
Bornyl-Containing Derivatives of Benzyloxyphenylpropanoic Acid as FFAR1 Agonists: In Vitro and In Vivo Studies
 , , , , , ,
, , , , , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
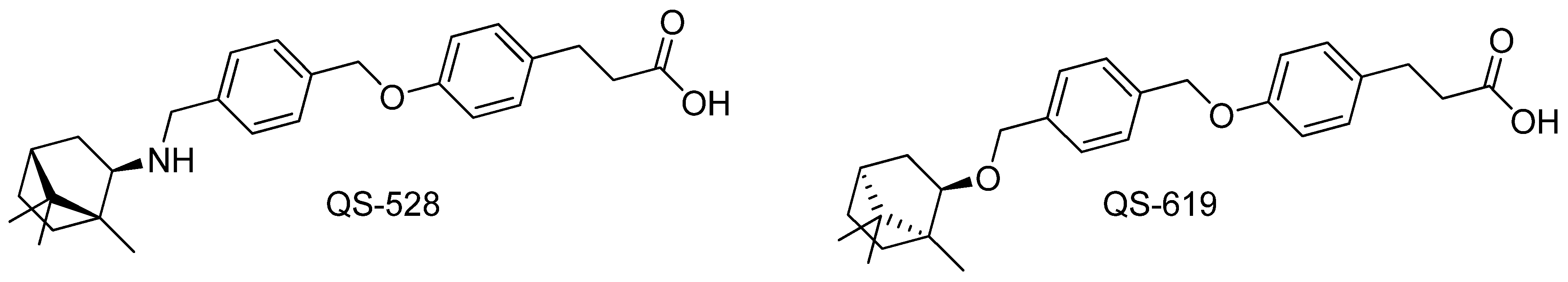
2.1. Investigated Compound
2.2. Animals
2.3. Oral Glucose Tolerance Test (OGTT)
2.4. Diet-Induced Model of Type 2 Diabetes Mellitus
2.5. Biochemical Assays
2.6. Histological Liver Examination
2.7. In Vitro FFAR1 Activation Assay
2.8. In Vitro Inhibition of DPP4 Assay
2.9. Insulin ELISA Examination
2.10. GIP ELISA Examination
2.11. Toxicology Study
2.12. Statistical Analysis
3. Results
3.1. OGTT in C57Bl6/J Mice
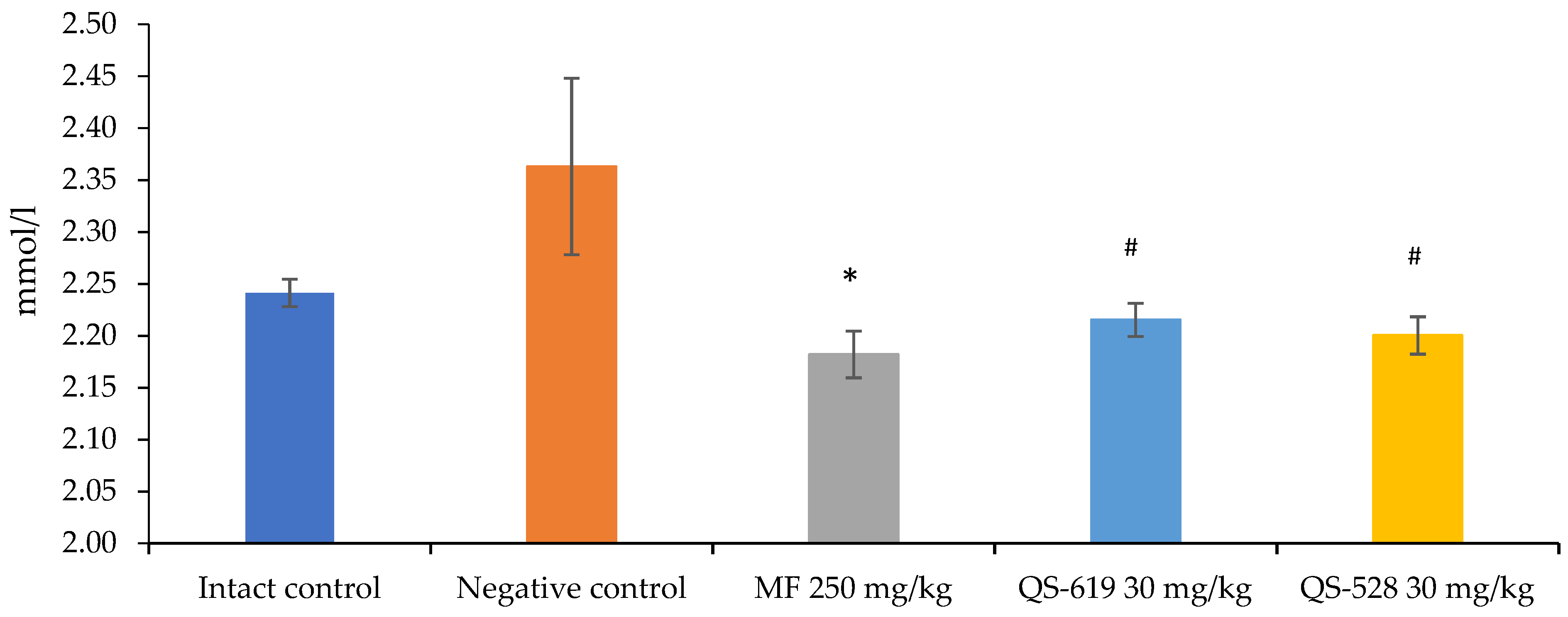
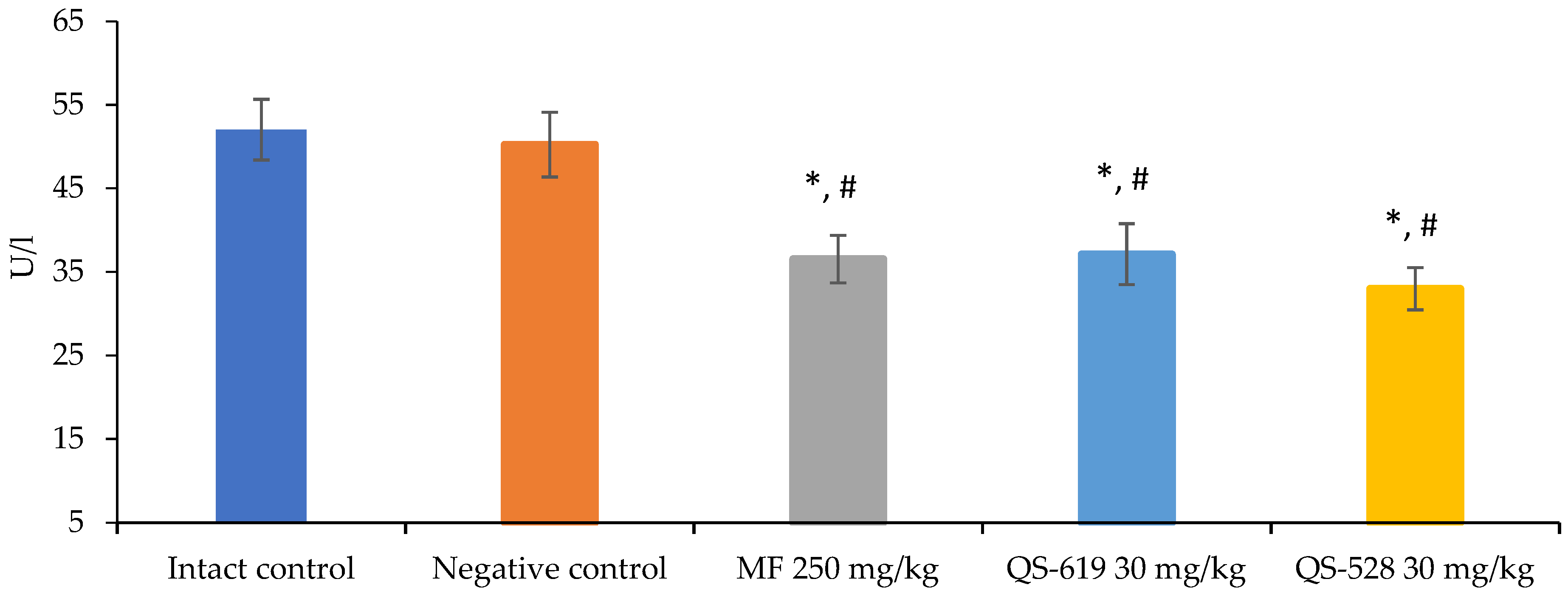
3.2. A Biochemical Blood Assay
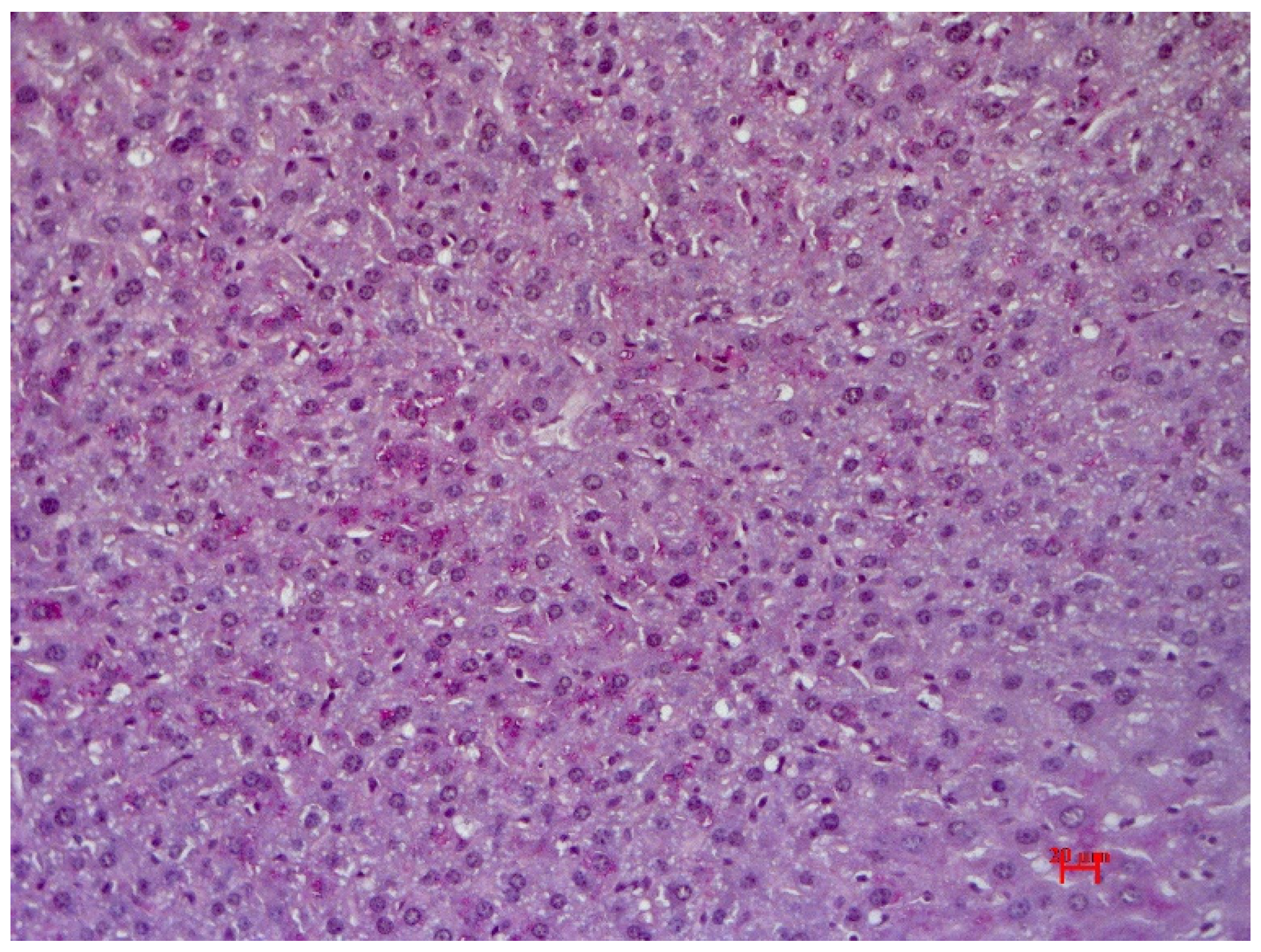
3.3. Histological Examination
3.4. In Vitro FFAR1 Activation and DPP IV Inhibition Assay
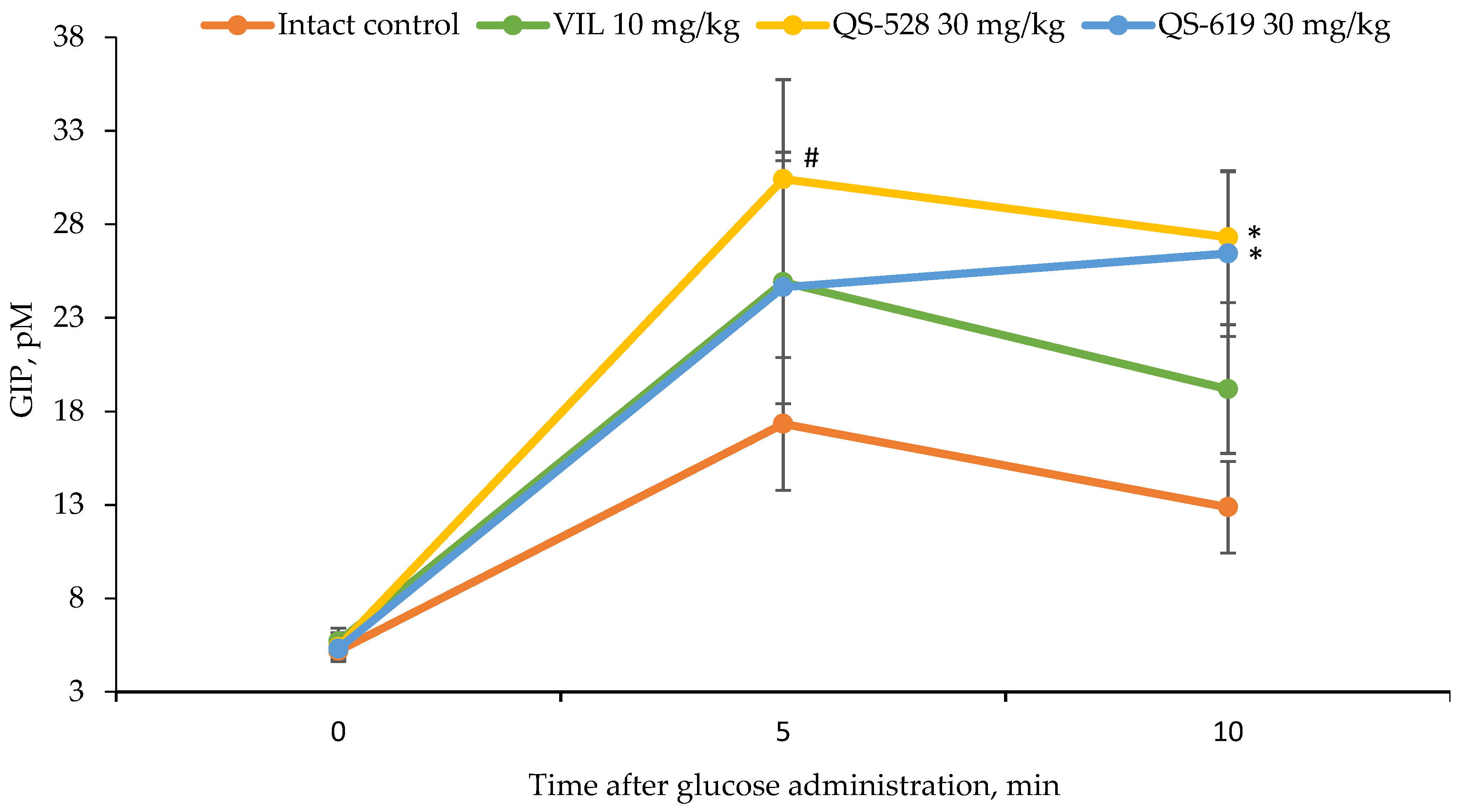
3.5. Measurement of Serum Insulin and GIP Concentrations
3.6. Acute Toxicity Study
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Petersen, M.C.; Shulman, G.I. Mechanisms of Insulin Action and Insulin Resistance. Physiol. Rev. 2018, 98, 2133–2223. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faselis, C.; Katsimardou, A.; Imprialos, K.; Deligkaris, P.; Kallistratos, M.; Dimitriadis, K. Microvascular Complications of Type 2 Diabetes Mellitus. Curr. Vasc. Pharmacol. 2020, 18, 117–124. [Google Scholar] [CrossRef] [PubMed]
