
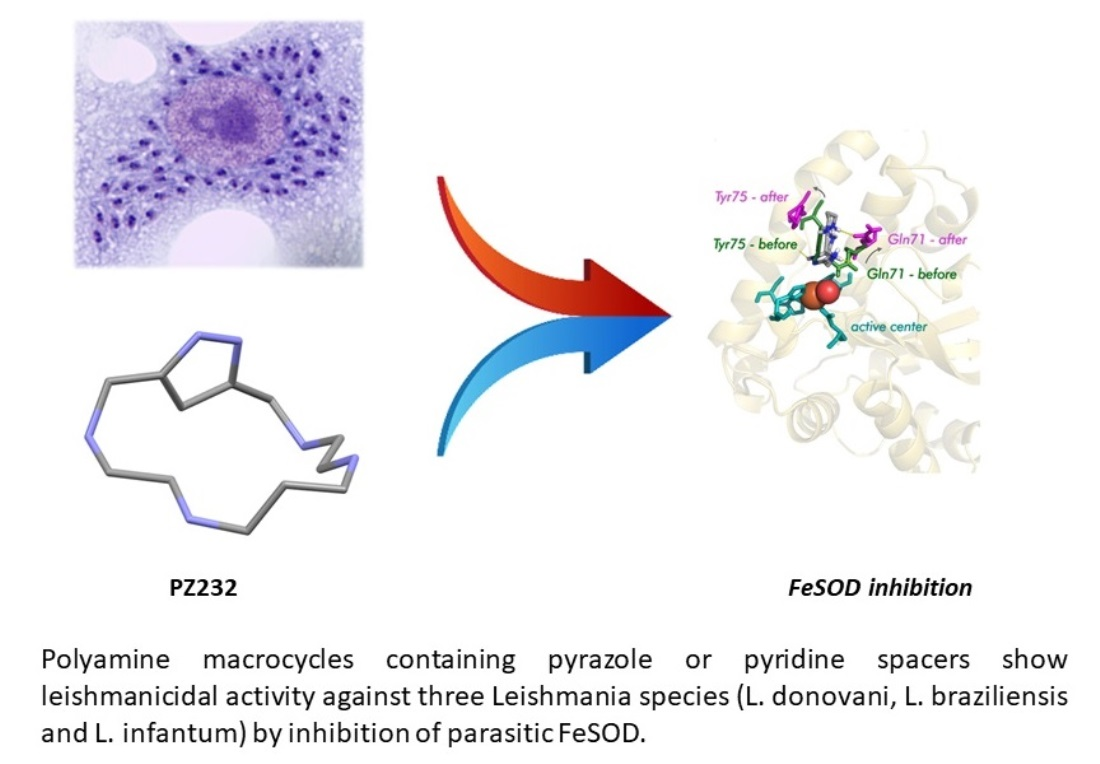
The Bioactivity of Xylene, Pyridine, and Pyrazole Aza Macrocycles against Three Representative Leishmania Species †
