
ApoE Mimetic Peptides to Improve the Vicious Cycle of Malnutrition and Enteric Infections by Targeting the Intestinal and Blood-Brain Barriers
 , , ,
, , ,
Abstract
:1. Introduction
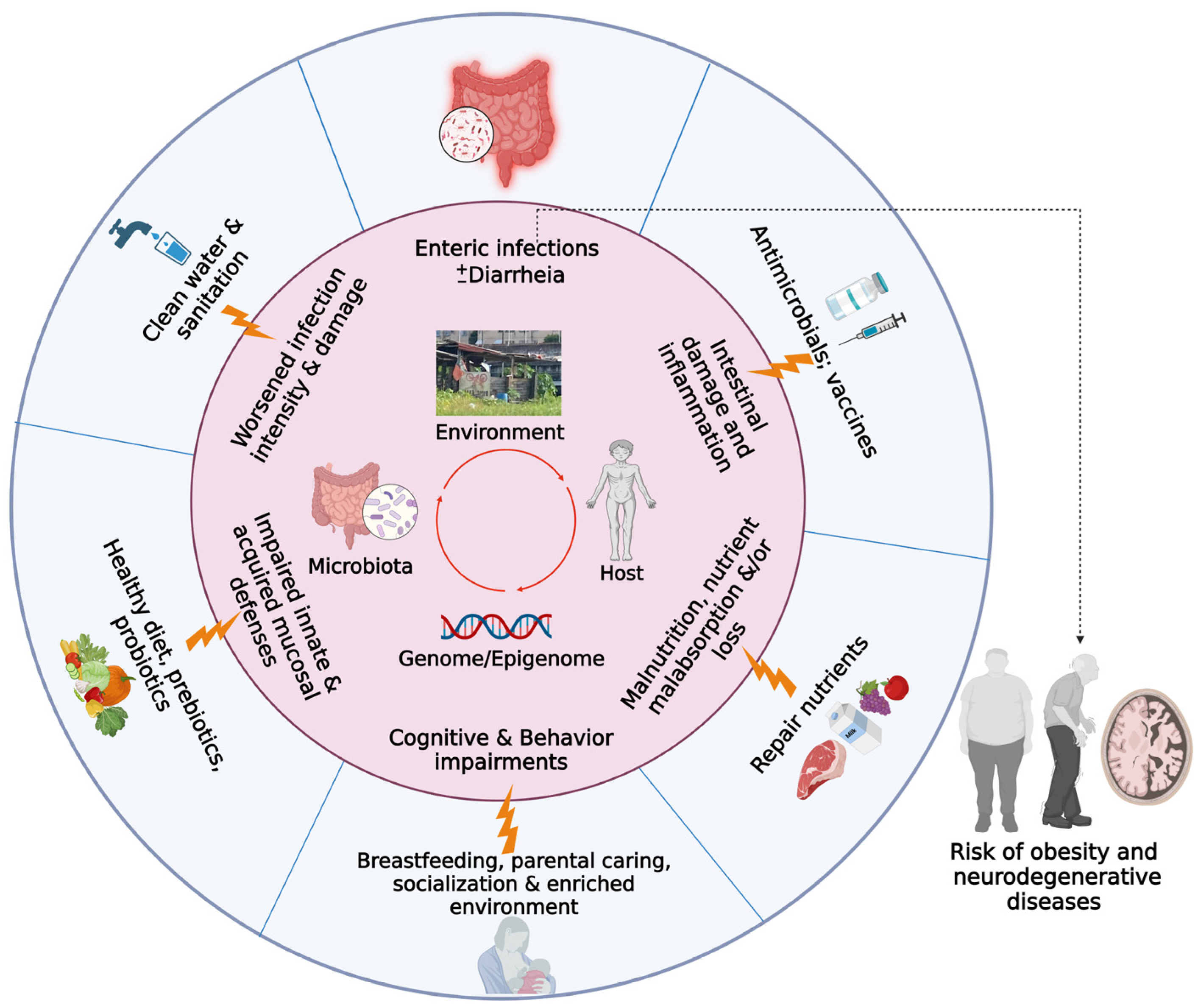
2. Vicious Cycle of Malnutrition and Enteric Diseases
2.1. Prolonged Malnutrition/Enteric Infections Early in Life as a Cause of Environmental Enteric Dysfunction: Mechanisms and Burdens of Disease
2.2. Effect of Enteropathy and Gut Dysbiosis on Neurological/Cognitive Outcomes
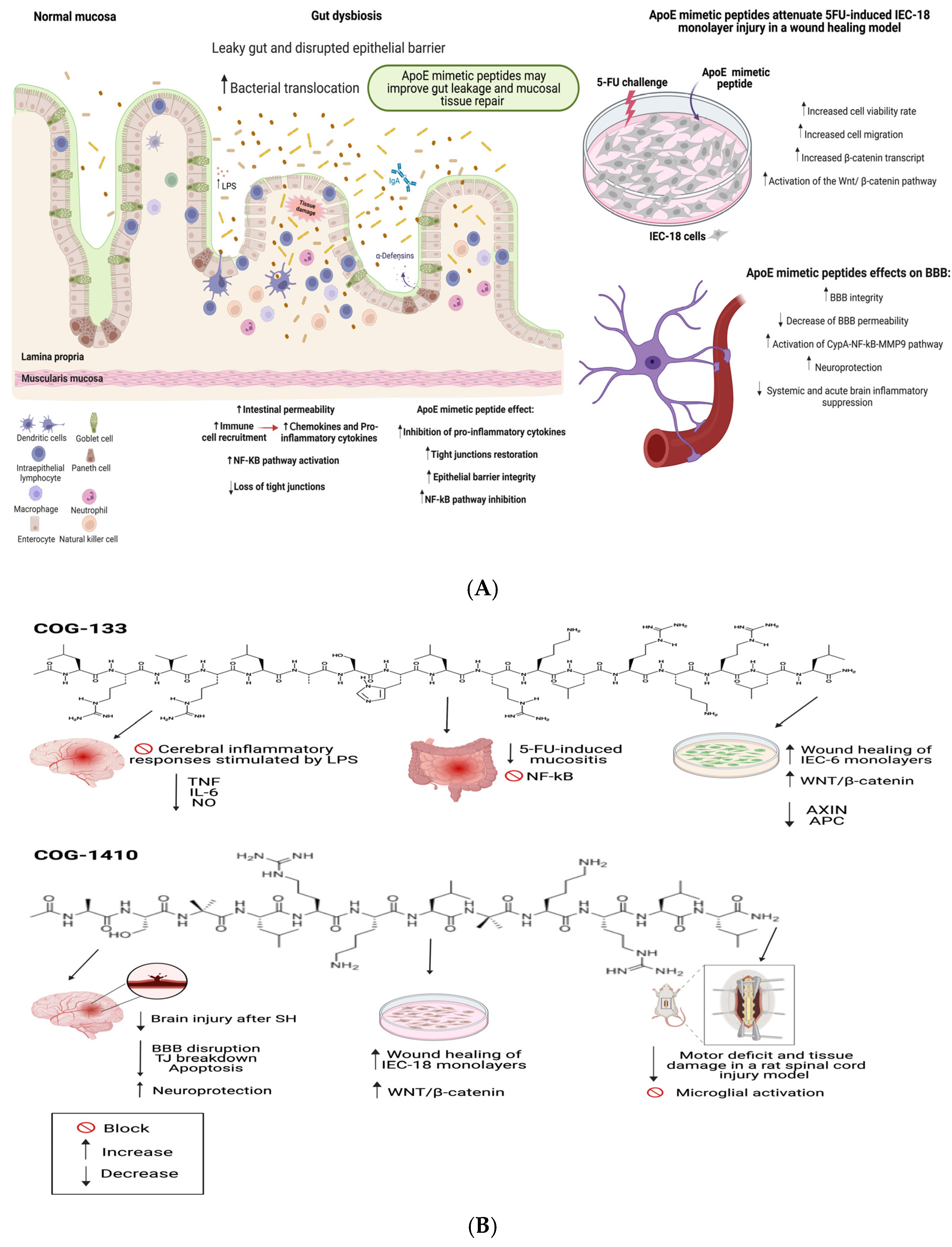
3. Effect of ApoE Mimetic Peptides on Intestinal and Brain Injuries
3.1. ApoE and ApoE Mimetic Peptides Biology
3.2. The Role of ApoE and ApoE Mimetic Peptides on the Intestinal Barrier, Microbiota and Inflammation
3.3. The Role of ApoE and ApoE Mimetic Peptides in the Blood-Brain Barrier and Neuroprotection

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Authors | ApoE Mimetic Peptides | Effects on the Small Intestine or Brain/Blood Brain Barrier | Mechanism of Action |
|---|---|---|---|
| Laskowitz et al. [94] | COG133 | Suppresses inflammatory responses | Inhibits the production of TNF and nitric oxide stimulated by LPS in microglial cells of ApoE-deficient mouse pups |
| Lynch et al. [95] | COG133 | Modifies systemic and cerebral inflammatory responses | Cerebral and systemic elevations of TNF and IL-6 were identified in APOE4-TR mice after intravenous LPS injection. Improved expression of these cytokines when treated with the mimetic peptide |
| Azevedo et al. [96] | COG 133 | Displays anti-inflammatory effects | Recovery of viability and migration of intestinal cells, reduction of intestinal myeloperoxidase levels and expression of IL-1β, TNF, iNOS mRNAs with improvement in IL-10 in a model of intestinal mucositis in mice |
| Singh et al. [100] | COG112 | Attenuates colon inflammation | Inhibits bacterial-induced expression of iNOS mRNA and protein and NO production in colonic epithelial cells using rodent model studies |
| Pessoa et al. [128] | COG1410 | Improves wound healing of IEC-18 monolayers | Restores intestinal IEC-18 cell monolayer after 5-FU injury through indirect activation of the Wnt/β-catenin pathway |
| Pessoa et al. [129] | COG133 | Improves wound healing of IEC-6 monolayers | Injury of intestinal IEC-6 cells by 5-FU resulted in decreased levels of APC transcript and Wnt/β-catenin pathway, which were recovered with mimetic peptide treatment |
| Wu et al. [144] | COG1410 | Attenuating tissue damage and reducing inflammation | Lessens neuronal apoptosis, microglia activation, and BBB leakage resulting in an increase of cerebral glucose uptake and promoting neurocognitive dysfunction attenuation in mice subjected to a subarachnoid hemorrhage model |
| Pang et al. [146] | COG1410 | Reduces early brain injury | Attenuates BBB disruption, causing less tight junction breakdown and vacuolization between endothelial cells, less endothelial cell apoptosis, in an experimental subarachnoid hemorrhage (SAH) model in mice |
| Qin et al. [147] | COG1410 | Displays neuroprotective effects | Increases cerebral glucose uptake with recovery of vestibulomotor deficits associated with rescued BBB |
| Li et al. [149] | COG1410 | Displays neuroprotective effects | In a model of neuronal autophagy after SAH, affecting the phosphorylation of GSK-3β in neuronal and brain tissue, resulting in an improvement in the neurological score |
| Wang et al. [153] | COG1410 | Attenuates tissue damage and reducing inflammation | Reduces cerebral edema, inflammation, and infarct area, also decreasing TNF RNA expression using a murine model of transient focal cerebral ischemia and reperfusion. |
| Zhang et al. [154] | COG133 | Plays a neuroprotective role by protecting blood-brain barrier function and inhibiting brain cell pyroptosis | Inhibits blood-brain barrier impairment through the proinflammatory CypA-NF-κB-MMP9 pathway and reduces neuronal pyroptosis by inhibiting NLRP3 inflammasome activation. |
| Liu et al. [156] | CN-105 | Reduces cerebral vasospasm and anti-inflammatory effect | Likely due to a reduction of FJB-positive cells, microgliosis, and an increase in the number of NeuN-positive cells in the hippocampus |
| Christensen et al. [157] | COG1410 | Reduces inflammatory response | Modulates positively protein phosphatase 2A (PP2A) activity, with an antagonistic action on SET binding, signaling by decreasing activation levels of phosphorylated signaling kinases |
| Zhao et al. [158] | COG1410 | Protects against brain injury-induced apoptosis and recovers learning and memory function | Improvements via extracellular signal-regulated mitochondrial apoptotic mechanism and Bax kinase1/2-Bax, attenuating the apoptotic signaling pathway by downregulating Bax protein and cytochrome C expression |
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Moreau, G.B.; Ramakrishnan, G.; Cook, H.L.; Fox, T.E.; Nayak, U.; Ma, J.Z.; Colgate, E.R.; Kirkpatrick, B.D.; Haque, R.; Petri, W.A.J. Childhood growth and neurocognition are associated with distinct sets of metabolites. EBioMedicine 2019, 44, 597–606. [Google Scholar] [CrossRef] [Green Version]
- Rogawski McQuade, E.T.; Scharf, R.J.; Svensen, E.; Huggins, A.; Maphula, A.; Bayo, E.; Blacy, L.; de Souza, P.P.E.; Costa, H.; Houpt, E.R.; et al. Impact of Shigella infections and inflammation early in life on child growth and school-aged cognitive outcomes: Findings from three birth cohorts over eight years. PLoS Negl. Trop. Dis. 2022, 16, e0010722. [Google Scholar] [CrossRef] [PubMed]
- MAL-ED Network Investigators. Early childhood cognitive development is affected by interactions among illness, diet, enteropathogens and the home environment: Findings from the MAL-ED birth cohort study. BMJ Glob. Health 2018, 3, e000752. [Google Scholar] [CrossRef]
- Oriá, R.B.; Murray-Kolb, L.E.; Scharf, R.J.; Pendergast, L.L.; Lang, D.R.; Kolling, G.L.; Guerrant, R.L. Early-life enteric infections: Relation between chronic systemic inflammation and poor cognition in children. Nutr. Rev. 2016, 74, 374–386. [Google Scholar] [CrossRef]
- Lima, A.A.M.; Kvalsund, M.P.; de Souza, P.P.E.; Figueiredo, Í.L.; Soares, A.M.; Mota, R.M.S.; Lima, N.L.; Pinkerton, R.C.; Patrick, P.P.; Guerrant, R.L.; et al. Zinc, vitamin A, and glutamine supplementation in Brazilian shantytown children at risk for diarrhea results in sex-specific improvements in verbal learning. Clinics 2013, 68, 351–358. [Google Scholar] [CrossRef]
