
Design of Folate-Containing Liposomal Nucleic Acid Delivery Systems for Antitumor Therapy

Abstract
:1. Introduction
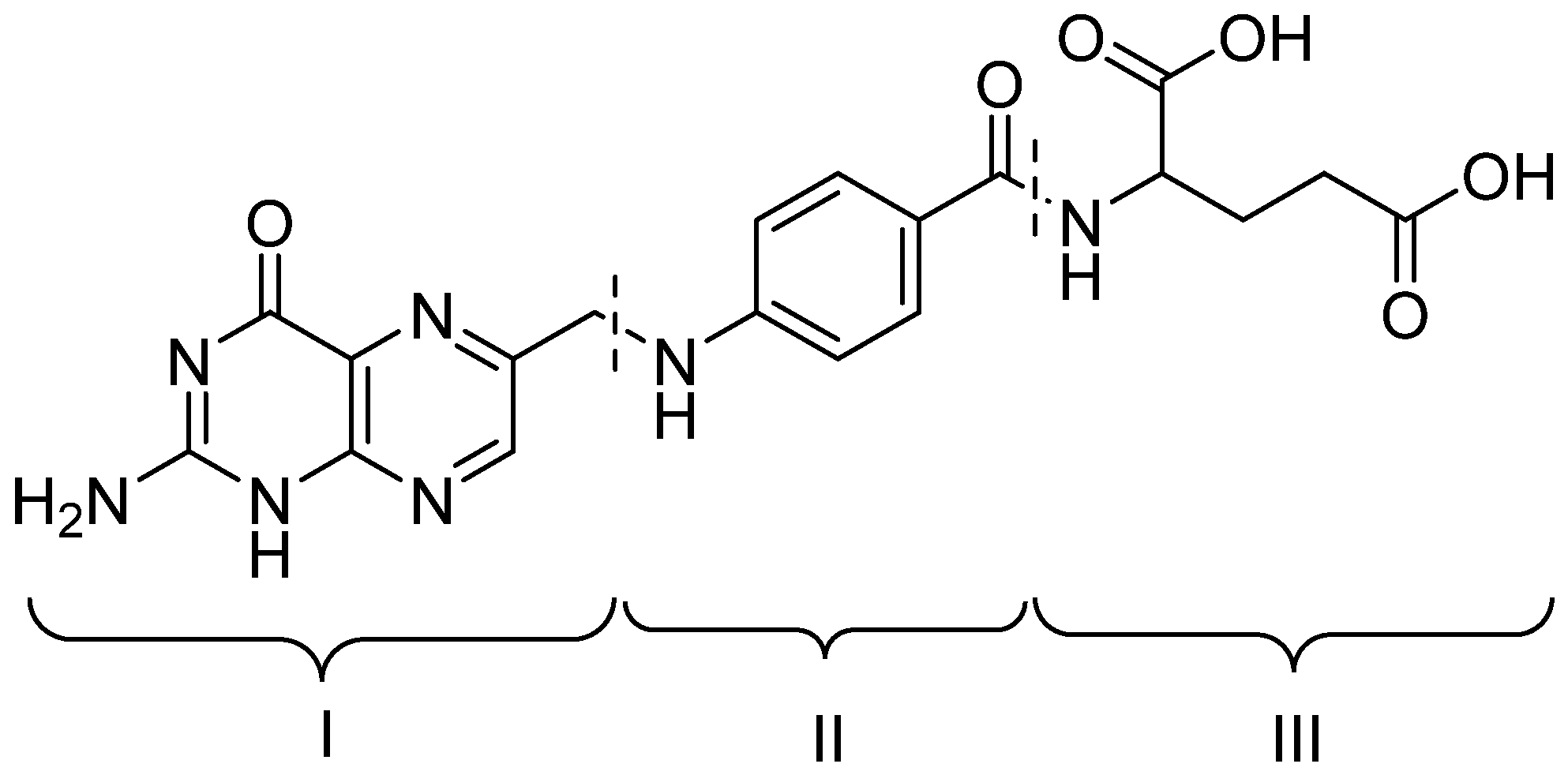
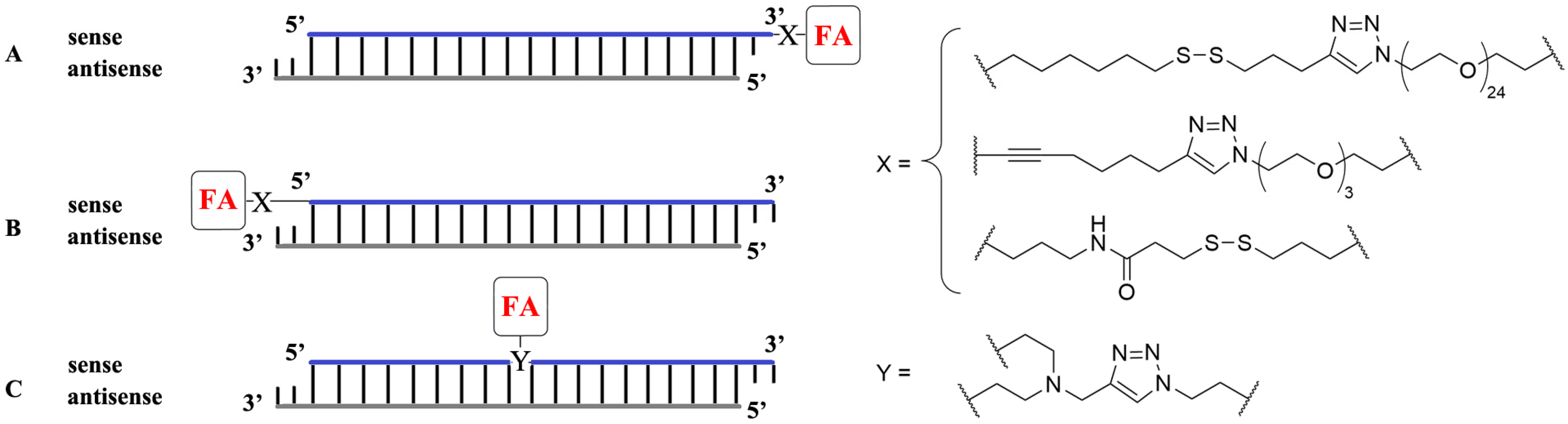
2. Conjugation of Folic Acid to Nucleic Acids
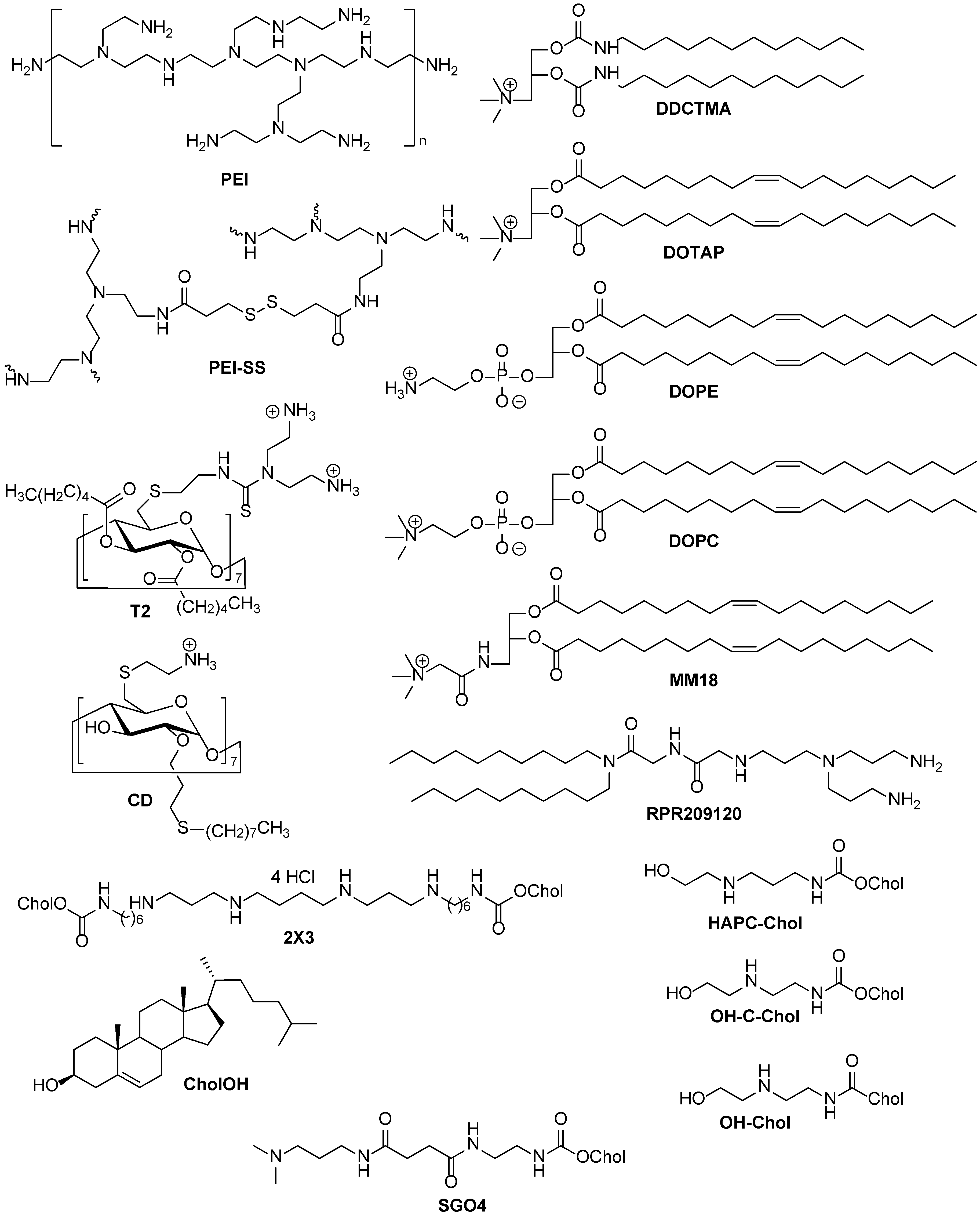
3. Noncovalent Interaction between Folic Acid and Nonviral Delivery Systems

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Delivered NA | Delivery System | FA Incorporation | Optimized FA Amount | Cell Line | Ref. |
|---|---|---|---|---|---|
| pCMV-luc pDNA | PEI 1 | adding to culture media | 1 mM | KB, F98, CHO | [40] |
| mixing of FA and NA with subsequent complexation | 0.5 mM | ||||
| adding to preformed complexes | Charge ratio FA/PEI/pDNA 60:8:1 | B16-F10 | [41] | ||
| survivin siRNA | PEI-SS 1 | adding to preformed complexes | Charge ratio FA/PEI-SS/siRNA 20:8:1 | HeLa, A549 | [42] |
| pGFP-N2 and pGL3 pDNA | DDCTMA 1–CholOH 1 (3:1) | adding to preformed complexes | 100 μg/mL | NCI-H460 | [43] |
| pEGFP-C2 pDNA | 2X3 1–DOPE 1 (1:2) | adding to preformed complexes | 1 mM | KB-3-1 | [44] |

4. Folate Lipoconjugates as Components for Liposomal Delivery System
4.1. FA Lipoconjugate Structure
4.2. Optimal Content of FA Lipoconjugates with Different PEG Spacers in Cationic Liposomes
4.3. Optimal Physicochemical Properties of Folate-Containing Liposomal NA Delivery Systems
5. Conclusions and Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Crooke, S.T.; Witztum, J.L.; Bennett, C.F.; Baker, B.F. RNA-Targeted Therapeutics. Cell Metab. 2018, 27, 714–739. [Google Scholar] [CrossRef]
- Blanco, E.; Shen, H.; Ferrari, M. Principles of Nanoparticle Design for Overcoming Biological Barriers to Drug Delivery. Nat. Biotechnol. 2015, 33, 941–951. [Google Scholar] [CrossRef]
- Assaraf, Y.G.; Leamon, C.P.; Reddy, J.A. The Folate Receptor as a Rational Therapeutic Target for Personalized Cancer Treatment. Drug Resist. Updat. 2014, 17, 89–95. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.-H.; Kim, J.-K.; Lim, S.-J.; Park, J.-S.; Lee, M.-K.; Kim, C.-K. Folate-Tethered Emulsion for the Target Delivery of Retinoids to Cancer Cells. Eur. J. Pharm. Biopharm. 2008, 68, 618–625. [Google Scholar] [CrossRef] [PubMed]
- Siwowska, K.; Schmid, R.M.; Cohrs, S.; Schibli, R.; Müller, C. Folate Receptor-Positive Gynecological Cancer Cells: In Vitro and In Vivo Characterization. Pharmaceuticals 2017, 10, 72. [Google Scholar] [CrossRef] [PubMed]
- Shen, J.; Hu, Y.; Putt, K.S.; Singhal, S.; Han, H.; Visscher, D.W.; Murphy, L.M.; Low, P.S. Assessment of Folate Receptor Alpha and Beta Expression in Selection of Lung and Pancreatic Cancer Patients for Receptor Targeted Therapies. Oncotarget 2018, 9, 4485–4495. [Google Scholar] [CrossRef] [PubMed]
- Kalber, T.L.; Kamaly, N.; So, P.W.; Pugh, J.A.; Bunch, J.; McLeod, C.W.; Jorgensen, M.R.; Miller, A.D.; Bell, J.D. A Low Molecular Weight Folate Receptor Targeted Contrast Agent for Magnetic Resonance Tumor Imaging. Mol. Imaging Biol. 2011, 13, 653–662. [Google Scholar] [CrossRef] [PubMed]
- Antony, A.C. The Biological Chemistry of Folate Receptors. Blood 1992, 79, 2807–2820. [Google Scholar] [CrossRef]
- Liang, X.; Luo, M.; Wei, X.W.; Ma, C.C.; Yang, Y.H.; Shao, B.; Liu, Y.T.; Liu, T.; Ren, J.; Liu, L.; et al. A Folate Receptor-Targeted Lipoplex Delivering Interleukin-15 Gene for Colon Cancer Immunotherapy. Oncotarget 2016, 7, 52207–52217. [Google Scholar] [CrossRef]
- Ledermann, J.A.; Canevari, S.; Thigpen, T. Targeting the Folate Receptor: Diagnostic and Therapeutic Approaches to Personalize Cancer Treatments. Ann. Oncol. 2015, 26, 2034–2043. [Google Scholar] [CrossRef]