- Mastrototaro, L.; Roden, M. Insulin Resistance and Insulin Sensitizing Agents. Metabolism 2021, 125, 154892. [Google Scholar] [CrossRef] [PubMed]
- Rendell, M.S. Current and Emerging Gluconeogenesis Inhibitors for the Treatment of Type 2 Diabetes. Expert Opin. Pharmacother. 2021, 22, 2167–2179. [Google Scholar] [CrossRef]
- Chen, C.; Li, H.; Long, Y.Q. GPR40 Agonists for the Treatment of Type 2 Diabetes Mellitus: The Biological Characteristics and the Chemical Space. Bioorg. Med. Chem. Lett. 2016, 26, 5603–5612. [Google Scholar] [CrossRef]
- Mancini, A.D.; Poitout, V. GPR40 Agonists for the Treatment of Type 2 Diabetes: Life after “TAKing” a Hit. Diabetes Obes. Metab. 2015, 17, 622–629. [Google Scholar] [CrossRef]
- Li, Z.; Zhou, Z.; Zhang, L. Current Status of GPR40/FFAR1 Modulators in Medicinal Chemistry (2016–2019): A Patent Review. Expert Opin. Ther. Pat. 2020, 30, 27–38. [Google Scholar] [CrossRef]
- Marcinak, J.F.; Munsaka, M.S.; Watkins, P.B.; Ohira, T.; Smith, N. Liver Safety of Fasiglifam (TAK-875) in Patients with Type 2 Diabetes: Review of the Global Clinical Trial Experience. Drug Saf. 2018, 41, 625–640. [Google Scholar] [CrossRef]
- Governa, P.; Caroleo, M.C.; Carullo, G.; Aiello, F.; Cione, E.; Manetti, F. FFAR1/GPR40: One Target, Different Binding Sites, Many Agonists, No Drugs, but a Continuous and Unprofitable Tug-of-War between Ligand Lipophilicity, Activity, and Toxicity. Bioorg. Med. Chem. Lett. 2021, 41, 127969. [Google Scholar] [CrossRef]
