
The Oral Delivery System of Modified GLP-1 by Probiotics for T2DM
Abstract
:1. Introduction
2. Materials and Methods
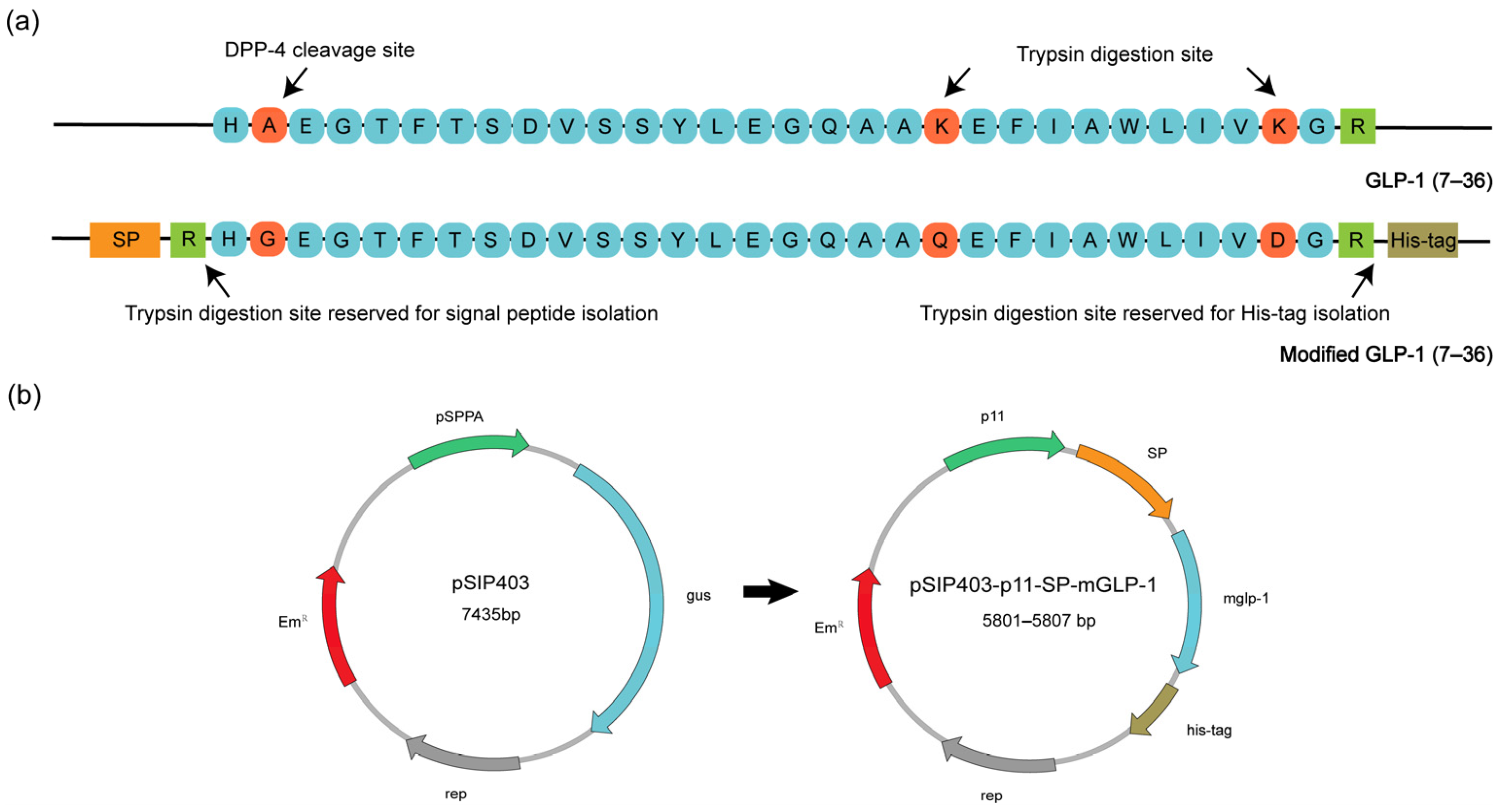
2.1. Bacterial Strains, Plasmids, and Growth Conditions
2.2. DNA Manipulation and Transformation
2.3. Expression of Secreted mGLP-1
2.4. Scanning Electron Microscopy (SEM)
2.5. Animals and Experimental Design
2.6. OGTT and Insulin Sensitivity Tests
2.7. Plasma Biochemical Parameter Measurements
2.8. Histopathological Examination and Bacterial Counts
2.9. Data Analysis
3. Results
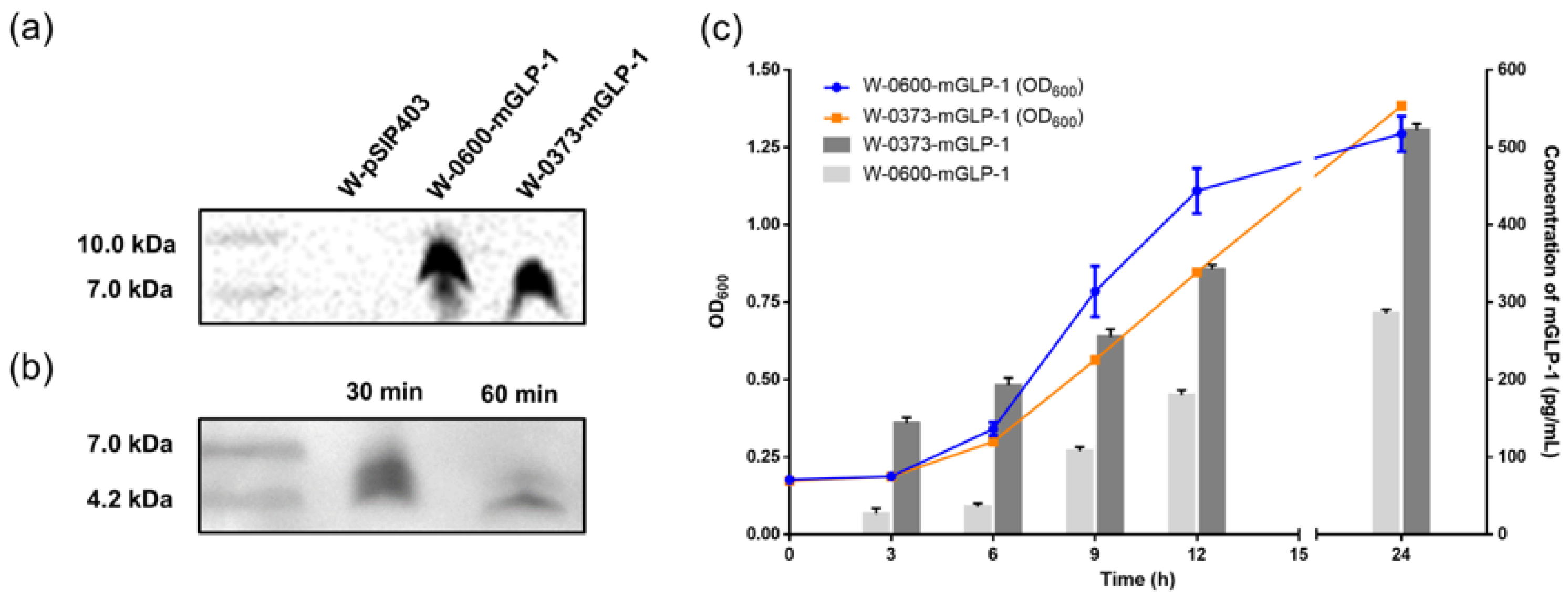
3.1. Construction and Expression of W-0373-mGLP-1
3.2. Scanning Electron Microscopy
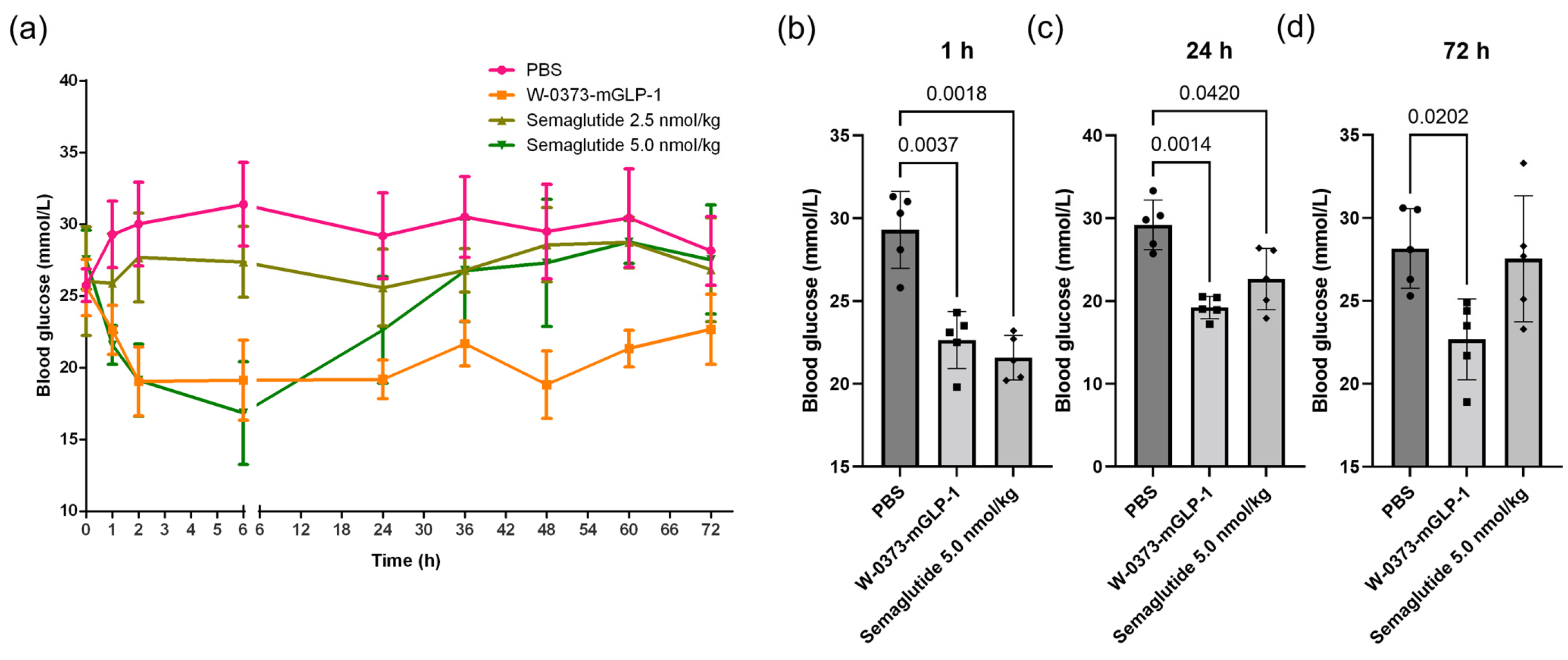
3.3. Effects of a Single Dose of W-0373-mGLP-1 and Different Doses of Semaglutide on Blood Glucose
3.4. Effects of 14-Day W-0373-mGLP-1 Administration on Blood Glucose, Random Water Intake, Fasting Blood Glucose, OGTT and ITT
3.5. Effects of 14-Day W-0373-mGLP-1 Administration on Plasma Lipid Levels
3.6. Effects of W-0373-mGLP-1 Administration on Pancreas
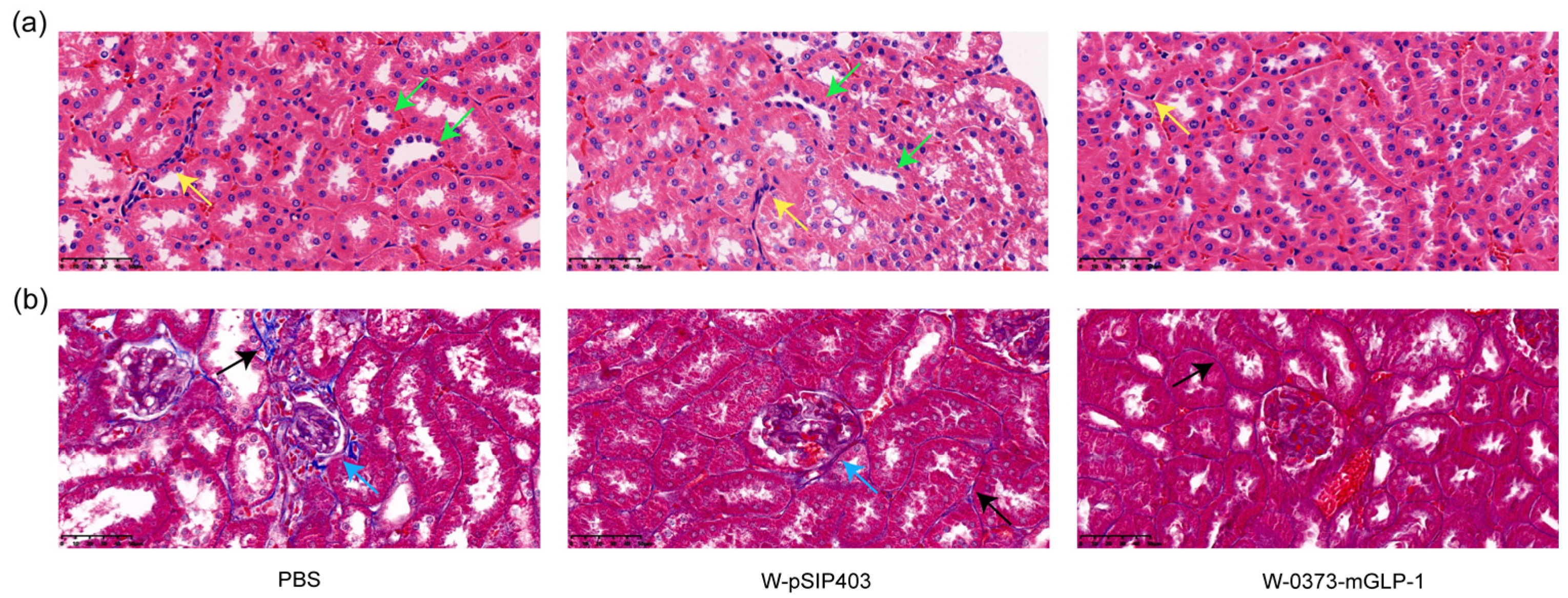
3.7. Effects of 14-Day W-0373-mGLP-1 Administration on Kidney
3.8. Colonization of L. plantarum in the Colorectum
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- IDF. IDF Diabetes Atlas, 10th ed.; International Diabetes Federation: Brussels, Belgium, 2021. [Google Scholar]