- Kabilova, T.O.; Shmendel, E.V.; Gladkikh, D.V.; Chernolovskaya, E.L.; Markov, O.V.; Morozova, N.G.; Maslov, M.A.; Zenkova, M.A. Targeted Delivery of Nucleic Acids into Xenograft Tumors Mediated by Novel Folate-Equipped Liposomes. Eur. J. Pharm. Biopharm. 2018, 123, 59–70. [Google Scholar] [CrossRef] [PubMed]
- Tie, Y.; Zheng, H.; He, Z.; Yang, J.; Shao, B.; Liu, L.; Luo, M.; Yuan, X.; Liu, Y.; Zhang, X.; et al. Targeting Folate Receptor β Positive Tumor-Associated Macrophages in Lung Cancer with a Folate-Modified Liposomal Complex. Signal Transduct. Target. Ther. 2020, 5, 6. [Google Scholar] [CrossRef] [PubMed]
- Sánchez-Del-Campo, L.; Montenegro, M.F.; Cabezas-Herrera, J.; Rodríguez-López, J.N. The Critical Role of Alpha-Folate Receptor in the Resistance of Melanoma to Methotrexate. Pigment Cell Melanoma Res. 2009, 22, 588–600. [Google Scholar] [CrossRef] [PubMed]
- Motoyama, K.; Onodera, R.; Tanaka, N.; Kameyama, K.; Higashi, T.; Kariya, R.; Okada, S.; Arima, H. Evaluation of Antitumor Effects of Folate-Conjugated Methyl-β-Cyclodextrin in Melanoma. Biol. Pharm. Bull. 2015, 38, 374–379. [Google Scholar] [CrossRef]
- Norton, N.; Youssef, B.; Hillman, D.W.; Nassar, A.; Geiger, X.J.; Necela, B.M.; Liu, H.; Ruddy, K.J.; Polley, M.Y.C.; Ingle, J.N.; et al. Folate Receptor Alpha Expression Associates with Improved Disease-Free Survival in Triple Negative Breast Cancer Patients. npj Breast Cancer 2020, 6, 4. [Google Scholar] [CrossRef]
- Pierce, J.T.; Cho, S.S.; Nag, S.; Zeh, R.; Jeon, J.; Holt, D.; Durham, A.; Nasrallah, M.P.; Singhal, S.; Lee, J.Y.K. Folate Receptor Overexpression in Human and Canine Meningiomas-Immunohistochemistry and Case Report of Intraoperative Molecular Imaging. Clin. Neurosurg. 2019, 85, 359–368. [Google Scholar] [CrossRef]
- Yang, R.; Kolb, E.A.; Qin, J.; Chou, A.; Sowers, R.; Hoang, B.; Healey, J.H.; Huvos, A.G.; Meyers, P.A.; Gorlick, R. The Folate Receptor α Is Frequently Overexpressed in Osteosarcoma Samples and Plays a Role in the Uptake of the Physiologic Substrate 5-Methyltetrahydrofolate. Clin. Cancer Res. 2007, 13, 2557–2567. [Google Scholar] [CrossRef]
- Cheung, A.; Bax, H.J.; Josephs, D.H.; Ilieva, K.M.; Pellizzari, G.; Opzoomer, J.; Bloomfield, J.; Fittall, M.; Grigoriadis, A.; Figini, M.; et al. Targeting Folate Receptor Alpha for Cancer Treatment. Oncotarget 2016, 7, 52553–52574. [Google Scholar] [CrossRef]
- Scaranti, M.; Cojocaru, E.; Banerjee, S.; Banerji, U. Exploiting the Folate Receptor α in Oncology. Nat. Rev. Clin. Oncol. 2020, 17, 349–359. [Google Scholar] [CrossRef]
- Loenen, W.A.M. S-Adenosylmethionine: Jack of All Trades and Master of Everything? Biochem. Soc. Trans. 2006, 34, 330–333. [Google Scholar] [CrossRef]
- Rana, A.; Bhatnagar, S. Advancements in Folate Receptor Targeting for Anti-Cancer Therapy: A Small Molecule-Drug Conjugate Approach. Bioorg. Chem. 2021, 112, 104946. [Google Scholar] [CrossRef]
- Kharkar, P.S.; Soni, G.; Rathod, V.; Shetty, S.; Gupta, M.K.; Yadav, K.S. An Outlook on Procedures of Conjugating Folate to (Co)Polymers and Drugs for Effective Cancer Targeting. Drug Dev. Res. 2020, 81, 823–836. [Google Scholar] [CrossRef]
- Mornet, E.; Carmoy, N.; Lainé, C.; Lemiègre, L.; Le Gall, T.; Laurent, I.; Marianowski, R.; Férec, C.; Lehn, P.; Benvegnu, T.; et al. Folate-Equipped Nanolipoplexes Mediated Efficient Gene Transfer into Human Epithelial Cells. Int. J. Mol. Sci. 2013, 14, 1477–1501. [Google Scholar] [CrossRef]