 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemistry
2.2. Parasite Strain and Culture
2.3. Cell Culture and Cytotoxicity Studies
2.4. In Vitro Activity Assays
2.5. Infectivity Assays
2.6. SOD Inhibition Assay
2.7. Metabolite Excretion
2.8. Rhodamine Assay
2.9. Theoretical Calculations
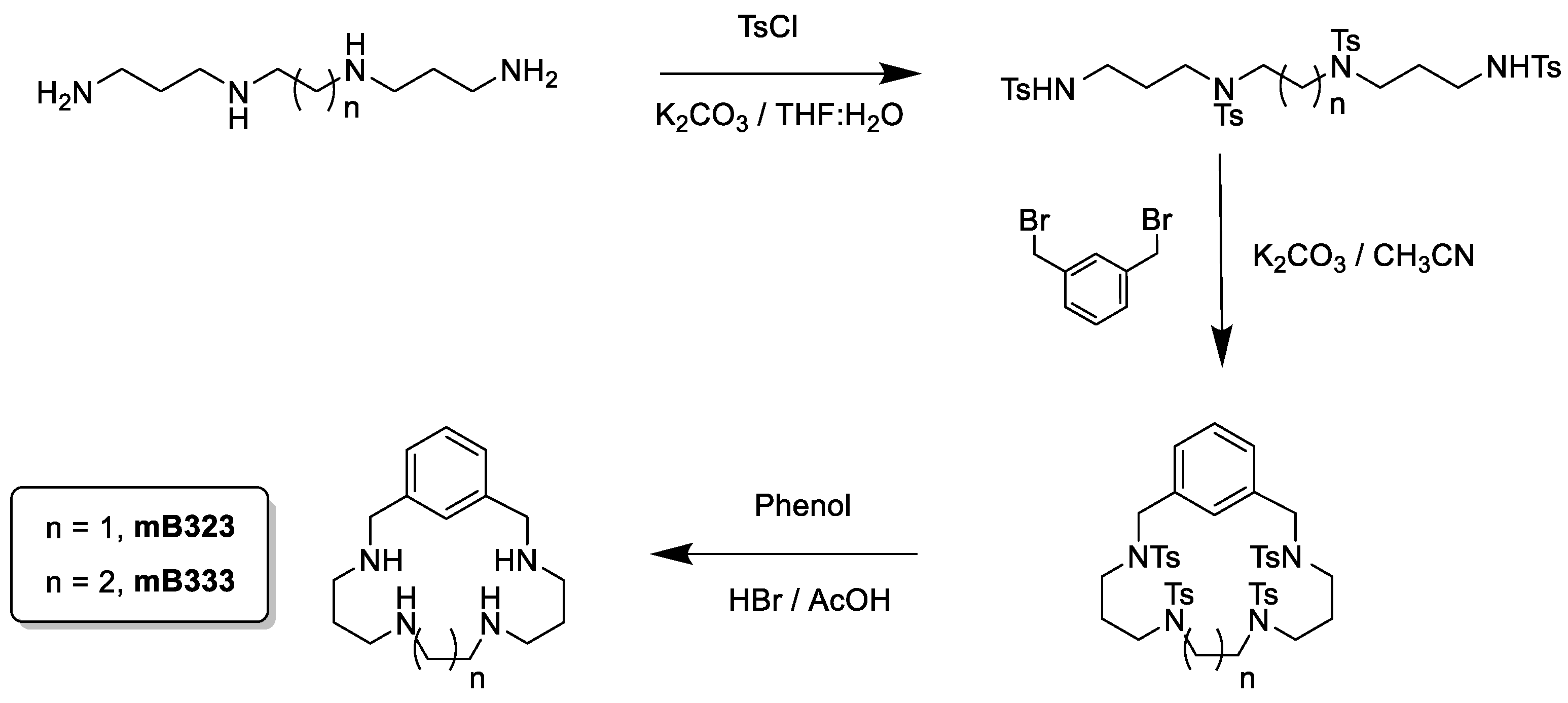
2.10. Synthesis of mB323 and mB333
3. Results and Discussions