- Zhou, Z.; Sun, B.; Yu, D.; Zhu, C. Gut Microbiota: An Important Player in Type 2 Diabetes Mellitus. Front. Cell. Infect. Microbiol. 2022, 12, 834485. [Google Scholar] [CrossRef] [PubMed]
- Talchai, C.; Xuan, S.; Lin, H.V.; Sussel, L.; Accili, D. Pancreatic beta cell dedifferentiation as a mechanism of diabetic beta cell failure. Cell 2012, 150, 1223–1234. [Google Scholar] [CrossRef] [Green Version]
- Artasensi, A.; Pedretti, A.; Vistoli, G.; Fumagalli, L. Type 2 Diabetes Mellitus: A Review of Multi-Target Drugs. Molecules 2020, 25, 1987. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, E.; Lim, S.; Lamptey, R.; Webb, D.R.; Davies, M.J. Type 2 diabetes. Lancet 2022, 400, 1803–1820. [Google Scholar] [CrossRef]
- Ferguson, D.; Finck, B.N. Emerging therapeutic approaches for the treatment of NAFLD and type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2021, 17, 484–495. [Google Scholar] [CrossRef]
- Meier, J.J. GLP-1 receptor agonists for individualized treatment of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2012, 8, 728–742. [Google Scholar] [CrossRef]
- Smith, N.K.; Hackett, T.A.; Galli, A.; Flynn, C.R. GLP-1: Molecular mechanisms and outcomes of a complex signaling system. Neurochem. Int. 2019, 128, 94–105. [Google Scholar] [CrossRef]
