolecular therapeutics. Bioconjug. Chem. 1999, 10, 1068–1074. [Google Scholar] [CrossRef]
- Hwang, S.J.; Bellocq, N.C.; Davis, M.E. Effects of structure of β-cyclodextrin-containing polymers on gene delivery. Bioconjug. Chem. 2001, 12, 280–290. [Google Scholar] [CrossRef]
- Pun, S.H.; Tack, F.; Bellocq, N.C.; Cheng, J.; Grubbs, B.H.; Jensen, G.S.; Davis, M.E.; Brewster, M.; Janicot, M.; Janssens, B.; Floren, W.; Bakker, A. Targeted delivery of RNA-cleaving DNA enzyme (DNAzyme) to tumor tissue by transferrin-modified, cyclodextrin-based particles. Cancer Biol. Ther. 2004, 3, 641–650. [Google Scholar] [CrossRef]
- Bartlett, D.W.; Davis, M.E. Impact of tumor-specific targeting and dosing schedule on tumor growth inhibition after intravenous administration of siRNA-containing nanoparticles. Biotechnol. Bioeng. 2008, 99, 975–985. [Google Scholar] [CrossRef]
- Davis, M.E. The first targeted delivery of siRNA in humans via a self-assembling, cyclodextrin polymer-based nanoparticle: From concept to clinic. Mol. Pharm. 2009, 6, 659–668. [Google Scholar] [CrossRef]
- Davis, M.E.; Zuckerman, J.E.; Choi, C.H.; Seligson, D.; Tolcher, A.; Alabi, C.A.; Yen, Y.; Heidel, J.D.; Ribas, A. Evidence of RNAi in humans from systemically administered siRNA via targeted nanoparticles. Nature 2010, 464, 1067–1070. [Google Scholar]
- Li, J.J.; Zhao, F.; Li, J. Supramolecular polymers based on cyclodextrins for drug and gene delivery. Adv. Biochem. Eng. Biotechnol. 2011, 125, 207–249. [Google Scholar]
- Arima, H.; Kihara, F.; Hirayama, F.; Uekama, K. Enhancement of gene expression by polyamidoamine dendrimer conjugates with α-, β-, and γ-cyclodextrins. Bioconjug. Chem. 2001, 12, 476–484. [Google Scholar] [CrossRef]
- Kihara, F.; Arima, H.; Tsutsumi, T.; Hirayama, F.; Uekama, K. Effects of structure of polyamidoamine dendrimer on gene transfer efficiency of the dendrimer conjugate with α-cyclodextrin. Bioconjug. Chem. 2002, 13, 1211–1219. [Google Scholar] [CrossRef]
- Kihara, F.; Arima, H.; Tsutsumi, T.; Hirayama, F.; Uekama, K. In vitro and in vivo gene transfer by an optimized α-cyclodextrin conjugate with polyamidoamine dendrimer. Bioconjug. Chem. 2003, 14, 342–350. [Google Scholar] [CrossRef]
- Ouyang, D.; Zhang, H.; Parekh, H.S.; Smith, S.C. The effect of pH on PAMAM dendrimer-siRNA complexation—Endosomal considerations as determined by molecular dynamics simulation. Biophys. Chem. 2011, 158, 126–133. [Google Scholar] [CrossRef]
- Arima, H. Recent findings of dendrimers and their conjugates as non-viral vectors. Recent Res. Devel. Bioconj. Chem. 2005, 2, 109–126. [Google Scholar]
- Wada, K.; Arima, H.; Tsutsumi, T.; Chihara, Y.; Hattori, K.; Hirayama, F.; Uekama, K. Improvement of gene delivery mediated by mannosylated dendrimer/α-cyclodextrin conjugates. J. Control. Release 2005, 104, 397–413. [Google Scholar] [CrossRef]
- Xu, J.; Ganesh, S.; Amiji, M. Non-condensing polymeric nanoparticles for targeted gene and siRNA delivery. Int. J. Pharm. 2011, in press. [Google Scholar]
- Arima, H.; Yamashita, S.; Mori, Y.; Hayashi, Y.; Motoyama, K.; Hattori, K.; Takeuchi, T.; Jono, H.; Ando, Y.; Hirayama, F.; et al. In vitro and in vivo gene delivery mediated by Lactosylated dendrimer/α-cyclodextrin conjugates (G2) into hepatocytes. J. Control. Release 2010, 146, 106–117. [Google Scholar] [CrossRef]
- Yoshimatsu, A.; Arizono, M.; Motoyama, K.; Hattori, K.; Takeuchi, T.; Hirayama, F.; Uekama, K.; Arima, H. Folate-PEG-Appended Dendrimer Conjugate with α-Cyclodextrin as a Novel Cancer Cell-Selective Gene Transfer Carrier. In Proceedings of the 14th International Cyclodextrins Symposium, Kyoto, Japan, 8-11 May 2008; pp. 276–281.