3.1. Synthesis of the Compounds
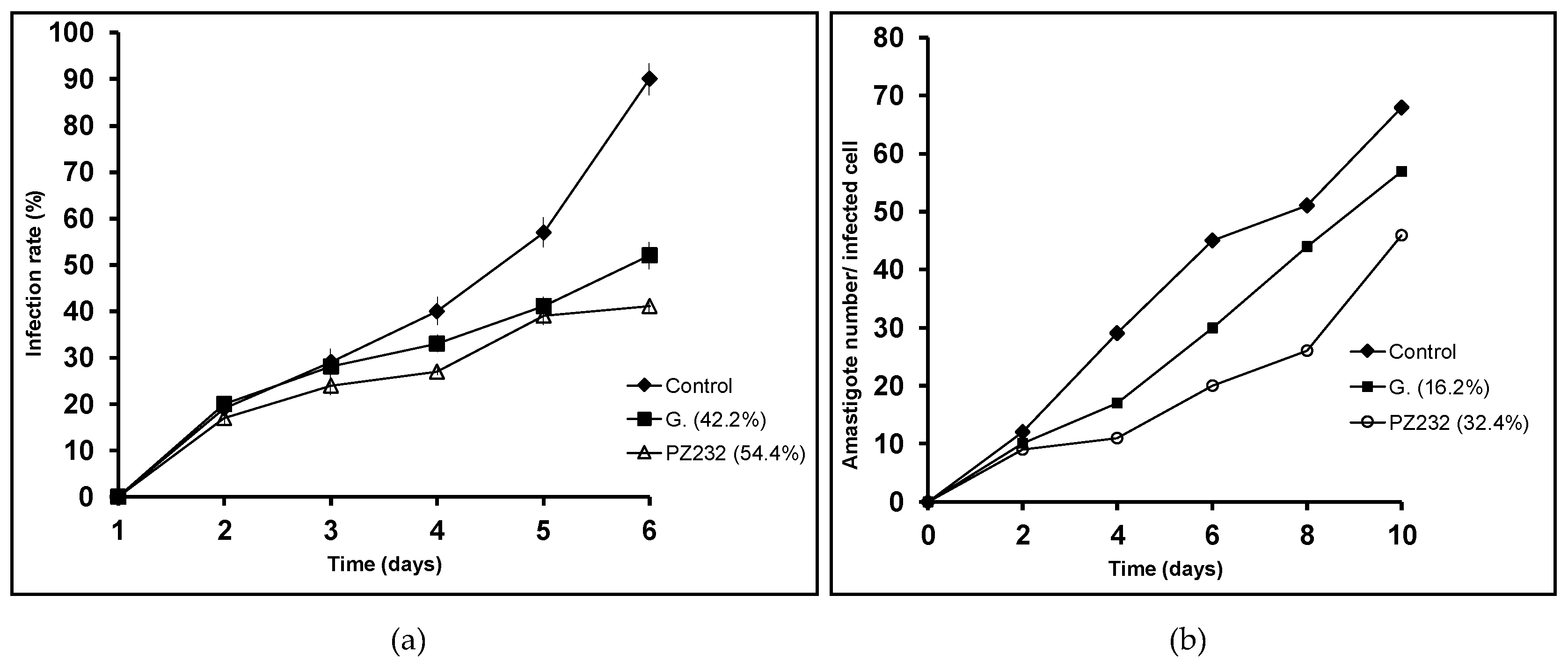
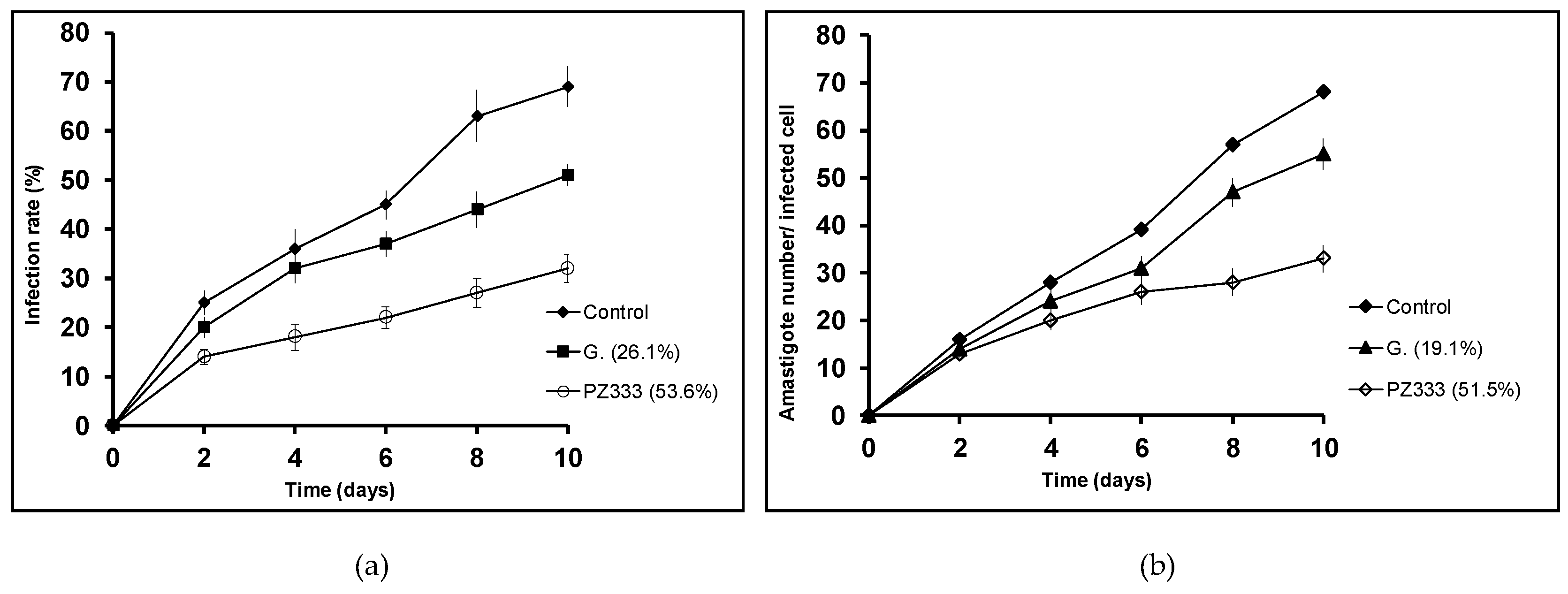