- Deacon, C.F. Dipeptidyl peptidase 4 inhibitors in the treatment of type 2 diabetes mellitus. Nat. Rev. Endocrinol. 2020, 16, 642–653. [Google Scholar] [CrossRef] [PubMed]
- Rosenstock, J.; Wysham, C.; Frias, J.P.; Kaneko, S.; Lee, C.J.; Fernandez Lando, L.; Mao, H.; Cui, X.; Karanikas, C.A.; Thieu, V.T. Efficacy and safety of a novel dual GIP and GLP-1 receptor agonist tirzepatide in patients with type 2 diabetes (SURPASS-1): A double-blind, randomised, phase 3 trial. Lancet 2021, 398, 143–155. [Google Scholar] [CrossRef] [PubMed]
- Frias, J.P.; Davies, M.J.; Rosenstock, J.; Perez Manghi, F.C.; Fernandez Lando, L.; Bergman, B.K.; Liu, B.; Cui, X.; Brown, K.; SURPASS-2 Investigators. Tirzepatide versus Semaglutide Once Weekly in Patients with Type 2 Diabetes. N. Engl. J. Med. 2021, 385, 503–515. [Google Scholar] [CrossRef]
- Arslanian, S.A.; Hannon, T.; Zeitler, P.; Chao, L.C.; Boucher-Berry, C.; Barrientos-Perez, M.; Bismuth, E.; Dib, S.; Cho, J.I.; Cox, D.; et al. Once-Weekly Dulaglutide for the Treatment of Youths with Type 2 Diabetes. N. Engl. J. Med. 2022, 387, 433–443. [Google Scholar] [CrossRef] [PubMed]
- Jacobsen, L.V.; Flint, A.; Olsen, A.K.; Ingwersen, S.H. Liraglutide in Type 2 Diabetes Mellitus: Clinical Pharmacokinetics and Pharmacodynamics. Clin. Pharm. 