- Abbasi, H.; Rahbar, N.; Kouchak, M.; Khalil Dezfuli, P.; Handali, S. Functionalized Liposomes as Drug Nanocarriers for Active Targeted Cancer Therapy: A Systematic Review. J. Liposome Res. 2022, 32, 195–210. [Google Scholar] [CrossRef]
- Saraf, S.; Jain, A.; Tiwari, A.; Verma, A.; Panda, P.K.; Jain, S.K. Advances in Liposomal Drug Delivery to Cancer: An Overview. J. Drug Deliv. Sci. Technol. 2020, 56, 101549. [Google Scholar] [CrossRef]
- Mirzavi, F.; Barati, M.; Soleimani, A.; Vakili-Ghartavol, R.; Jaafari, M.R.; Soukhtanloo, M. A Review on Liposome-Based Therapeutic Approaches against Malignant Melanoma. Int. J. Pharm. 2021, 599, 120413. [Google Scholar] [CrossRef]
- Ridha, A.A.; Kashanian, S.; Azandaryani, A.H.; Rafipour, R.; Mahdavian, E. New Folate-Modified Human Serum Albumin Conjugated to Cationic Lipid Carriers for Dual Targeting of Mitoxantrone against Breast Cancer. Curr. Pharm. Biotechnol. 2019, 21, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Gangopadhyay, S.; Nikam, R.R.; Gore, K.R. Folate Receptor-Mediated siRNA Delivery: Recent Developments and Future Directions for RNAi Therapeutics. Nucleic Acid Ther. 2021, 31, 245–270. [Google Scholar] [CrossRef] [PubMed]
- Sumohan Pillai, A.; Alexander, A.; Sri Varalakshmi, G.; Manikantan, V.; Allben Akash, B.; Enoch, I.V.M.V. Cyclodextrin and Folate Functionalized Polymer Nanocarriers: Chain Length Dependent Properties. Eur. Polym. J. 2022, 179, 111550. [Google Scholar] [CrossRef]
- Sharma, S.; Pukale, S.; Sahel, D.K.; Singh, P.; Mittal, A.; Chitkara, D. Folate Targeted Hybrid Lipo-Polymeric Nanoplexes Containing Docetaxel and miRNA-34a for Breast Cancer Treatment. Mater. Sci. Eng. C 2021, 128, 112305. [Google Scholar] [CrossRef]
- Reddy, J.A.; Clapp, D.W.; Low, P.S. Retargeting of Viral Vectors to the Folate Receptor Endocytic Pathway. J. Control. Release 2001, 74, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Lawler, S.E.; Speranza, M.C.; Cho, C.F.; Chiocca, E.A. Oncolytic Viruses in Cancer Treatment a Review. JAMA Oncol. 2017, 3, 841–849. [Google Scholar] [CrossRef] [PubMed]
- Goradel, N.H.; Negahdari, B.; Ghorghanlu, S.; Jahangiri, S.; Arashkia, A. Strategies for Enhancing Intratumoral Spread of Oncolytic Adenoviruses. Pharmacol. Ther. 2020, 213, 107586. [Google Scholar] [CrossRef]
- Ultav, G.; Tonbul, H.; Salva, E. An Effective VEGF-siRNA Delivery via Folic Acid Decorated and Pegylated Silica Nanoparticles. J. Drug Deliv. Sci. Technol. 2022, 76, 103828. [Google Scholar] [CrossRef]
- Maiyo, F.; Singh, M. Polymerized Selenium Nanoparticles for Folate-Receptor-Targeted Delivery of Anti-Luc-siRNA: Potential for Gene Silencing. Biomedicines 2020, 8, 76. [Google Scholar] [CrossRef]
- Willibald, J.; Harder, J.; Sparrer, K.; Conzelmann, K.K.; Carell, T. Click-Modified Anandamide siRNA Enables Delivery and Gene Silencing in Neuronal and Immune Cells. J. Am. Chem. Soc. 2012, 134, 12330–12333. [Google Scholar] [CrossRef]
- Thomas, M.; Kularatne, S.A.; Qi, L.; Kleindl, P.; Leamon, C.P.; Hansen, M.J.; Low, P.S. Ligand-Targeted Delivery of Small Interfering RNAs to Malignant Cells and Tissues. Ann. N. Y. Acad. Sci. 2009, 1175, 32–39. [Google Scholar] [CrossRef]
- Dohmen, C.; Fröhlich, T.; Lächelt, U.; Röhl, I.; Vornlocher, H.P.; Hadwiger, P.; Wagner, E. Defined Folate-PEG-siRNA Conjugates for Receptor-Specific Gene Silencing. Mol. Ther. Nucleic Acids 2012, 1, e7. [Google Scholar] [CrossRef]
- Salim, L.; Islam, G.; Desaulniers, J.P. Targeted Delivery and Enhanced Gene-Silencing Activity of Centrally Modified Folic Acid-siRNA Conjugates. Nucleic Acids Res. 2020, 48, 75–85. [Google Scholar] [CrossRef]