3.2. In Vitro Activity and Infectivity Assays
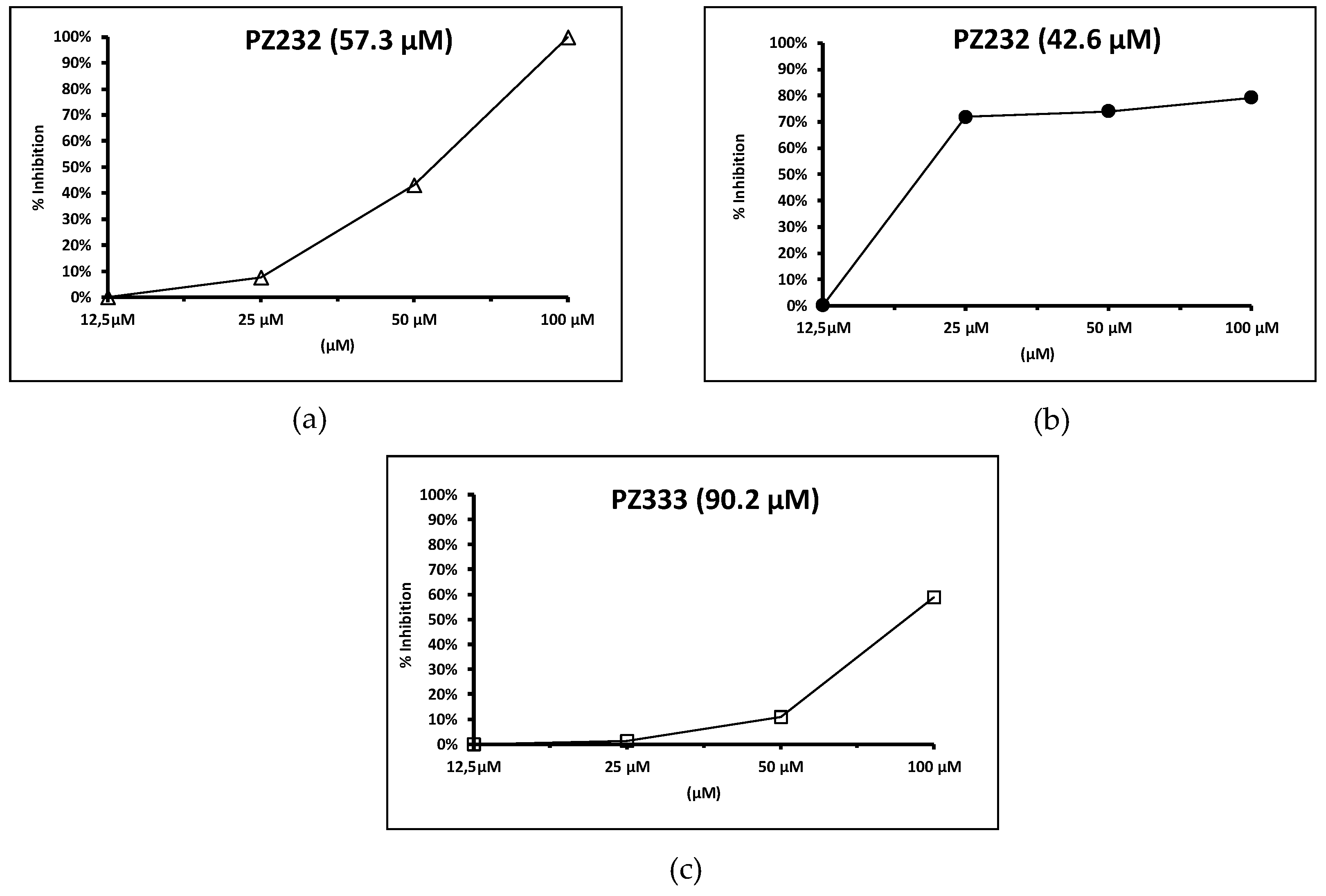
3.3. SOD Inhibition Assays
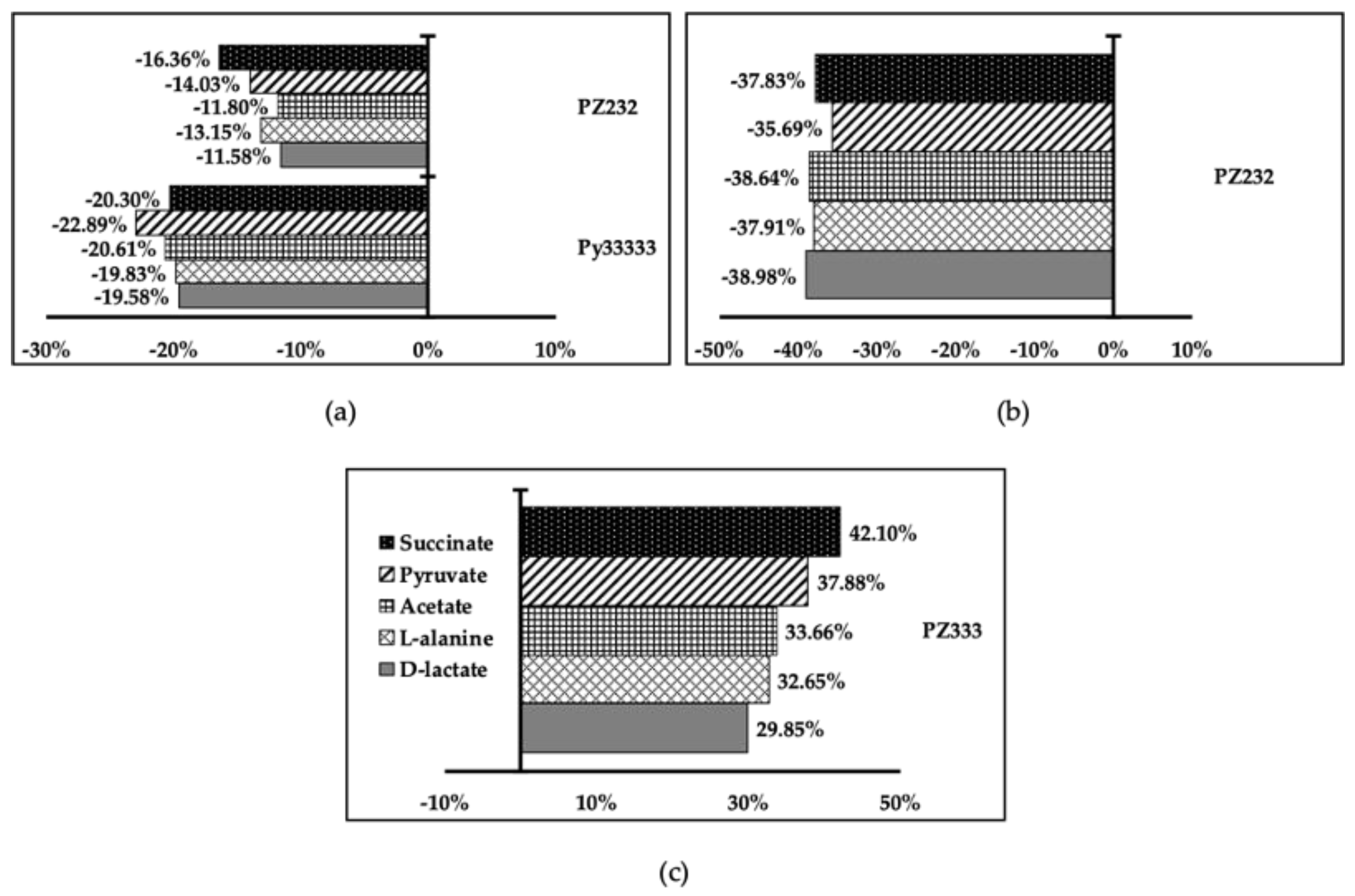