2016, 55, 657–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Christou, G.A.; Katsiki, N.; Blundell, J.; Fruhbeck, G.; Kiortsis, D.N. Semaglutide as a promising antiobesity drug. Obes Rev. 2019, 20, 805–815. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Jin, T. The incretin hormone GLP-1 and mechanisms underlying its secretion. J. Diabetes 2016, 8, 753–765. [Google Scholar] [CrossRef] [Green Version]
- Bucheit, J.D.; Pamulapati, L.G.; Carter, N.; Malloy, K.; Dixon, D.L.; Sisson, E.M. Oral Semaglutide: A Review of the First Oral Glucagon-Like Peptide 1 Receptor Agonist. Diabetes Technol. 2020, 22, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Anselmo, A.C.; Gokarn, Y.; Mitragotri, S. Non-invasive delivery strategies for biologics. Nat. Rev. Drug Discov. 2019, 18, 19–40. [Google Scholar] [CrossRef]
- Pratley, R.; Amod, A.; Hoff, S.T.; Kadowaki, T.; Lingvay, I.; Nauck, M.; Pedersen, K.B.; Saugstrup, T.; Meier, J.J.; PIONEER 4 Investigators. Oral semaglutide versus subcutaneous liraglutide and placebo in type 2 diabetes (PIONEER 4): A randomised, double-blind, phase 3a trial. Lancet 2019, 394, 39–50. [Google Scholar] [CrossRef]
- Mathur, H.; Beresford, T.P.; Cotter, P.D. Health Benefits of Lactic Acid Bacteria (LAB) Fermentates. Nutrients 2020, 12, 1679. [Google Scholar] [CrossRef]
- GRAS Notices. Available online: https://www.accessdata.fda.gov/scripts/fdcc/?set=GRASNotices (accessed on 28 March 2023).