- Guo, W.; Lee, R.J. Efficient Gene Delivery via Non-Covalent Complexes of Folic Acid and Polyethylenimine. J. Control. Release 2001, 77, 131–138. [Google Scholar] [CrossRef] [PubMed]
- Kurosaki, T.; Morishita, T.; Kodama, Y.; Sato, K.; Nakagawa, H.; Higuchi, N.; Nakamura, T.; Hamamoto, T.; Sasaki, H.; Kitahara, T. Nanoparticles Electrostatically Coated with Folic Acid for Effective Gene Therapy. Mol. Pharm. 2011, 8, 913–919. [Google Scholar] [CrossRef]
- Zheng, B.; Yang, S.; Wang, M.; Yang, X.; Teng, L.; Xie, J.; Teng, L.; Lee, R.J. Non-Covalent Nanocomplexes of Folic Acid and Reducible Polyethylenimine for Survivin siRNA Delivery. Anticancer Res. 2015, 35, 5433–5442. [Google Scholar]
- Cui, S.; Zhi, D.; Zhao, Y.; Chen, H.; Meng, Y.; Zhang, C.; Zhang, S. Cationic Lioposomes with Folic Acid as Targeting Ligand for Gene Delivery. Bioorg. Med. Chem. Lett. 2016, 26, 4025–4029. [Google Scholar] [CrossRef]
- Shmendel, E.V.; Kabilova, T.O.; Morozova, N.G.; Zenkova, M.A.; Maslov, M.A. Targeted Delivery of Nucleic Acids by Folate-Containing Liposomes into KB-3-1 and HEK 293 Cells. Russ. J. Bioorg. Chem. 2019, 45, 719–725. [Google Scholar] [CrossRef]
- Aranda, C.; Urbiola, K.; Méndez Ardoy, A.; García Fernández, J.M.; Ortiz Mellet, C.; De Ilarduya, C.T. Targeted Gene Delivery by New Folate-Polycationic Amphiphilic Cyclodextrin-DNA Nanocomplexes in Vitro and in Vivo. Eur. J. Pharm. Biopharm. 2013, 85, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Puchkov, P.A.; Maslov, M.A. Lipophilic Polyamines as Promising Components of Liposomal Gene Delivery Systems. Pharmaceutics 2021, 13, 920. [Google Scholar] [CrossRef] [PubMed]
- Mikheev, A.A.; Shmendel, E.V.; Zhestovskaya, E.S.; Nazarov, G.V.; Maslov, M.A. Cationic Liposomes As Delivery Systems for Nucleic Acids. Fine Chem. Technol. 2020, 15, 7–27. [Google Scholar] [CrossRef]
- Wang, W.; Zhou, F.; Ge, L.; Liu, X.; Kong, F. A Promising Targeted Gene Delivery System: Folate-Modified Dexamethasone-Conjugated Solid Lipid Nanoparticles. Pharm. Biol. 2014, 52, 1039–1044. [Google Scholar] [CrossRef]
- Shmendel, E.; Kabilova, T.; Morozova, N.; Zenkova, M.; Maslov, M. Effects of Spacers within a Series of Novel Folate-Containing Lipoconjugates on the Targeted Delivery of Nucleic Acids. J. Drug Deliv. Sci. Technol. 2020, 57, 101609. [Google Scholar] [CrossRef]
- Bhattacharya, S.; Franz, A.; Li, X.; Jasti, B. Synthesis of Folate-Conjugated Amphiphiles for Tumor-Targeted Drug Delivery. J. Drug Target. 2008, 16, 780–789. [Google Scholar] [CrossRef]
- Poh, S.; Chelvam, V.; Ayala-López, W.; Putt, K.S.; Low, P.S. Selective Liposome Targeting of Folate Receptor Positive Immune Cells in Inflammatory Diseases. Nanomed. Nanotechnol. Biol. Med. 2018, 14, 1033–1043. [Google Scholar] [CrossRef] [PubMed]
- Ran, R.; Middelberg, A.P.J.; Zhao, C.X. Microfluidic Synthesis of Multifunctional Liposomes for Tumour Targeting. Colloids Surf. B Biointerfaces 2016, 148, 402–410. [Google Scholar] [CrossRef]
- Evans, J.C.; Malhotra, M.; Guo, J.; O’Shea, J.P.; Hanrahan, K.; O’Neill, A.; Landry, W.D.; Griffin, B.T.; Darcy, R.; Watson, R.W.; et al. Folate-Targeted Amphiphilic Cyclodextrin.siRNA Nanoparticles for Prostate Cancer Therapy Exhibit PSMA Mediated Uptake, Therapeutic Gene Silencing In Vitro and Prolonged Circulation In Vivo. Nanomed. Nanotechnol. Biol. Med. 2016, 12, 2341–2351. [Google Scholar] [CrossRef] [PubMed]
- Lohade, A.A.; Jain, R.R.; Iyer, K.; Roy, S.K.; Shimpi, H.H.; Pawar, Y.; Rajan, M.G.R.; Menon, M.D. A Novel Folate-Targeted Nanoliposomal System of Doxorubicin for Cancer Targeting. AAPS PharmSciTech 2016, 17, 1298–1311. [Google Scholar] [CrossRef]