- Hauge, M.; Vestmar, M.A.; Husted, A.S.; Ekberg, J.P.; Wright, M.J.; di Salvo, J.; Weinglass, A.B.; Engelstoft, M.S.; Madsen, A.N.; Lückmann, M.; et al. GPR40 (FFAR1)—Combined Gs and Gq Signaling Invitro Is Associated with Robust Incretin Secretagogue Action Ex Vivo and in Vivo. Mol. Metab. 2015, 4, 3–14. [Google Scholar] [CrossRef]
- Ookawara, M.; Matsuda, K.; Watanabe, M.; Moritoh, Y. The GPR40 Full Agonist SCO-267 Improves Liver Parameters in a Mouse Model of Nonalcoholic Fatty Liver Disease without Affecting Glucose or Body Weight. J. Pharmacol. Exp. Ther. 2020, 375, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Secor, J.D.; Fligor, S.C.; Tsikis, S.T.; Yu, L.J.; Puder, M. Free Fatty Acid Receptors as Mediators and Therapeutic Targets in Liver Disease. Front. Physiol. 2021, 12, 656441. [Google Scholar] [CrossRef] [PubMed]
- Vergani, L. Fatty Aids and Effects on in Vitro and in Vivo Models of Liver Steatosis. Curr. Med. Chem. 2017, 26, 3439–3456. [Google Scholar] [CrossRef] [PubMed]
- Kuranov, S.O.; Luzina, O.A.; Onopchenko, O.; Pishel, I.; Zozulya, S.; Gureev, M.; Salakhutdinov, N.F.; Krasavin, M. Exploring Bulky Natural and Natural-like Periphery in the Design of p-(Benzyloxy)Phenylpropionic Acid Agonists of Free Fatty Acid Receptor 1 (GPR40). Bioorg. Chem. 2020, 99, 103830. [Google Scholar] [CrossRef]