- Gareau, M.G.; Sherman, P.M.; Walker, W.A. Probiotics and the gut microbiota in intestinal health and disease. Nat. Rev. Gastroenterol. Hepatol. 2010, 7, 503–514. [Google Scholar] [CrossRef] [Green Version]
- Garbacz, K. Anticancer activity of lactic acid bacteria. Semin. Cancer Biol. 2022, 86, 356–366. [Google Scholar] [CrossRef]
- Kamiya, T.; Watanabe, Y.; Makino, S.; Kano, H.; Tsuji, N.M. Improvement of Intestinal Immune Cell Function by Lactic Acid Bacteria for Dairy Products. Microorganisms 2016, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Riaz Rajoka, M.S.; Shi, J.; Zhu, J.; Shao, D.; Huang, Q.; Yang, H.; Jin, M. Capacity of lactic acid bacteria in immunity enhancement and cancer prevention. Appl. Microbiol. Biotechnol. 2017, 101, 35–45. [Google Scholar] [CrossRef]
- Ivanovic, N.; Minic, R.; Dimitrijevic, L.; Radojevic Skodric, S.; Zivkovic, I.; Djordjevic, B. Lactobacillus rhamnosus LA68 and Lactobacillus plantarum WCFS1 differently influence metabolic and immunological parameters in high fat diet-induced hypercholesterolemia and hepatic steatosis. Food Funct. 2015, 6, 558–565. [Google Scholar] [CrossRef] [PubMed]
- Lee, E.; Jung, S.R.; Lee, S.Y.; Lee, N.K.; Paik, H.D.; Lim, S.I. Lactobacillus plantarum Strain Ln4 Attenuates Diet-Induced Obesity, Insulin Resistance, and Changes in Hepatic mRNA Levels Associated with Glucose and Lipid Metabolism. Nutrients 2018, 10, 643. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.; Jin, G.; Pang, X.; Mo, Q.; Bao, J.; Liu, T.; Wu, J.; Xie, R.; Liu, X.; Liu, J.; et al. Lactobacillus rhamnosus GG colonization in early life regulates gut-brain axis and relieves anxiety-like behavior in adulthood. Pharm. Res. 2022, 177, 106090. [Google Scholar] [CrossRef] [PubMed]
- Valeur, N.; Engel, P.; Carbajal, N.; Connolly, E.; Ladefoged, K. Colonization and immunomodulation by Lactobacillus reuteri ATCC 55730 in the human gastrointestinal tract. Appl. Environ. Microbiol. 2004, 70, 1176–1181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wells, J.M.; Mercenier, A. Mucosal delivery of therapeutic and prophylactic molecules using lactic acid bacteria. Nat. Rev. Microbiol. 2008, 6, 349–362. [Google Scholar] [CrossRef] [PubMed]
- Welman, A.D.; Maddox, I.S. Exopolysaccharides from lactic acid bacteria: Perspectives and challenges. Trends Biotechnol. 2003, 21, 269–274. [Google Scholar] [CrossRef] [PubMed]
- De Filippis, F.; Pasolli, E.; Ercolini, D. The food-gut axis: Lactic acid bacteria and their link to food, the gut microbiome and human health. FEMS Microbiol. Rev. 2020, 44, 454–489. [Google Scholar] [CrossRef]
- Mobergslien, A.; Vasovic, V.; Mathiesen, G.; Fredriksen, L.; Westby, P.; Eijsink, V.G.; Peng, Q.; Sioud, M. Recombinant Lactobacillus plantarum induces immune responses to cancer testis antigen NY-ESO-1 and maturation of dendritic cells. Hum. Vaccin. Immunother. 2015, 11, 2664–2673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skrlec, K.; Rucman, R.; Jarc, E.; Sikiric, P.; Svajger, U.; Petan, T.; Perisic Nanut, M.; Strukelj, B.; Berlec, A. Engineering recombinant Lactococcus lactis as a delivery vehicle for BPC-157 peptide with antioxidant activities. Appl. Microbiol. Biotechnol. 2018, 102, 10103–10117. [Google Scholar] [CrossRef]
- Sak-Ubol, S.; Namvijitr, P.; Pechsrichuang, P.; Haltrich, D.; Nguyen, T.H.; Mathiesen, G.; Eijsink, V.G.; Yamabhai, M. Secretory production of a beta-mannanase and a chitosanase using a Lactobacillus plantarum expression system. Microb. Cell. Fact. 2016, 15, 81. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Braat, H.; Rottiers, P.; Hommes, D.W.; Huyghebaert, N.; Remaut, E.; Remon, J.P.; van Deventer, S.J.; Neirynck, S.; Peppelenbosch, M.P.; Steidler, L. A phase I trial with transgenic bacteria expressing interleukin-10 in Crohn’s disease. Clin. Gastroenterol. Hepatol. 2006, 4, 754–759. [Google Scholar] [CrossRef] [PubMed]
- Kuwahara, A.; Arita, M.; Kushiro, A.; Sakube, Y.; Sisido, M.; Ohtsuki, T. Lactobacillus-mediated RNA interference in nematode. J. Biosci. Bioeng. 2010, 109, 189–192. [Google Scholar] [CrossRef]
- Kleerebezem, M.; Boekhorst, J.; van Kranenburg, R.; Molenaar, D.; Kuipers, O.P.; Leer, R.; Tarchini, R.; Peters, S.A.; Sandbrink, H.M.; Fiers, M.W.; et al. Complete genome sequence of Lactobacillus plantarum WCFS1. Proc. Natl. Acad. Sci. USA 2003, 100, 1990–1995. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Siezen, R.J.; Francke, C.; Renckens, B.; Boekhorst, J.; Wels, M.; Kleerebezem, M.; van Hijum, S.A. Complete resequencing and reannotation of the Lactobacillus plantarum WCFS1 genome. J. Bacteriol. 2012, 194, 195–196. [Google Scholar] [CrossRef] [Green Version]