- Muhammad, F.; Fan, B.; Wang, R.; Ren, J.; Jia, S.; Wang, L.; Chen, Z.; Liu, X.A. The Molecular Gut-Brain Axis in Early Brain Development. Int. J. Mol. Sci. 2022, 23, 15389. [Google Scholar] [CrossRef] [PubMed]
- Patrick, P.D.; Oriá, R.B.; Madhavan, V.; Pinkerton, R.C.; Lorntz, B.; Lima, A.A.M.; Guerrant, R.L. Limitations in verbal fluency following heavy burdens of early childhood diarrhea in Brazilian shantytown children. Child Neuropsychol. 2005, 11, 233–244. [Google Scholar] [CrossRef]
- Niehaus, M.D.; Moore, S.R.; Patrick, P.D.; Derr, L.L.; Lorntz, B.; Lima, A.A.; Guerrant, R.L. Early childhood diarrhea is associated with diminished cognitive function 4 to 7 years later in children in a northeast Brazilian shantytown. Am. J. Trop. Med. Hyg. 2002, 66, 590–593. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerrant, D.I.; Moore, S.R.; Lima, A.A.; Patrick, P.D.; Schorling, J.B.; Guerrant, R.L. Association of early childhood diarrhea and cryptosporidiosis with impaired physical fitness and cognitive function four-seven years later in a poor urban community in northeast Brazil. Am. J. Trop. Med. Hyg. 1999, 61, 707–713. [Google Scholar] [CrossRef]
- Colston, J.M.; Chen, Y.T.; Hinson, P.; Nguyen, N.-L.H.; Peñataro Yori, P.; Olortegui, M.P.; Rengifo Trigoso, D.; Siguas Salas, M.; Guerrant, R.L.; François, R.; et al. Enteropathy Markers in Early Life Were Associated with Adipokine, Apolipoprotein, and Cytokine Profiles Consistent with an Adverse Cardiometabolic Disease Risk Profile Later in Childhood in a Peruvian Birth Cohort. Am. J. Trop. Med. Hyg. 2022, 107, 754–765. [Google Scholar] [CrossRef]
- Guerrant, R.L.; DeBoer, M.D.; Moore, S.R.; Scharf, R.J.; Lima, A.A.M. The impoverished gut--a triple burden of diarrhoea, stunting and chronic disease. Nat. Rev. Gastroenterol. Hepatol. 2013, 10, 220–229. [Google Scholar] [CrossRef]
- DeBoer, M.D.; Lima, A.A.M.; Oría, R.B.; Scharf, R.J.; Moore, S.R.; Luna, M.A.; Guerrant, R.L. Early childhood growth failure and the developmental origins of adult disease: Do enteric infections and malnutrition increase risk for the metabolic syndrome? Nutr. Rev. 2012, 70, 642–653. [Google Scholar] [CrossRef] [Green Version]
- Guerrant, R.L.; Kosek, M.; Moore, S.; Lorntz, B.; Brantley, R.; Lima, A.A.M. Magnitude and impact of diarrheal diseases. Arch. Med. Res. 2002, 33, 351–355. [Google Scholar] [CrossRef] [PubMed]
- Behera, D.K.; Mishra, S. The burden of diarrhea, etiologies, and risk factors in India from 1990 to 2019: Evidence from the global burden of disease study. BMC Public Health 2022, 22, 92. [Google Scholar] [CrossRef] [PubMed]
- Thornburg, K.L.; Shannon, J.; Thuillier, P.; Turker, M.S. In utero life and epigenetic predisposition for disease. Adv. Genet. 2010, 71, 57–78. [Google Scholar] [PubMed] [Green Version]
- Vaiserman, A.; Lushchak, O. Prenatal Malnutrition-Induced Epigenetic Dysregulation as a Risk Factor for Type 2 Diabetes. Int. J. Genom. 2019, 2019, 3821409. [Google Scholar] [CrossRef] [Green Version]
- Takahashi, Y.; Morales Valencia, M.; Yu, Y.; Ouchi, Y.; Takahashi, K.; Shokhirev, M.N.; Lande, K.; Williams, A.E.; Fresia, C.; Kurita, M.; et al. Transgenerational inheritance of acquired epigenetic signatures at CpG islands in mice. Cell 2023, 186, 715–731.e19. [Google Scholar] [CrossRef] [PubMed]
- Alam, M.J.; Islam, M.M.; Maniruzzaman, M.; Ahmed, N.A.M.F.; Tawabunnahar, M.; Rahman, M.J.; Roy, D.C.; Mydam, J. Socioeconomic inequality in the prevalence of low birth weight and its associated determinants in Bangladesh. PLoS ONE 2022, 17, e0276718. [Google Scholar] [CrossRef] [PubMed]
- DeBoer, M.D.; Scharf, R.J.; Leite, A.M.; Férrer, A.; Havt, A.; Pinkerton, R.; Lima, A.A.; Guerrant, R.L. Systemic inflammation, growth factors, and linear growth in the setting of infection and malnutrition. Nutrition 2017, 33, 248–253. [Google Scholar] [CrossRef] [Green Version]
- Leocádio, P.C.L.; Lopes, S.C.; Dias, R.P.; Alvarez-Leite, J.I.; Guerrant, R.L.; Malva, J.O.; Oriá, R.B. The Transition From Undernutrition to Overnutrition Under Adverse Environments and Poverty: The Risk for Chronic Diseases. Front. Nutr. 2021, 8, 676044. [Google Scholar] [CrossRef] [PubMed]
- Upadhyay, R.P.; Naik, G.; Choudhary, T.S.; Chowdhury, R.; Taneja, S.; Bhandari, N.; Martines, J.C.; Bahl, R.; Bhan, M.K. Cognitive and motor outcomes in children born low birth weight: A systematic review and meta-analysis of studies from South Asia. BMC Pediatr. 2019, 19, 35. [Google Scholar] [CrossRef] [PubMed]
- Gomes, C.F.; Sousa, M.; Lourenço, I.; Martins, D.; Torres, J. Gastrointestinal diseases during pregnancy: What does the gastroenterologist need to know? Ann. Gastroenterol. 2018, 31, 385–394. [Google Scholar] [PubMed]
- Gopalakrishna, K.P.; Hand, T.W. Influence of Maternal Milk on the Neonatal Intestinal Microbiome. Nutrients 2020, 12, 823. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iddrisu, I.; Monteagudo-Mera, A.; Poveda, C.; Pyle, S.; Shahzad, M.; Andrews, S.; Walton, G.E. Malnutrition and Gut Microbiota in Children. Nutrients 2021, 13, 2727. [Google Scholar] [CrossRef]
- Blanton, L.V.; Charbonneau, M.R.; Salih, T.; Barratt, M.J.; Venkatesh, S.; Ilkaveya, O.; Subramanian, S.; Manary, M.J.; Trehan, I.; Jorgensen, J.M.; et al. Gut bacteria that prevent growth impairments transmitted by microbiota from malnourished children. Science 2016, 351, 6275. [Google Scholar] [CrossRef] [Green Version]
- Bäckhed, F.; Ding, H.; Wang, T.; Hooper, L.V.; Koh, G.Y.; Nagy, A.; Semenkovich, C.F.; Gordon, J.I. The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. USA 2004, 101, 15718–15723. [Google Scholar] [CrossRef] [Green Version]
- McCormick, B.J.J.; Richard, S.A.; Murray-Kolb, L.E.; Kang, G.; Lima, A.A.M.; Mduma, E.; Kosek, M.N.; Rogawski McQuade, E.T.; Houpt, E.R.; Bessong, P.; et al. Full breastfeeding protection against common enteric bacteria and viruses: Results from the MAL-ED cohort study. Am. J. Clin. Nutr. 2022, 115, 759–769. [Google Scholar] [CrossRef]