- He, Z.; Yu, Y.; Zhang, Y.; Yan, Y.; Zheng, Y.; He, J.; Xie, Y.; He, G.; Wei, Y.; Song, X. Gene Delivery with Active Targeting to Ovarian Cancer Cells Mediated by Folate Receptor α. J. Biomed. Nanotechnol. 2013, 9, 833–844. [Google Scholar] [CrossRef]
- Hofland, H.E.J.; Masson, C.; Iginla, S.; Osetinsky, I.; Reddy, J.A.; Leamon, C.P.; Scherman, D.; Bessodes, M.; Wils, P. Folate-Targeted Gene Transfer In Vivo. Mol. Ther. 2002, 5, 739–744. [Google Scholar] [CrossRef]
- Kim, S.H.; Jeong, J.H.; Mok, H.; Lee, S.H.; Kim, S.W.; Park, T.G. Folate Receptor Targeted Delivery of Polyelectrolyte Complex Micelles Prepared from ODN-PEG-Folate Conjugate and Cationic Lipids. Biotechnol. Prog. 2007, 23, 232–237. [Google Scholar] [CrossRef]
- Lee, M.; Kim, S.W. Polyethylene Glycol-Conjugated Copolymers for Plasmid DNA Delivery. Pharm. Res. 2005, 22, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Suk, J.S.; Xu, Q.; Kim, N.; Hanes, J.; Ensign, L.M. PEGylation as a Strategy for Improving Nanoparticle-Based Drug and Gene Delivery. Adv. Drug Deliv. Rev. 2016, 99, 28–51. [Google Scholar] [CrossRef]
- Tagalakis, A.D.; Kenny, G.D.; Bienemann, A.S.; McCarthy, D.; Munye, M.M.; Taylor, H.; Wyatt, M.J.; Lythgoe, M.F.; White, E.A.; Hart, S.L. PEGylation Improves the Receptor-Mediated Transfection Efficiency of Peptide-Targeted, Self-Assembling, Anionic Nanocomplexes. J. Control. Release 2014, 174, 177–187. [Google Scholar] [CrossRef]
- Kabilova, T.; Shmendel, E.; Gladkikh, D.; Morozova, N.; Maslov, M.; Chernolovskaya, E.; Vlassov, V.; Zenkova, M. Novel PEGylated Liposomes Enhance Immunostimulating Activity of IsRNA. Molecules 2018, 23, 3101. [Google Scholar] [CrossRef] [PubMed]
- Xia, Y.; Tian, J.; Chen, X. Effect of Surface Properties on Liposomal siRNA Delivery. Biomaterials 2016, 79, 56–68. [Google Scholar] [CrossRef]
- Abu Lila, A.S.; Kiwada, H.; Ishida, T. The Accelerated Blood Clearance (ABC) Phenomenon: Clinical Challenge and Approaches to Manage. J. Control. Release 2013, 172, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Tagami, T.; Nakamura, K.; Shimizu, T.; Yamazaki, N.; Ishida, T.; Kiwada, H. CpG Motifs in pDNA-Sequences Increase Anti-PEG IgM Production Induced by PEG-Coated pDNA-Lipoplexes. J. Control. Release 2010, 142, 160–166. [Google Scholar] [CrossRef]
- Yang, Q.; Lai, S.K. Anti-PEG Immunity: Emergence, Characteristics, and Unaddressed Questions. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2015, 7, 655–677. [Google Scholar] [CrossRef]
- Gorle, S.; Ariatti, M.; Singh, M. Novel Serum-Tolerant Lipoplexes Target the Folate Receptor Efficiently. Eur. J. Pharm. Sci. 2014, 59, 83–93. [Google Scholar] [CrossRef] [PubMed]
- Hattori, Y.; Shimizu, S.; Ozaki, K.; Onishi, H. Effect of Cationic Lipid Type in Folate-PEG-Modified Cationic Liposomes on Folate Receptor-Mediated siRNA Transfection in Tumor Cells. Pharmaceutics 2019, 11, 181. [Google Scholar] [CrossRef]
- Reddy, J.A.; Abburi, C.; Hofland, H.; Howard, S.J.; Vlahov, I.; Wils, P.; Leamon, C.P. Folate-Targeted, Cationic Liposome-Mediated Gene Transfer into Disseminated Peritoneal Tumors. Gene Ther. 2002, 9, 1542–1560. [Google Scholar] [CrossRef]
- Tang, M.; Sakasai, S.; Onishi, H.; Kawano, K.; Hattori, Y. Effect of PEG Anchor in PEGylation of Folate-Modified Cationic Liposomes with PEG-Derivatives on Systemic siRNA Delivery into the Tumor. J. Drug Target. 2023, 31, 74–88. [Google Scholar] [CrossRef] [PubMed]