- Sorvig, E.; Mathiesen, G.; Naterstad, K.; Eijsink, V.G.H.; Axelsson, L. High-level, inducible gene expression in Lactobacillus sakei and Lactobacillus plantarum using versatile expression vectors. Microbiology 2005, 151, 2439–2449. [Google Scholar] [CrossRef] [Green Version]
- Karlskas, I.L.; Maudal, K.; Axelsson, L.; Rud, I.; Eijsink, V.G.; Mathiesen, G. Heterologous protein secretion in Lactobacilli with modified pSIP vectors. PLoS ONE 2014, 9, e91125. [Google Scholar] [CrossRef] [Green Version]
- Rud, I.; Jensen, P.R.; Naterstad, K.; Axelsson, L. A synthetic promoter library for constitutive gene expression in Lactobacillus plantarum. Microbiology 2006, 152, 1011–1019. [Google Scholar] [CrossRef] [Green Version]
- Mathiesen, G.; Sveen, A.; Piard, J.C.; Axelsson, L.; Eijsink, V.G. Heterologous protein secretion by Lactobacillus plantarum using homologous signal peptides. J. Appl. Microbiol. 2008, 105, 215–226. [Google Scholar] [CrossRef]
- Mathiesen, G.; Sveen, A.; Brurberg, M.B.; Fredriksen, L.; Axelsson, L.; Eijsink, V.G. Genome-wide analysis of signal peptide functionality in Lactobacillus plantarum WCFS1. BMC Genom. 2009, 10, 425. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Luo, L.; Dong, C.; Zheng, X.; Guo, B.; Xia, Y.; Tao, L.; Ai, L. Polysaccharides can improve the survival of Lactiplantibacillus plantarum subjected to freeze-drying. J. Dairy Sci. 2021, 104, 2606–2614. [Google Scholar] [CrossRef] [PubMed]
- Marco, M.L.; Peters, T.H.; Bongers, R.S.; Molenaar, D.; van Hemert, S.; Sonnenburg, J.L.; Gordon, J.I.; Kleerebezem, M. Lifestyle of Lactobacillus plantarum in the mouse caecum. Environ. Microbiol. 2009, 11, 2747–2757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Bokhorst-van de Veen, H.; van Swam, I.; Wels, M.; Bron, P.A.; Kleerebezem, M. Congruent strain specific intestinal persistence of Lactobacillus plantarum in an intestine-mimicking in vitro system and in human volunteers. PLoS ONE 2012, 7, e44588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferreira-Lazarte, A.; Plaza-Vinuesa, L.; de Las Rivas, B.; Villamiel, M.; Munoz, R.; Moreno, F.J. Production of alpha-rhamnosidases from Lactobacillus plantarum WCFS1 and their role in deglycosylation of dietary flavonoids naringin and rutin. Int. J. Biol. Macromol. 2021, 193, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Sasikumar, P.; Gomathi, S.; Anbazhagan, K.; Selvam, G.S. Secretion of biologically active heterologous oxalate decarboxylase (OxdC) in Lactobacillus plantarum WCFS1 using homologous signal peptides. Biomed. Res. Int. 2013, 2013, 280432. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sorvig, E.; Gronqvist, S.; Naterstad, K.; Mathiesen, G.; Eijsink, V.G.; Axelsson, L. Construction of vectors for inducible gene expression in Lactobacillus sakei and L plantarum. FEMS Microbiol. Lett. 2003, 229, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Dower, W.J.; Miller, J.F.; Ragsdale, C.W. High efficiency transformation of E. coli by high voltage electroporation. Nucleic. Acids Res. 1988, 16, 6127–6145. [Google Scholar] [CrossRef] [Green Version]
- van den Nieuwboer, M.; van Hemert, S.; Claassen, E.; de Vos, W.M. Lactobacillus plantarum WCFS1 and its host interaction: A dozen years after the genome. Microb. Biotechnol. 2016, 9, 452–465. [Google Scholar] [CrossRef] [Green Version]
- Omori, K.; Nakamura, A.; Miyoshi, H.; Yamauchi, Y.; Kawata, S.; Takahashi, K.; Kitao, N.; Nomoto, H.; Kameda, H.; Cho, K.Y.; et al. Glucokinase Inactivation Paradoxically Ameliorates Glucose Intolerance by Increasing beta-Cell Mass in db/db Mice. Diabetes 2021, 70, 917–931. [Google Scholar] [CrossRef]
- Katz, A.; Nambi, S.S.; Mather, K.; Baron, A.D.; Follmann, D.A.; Sullivan, G.; Quon, M.J. Quantitative insulin sensitivity check index: A simple, accurate method for assessing insulin sensitivity in humans. J. Clin. Endocrinol. Metab. 2000, 85, 2402–2410. [Google Scholar] [CrossRef] [PubMed]
- Lin, Y.; Krogh-Andersen, K.; Pelletier, J.; Marcotte, H.; Ostenson, C.G.; Hammarstrom, L. Oral Delivery of Pentameric Glucagon-Like Peptide-1 by Recombinant Lactobacillus in Diabetic Rats. PLoS ONE 2016, 11, e0162733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.Z.W.; Huan, H.; Xia, W.; Chen, Y.; Wang, P.; Liu, Y. Construction of an Integrated mCherry Red Fluorescent Protein Expression System for Labeling and Tracing in Lactiplantibacillus plantarum WCFS1. Front. Microbiol. 2021, 22, 690270. [Google Scholar] [CrossRef] [PubMed]