- Watanabe, K.; Petri, W.A.J. Environmental Enteropathy: Elusive but Significant Subclinical Abnormalities in Developing Countries. EBioMedicine 2016, 10, 25–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bartelt, L.A.; Bolick, D.T.; Guerrant, R.L. Disentangling Microbial Mediators of Malnutrition: Modeling Environmental Enteric Dysfunction. Cell. Mol. Gastroenterol. Hepatol. 2019, 7, 692–707. [Google Scholar] [CrossRef] [Green Version]
- Syed, S.; Ali, A.; Duggan, C. Environmental Enteric Dysfunction in Children. J. Pediatr. Gastroenterol. Nutr. 2016, 63, 6–14. [Google Scholar] [CrossRef] [Green Version]
- Guerrant, R.L.; Leite, A.M.; Pinkerton, R.; Medeiros, P.H.Q.S.; Cavalcante, P.A.; DeBoer, M.; Kosek, M.; Duggan, C.; Gewirtz, A.; Kagan, J.C.; et al. Biomarkers of Environmental Enteropathy, Inflammation, Stunting, and Impaired Growth in Children in Northeast Brazil. PLoS ONE 2016, 11, e0158772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vaz Nery, S.; Bennett, I.; Clarke, N.E.; Lin, A.; Rahman, Z.; Rahman, M.; Clements, A.C.A. Characterisation of environmental enteropathy biomarkers and associated risk factors in children in the context of a WASH trial in Timor-Leste. Int. J. Hyg. Environ. Health 2018, 221, 901–906. [Google Scholar] [CrossRef] [PubMed]
- Jones, K.D.; Thitiri, J.; Ngari, M.; Berkley, J.A. Childhood malnutrition: Toward an understanding of infections, inflammation, and antimicrobials. Food Nutr. Bull. 2014, 35, S64–S70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grosso, G.; Mateo, A.; Rangelov, N.; Buzeti, T.; Birt, C. Nutrition in the context of the Sustainable Development Goals. Eur. J. Public Health 2020, 30, i19–i23. [Google Scholar] [CrossRef]
- Hodges, P.; Tembo, M.; Kelly, P. Intestinal Biopsies for the Evaluation of Environmental Enteropathy and Environmental Enteric Dysfunction. J. Infect. Dis. 2021, 224, S856–S863. [Google Scholar] [CrossRef]
- Subramanian, S.; Huq, S.; Yatsunenko, T.; Haque, R.; Mahfuz, M.; Alam, M.A.; Benezra, A.; DeStefano, J.; Meier, M.F.; Muegge, B.D.; et al. Persistent gut microbiota immaturity in malnourished Bangladeshi children. Nature 2014, 510, 417–421. [Google Scholar] [CrossRef] [Green Version]
- Marungruang, N.; Kovalenko, T.; Osadchenko, I.; Voss, U.; Huang, F.; Burleigh, S.; Ushakova, G.; Skibo, G.; Nyman, M.; Prykhodko, O.; et al. Lingonberries and their two separated fractions differently alter the gut microbiota, improve metabolic functions, reduce gut inflammatory properties, and improve brain function in ApoE−/− mice fed high-fat diet. Nutr. Neurosci. 2020, 23, 600–612. [Google Scholar] [CrossRef] [Green Version]
- Phuah, C.-L.; Dave, T.; Malik, R.; Raffeld, M.R.; Ayres, A.M.; Goldstein, J.N.; Viswanathan, A.; Greenberg, S.M.; Jagiella, J.M.; Hansen, B.M.; et al. Genetic variants influencing elevated myeloperoxidase levels increase risk of stroke. Brain 2017, 140, 2663–2672. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, T.; Rogers, K.A.; Rutt, B.K.; Ronald, J.A. Close Association of Myeloperoxidase-Producing Activated Microglia with Amyloid Plaques in Hypercholesterolemic Rabbits. J. Alzheimer’s Dis. 2019, 67, 1221–1234. [Google Scholar] [CrossRef]
- Nasrin, D.; Blackwelder, W.C.; Sommerfelt, H.; Wu, Y.; Farag, T.H.; Panchalingam, S.; Biswas, K.; Saha, D.; Jahangir Hossain, M.; Sow, S.O.; et al. Pathogens Associated With Linear Growth Faltering in Children With Diarrhea and Impact of Antibiotic Treatment: The Global Enteric Multicenter Study. J. Infect. Dis. 2021, 224, S848–S855. [Google Scholar] [CrossRef]
- Pane, S.; Putignani, L. Cryptosporidium: Still Open Scenarios. Pathogens 2022, 11, 515. [Google Scholar] [CrossRef] [PubMed]
- Bolick, D.T.; Mayneris-Perxachs, J.; Medlock, G.L.; Kolling, G.L.; Papin, J.A.; Swann, J.R.; Guerrant, R.L. Increased Urinary Trimethylamine N-Oxide Following Cryptosporidium Infection and Protein Malnutrition Independent of Microbiome Effects. J. Infect. Dis. 2017, 216, 64–71. [Google Scholar] [CrossRef]
- Lima, A.A.M.; Oriá, R.B.; Soares, A.M.; Filho, J.Q.; de Sousa, F.J.; Abreu, C.B.; Bindá, A.; Lima, I.; Quetz, J.; Moraes, M.; et al. Geography, population, demography, socioeconomic, anthropometry, and environmental status in the MAL-ED cohort and case-control study Sites in Fortaleza, Ceará, Brazil. Clin. Infect. Dis. 2014, 59 (Suppl. 4), S287–S294. [Google Scholar] [CrossRef] [Green Version]
- Bushen, O.Y.; Kohli, A.; Pinkerton, R.C.; Dupnik, K.; Newman, R.D.; Sears, C.L.; Fayer, R.; Lima, A.A.M.; Guerrant, R.L. Heavy cryptosporidial infections in children in northeast Brazil: Comparison of Cryptosporidium hominis and Cryptosporidium parvum. Trans. R. Soc. Trop. Med. Hyg. 2007, 101, 378–384. [Google Scholar] [CrossRef]
- Kirkpatrick, B.D.; Daniels, M.M.; Jean, S.S.; Pape, J.W.; Karp, C.; Littenberg, B.; Fitzgerald, D.W.; Lederman, H.M.; Nataro, J.P.; Sears, C.L. Cryptosporidiosis stimulates an inflammatory intestinal response in malnourished Haitian children. J. Infect. Dis. 2002, 186, 94–101. [Google Scholar] [CrossRef] [Green Version]
- de Medeiros, P.H.Q.S.; Pinto, D.V.; de Almeida, J.Z.; Rêgo, J.M.C.; Rodrigues, F.A.P.; Lima, A.Â.M.; Bolick, D.T.; Guerrant, R.L.; Oriá, R.B. Modulation of Intestinal Immune and Barrier Functions by Vitamin A: Implications for Current Understanding of Malnutrition and Enteric Infections in Children. Nutrients 2018, 10, 1128. [Google Scholar] [CrossRef] [Green Version]
- Pinkerton, R.; Oriá, R.B.; Lima, A.A.M.; Rogawski, E.T.; Oriá, M.O.B.; Patrick, P.D.; Moore, S.R.; Wiseman, B.L.; Niehaus, M.D.; Guerrant, R.L. Early Childhood Diarrhea Predicts Cognitive Delays in Later Childhood Independently of Malnutrition. Am. J. Trop. Med. Hyg. 2016, 95, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Murray-Kolb, L.E.; Rasmussen, Z.A.; Scharf, R.J.; Rasheed, M.A.; Svensen, E.; Seidman, J.C.; Tofail, F.; Koshy, B.; Shrestha, R.; Maphula, A.; et al. The MAL-ED cohort study: Methods and lessons learned when assessing early child development and caregiving mediators in infants and young children in 8 low- and middle-income countries. Clin. Infect. Dis. 2014, 59 (Suppl. 4), S261–S272. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- MAL-ED Network Investigators The MAL-ED study: A multinational and multidisciplinary approach to understand the relationship between enteric pathogens, malnutrition, gut physiology, physical growth, cognitive development, and immune responses in infants and children up to 2 years of. Clin. Infect. Dis. 2014, 59 (Suppl. 4), S193–S206.