- Kuranov, S.; Luzina, O.; Khvostov, M.; Baev, D.; Kuznetsova, D.; Zhukova, N.; Vassiliev, P.; Kochetkov, A.; Tolstikova, T.; Salakhutdinov, N. Bornyl Derivatives of P-(Benzyloxy)Phenylpropionic Acid: In Vivo Evaluation of Antidiabetic Activity. Pharmaceuticals 2020, 13, 404. [Google Scholar] [CrossRef]
- Tai, M.M. A Mathematical Model for the Determination of Total Area under Glucose Tolerance and Other Metabolic Curves. Diabetes Care 1994, 17, 152–154. [Google Scholar] [CrossRef] [Green Version]
- Scheen, A.J. Investigational Insulin Secretagogues for Type 2 Diabetes. Expert Opin. Investig. Drugs 2016, 25, 405–422. [Google Scholar] [CrossRef]
- Edfalk, S.; Steneberg, P.; Edlund, H. Gpr40 Is Expressed in Enteroendocrine Cells and Mediates Free Fatty Acid Stimulation of Incretin Secretion. Diabetes 2008, 57, 2280–2287. [Google Scholar] [CrossRef] [Green Version]
- Nagarajan, P.; Jerald Mahesh Kumar, M.; Venkatesan, R.; Majundar, S.S.; Juyal, R.C. Genetically Modified Mouse Models for the Study of Nonalcoholic Fatty Liver Disease. World J. Gastroenterol. 2012, 18, 1141–1153. [Google Scholar] [CrossRef]
- Lebedev, P.; Peresypkina, A.; Gustinovich, V.; Godunov, O.; Gustinovich, V.; Zatolokina, M.; Goliusov, A.; Pokrovskii, M. Correction of Metabolic Disturbances by Functional Food Compositions in Experimental Obesity in CD-1 and Agouti-Yellow Mice. Funct. Foods Health Dis. 2022, 12, 26–45. [Google Scholar] [CrossRef]