- Tavornvipas, S.; Hirayama, F.; Arima, H.; Uekama, K.; Ishiguro, T.; Oka, M.; Hamayasu, K.; Hashimoto, H. 6-O-α-(4-O-α-D-glucuronyl)-D-glucosyl-β-cyclodextrin: Solubilizing ability and some cellular effects. Int. J. Pharm. 2002, 249, 199–209. [Google Scholar] [CrossRef]
- Anno, T.; Motoyama, K.; Higashi, T.; Hirayama, F.; Uekama, K.; Arima, H. Preparation and evaluation of polyamidoamine dendrimer (G2)/branched-β-cyclodextrin conjugate as a novel gene transfer carrier. J. Incl. Phenom. Macrocycl. Chem. 2011, 70, 339–344. [Google Scholar] [CrossRef]
- Schroder, H.C.; Ushijima, H.; Theis, C.; Seve, A.P.; Hubert, J.; Muller, W.E. Expression of nuclear lectin carbohydrate-binding protein 35 in human immunodeficiency virus type 1-infected Molt-3 cells. J. Acquir. Immune Defic. Syndr. Hum. Retrovirol. 1995, 9, 340–348. [Google Scholar]
- Haudek, K.C.; Spronk, K.J.; Voss, P.G.; Patterson, R.J.; Wang, J.L.; Arnoys, E.J. Dynamics of galectin-3 in the nucleus and cytoplasm. Biochim. Biophys. Acta 1800, 181–189. [Google Scholar]
- Seve, A.P.; Felin, M.; Doyennette-Moyne, M.A.; Sahraoui, T.; Aubery, M.; Hubert, J. Evidence for a lactose-mediated association between two nuclear carbohydrate-binding proteins. Glycobiology 1993, 3, 23–30. [Google Scholar] [CrossRef]
- Rousseau, C.; Felin, M.; Doyennette-Moyne, M.A.; Seve, A.P. CBP70, a glycosylated nuclear lectin. J. Cell Biochem. 1997, 66, 370–385. [Google Scholar] [CrossRef]
- Ko, Y.T.; Kale, A.; Hartner, W.C.; Papahadjopoulos-Sternberg, B.; Torchilin, V.P. Self-assembling micelle-like nanoparticles based on phospholipid-polyethyleneimine conjugates for systemic gene delivery. J. Control. Release 2009, 133, 132–138. [Google Scholar] [CrossRef]
- Anno, T.; Higashi, T.; Motoyama, K.; Hirayama, F.; Uekama, K.; Arima, H. Potential use of glucuronylglucosyl-β-cyclodextrin/dendrimer conjugate (G2) as a DNA carrier in vitro and in vivo. J. Drug Target. 2011, in press.. [Google Scholar]
- Alisky, J.M.; Davidson, B.L. Towards therapy using RNA interference. Am. J. Pharmacogenomics 2004, 4, 45–51. [Google Scholar] [CrossRef]
- McManus, M.T.; Sharp, P.A. Gene silencing in mammals by small interfering RNAs. Nat. Rev. Genet. 2002, 3, 737–747. [Google Scholar] [CrossRef]
- Zender, L.; Kubicka, S. Suppression of apoptosis in the liver by systemic and local delivery of small-interfering RNAs. Methods Mol. Biol. 2007, 361, 217–226. [Google Scholar]
- Thomas, M.; Klibanov, A.M. Non-viral gene therapy: Polycation-mediated DNA delivery. Appl. Microbiol. Biotechnol. 2003, 62, 27–34. [Google Scholar] [CrossRef]
- Tsutsumi, T.; Hirayama, F.; Uekama, K.; Arima, H. Evaluation of polyamidoamine dendrimer/α-cyclodextrin conjugate (generation 3, G3) as a novel carrier for small interfering RNA (siRNA). J. Control. Release 2007, 119, 349–359. [Google Scholar] [CrossRef] [Green Version]