- Platts-Mills, J.A.; Babji, S.; Bodhidatta, L.; Gratz, J.; Haque, R.; Havt, A.; McCormick, B.J.; McGrath, M.; Olortegui, M.P.; Samie, A.; et al. Pathogen-specific burdens of community diarrhoea in developing countries: A multisite birth cohort study (MAL-ED). Lancet Glob. Health 2015, 3, e564–e575. [Google Scholar] [CrossRef] [Green Version]
- Kosek, M.; Haque, R.; Lima, A.; Babji, S.; Shrestha, S.; Qureshi, S.; Amidou, S.; Mduma, E.; Lee, G.; Yori, P.P.; et al. Fecal markers of intestinal inflammation and permeability associated with the subsequent acquisition of linear growth deficits in infants. Am. J. Trop. Med. Hyg. 2013, 88, 390–396. [Google Scholar] [CrossRef] [Green Version]
- Marques, O.; Weiss, G.; Muckenthaler, M.U. The role of iron in chronic inflammatory diseases: From mechanisms to treatment options in anemia of inflammation. Blood 2022, 140, 2011–2023. [Google Scholar] [CrossRef]
- Lauer, J.M.; Ghosh, S.; Ausman, L.M.; Webb, P.; Bashaasha, B.; Agaba, E.; Turyashemererwa, F.M.; Tran, H.Q.; Gewirtz, A.T.; Erhardt, J.; et al. Markers of Environmental Enteric Dysfunction Are Associated with Poor Growth and Iron Status in Rural Ugandan Infants. J. Nutr. 2020, 150, 2175–2182. [Google Scholar] [CrossRef]
- Loukas, A.; Hotez, P.J.; Diemert, D.; Yazdanbakhsh, M.; McCarthy, J.S.; Correa-Oliveira, R.; Croese, J.; Bethony, J.M. Hookworm infection. Nat. Rev. Dis. Prim. 2016, 2, 16088. [Google Scholar] [CrossRef] [PubMed]
- Opoka, R.O.; Conroy, A.L.; Waiswa, A.; Wasswa, R.; Tumwine, J.K.; Karamagi, C.; John, C.C. Severe Anemia Is Associated with Systemic Inflammation in Young Children Presenting to a Tertiary Hospital in Uganda. Am. J. Trop. Med. Hyg. 2020, 103, 2574–2580. [Google Scholar] [CrossRef]
- Sangkhae, V.; Nemeth, E. Regulation of the Iron Homeostatic Hormone Hepcidin. Adv. Nutr. Int. Rev. J. 2017, 8, 126–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lam, L.K.M.; Murphy, S.; Kokkinaki, D.; Venosa, A.; Sherrill-Mix, S.; Casu, C.; Rivella, S.; Weiner, A.; Park, J.; Shin, S.; et al. DNA binding to TLR9 expressed by red blood cells promotes innate immune activation and anemia. Sci. Transl. Med. 2021, 13, eabj1008. [Google Scholar] [CrossRef] [PubMed]
- Nampijja, M.; Mutua, A.M.; Elliott, A.M.; Muriuki, J.M.; Abubakar, A.; Webb, E.L.; Atkinson, S.H. Low Hemoglobin Levels Are Associated with Reduced Psychomotor and Language Abilities in Young Ugandan Children. Nutrients 2022, 14, 1452. [Google Scholar] [CrossRef] [PubMed]
- van Tilborg, E.; de Theije, C.G.M.; van Hal, M.; Wagenaar, N.; de Vries, L.S.; Benders, M.J.; Rowitch, D.H.; Nijboer, C.H. Origin and dynamics of oligodendrocytes in the developing brain: Implications for perinatal white matter injury. Glia 2018, 66, 221–238. [Google Scholar] [CrossRef]
- Graf, A.E.; Lallier, S.W.; Waidyaratne, G.; Thompson, M.D.; Tipple, T.E.; Hester, M.E.; Trask, A.J.; Rogers, L.K. Maternal high fat diet exposure is associated with increased hepcidin levels, decreased myelination, and neurobehavioral changes in male offspring. Brain Behav. Immun. 2016, 58, 369–378. [Google Scholar] [CrossRef] [Green Version]
- Nannan, N.; Laubscher, R.; Nel, J.H.; Neethling, I.; Dhansay, M.A.; Turawa, E.B.; Labadarios, D.; Pacella, R.; Bradshaw, D.; Pillay van-Wyk, V. Estimating the changing burden of disease attributable to childhood stunting, wasting and underweight in South Africa for 2000, 2006 and 2012. S. Afr. Med. J. 2022, 112, 676–683. [Google Scholar] [CrossRef] [PubMed]
- Gona, P.N.; Gona, C.M.; Chikwasha, V.; Haruzivishe, C.; Rao, S.R.; Mapoma, C.C. Oral rehydration solution coverage in under 5 children with diarrhea: A tri-country, subnational, cross-sectional comparative analysis of two demographic health surveys cycles. BMC Public Health 2020, 20, 1716. [Google Scholar] [CrossRef] [PubMed]
- Avelar Rodriguez, D.; Ryan, P.M.; Toro Monjaraz, E.M.; Ramirez Mayans, J.A.; Quigley, E.M. Small Intestinal Bacterial Overgrowth in Children: A State-Of-The-Art Review. Front. Pediatr. 2019, 7, 363. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Donowitz, J.R.; Pu, Z.; Lin, Y.; Alam, M.; Ferdous, T.; Shama, T.; Taniuchi, M.; Islam, M.O.; Kabir, M.; Nayak, U.; et al. Small Intestine Bacterial Overgrowth in Bangladeshi Infants Is Associated With Growth Stunting in a Longitudinal Cohort. Am. J. Gastroenterol. 2022, 117, 167–175. [Google Scholar] [CrossRef]
- Rogawski, E.T.; Guerrant, R.L. The Burden of Enteropathy and “Subclinical” Infections. Pediatr. Clin. N. Am. 2017, 64, 815–836. [Google Scholar] [CrossRef]
- Hennessey, C.; Keogh, C.E.; Barboza, M.; Brust-Mascher, I.; Knotts, T.A.; Sladek, J.A.; Pusceddu, M.M.; Stokes, P.; Rabasa, G.; Honeycutt, M.; et al. Neonatal Enteropathogenic Escherichia coli Infection Disrupts Microbiota-Gut-Brain Axis Signaling. Infect. Immun. 2021, 89, e0005921. [Google Scholar] [CrossRef]
- McCormick, B.J.J.; Caulfield, L.E.; Richard, S.A.; Pendergast, L.; Seidman, J.C.; Maphula, A.; Koshy, B.; Blacy, L.; Roshan, R.; Nahar, B.; et al. Early Life Experiences and Trajectories of Cognitive Development. Pediatrics 2020, 146, e20193660. [Google Scholar] [CrossRef]
- Platts-Mills, J.A.; Liu, J.; Rogawski, E.T.; Kabir, F.; Lertsethtakarn, P.; Siguas, M.; Khan, S.S.; Praharaj, I.; Murei, A.; Nshama, R.; et al. Use of quantitative molecular diagnostic methods to assess the aetiology, burden, and clinical characteristics of diarrhoea in children in low-resource settings: A reanalysis of the MAL-ED cohort study. Lancet Glob. Health 2018, 6, e1309–e1318. [Google Scholar] [CrossRef] [Green Version]
- Schwabkey, Z.I.; Wiesnoski, D.H.; Chang, C.-C.; Tsai, W.-B.; Pham, D.; Ahmed, S.S.; Hayase, T.; Ortega Turrubiates, M.R.; El-Himri, R.K.; Sanchez, C.A.; et al. Diet-derived metabolites and mucus link the gut microbiome to fever after cytotoxic cancer treatment. Sci. Transl. Med. 2022, 14, eabo3445. [Google Scholar] [CrossRef]
- Laval, L.; Martin, R.; Natividad, J.N.; Chain, F.; Miquel, S.; Desclée de Maredsous, C.; Capronnier, S.; Sokol, H.; Verdu, E.F.; van Hylckama Vlieg, J.E.T.; et al. Lactobacillus rhamnosus CNCM I-3690 and the commensal bacterium Faecalibacterium prausnitzii A2-165 exhibit similar protective effects to induced barrier hyper-permeability in mice. Gut Microbes 2015, 6, 1–9. [Google Scholar] [CrossRef] [Green Version]