- Geerling, J.J.; Boon, M.R.; van der Zon, G.C.; van den Berg, S.A.A.; van den Hoek, A.M.; Lombès, M.; Princen, H.M.G.; Havekes, L.M.; Rensen, P.C.N.; Guigas, B. Metformin Lowers Plasma Triglycerides by Promoting Vldl-Triglyceride Clearance by Brown Adipose Tissue in Mice. Diabetes 2014, 63, 880–891. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.R.; Liu, J.; Cheng, L.D.; Liu, Z.Y.; Zheng, X.B.; Liang, H.; Xu, F. Metformin Alleviates Steatohepatitis in Diet-Induced Obese Mice in a SIRT1-Dependent Way. Front. Pharmacol. 2021, 12, 704112. [Google Scholar] [CrossRef] [PubMed]
- Saeedi Saravi, S.S.; Hasanvand, A.; Shahkarami, K.; Dehpour, A.R. The Protective Potential of Metformin against Acetaminophen-Induced Hepatotoxicity in BALB/C Mice. Pharm. Biol. 2016, 54, 2830–2837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gilbert, M.P.; Pratley, R.E. GLP-1 Analogs and DPP-4 Inhibitors in Type 2 Diabetes Therapy: Review of Head-to-Head Clinical Trials. Front. Endocrinol. 2020, 11, 178. [Google Scholar] [CrossRef] [PubMed]
- Ahrén, B.; Schweizer, A.; Dejager, S.; Villhauer, E.B.; Dunning, B.E.; Foley, J.E. Mechanisms of Action of the Dipeptidyl Peptidase-4 Inhibitor Vildagliptin in Humans. Diabetes Obes. Metab. 2011, 13, 775–783. [Google Scholar] [CrossRef] [PubMed]