- Leamon, C.P.; Cooper, S.R.; Hardee, G.E. Folate-Liposome-Mediated Antisense Oligodeoxynucleotide Targeting to Cancer Cells: Evaluation In Vitro and In Vivo. Bioconjug. Chem. 2003, 14, 738–747. [Google Scholar] [CrossRef]
- Lee, R.J.; Low, P.S. Delivery of Liposomes into Cultured KB Cells via Folate Receptor-Mediated Endocytosis. J. Biol. Chem. 1994, 269, 3198–3204. [Google Scholar] [CrossRef] [PubMed]
- Darwitan, A.; Tan, Y.F.; Wong, Y.S.; Nedumaran, A.M.; Czarny, B.; Venkatraman, S. Targeting Efficiency of Nanoliposomes on Atherosclerotic Foam Cells: Polyethylene Glycol-to-Ligand Ratio Effects. Expert Opin. Drug Deliv. 2020, 17, 1165–1176. [Google Scholar] [CrossRef]
- Zhang, W.; Cui, J.; Xia, Y.; Zhang, X.; Duan, S. Synthesis of Two Folate Conjugates and Their Targeting Effect In Vitro. J China Pharm. Univ. 2021, 52, 447–454. [Google Scholar] [CrossRef]
- Yoshizawa, T.; Hattori, Y.; Hakoshima, M.; Koga, K.; Maitani, Y. Folate-Linked Lipid-Based Nanoparticles for Synthetic siRNA Delivery in KB Tumor Xenografts. Eur. J. Pharm. Biopharm. 2008, 70, 718–725. [Google Scholar] [CrossRef] [PubMed]
- Krzysztoń, R.; Salem, B.; Lee, D.J.; Schwake, G.; Wagner, E.; Rädler, J.O. Microfluidic Self-Assembly of Folate-Targeted Monomolecular siRNA-Lipid Nanoparticles. Nanoscale 2017, 9, 7442–7453. [Google Scholar] [CrossRef] [PubMed]
- Gaponova, S.; Patutina, O.; Sen’kova, A.; Burakova, E.; Savin, I.; Markov, A.; Shmendel, E.; Maslov, M.; Stetsenko, D.; Vlassov, V.; et al. Single Shot vs. Cocktail: A Comparison of Mono- and Combinative Application of miRNA-Targeted Mesyl Oligonucleotides for Efficient Antitumor Therapy. Cancers 2022, 14, 4396. [Google Scholar] [CrossRef]
- Patutina, O.A.; Gaponova, S.K.; Sen’kova, A.V.; Savin, I.A.; Gladkikh, D.V.; Burakova, E.A.; Fokina, A.A.; Maslov, M.A.; Shmendel’, E.V.; Wood, M.J.A.; et al. Mesyl Phosphoramidate Backbone Modified Antisense Oligonucleotides Targeting miR-21 with Enhanced In Vivo Therapeutic Potency. Proc. Natl. Acad. Sci. USA 2020, 117, 32370–32379. [Google Scholar] [CrossRef]
- Gladkikh, D.V.; Sen′ Kova, A.V.; Chernikov, I.V.; Kabilova, T.O.; Popova, N.A.; Nikolin, V.P.; Shmendel, E.V.; Maslov, M.A.; Vlassov, V.V.; Zenkova, M.A.; et al. Folate-Equipped Cationic Liposomes Deliver Anti-Mdr1-siRNA to the Tumor and Increase the Efficiency of Chemotherapy. Pharmaceutics 2021, 13, 1252. [Google Scholar] [CrossRef]
- Dan, N. Structure and kinetics of synthetic, lipid-based nucleic acid carriers: Lipoplexes. In Lipid Nanocarriers for Drug Targeting; Grumezescu, A.M., Ed.; William Andrew Publishing: Norwich, NY, USA, 2018; pp. 529–562. [Google Scholar] [CrossRef]
- Gaspar, R.; Coelho, F.; Silva, B.F.B. Lipid-Nucleic Acid Complexes: Physicochemical Aspects and Prospects for Cancer Treatment. Molecules 2020, 25, 5006. [Google Scholar] [CrossRef]
- Martin-Herranz, A.; Ahmad, A.; Evans, H.M.; Ewert, K.; Schulze, U.; Safinya, C.R. Surface Functionalized Cationic Lipid-DNA Complexes for Gene Delivery: PEGylated Lamellar Complexes Exhibit Distinct DNA/DNA Interaction Regimes. Biophys. J. 2004, 86, 1160–1168. [Google Scholar] [CrossRef]
- Nordling-David, M.M.; Golomb, G. Gene Delivery by Liposomes. Isr. J. Chem. 2013, 53, 737–747. [Google Scholar] [CrossRef]
- Yang, Y.; He, L.; Liu, Y.; Xia, S.; Fang, A.; Xie, Y.; Gan, L.; He, Z.; Tan, X.; Jiang, C.; et al. Promising Nanocarriers for PEDF Gene Targeting Delivery to Cervical Cancer Cells Mediated by the Over-Expressing FRα. Sci. Rep. 2016, 6, 32427. [Google Scholar] [CrossRef] [PubMed]