- Braniste, V.; Al-Asmakh, M.; Kowal, C.; Anuar, F.; Abbaspour, A.; Tóth, M.; Korecka, A.; Bakocevic, N.; Ng, L.G.; Kundu, P.; et al. The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 2014, 6, 263ra158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, S.; Yi, J.; Zhang, Y.G.; Zhou, J.; Sun, J. Leaky intestine and impaired microbiome in an amyotrophic lateral sclerosis mouse model. Physiol. Rep. 2015, 3, e12356. [Google Scholar] [CrossRef] [Green Version]
- Parker, A.; Fonseca, S.; Carding, S.R. Gut microbes and metabolites as modulators of blood-brain barrier integrity and brain health. Gut Microbes 2020, 11, 135–157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jin, J.; Xu, Z.; Zhang, L.; Zhang, C.; Zhao, X.; Mao, Y.; Zhang, H.; Liang, X.; Wu, J.; Yang, Y.; et al. Gut-derived β-amyloid: Likely a centerpiece of the gut-brain axis contributing to Alzheimer’s pathogenesis. Gut Microbes 2023, 15, 2167172. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Y.; Jaber, V.R.; Pogue, A.I.; Sharfman, N.M.; Taylor, C.; Lukiw, W.J. Lipopolysaccharides (LPSs) as Potent Neurotoxic Glycolipids in Alzheimer’s Disease (AD). Int. J. Mol. Sci. 2022, 23, 12671. [Google Scholar] [CrossRef] [PubMed]
- Kesika, P.; Suganthy, N.; Sivamaruthi, B.S.; Chaiyasut, C. Role of gut-brain axis, gut microbial composition, and probiotic intervention in Alzheimer’s disease. Life Sci. 2021, 264, 118627. [Google Scholar] [CrossRef]
- Alexandrov, P.; Zhai, Y.; Li, W.; Lukiw, W. Lipopolysaccharide-stimulated, NF-kB-, miRNA-146a- and miRNA-155-mediated molecular-genetic communication between the human gastrointestinal tract microbiome and the brain. Folia Neuropathol. 2019, 57, 211–219. [Google Scholar] [CrossRef]
- Kalyan, M.; Tousif, A.H.; Sonali, S.; Vichitra, C.; Sunanda, T.; Praveenraj, S.S.; Ray, B.; Gorantla, V.R.; Rungratanawanich, W.; Mahalakshmi, A.M.; et al. Role of Endogenous Lipopolysaccharides in Neurological Disorders. Cells 2022, 11, 4038. [Google Scholar] [CrossRef]
- Pistollato, F.; Sumalla Cano, S.; Elio, I.; Masias Vergara, M.; Giampieri, F.; Battino, M. Role of gut microbiota and nutrients in amyloid formation and pathogenesis of Alzheimer disease. Nutr. Rev. 2016, 74, 624–634. [Google Scholar] [CrossRef] [Green Version]
- Wiesinger, A.; Peters, W.; Chappell, D.; Kentrup, D.; Reuter, S.; Pavenstädt, H.; Oberleithner, H.; Kümpers, P. Nanomechanics of the endothelial glycocalyx in experimental sepsis. PLoS ONE 2013, 8, e80905. [Google Scholar] [CrossRef] [Green Version]
- Mou, Y.; Du, Y.; Zhou, L.; Yue, J.; Hu, X.; Liu, Y.; Chen, S.; Lin, X.; Zhang, G.; Xiao, H.; et al. Gut Microbiota Interact With the Brain Through Systemic Chronic Inflammation: Implications on Neuroinflammation, Neurodegeneration, and Aging. Front. Immunol. 2022, 13, 796288. [Google Scholar] [CrossRef] [PubMed]
- Haruwaka, K.; Ikegami, A.; Tachibana, Y.; Ohno, N.; Konishi, H.; Hashimoto, A.; Matsumoto, M.; Kato, D.; Ono, R.; Kiyama, H.; et al. Dual microglia effects on blood brain barrier permeability induced by systemic inflammation. Nat. Commun. 2019, 10, 5816. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Husain, M.A.; Laurent, B.; Plourde, M. APOE and Alzheimer’s Disease: From Lipid Transport to Physiopathology and Therapeutics. Front. Neurosci. 2021, 15, 630502. [Google Scholar] [CrossRef] [PubMed]
- Kelly, M.E.; Clay, M.A.; Mistry, M.J.; Hsieh-Li, H.M.; Harmony, J.A. Apolipoprotein E inhibition of proliferation of mitogen-activated T lymphocytes: Production of interleukin 2 with reduced biological activity. Cell. Immunol. 1994, 159, 124–139. [Google Scholar] [CrossRef]
- Baitsch, D.; Bock, H.H.; Engel, T.; Telgmann, R.; Müller-Tidow, C.; Varga, G.; Bot, M.; Herz, J.; Robenek, H.; von Eckardstein, A.; et al. Apolipoprotein E induces antiinflammatory phenotype in macrophages. Arter. Thromb. Vasc. Biol. 2011, 31, 1160–1168. [Google Scholar] [CrossRef] [Green Version]
- Mahley, R.W.; Rall, S.C.J. Apolipoprotein E: Far more than a lipid transport protein. Annu. Rev. Genom. Hum. Genet. 2000, 1, 507–537. [Google Scholar] [CrossRef]
- Tudorache, I.F.; Trusca, V.G.; Gafencu, A.V. Apolipoprotein E-A Multifunctional Protein with Implications in Various Pathologies as a Result of Its Structural Features. Comput. Struct. Biotechnol. J. 2017, 15, 359–365. [Google Scholar] [CrossRef]
- Azevedo, O.G.; Bolick, D.T.; Roche, J.K.; Pinkerton, R.F.; Lima, A.A.; Vitek, M.P.; Warren, C.A.; Oriá, R.B.; Guerrant, R.L. Apolipoprotein E plays a key role against cryptosporidial infection in transgenic undernourished mice. PLoS ONE 2014, 9, e89562. [Google Scholar]
- Das, S.; Stevens, T.; Castillo, C.; Villasenõr, A.; Arredondo, H.; Reddy, K. Lipid metabolism in mucous-dwelling amitochondriate protozoa. Int. J. Parasitol. 2002, 32, 655–675. [Google Scholar] [CrossRef]
- Oriá, R.B.; Patrick, P.D.; Blackman, J.A.; Lima, A.A.M.; Guerrant, R.L. Role of apolipoprotein E4 in protecting children against early childhood diarrhea outcomes and implications for later development. Med. Hypotheses 2007, 68, 1099–1107. [Google Scholar] [CrossRef] [Green Version]
- Nankar, S.A.; Ahmed, S.; Sharma, S.S.; Pande, A.H. Apolipoprotein-mimetic Peptides: Current and Future Prospectives. Curr. Protein Pept. Sci. 2022, 23, 757–772. [Google Scholar] [PubMed]
- Chen, Y.; Strickland, M.R.; Soranno, A.; Holtzman, D.M. Apolipoprotein E: Structural Insights and Links to Alzheimer Disease Pathogenesis. Neuron 2021, 109, 205–221. [Google Scholar] [CrossRef] [PubMed]
- Mahley, R.W. Apolipoprotein E: From cardiovascular disease to neurodegenerative disorders. J. Mol. Med. 2016, 94, 739–746. [Google Scholar] [CrossRef] [Green Version]
- Laskowitz, D.T.; Thekdi, A.D.; Thekdi, S.D.; Han, S.K.; Myers, J.K.; Pizzo, S.V.; Bennett, E.R. Downregulation of microglial activation by apolipoprotein E and apoE-mimetic peptides. Exp. Neurol. 2001, 167, 74–85. [Google Scholar] [CrossRef] [PubMed]