- Burant, C.F. Activation of GPR40 as a Therapeutic Target for the Treatment of Type 2 Diabetes. Diabetes Care 2013, 36, S175–S179. [Google Scholar] [CrossRef] [Green Version]
- On, S.; Kim, H.Y.; Kim, H.S.; Park, J.; Kang, K.W. Involvement of G-Protein-Coupled Receptor 40 in the Inhibitory Effects of Docosahexaenoic Acid on SREBP1-Mediated Lipogenic Enzyme Expression in Primary Hepatocytes. Int. J. Mol. Sci. 2019, 20, 2625. [Google Scholar] [CrossRef] [Green Version]
- Yang, M.; Zhang, C.Y. G Protein-Coupled Receptors as Potential Targets for Nonalcoholic Fatty Liver Disease Treatment. World J. Gastroenterol. 2021, 27, 677–691. [Google Scholar] [CrossRef]
- Pon’kina, D.; Kuranov, S.; Khvostov, M.; Zhukova, N.; Meshkova, Y.; Marenina, M.; Luzina, O.; Tolstikova, T.; Salakhutdinov, N. Hepatoprotective Effect of a New FFAR1 Agonist—N-Alkylated Isobornylamine. Molecules 2023, 28, 396. [Google Scholar] [CrossRef]
- Ueno, H.; Ito, R.; Abe, S.I.; Ookawara, M.; Miyashita, H.; Ogino, H.; Miyamoto, Y.; Yoshihara, T.; Kobayashi, A.; Tsujihata, Y.; et al. SCO-267, a GPR40 Full Agonist, Improves Glycemic and Body Weight Control in Rat Models of Diabetes and Obesity. J. Pharmacol. Exp. Ther. 2019, 370, 172–181. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Meng, X.; Xu, J.; Huang, X.; Li, H.; Li, G.; Wang, S.; Man, Y.; Tang, W.; Li, J. GPR40 Agonist Ameliorates Liver X Receptor-Induced Lipid Accumulation in Liver by Activating AMPK Pathway. Sci. Rep. 2016, 6, 