- He, Z.Y.; Zhang, Y.G.; Yang, Y.H.; Ma, C.C.; Wang, P.; Du, W.; Li, L.; Xiang, R.; Song, X.R.; Zhao, X.; et al. In Vivo Ovarian Cancer Gene Therapy Using CRISPR-Cas9. Hum. Gene Ther. 2018, 29, 223–233. [Google Scholar] [CrossRef]
- Li, W.; Yan, R.; Liu, Y.; He, C.; Zhang, X.; Lu, Y.; Khan, M.W.; Xu, C.; Yang, T.; Xiang, G. Co-Delivery of Bmi1 Small Interfering RNA with Ursolic Acid by Folate Receptor-Targeted Cationic Liposomes Enhances Anti-Tumor Activity of Ursolic Acid In Vitro and In Vivo. Drug Deliv. 2019, 26, 794–802. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Zhang, T.; Wu, B.; Zhang, X. Insights into the Therapeutic Potential of Hypoxia-Inducible Factor-1α Small Interfering RNA in Malignant Melanoma Delivered via Folate-Decorated Cationic Liposomes. Int. J. Nanomed. 2016, 11, 991–1002. [Google Scholar] [CrossRef]
- Hoffmann, M.; Gerlach, S.; Hoffmann, C.; Richter, N.; Hersch, N.; Csiszár, A.; Merkel, R.; Hoffmann, B. PEGylation and Folic-Acid Functionalization of Cationic Lipoplexes—Improved Nucleic Acid Transfer into Cancer Cells. Front. Bioeng. Biotechnol. 2022, 10, 1066887. [Google Scholar] [CrossRef] [PubMed]
- Metwally, A.A.; Blagbrough, I.S.; Mantell, J.M. Quantitative Silencing of EGFP Reporter Gene by Self-Assembled siRNA Lipoplexes of LinOS and Cholesterol. Mol. Pharm. 2012, 9, 3384–3395. [Google Scholar] [CrossRef] [PubMed]
- Karpuz, M.; İlhan, M.; Gültekin, H.E.; Ozgenc, E.; Şenyiğit, Z.; Atlihan-Gundogdu, E. Nanovesicles for Tumor-Targeted Drug Delivery; Academic Press: Cambridge, MA, USA, 2022; ISBN 9780323918657. [Google Scholar]
- Kumar, R.; Dkhar, D.S.; Kumari, R.; Divya; Mahapatra, S.; Srivastava, A.; Dubey, V.K.; Chandra, P. Ligand Conjugated Lipid-Based Nanocarriers for Cancer Theranostics. Biotechnol. Bioeng. 2022, 119, 3022–3043. [Google Scholar] [CrossRef] [PubMed]
- Dennahy, I.S.; Han, Z.; Maccuaig, W.M.; Chalfant, H.M.; Condacse, A.; Hagood, J.M.; Claros-Sorto, J.C.; Razaq, W.; Holter-Chakrabarty, J.; Squires, R.; et al. Nanotheranostics for Image-Guided Cancer Treatment. Pharmaceutics 2022, 14, 917. [Google Scholar] [CrossRef]
- Zheng, Z.; Li, Z.; Xu, C.; Guo, B.; Guo, P. Folate-Displaying Exosome Mediated Cytosolic Delivery of siRNA Avoiding Endosome Trapping. J. Control. Release 2019, 311–312, 43–49. [Google Scholar] [CrossRef]
- Feng, C.; Xiong, Z.; Wang, C.; Xiao, W.; Xiao, H.; Xie, K.; Chen, K.; Liang, H.; Zhang, X.; Yang, H. Folic Acid-Modified Exosome-PH20 Enhances the Efficiency of Therapy via Modulation of the Tumor Microenvironment and Directly Inhibits Tumor Cell Metastasis. Bioact. Mater. 2021, 6, 963–974. [Google Scholar] [CrossRef] [PubMed]



| Liposomal Composition | FA Lipoconjugate | Optimal Content of FA Lipoconjugate | NA | Target | Results | Ref. |
|---|---|---|---|---|---|---|
| DOPC 1–CholOH (65:35 mol.) | DSPE-PEG3400-Cys-FA | 0.03% from the range 0.01–2.0% | pDNA contained the luciferase reporter gene under the CMV promoter. | FR-positive M109 cells | 54 fluorescence units/mg cell lysate | [68] |
| RPR209120 1–DOPE (1:1 mol.) | 0.1% from the range 0.01–3.0% | FR-positive M109 cells | 1500 ng luciferase/well | |||
| 0.01% from the range 0.01–0.03% | L1210A intraperitoneal tumor model | 125 ng/mg protein | ||||
| DOTAP 1–CholOH–mPEG2000-Chol (50:45:5) | Chol-suc-PEG2000-FA | 0.1% from the range 0.01–1.0% | pEGFP | SKOV-3 | 14% cells | [55] |
| KB | 30% cells | |||||
| OH-Chol 1–DOPE (3:1 mol.) | DSPE-PEG2000-FA | 1.0% from the range 1.0–3.0% | Luc siRNA/ EGFP siRNA | KB-Luc KB-EGFP | 36% of untreated cells/ 75% EGFP expression level 4 | [67] |