- Lynch, J.R.; Tang, W.; Wang, H.; Vitek, M.P.; Bennett, E.R.; Sullivan, P.M.; Warner, D.S.; Laskowitz, D.T. APOE genotype and an ApoE-mimetic peptide modify the systemic and central nervous system inflammatory response. J. Biol. Chem. 2003, 278, 48529–48533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Azevedo, O.G.R.; Oliveira, R.A.C.; Oliveira, B.C.; Zaja-Milatovic, S.; Araújo, C.V.; Wong, D.V.T.; Costa, T.B.; Lucena, H.B.M.; Lima, R.C.P.J.; Ribeiro, R.A.; et al. Apolipoprotein E COG 133 mimetic peptide improves 5-fluorouracil-induced intestinal mucositis. BMC Gastroenterol. 2012, 12, 35. [Google Scholar] [CrossRef] [Green Version]
- Jofre-Monseny, L.; Minihane, A.-M.; Rimbach, G. Impact of apoE genotype on oxidative stress, inflammation and disease risk. Mol. Nutr. Food Res. 2008, 52, 131–145. [Google Scholar] [CrossRef]
- Chuang, K.; Elford, E.L.; Tseng, J.; Leung, B.; Harris, H.W. An expanding role for apolipoprotein E in sepsis and inflammation. Am. J. Surg. 2010, 200, 391–397. [Google Scholar] [CrossRef] [Green Version]
- de Bont, N.; Netea, M.G.; Demacker, P.N.; Verschueren, I.; Kullberg, B.J.; van Dijk, K.W.; van der Meer, J.W.; Stalenhoef, A.F. Apolipoprotein E knock-out mice are highly susceptible to endotoxemia and Klebsiella pneumoniae infection. J. Lipid Res. 1999, 40, 680–685. [Google Scholar] [CrossRef]
- Singh, K.; Chaturvedi, R.; Asim, M.; Barry, D.P.; Lewis, N.D.; Vitek, M.P.; Wilson, K.T. The apolipoprotein E-mimetic peptide COG112 inhibits the inflammatory response to Citrobacter rodentium in colonic epithelial cells by preventing NF-kappaB activation. J. Biol. Chem. 2008, 283, 16752–16761. [Google Scholar] [CrossRef] [Green Version]
- Singh, K.; Chaturvedi, R.; Barry, D.P.; Coburn, L.A.; Asim, M.; Lewis, N.D.; Piazuelo, M.B.; Washington, M.K.; Vitek, M.P.; Wilson, K.T. The apolipoprotein E-mimetic peptide COG112 inhibits NF-kappaB signaling, proinflammatory cytokine expression, and disease activity in murine models of colitis. J. Biol. Chem. 2011, 286, 3839–3850. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, J.; Zheng, M.; Liang, P.; Wei, Y.; Yin, X.; Tang, Y.; Xue, Y. Apolipoprotein E and its mimetic peptide suppress Th1 and Th17 responses in experimental autoimmune encephalomyelitis. Neurobiol. Dis. 2013, 56, 59–65. [Google Scholar] [CrossRef]
- Petruk, G.; Elvén, M.; Hartman, E.; Davoudi, M.; Schmidtchen, A.; Puthia, M.; Petrlova, J. The role of full-length apoE in clearance of Gram-negative bacteria and their endotoxins. J. Lipid Res. 2021, 62, 100086. [Google Scholar] [CrossRef]
- Puthia, M.; Marzinek, J.K.; Petruk, G.; Bergdahl, G.E.; Bond, P.J.; Petrlova, J. Antibacterial and Anti-Inflammatory Effects of Apolipoprotein E. Biomedicines 2022, 10, 1430. [Google Scholar] [CrossRef]
- Shao, Y.; Zhao, T.; Zhang, W.; He, J.; Lu, F.; Cai, Y.; Lai, Z.; Wei, N.; Liang, C.; Liu, L.; et al. Presence of the apolipoprotein E-ε4 allele is associated with an increased risk of sepsis progression. Sci. Rep. 2020, 10, 15735. [Google Scholar] [CrossRef] [PubMed]
- Freitas, R.S.; Roque, C.R.; Matos, G.A.; Belayev, L.; de Azevedo, O.G.R.; Alvarez-Leite, J.I.; Guerrant, R.L.; Oriá, R.B. Immunoinflammatory role of apolipoprotein E4 in malnutrition and enteric infections and the increased risk for chronic diseases under adverse environments. Nutr. Rev. 2022, 80, 1001–1012. [Google Scholar] [CrossRef] [PubMed]
- Petropoulou, P.-I.; Berbée, J.F.P.; Theodoropoulos, V.; Hatziri, A.; Stamou, P.; Karavia, E.A.; Spyridonidis, A.; Karagiannides, I.; Kypreos, K.E. Lack of LCAT reduces the LPS-neutralizing capacity of HDL and enhances LPS-induced inflammation in mice. Biochim. Biophys. Acta (BBA)-Mol. Basis Dis. 2015, 1852, 2106–2115. [Google Scholar] [CrossRef] [Green Version]
- Whitson, H.E.; Colton, C.; El Khoury, J.; Gate, D.; Goate, A.; Heneka, M.T.; Kaddurah-Daouk, R.; Klein, R.S.; Shinohara, M.L.; Sisodia, S.; et al. Infection and inflammation: New perspectives on Alzheimer’s disease. Brain Behav. Immun.-Health 2022, 22, 100462. [Google Scholar] [CrossRef]
- Zhang, P.; Wang, T.; Zhu, X.; Feng, L.; Wang, J.; Li, Y.; Zhang, X.; Cui, T.; Li, M. Jiedu Yizhi Formula Improves Cognitive Function by Regulating the Gut Dysbiosis and TLR4/NF-κB Signaling Pathway. Neuropsychiatr. Dis. Treat 2023, 19, 49–62. [Google Scholar] [CrossRef]
- Zhu, Z.; Ma, X.; Wu, J.; Xiao, Z.; Wu, W.; Ding, S.; Zheng, L.; Liang, X.; Luo, J.; Ding, D.; et al. Altered Gut Microbiota and Its Clinical Relevance in Mild Cognitive Impairment and Alzheimer’s Disease: Shanghai Aging Study and Shanghai Memory Study. Nutrients 2022, 14, 3959. [Google Scholar] [CrossRef]
- Pires, M.; Rego, A.C. Apoe4 and Alzheimer’s Disease Pathogenesis-Mitochondrial Deregulation and Targeted Therapeutic Strategies. Int. J. Mol. Sci. 2023, 24, 778. [Google Scholar] [CrossRef] [PubMed]
- Ungar, L.; Altmann, A.; Greicius, M.D. Apolipoprotein E, gender, and Alzheimer’s disease: An overlooked, but potent and promising interaction. Brain Imaging Behav. 2014, 8, 262–273. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tran, T.T.T.; Corsini, S.; Kellingray, L.; Hegarty, C.; Le Gall, G.; Narbad, A.; Müller, M.; Tejera, N.; O’Toole, P.W.; Minihane, A.M.; et al. APOE genotype influences the gut microbiome structure and function in humans and mice: Relevance for Alzheimer’s disease pathophysiology. FASEB J. 2019, 33, 8221–8231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vitek, M.P.; Brown, C.M.; Colton, C.A. APOE genotype-specific differences in the innate immune response. Neurobiol. Aging 2009, 30, 1350–1360. [Google Scholar] [CrossRef] [Green Version]
- Raulin, A.-C.; Doss, S.V.; Trottier, Z.A.; Ikezu, T.C.; Bu, G.; Liu, C.-C. ApoE in Alzheimer’s disease: Pathophysiology and therapeutic strategies. Mol. Neurodegener. 2022, 17, 72. [Google Scholar] [CrossRef]
- Seo, D.-O.; O’Donnell, D.; Jain, N.; Ulrich, J.D.; Herz, J.; Li, Y.; Lemieux, M.; Cheng, J.; Hu, H.; Serrano, J.R.; et al. ApoE isoform- and microbiota-dependent progression of neurodegeneration in a mouse model of tauopathy. Science 2023, 379, eadd1236. [Google Scholar] [CrossRef]
- Gale, S.C.; Gao, L.; Mikacenic, C.; Coyle, S.M.; Rafaels, N.; Murray Dudenkov, T.; Madenspacher, J.H.; Draper, D.W.; Ge, W.; Aloor, J.J.; et al. APOε4 is associated with enhanced in vivo innate immune responses in human subjects. J. Allergy Clin. Immunol. 2014, 134, 127–134. [Google Scholar] [CrossRef] [Green Version]