25237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sahgal, G.; Ramanathan, S.; Sasidharan, S.; Mordi, M.N.; Ismail, S.; Mansor, S.M. Brine Shrimp Lethality and Acute Oral Toxicity Studies on Swietenia Mahagoni (Linn.) Jacq. Seed Methanolic Extract. Pharmacogn. Res. 2010, 2, 215–220. [Google Scholar]













Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pon’kina, D.A.; Kuranov, S.O.; Marenina, M.K.; Meshkova, Y.V.; Zhukova, N.A.; Khvostov, M.V.; Luzina, O.A.; Tolstikova, T.G.; Salakhutdinov, N.F. Bornyl-Containing Derivatives of Benzyloxyphenylpropanoic Acid as FFAR1 Agonists: In Vitro and In Vivo Studies. Pharmaceutics 2023, 15, 1670. https://doi.org/10.3390/pharmaceutics15061670
Pon’kina DA, Kuranov SO, Marenina MK, Meshkova YV, Zhukova NA, Khvostov MV, Luzina OA, Tolstikova TG, Salakhutdinov NF. Bornyl-Containing Derivatives of Benzyloxyphenylpropanoic Acid as FFAR1 Agonists: In Vitro and In Vivo Studies. Pharmaceutics. 2023; 15(6):1670. https://doi.org/10.3390/pharmaceutics15061670
Chicago/Turabian StylePon’kina, Darya A., Sergey O. Kuranov, Mariya K. Marenina, Yulia V. Meshkova, Nataliya A. Zhukova, Mikhail V. Khvostov, Olga A. Luzina, Tatiana G. Tolstikova, and Nariman F. Salakhutdinov. 2023. "Bornyl-Containing Derivatives of Benzyloxyphenylpropanoic Acid as FFAR1 Agonists: In Vitro and In Vivo Studies" Pharmaceutics 15, no. 6: 1670. https://doi.org/10.3390/pharmaceutics15061670