3.4. Metabolite Excretion Assays
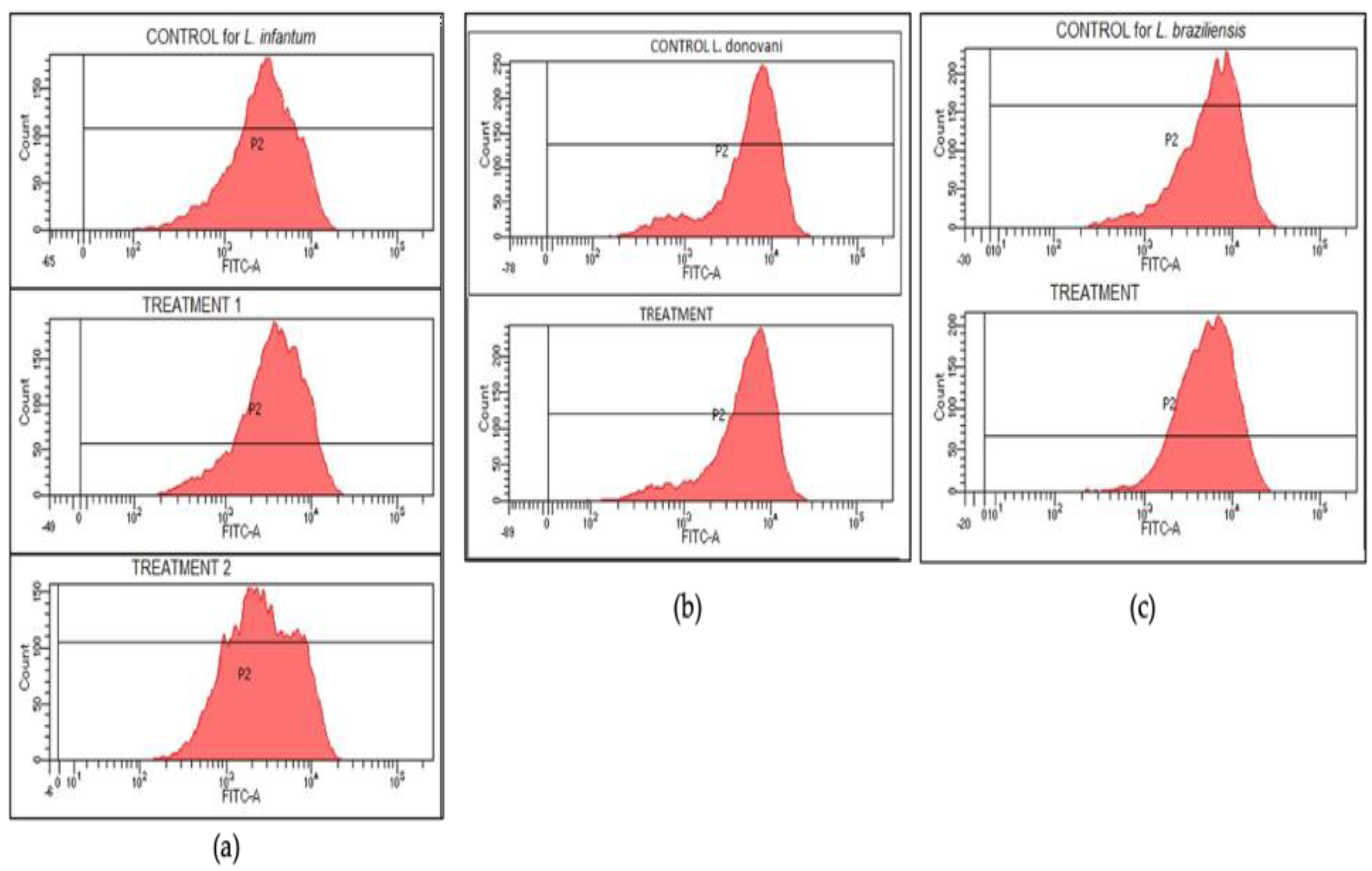
3.5. Rhodamine Assay
3.6. Theoretical Calculations