- Oriá, R.B.; Patrick, P.D.; Zhang, H.; Lorntz, B.; de Castro Costa, C.M.; Brito, G.A.C.; Barrett, L.J.; Lima, A.A.M.; Guerrant, R.L. APOE4 protects the cognitive development in children with heavy diarrhea burdens in Northeast Brazil. Pediatr. Res. 2005, 57, 310–316. [Google Scholar] [CrossRef] [Green Version]
- Vasunilashorn, S.; Finch, C.E.; Crimmins, E.M.; Vikman, S.A.; Stieglitz, J.; Gurven, M.; Kaplan, H.; Allayee, H. Inflammatory gene variants in the Tsimane, an indigenous Bolivian population with a high infectious load. Biodemography Soc. Biol. 2011, 57, 33–52. [Google Scholar] [CrossRef] [Green Version]
- Garcia, A.R.; Finch, C.; Gatz, M.; Kraft, T.; Rodriguez, D.E.; Cummings, D.; Charifson, M.; Buetow, K.; Beheim, B.A.; Allayee, H.; et al. APOE4 is associated with elevated blood lipids and lower levels of innate immune biomarkers in a tropical Amerindian subsistence population. eLife 2021, 10, e68231. [Google Scholar] [CrossRef]
- Trumble, B.C.; Stieglitz, J.; Blackwell, A.D.; Allayee, H.; Beheim, B.; Finch, C.E.; Gurven, M.; Kaplan, H. Apolipoprotein E4 is associated with improved cognitive function in Amazonian forager-horticulturalists with a high parasite burden. FASEB J. 2017, 31, 1508–1515. [Google Scholar] [CrossRef] [Green Version]
- Karlsson, E.K.; Kwiatkowski, D.P.; Sabeti, P.C. Natural selection and infectious disease in human populations. Nat. Rev. Genet. 2014, 15, 379–393. [Google Scholar] [CrossRef] [PubMed]
- Corbett, S.; Courtiol, A.; Lummaa, V.; Moorad, J.; Stearns, S. The transition to modernity and chronic disease: Mismatch and natural selection. Nat. Rev. Genet. 2018, 19, 419–430. [Google Scholar] [CrossRef] [PubMed]
- Weksler, M.E.; Relkin, N.; Turkenich, R.; LaRusse, S.; Zhou, L.; Szabo, P. Patients with Alzheimer disease have lower levels of serum anti-amyloid peptide antibodies than healthy elderly individuals. Exp. Gerontol. 2002, 37, 943–948. [Google Scholar] [CrossRef]
- Kowalski, K.; Mulak, A. Brain-Gut-Microbiota Axis in Alzheimer’s Disease. J. Neurogastroenterol. Motil. 2019, 25, 48–60. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, A.; Matsushita, M.; Zhang, L.; Wang, H.; Shi, X.; Gu, H.; Xia, Z.; Cui, J.Y. Cadmium exposure modulates the gut-liver axis in an Alzheimer’s disease mouse model. Commun. Biol. 2021, 4, 1398. [Google Scholar] [CrossRef]
- Mitter, S.S.; Oriá, R.B.; Kvalsund, M.P.; Pamplona, P.; Joventino, E.S.; Mota, R.M.S.; Gonçalves, D.C.; Patrick, P.D.; Guerrant, R.L.; Lima, A.A.M. Apolipoprotein E4 influences growth and cognitive responses to micronutrient supplementation in shantytown children from northeast Brazil. Clinics 2012, 67, 11–18. [Google Scholar] [CrossRef]
- Pessoa, T.; Carneiro, I.; Bertolinni, L.; Oria, R.; Abreu Junior Jose, G. Modulation of the wnt/β-catenin signal pathway in vitro by apoE COG1410 in IEC-18 after 5-FU injury. Ann. Oncol. 2017, 28, iii128. [Google Scholar] [CrossRef]
- Pessoa, T.; Carneiro, I.; Aguiar, P.; Vieira, L.; Vitek, M.; Guerrant, R.; Oria, R.; Abreu Junior, J. WNT/β-catenin pathway in recovering IEC-6 intestinal cells after 5-FU induced injury and treatment with mimetic peptide APOE COG133. Ann. Oncol. 2019, 30, iv102. [Google Scholar] [CrossRef]
- Mauch, D.H.; Nägler, K.; Schumacher, S.; Göritz, C.; Müller, E.C.; Otto, A.; Pfrieger, F.W. CNS synaptogenesis promoted by glia-derived cholesterol. Science 2001, 294, 1354–1357. [Google Scholar] [CrossRef]
- Yu, T.-S.; Tensaouti, Y.; Stephanz, E.P.; Chintamen, S.; Rafikian, E.E.; Yang, M.; Kernie, S.G. Astrocytic ApoE underlies maturation of hippocampal neurons and cognitive recovery after traumatic brain injury in mice. Commun. Biol. 2021, 4, 1303. [Google Scholar] [CrossRef]
- Hirschfeld, L.R.; Risacher, S.L.; Nho, K.; Saykin, A.J. Myelin repair in Alzheimer’s disease: A review of biological pathways and potential therapeutics. Transl. Neurodegener. 2022, 11, 47. [Google Scholar] [CrossRef]
- Ahmed, S.; Pande, A.H.; Sharma, S.S. Therapeutic potential of ApoE-mimetic peptides in CNS disorders: Current perspective. Exp. Neurol. 2022, 353, 114051. [Google Scholar] [CrossRef]
- Cristóvão, A.C.; Bernardino, L.; Ferreira, R. Advances in Brain Vascular Research; Cambridge Scholars Publisher: Newcastle upon Tyne, UK, 2020. [Google Scholar]
- Zlokovic, B.V. Neurovascular pathways to neurodegeneration in Alzheimer’s disease and other disorders. Nat. Rev. Neurosci. 2011, 12, 723–738. [Google Scholar] [CrossRef] [PubMed]
- Verghese, P.B.; Castellano, J.M.; Holtzman, D.M. Apolipoprotein E in Alzheimer’s disease and other neurological disorders. Lancet Neurol. 2011, 10, 241–252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bell, R.D.; Winkler, E.A.; Singh, I.; Sagare, A.P.; Deane, R.; Wu, Z.; Holtzman, D.M.; Betsholtz, C.; Armulik, A.; Sallstrom, J.; et al. Apolipoprotein E controls cerebrovascular integrity via cyclophilin A. Nature 2012, 485, 512–516. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hays, C.C.; Zlatar, Z.Z.; Meloy, M.J.; Bondi, M.W.; Gilbert, P.E.; Liu, T.T.; Helm, J.L.; Wierenga, C.E. APOE modifies the interaction of entorhinal cerebral blood flow and cortical thickness on memory function in cognitively normal older adults. Neuroimage 2019, 202, 116162. [Google Scholar] [CrossRef] [PubMed]
- Bangen, K.J.; Beiser, A.; Delano-Wood, L.; Nation, D.A.; Lamar, M.; Libon, D.J.; Bondi, M.W.; Seshadri, S.; Wolf, P.A.; Au, R. APOE genotype modifies the relationship between midlife vascular risk factors and later cognitive decline. J. Stroke Cerebrovasc. Dis. 2013, 22, 1361–1369. [Google Scholar] [CrossRef] [Green Version]
- Lin, A.-L.; Jahrling, J.B.; Zhang, W.; DeRosa, N.; Bakshi, V.; Romero, P.; Galvan, V.; Richardson, A. Rapamycin rescues vascular, metabolic and learning deficits in apolipoprotein E4 transgenic mice with pre-symptomatic Alzheimer’s disease. J. Cerebrovasc. Blood Flow Metab. 2017, 37, 217–226. [Google Scholar] [CrossRef] [Green Version]
- Alata, W.; Ye, Y.; St-Amour, I.; Vandal, M.; Calon, F. Human apolipoprotein E ɛ4 expression impairs cerebral vascularization and blood-brain barrier function in mice. J. Cerebrovasc. Blood Flow Metab. 2015, 35, 86–94. [Google Scholar] [CrossRef]
- Jackson, R.J.; Meltzer, J.C.; Nguyen, H.; Commins, C.; Bennett, R.E.; Hudry, E.; Hyman, B.T. APOE4 derived from astrocytes leads to blood-brain barrier impairment. Brain 2022, 145, 3582–3593. [Google Scholar] [CrossRef]