- Tauer, C.; Heinl, S.; Egger, E.; Heiss, S.; Grabherr, R. Tuning constitutive recombinant gene expression in Lactobacillus plantarum. Microb. Cell Fact. 2014, 13, 150. [Google Scholar] [CrossRef]
- Zeng, Z.; Yu, R.; Zuo, F.; Zhang, B.; Peng, D.; Ma, H.; Chen, S. Heterologous Expression and Delivery of Biologically Active Exendin-4 by Lactobacillus paracasei L14. PLoS ONE 2016, 11, e0165130. [Google Scholar] [CrossRef] [Green Version]
- Monnier, L.; Lapinski, H.; Colette, C. Contributions of fasting and postprandial plasma glucose increments to the overall diurnal hyperglycemia of type 2 diabetic patients: Variations with increasing levels of HbA(1c). Diabetes Care 2003, 26, 881–885. [Google Scholar] [CrossRef] [Green Version]
- de Boer, S.A.; Lefrandt, J.D.; Petersen, J.F.; Boersma, H.H.; Mulder, D.J.; Hoogenberg, K. The effects of GLP-1 analogues in obese, insulin-using type 2 diabetes in relation to eating behaviour. Int. J. Clin. Pharm. 2016, 38, 144–151. [Google Scholar] [CrossRef] [Green Version]
- Bahiru, E.; Hsiao, R.; Phillipson, D.; Watson, K.E. Mechanisms and Treatment of Dyslipidemia in Diabetes. Curr. Cardiol. Rep. 2021, 23, 26. [Google Scholar] [CrossRef]
- Marshall, S.M. The pancreas in health and in diabetes. Diabetologia 2020, 63, 1962–1965. [Google Scholar] [CrossRef]
- Gromada, J.; Chabosseau, P.; Rutter, G.A. The alpha-cell in diabetes mellitus. Nat. Rev. Endocrinol. 2018, 14, 694–704. [Google Scholar] [CrossRef]
- Braunwald, E. Diabetes, heart failure, and renal dysfunction: The vicious circles. Prog. Cardiovasc. Dis. 2019, 62, 298–302. [Google Scholar] [CrossRef] [PubMed]
- Karczewski, J.; Troost, F.J.; Konings, I.; Dekker, J.; Kleerebezem, M.; Brummer, R.J.; Wells, J.M. Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest. Liver Physiol. 2010, 298, G851–G859. [Google Scholar] [CrossRef] [Green Version]
- Park, Y.E.; Kim, M.S.; Shim, K.W.; Kim, Y.I.; Chu, J.; Kim, B.K.; Choi, I.S.; Kim, J.Y. Effects of Lactobacillus plantarum Q180 on Postprandial Lipid Levels and Intestinal Environment: A Double-Blind, Randomized, Placebo-Controlled, Parallel Trial. Nutrients 2020, 12, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donnelly, D. The structure and function of the glucagon-like peptide-1 receptor and its ligands. Br. J. Pharm. 2012, 166, 27–41. [Google Scholar] [CrossRef] [Green Version]
- Ozempic (semaglutide) Injection. 2018. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/nda/2017/209637Orig1s000TOC.cfm (accessed on 28 March 2023).
- Nauck, M.A.; Quast, D.R.; Wefers, J.; Meier, J.J. GLP-1 receptor agonists in the treatment of type 2 diabetes—State-of-the-art. Mol. Metab. 2021, 46, 101102. [Google Scholar] [CrossRef]
- Johnson, E.C.; Bardis, C.N.; Jansen, L.T.; Adams, J.D.; Kirkland, T.W.; Kavouras, S.A. Reduced water intake deteriorates glucose regulation in patients with type 2 diabetes. Nutr. Res. 2017, 43, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Wu, L.; Parhofer, K.G. Diabetic dyslipidemia. Metabolism 2014, 63, 1469–1479. [Google Scholar] [CrossRef]
- Nauck, M.A.; Wefers, J.; Meier, J.J. Treatment of type 2 diabetes: Challenges, hopes, and anticipated successes. Lancet Diabetes Endocrinol. 2021, 9, 525–544. [Google Scholar] [CrossRef]
- Tomic, D.; Shaw, J.E.; Magliano, D.J. The burden and risks of emerging complications of diabetes mellitus. Nat. Rev. Endocrinol. 2022, 18, 525–539. [Google Scholar] [CrossRef]
- Iorga, R.A.; Bacalbasa, N.; Carsote, M.; Bratu, O.G.; Stanescu, A.M.A.; Bungau, S.; Pantis, C.; Diaconu, C.C. Metabolic and cardiovascular benefits of GLP-1 agonists, besides the hypoglycemic effect (Review). Exp. Med. 2020, 20, 2396–2400. [Google Scholar] [CrossRef]
- Xu, J.; Wang, J.; Cheng, Y.; Li, X.; He, M.; Zhu, J.; Han, H.; Wei, G.; Kong, H.; Xie, W.; et al. Glucagon-Like Peptide-1 Mediates the Protective Effect of the Dipeptidyl Peptidase IV Inhibitor on Renal Fibrosis via Reducing the Phenotypic Conversion of Renal Microvascular Cells in Monocrotaline-Treated Rats. Biomed. Res. Int 2018, 2018, 1864107. [Google Scholar] [CrossRef] [Green Version]
- Kawanami, D.; Takashi, Y. GLP-1 Receptor Agonists in Diabetic Kidney Disease: From Clinical Outcomes to Mechanisms. Front. Pharm. 2020, 11, 967. [Google Scholar] [CrossRef] [PubMed]
- Fang, C.; Kim, H.; Yanagisawa, L.; Bennett, W.; Sirven, M.A.; Alaniz, R.C.; Talcott, S.T.; Mertens-Talcott, S.U. Gallotannins and Lactobacillus plantarum WCFS1 Mitigate High-Fat Diet-Induced Inflammation and Induce Biomarkers for Thermogenesis in Adipose Tissue in Gnotobiotic Mice. Mol. Nutr. Food Res. 2019, 63, e1800937. [Google Scholar] [CrossRef]