| OH-C-Chol 1–DOPE (3:1 mol.) | 2.0% from the range 1.0–3.0% | 18% of untreated cells/ 65% EGFP expression level 4 | ||||
| HAPC-Chol 1–DOPE (3:1 mol.) | 2.0% from the range 1.0–3.0% | 20% of untreated cells/ 60% EGFP expression level 4 | ||||
| 2X3–DOPE (1:2 mol.) | Dg-PEG800-FA | 2.0% from the range 0.5–2.0% | FITC-ODN | KB-3-1 | 86 RFU 2 | [44,49] |
| HEK293 | 82 RFU | |||||
| MM18 1 | Diether-PEG570-FA | 10.0% from the range 2.0–10.0% | pDNA (pTG11033) | HeLa | 1 × 107 total RLU 3/mg of proteins | [23] |
| Liposomal Composition | Lipoplexes | In Vitro | In Vivo | |||||
|---|---|---|---|---|---|---|---|---|
| NA | N/P | Size (nm) | ζ-Potential (mV) | Results | Ref | Target | Ref. | |
| SGO4–DOPE–DSPE-PEG2000–DSPE-PEG2000-FA (1:0.96:0.04:0.01 mol.) | pCMV-luc | 3/1 | 147 ± 0.6 | −2.09 | 1 × 107 RLU 4/mg proteins (HeLa, KB) 1 × 106 RLU 4/mg proteins (HEK293) | [66] | n.d. 1 | |
| MM18–Diether-FA-PEG570 (9:1 wt) | pTG11033 | 1/1 | 521 | −49 | 1 × 107 RLU 4/mg proteins (HeLa, 16HBE14o) 1 × 106 RLU 4/mg proteins (A549) | [23] | Swiss mice (nasal airway epithelium) | [23] |
| DOTAP–CholOH–mPEG2000-Chol–Chol-suc-PEG2000-FA (50:45:5:0.1 mol.) | pEGFP | 5/1 2 | 199.37 ± 2.56 | 37.97 | 30% of KB, 14% of SKOV-3 | [55] | n.d. 1 | |
| pIL15 | 6/1 2 | 250–300 | 18 | n.d. 1 | CT26 colon cancer mouse model | [9] | ||
| pPEDF | 6/1 2 | 200 | 20 | 62% of HeLa | [83] | HeLa tumor xenografts | [83] | |
| gDNMT1 plasmid | 12/1 2 | 150 | 35 | 28.6% of indels (SKOV-3) | [84] | 5-fold decrease of SKOV-3 tumor xenografts weight | [84] | |
| DOTAP–CholOH–DSPE-mPEG2000– Chol-suc-PEG2000-FA (40:55:4.5:0.5 mol.) | Bmi1 siRNA | 200/1 2 | 165.1 ± 13.8 | 18.6 | 71% inhibition of KB cells | [85] | ~4-fold decrease of KB tumor xenografts | [85] |
| OH-Chol–DOPE–DSPE-PEG2000-FA (3:1 mol.:1 mol%) | PLK1 siRNA | 7/1 | 180.1 ± 2.8 3 | 35.2 | 50% of untreatment (PKL mRNA level in KB-Luc cells) | [67] | n.d. 1 | |
| OH-C-Chol–DOPE–DSPE-PEG2000-FA (3:1 mol.:2 mol%) | 188.8 ± 1.2 3 | 29.1 | 55% of untreatment (PKL mRNA level in KB-Luc cells) | n.d. 1 | ||||
| HAPC-Chol–DOPE–DSPE-PEG2000-FA (3:1 mol.:2 mol%) | 172.9 ± 1.2 3 | 28.3 | 54% of untreatment (PKL mRNA level in KB-Luc cells) | KB tumor xenografts | [67] | |||
| DDAB–DOPE–PEG1600-Chol–DSPE-PEG5000-FA (1:1 mol.:2.5 mol%:0.5 mol%) | 4/1 | 169.8 ± 1.6 | 39.3 | 10% of untreated KB-luc cells | [69] | [69] | ||
| 2X3–DOPE–Dg-PEG800-FA (1:2 mol.:2 mol%) | MDR1 siRNA | 1/1 | 175.2 ± 22.6 | −3 | n.d. 1 | L1210, LLC, KB-3-1, KB-8-5 tumor xenografts | [78] | |
| CD–DSPE-PEG5000-FA–DSPE-PEG5000-Methyl (2:1:1 mol.) | RelA siRNA | 10/1 | 207.20 ± 5.39 | 4.78 | 44% reduction for VCaP cells, 22% reduction for LNCaP cells | [53] | n.d. 1 | |
| Soybean lecithin S100–CholOH–oligochitosan derivative of FA (425 mg:75 mg:50 mg) | HIF-1α siRNA | - | 95.3 | 2.41 | 550 mean fluorescence intensity on human MM cells (A375) | [86] | n.d. 1 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shmendel, E.V.; Puchkov, P.A.; Maslov, M.A. Design of Folate-Containing Liposomal Nucleic Acid Delivery Systems for Antitumor Therapy. Pharmaceutics 2023, 15, 1400. https://doi.org/10.3390/pharmaceutics15051400
Shmendel EV, Puchkov PA, Maslov MA. Design of Folate-Containing Liposomal Nucleic Acid Delivery Systems for Antitumor Therapy. Pharmaceutics. 2023; 15(5):1400. https://doi.org/10.3390/pharmaceutics15051400
Chicago/Turabian StyleShmendel, Elena V., Pavel A. Puchkov, and Michael A. Maslov. 2023. "Design of Folate-Containing Liposomal Nucleic Acid Delivery Systems for Antitumor Therapy" Pharmaceutics 15, no. 5: 1400. https://doi.org/10.3390/pharmaceutics15051400