- Lynch, J.R.; Pineda, J.A.; Morgan, D.; Zhang, L.; Warner, D.S.; Benveniste, H.; Laskowitz, D.T. Apolipoprotein E affects the central nervous system response to injury and the development of cerebral edema. Ann. Neurol. 2002, 51, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Pang, J.; Peng, J.; Cao, F.; Vitek, M.P.; Li, F.; Jiang, Y.; Sun, X. An apoE-derived mimic peptide, COG1410, alleviates early brain injury via reducing apoptosis and neuroinflammation in a mouse model of subarachnoid hemorrhage. Neurosci. Lett. 2016, 627, 92–99. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; He, Q.; Wang, M.; Wang, X.; Gong, F.; Bai, L.; Zhang, J.; Wang, W. Quantifying blood-brain-barrier leakage using a combination of evans blue and high molecular weight FITC-Dextran. J. Neurosci. Methods 2019, 325, 108349. [Google Scholar] [CrossRef] [PubMed]
- Pang, J.; Chen, Y.; Kuai, L.; Yang, P.; Peng, J.; Wu, Y.; Chen, Y.; Vitek, M.P.; Chen, L.; Sun, X.; et al. Inhibition of Blood-Brain Barrier Disruption by an Apolipoprotein E-Mimetic Peptide Ameliorates Early Brain Injury in Experimental Subarachnoid Hemorrhage. Transl. Stroke Res. 2017, 8, 257–272. [Google Scholar] [CrossRef]
- Qin, X.; You, H.; Cao, F.; Wu, Y.; Peng, J.; Pang, J.; Xu, H.; Chen, Y.; Chen, L.; Vitek, M.P.; et al. Apolipoprotein E Mimetic Peptide Increases Cerebral Glucose Uptake by Reducing Blood-Brain Barrier Disruption after Controlled Cortical Impact in Mice: An (18)F-Fluorodeoxyglucose PET/CT Study. J. Neurotrauma 2017, 34, 943–951. [Google Scholar] [CrossRef] [Green Version]
- Cao, F.; Yin, A.; Wen, G.; Sheikh, A.M.; Tauqeer, Z.; Malik, M.; Nagori, A.; Schirripa, M.; Schirripa, F.; Merz, G.; et al. Retraction Note: Alteration of astrocytes and Wnt/β-catenin signaling in the frontal cortex of autistic subjects. J. Neuroinflamm. 2016, 13, 106. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Peng, J.; Pang, J.; Wu, Y.; Huang, X.; Li, Y.; Zhou, J.; Gu, L.; Sun, X.; Chen, L.; et al. Apolipoprotein E-Mimetic Peptide COG1410 Promotes Autophagy by Phosphorylating GSK-3β in Early Brain Injury Following Experimental Subarachnoid Hemorrhage. Front. Neurosci. 2018, 12, 127. [Google Scholar] [CrossRef] [Green Version]
- Navab, M.; Shechter, I.; Anantharamaiah, G.M.; Reddy, S.T.; Van Lenten, B.J.; Fogelman, A.M. Structure and function of HDL mimetics. Arter. Thromb. Vasc. Biol. 2010, 30, 164–168. [Google Scholar] [CrossRef] [Green Version]
- James, M.L.; Komisarow, J.M.; Wang, H.; Laskowitz, D.T. Therapeutic Development of Apolipoprotein E Mimetics for Acute Brain Injury: Augmenting Endogenous Responses to Reduce Secondary Injury. Neurotherapeutics 2020, 17, 475–483. [Google Scholar] [CrossRef]
- Guttman, M.; Prieto, J.H.; Handel, T.M.; Domaille, P.J.; Komives, E.A. Structure of the minimal interface between ApoE and LRP. J. Mol. Biol. 2010, 398, 306–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, R.; Hong, J.; Lu, M.; Neil, J.E.; Vitek, M.P.; Liu, X.; Warner, D.S.; Li, F.; Sheng, H. ApoE mimetic ameliorates motor deficit and tissue damage in rat spinal cord injury. J. Neurosci. Res. 2014, 92, 884–892. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Gao, B.; Ouyang, J.; Tai, B.; Zhou, S. COG133 Attenuates the Early Brain Injury Induced by Blood-Brain Barrier Disruption in Experimental Subarachnoid Hemorrhage. J. Health Eng. 2022, 2022, 4404039. [Google Scholar] [CrossRef]
- Wang, Q.; Kumar, V.; Lin, F.; Sethi, B.; Coulter, D.W.; McGuire, T.R.; Mahato, R.I. ApoE mimetic peptide targeted nanoparticles carrying a BRD4 inhibitor for treating Medulloblastoma in mice. J. Control. Release 2020, 323, 463–474. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, G.; Kolls, B.J.; Tan, Y.; Fang, C.; Wang, H.; Laskowitz, D.T. Apolipoprotein E mimetic peptide CN-105 improves outcome in a murine model of SAH. Stroke Vasc. Neurol. 2018, 3, 222–230. [Google Scholar] [CrossRef] [Green Version]
- Christensen, D.J.; Ohkubo, N.; Oddo, J.; Van Kanegan, M.J.; Neil, J.; Li, F.; Colton, C.A.; Vitek, M.P. Apolipoprotein E and peptide mimetics modulate inflammation by binding the SET protein and activating protein phosphatase 2A. J. Immunol. 2011, 186, 2535–2542. [Google Scholar] [CrossRef] [Green Version]
- Zhao, Y.; Li, J.; Tang, Q.; Gao, J.; Chen, C.; Jing, L.; Zhang, P.; Li, S. Apolipoprotein E mimetic peptide protects against diffuse brain injury. Neural Regen. Res. 2014, 9, 463–473. [Google Scholar] [CrossRef]
- Bergmann, S.; Lawler, S.E.; Qu, Y.; Fadzen, C.M.; Wolfe, J.M.; Regan, M.S.; Pentelute, B.L.; Agar, N.Y.R.; Cho, C.-F. Blood-brain-barrier organoids for investigating the permeability of CNS therapeutics. Nat. Protoc. 2018, 13, 2827–2843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oriá, R.B.; Malva, J.O.; Foley, P.L.; Freitas, R.S.; Bolick, D.T.; Guerrant, R.L. Revisiting Inbred Mouse Models to Study the Developing Brain: The Potential Role of Intestinal Microbiota. Front. Hum. Neurosci. 2018, 12, 358. [Google Scholar] [CrossRef] [Green Version]
- Wang, B.; Zhang, F.-W.; Wang, W.-X.; Zhao, Y.-Y.; Sun, S.-Y.; Yu, J.-H.; Vitek, M.P.; Li, G.F.; Ma, R.; Wang, S.; et al. Apolipoprotein E mimetic peptide COG1410 combats pandrug-resistant Acinetobacter baumannii. Front. Microbiol. 2022, 13, 934765. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oriá, R.B.; Freitas, R.S.; Roque, C.R.; Nascimento, J.C.R.; Silva, A.P.; Malva, J.O.; Guerrant, R.L.; Vitek, M.P. ApoE Mimetic Peptides to Improve the Vicious Cycle of Malnutrition and Enteric Infections by Targeting the Intestinal and Blood-Brain Barriers. Pharmaceutics 2023, 15, 1086. https://doi.org/10.3390/pharmaceutics15041086
Oriá RB, Freitas RS, Roque CR, Nascimento JCR, Silva AP, Malva JO, Guerrant RL, Vitek MP. ApoE Mimetic Peptides to Improve the Vicious Cycle of Malnutrition and Enteric Infections by Targeting the Intestinal and Blood-Brain Barriers. Pharmaceutics. 2023; 15(4):1086. https://doi.org/10.3390/pharmaceutics15041086
Chicago/Turabian StyleOriá, Reinaldo B., Raul S. Freitas, Cássia R. Roque, José Carlos R. Nascimento, Ana Paula Silva, João O. Malva, Richard L. Guerrant, and Michael P. Vitek. 2023. "ApoE Mimetic Peptides to Improve the Vicious Cycle of Malnutrition and Enteric Infections by Targeting the Intestinal and Blood-Brain Barriers" Pharmaceutics 15, no. 4: 1086. https://doi.org/10.3390/pharmaceutics15041086