- Tsutsumi, T.; Hirayama, F.; Uekama, K.; Arima, H. Potential use of polyamidoamine dendrimer/α-cyclodextrin conjugate (generation 3, G3) as a novel carrier for short hairpin RNA-expressing plasmid DNA. J. Pharm. Sci. 2008, 97, 3022–3034. [Google Scholar] [CrossRef]
- Arima, H.; Tsutsumi, T.; Yoshimatsu, A.; Ikeda, H.; Motoyama, K.; Higashi, T.; Hirayama, F.; Uekama, K. Inhibitory effect of siRNA complexes with polyamidoamine dendrimer/α-cyclodextrin conjugate (generation 3, G3) on endogenous gene expression. Eur. J. Pharm. Sci. 2011, in press. [Google Scholar]
- Li, J.J.; Zhao, F.; Li, J. Polyrotaxanes for applications in life science and biotechnology. Appl. Microbiol. Biotechnol. 2011, 90, 427–443. [Google Scholar] [CrossRef]
- Motoyama, K.; Hayashida, K.; Arima, H. Potential use of polypseudorotaxanes of pegylated polyamidoamine dendrimer with cyclodextrins as novel sustained release systems for DNA. Chem. Pharm. Bull. 2011, 59, 476–479. [Google Scholar] [CrossRef]
- Motoyama, K.; Hayashida, K.; Higashi, T.; Arima, H. Polypseudorotaxanes of pegylated α-cyclodextrin/polyamidoamine dendrimer conjugate with cyclodextrins as a sustained release system for DNA. Bioorg. Med. Chem. 2012, in press. [Google Scholar]
- Arima, H.; Chihara, Y.; Arizono, M.; Yamashita, S.; Wada, K.; Hirayama, F.; Uekama, K. Enhancement of gene transfer activity mediated by mannosylated dendrimer/α-cyclodextrin conjugate (generation 3, G3). J. Control. Release 2006, 116, 64–74. [Google Scholar] [CrossRef]
- Wada, K.; Arima, H.; Tsutsumi, T.; Hirayama, F.; Uekama, K. Enhancing effects of galactosylated dendrimer/α-cyclodextrin conjugates on gene transfer efficiency. Biol. Pharm. Bull. 2005, 28, 500–505. [Google Scholar] [CrossRef]
- Motoyama, K.; Mori, Y.; Yamashita, S.; Hayashi, Y.; Jono, H.; Ando, Y.; Hirayama, F.; Uekama, K.; Arima, H. In vitro gene delivery mediated by lactosylated dendrimer (generation 3, G3)/α-cyclodextrin conjugates into hepatocytes. J. Incl. Phenom. Macrocycl. Chem. 2011, 70, 333–338. [Google Scholar] [CrossRef]
- Arima, H.; Motoyama, K.; Higashi, T. Potential use of polyamidoamine dendrimer conjugates with cyclodextrins as novel carriers for siRNA. Pharmaceuticals 2012, 5, 61–78. [Google Scholar]
© 2012 by the authors; licensee MDPI, Basel, Switzerland. This article is an open-access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/3.0/).
Share and Cite
Arima, H.; Motoyama, K.; Higashi, T. Polyamidoamine Dendrimer Conjugates with Cyclodextrins as Novel Carriers for DNA, shRNA and siRNA. Pharmaceutics 2012, 4, 130-148. https://doi.org/10.3390/pharmaceutics4010130
Arima H, Motoyama K, Higashi T. Polyamidoamine Dendrimer Conjugates with Cyclodextrins as Novel Carriers for DNA, shRNA and siRNA. Pharmaceutics. 2012; 4(1):130-148. https://doi.org/10.3390/pharmaceutics4010130
Chicago/Turabian StyleArima, Hidetoshi, Keiichi Motoyama, and Taishi Higashi. 2012. "Polyamidoamine Dendrimer Conjugates with Cyclodextrins as Novel Carriers for DNA, shRNA and siRNA" Pharmaceutics 4, no. 1: 130-148. https://doi.org/10.3390/pharmaceutics4010130