- Plavec, T.V.; Berlec, A. Engineering of lactic acid bacteria for delivery of therapeutic proteins and peptides. Appl. Microbiol. Biotechnol. 2019, 103, 2053–2066. [Google Scholar] [CrossRef] [PubMed]
- Corb Aron, R.A.; Abid, A.; Vesa, C.M.; Nechifor, A.C.; Behl, T.; Ghitea, T.C.; Munteanu, M.A.; Fratila, O.; Andronie-Cioara, F.L.; Toma, M.M.; et al. Recognizing the Benefits of Pre-/Probiotics in Metabolic Syndrome and Type 2 Diabetes Mellitus Considering the Influence of Akkermansia muciniphila as a Key Gut Bacterium. Microorganisms 2021, 9, 618. [Google Scholar] [CrossRef] [PubMed]
- Mao, R.; Wu, D.; Hu, S.; Zhou, K.; Wang, M.; Wang, Y. Secretory expression and surface display of a new and biologically active single-chain insulin (SCI-59) analog by lactic acid bacteria. Appl. Microbiol. Biotechnol. 2017, 101, 3259–3271. [Google Scholar] [CrossRef]
- Agarwal, P.; Khatri, P.; Billack, B.; Low, W.K.; Shao, J. Oral delivery of glucagon like peptide-1 by a recombinant Lactococcus lactis. Pharm Res. 2014, 31, 3404–3414. [Google Scholar] [CrossRef]
- Ahn, S.; Lee, I.H.; Lee, E.; Kim, H.; Kim, Y.C.; Jon, S. Oral delivery of an anti-diabetic peptide drug via conjugation and complexation with low molecular weight chitosan. J. Control. Release 2013, 170, 226–232. [Google Scholar] [CrossRef] [PubMed]
- Shrestha, N.; Bouttefeux, O.; Vanvarenberg, K.; Lundquist, P.; Cunarro, J.; Tovar, S.; Khodus, G.; Andersson, E.; Keita, A.V.; Gonzalez Dieguez, C.; et al. The stimulation of GLP-1 secretion and delivery of GLP-1 agonists via nanostructured lipid carriers. Nanoscale 2018, 10, 603–613. [Google Scholar] [CrossRef]
- Araujo, F.; Shrestha, N.; Shahbazi, M.A.; Fonte, P.; Makila, E.M.; Salonen, J.J.; Hirvonen, J.T.; Granja, P.L.; Santos, H.A.; Sarmento, B. The impact of nanoparticles on the mucosal translocation and transport of GLP-1 across the intestinal epithelium. Biomaterials 2014, 35, 9199–9207. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Plasmids/Strains | Description | Source |
|---|---|---|
| Plasmids | ||
| pSIP403 | EmR, containing the pSPPA promoter | [49] |
| pSIP403-p11-0600-mGLP-1 | pSIP403 carrying promoter p11, signal peptide Lp_0600 and modified glp-1 gene | This study |
| pSIP403-p11-0373-mGLP-1 | pSIP403 carrying promoter p11, signal peptide Lp_0373 and modified glp-1 gene | This study |
| Strains | ||
| Escherichia coli (E. coli) DH5α | Host strain | [50] |
| Lactobacillus plantarum (L. plantarum) WCFS1 | Host strain | [37,51] |
| W-pSIP403 | L. plantarum WCFS1 containing empty plasmid pSIP403 | This study |
| W-0600-mGLP-1 | L. plantarum WCFS1 containing plasmid pSIP403-p11-0600-mGLP-1 | This study |
| W-0373-mGLP-1 | L. plantarum WCFS1 containing plasmid pSIP403-p11-0373-mGLP-1 | This study |
| Technique | Details of Formulation | Antidiabetic Peptides | Advantages | Reference |
|---|---|---|---|---|
| Permeation enhancer | sodium N-(8-[2-hydroxybenzoyl] amino) caprylate (SNAC) | Semaglutide | Promoting the transport of polypeptides across the gastrointestinal biofilm | [18] |
| Mucoadhesive polymer | Chitosan | Exendin-4 | Mucosal adhesion properties, high stability against enzymatic degradation | [81] |
| Nanostructured lipid carrier | Solid lipid | GLP-1/Exendin-4 | Activating the endogenous GLP-1 secretion, site-specific release | [82] |
| Nanoparticle | Chitosan—poly(lactide-co-glycolide) polymer | GLP-1 | Protecting peptides from degradation and promoting the permeation of GLP-1 across the intestinal | [83] |
| Lactic acid bacteria (LAB) delivery system | LAB | GLP-1/Exendin-4 | Functioning as a vehicle both for expression and delivery, bio-safety, colorectal colonization ability, intestinal transport capacity, treatment-associated probiotic effects | [57,80] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Q.; Guo, H.; Mao, W.; Qian, X.; Liu, Y. The Oral Delivery System of Modified GLP-1 by Probiotics for T2DM. Pharmaceutics 2023, 15, 1202. https://doi.org/10.3390/pharmaceutics15041202
Wang Q, Guo H, Mao W, Qian X, Liu Y. The Oral Delivery System of Modified GLP-1 by Probiotics for T2DM. Pharmaceutics. 2023; 15(4):1202. https://doi.org/10.3390/pharmaceutics15041202
Chicago/Turabian StyleWang, Qing, Haixin Guo, Wenwei Mao, Xiuping Qian, and Yangang Liu. 2023. "The Oral Delivery System of Modified GLP-1 by Probiotics for T2DM" Pharmaceutics 15, no. 4: 1202. https://doi.org/10.3390/pharmaceutics15041202