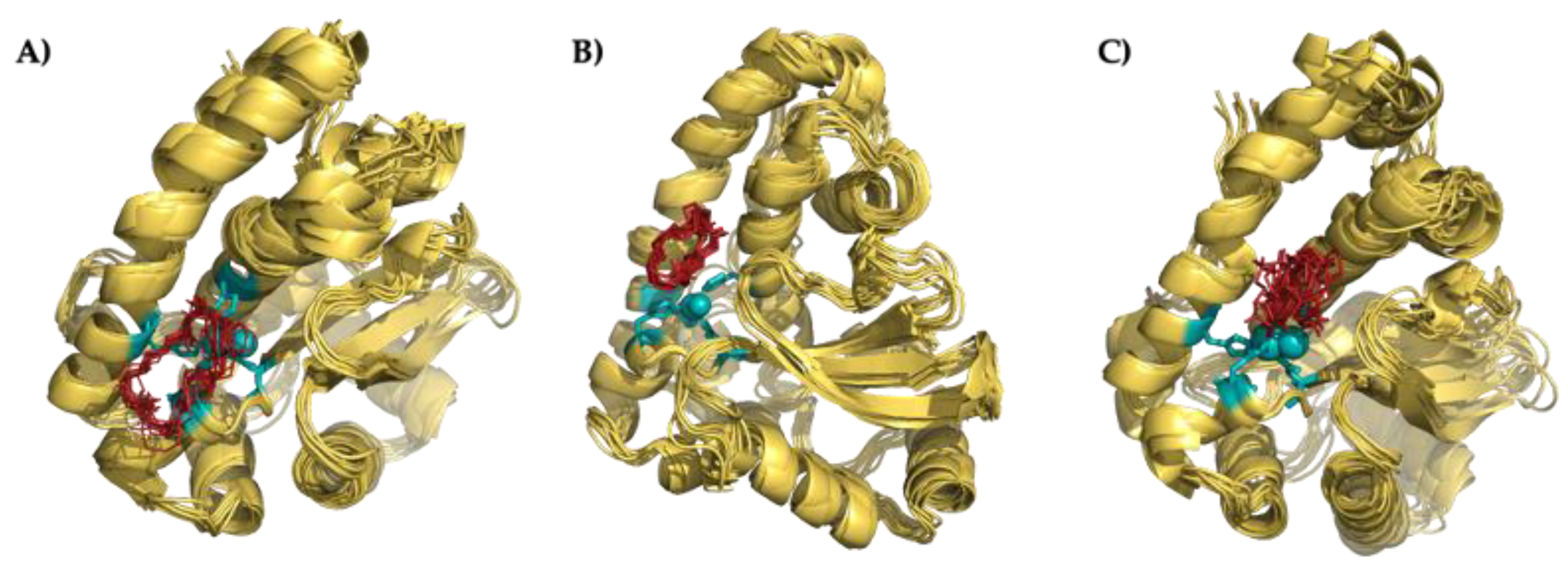
3.6.1. Preliminary Description of the Active Center of the Fe-SOD Enzyme
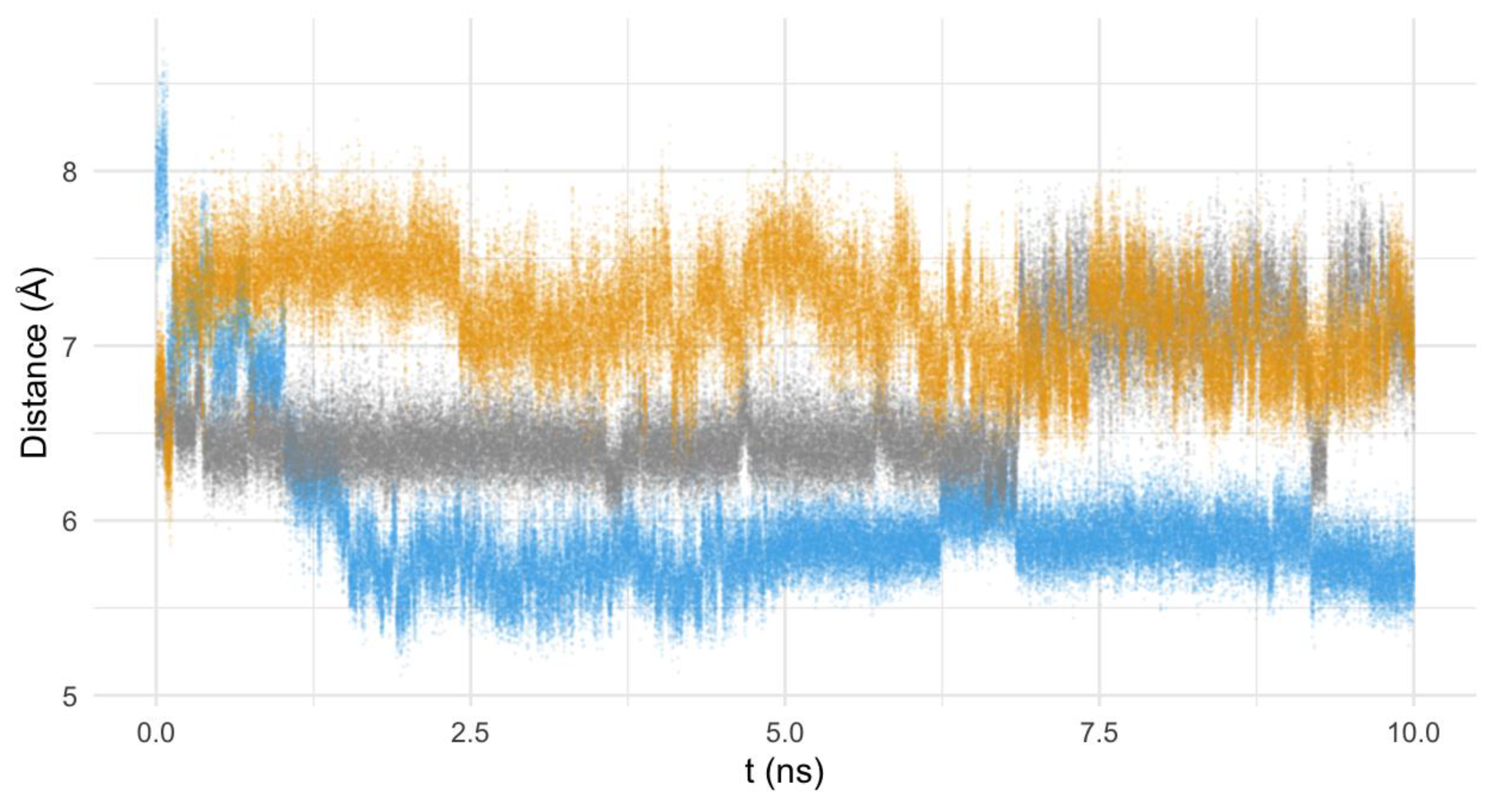
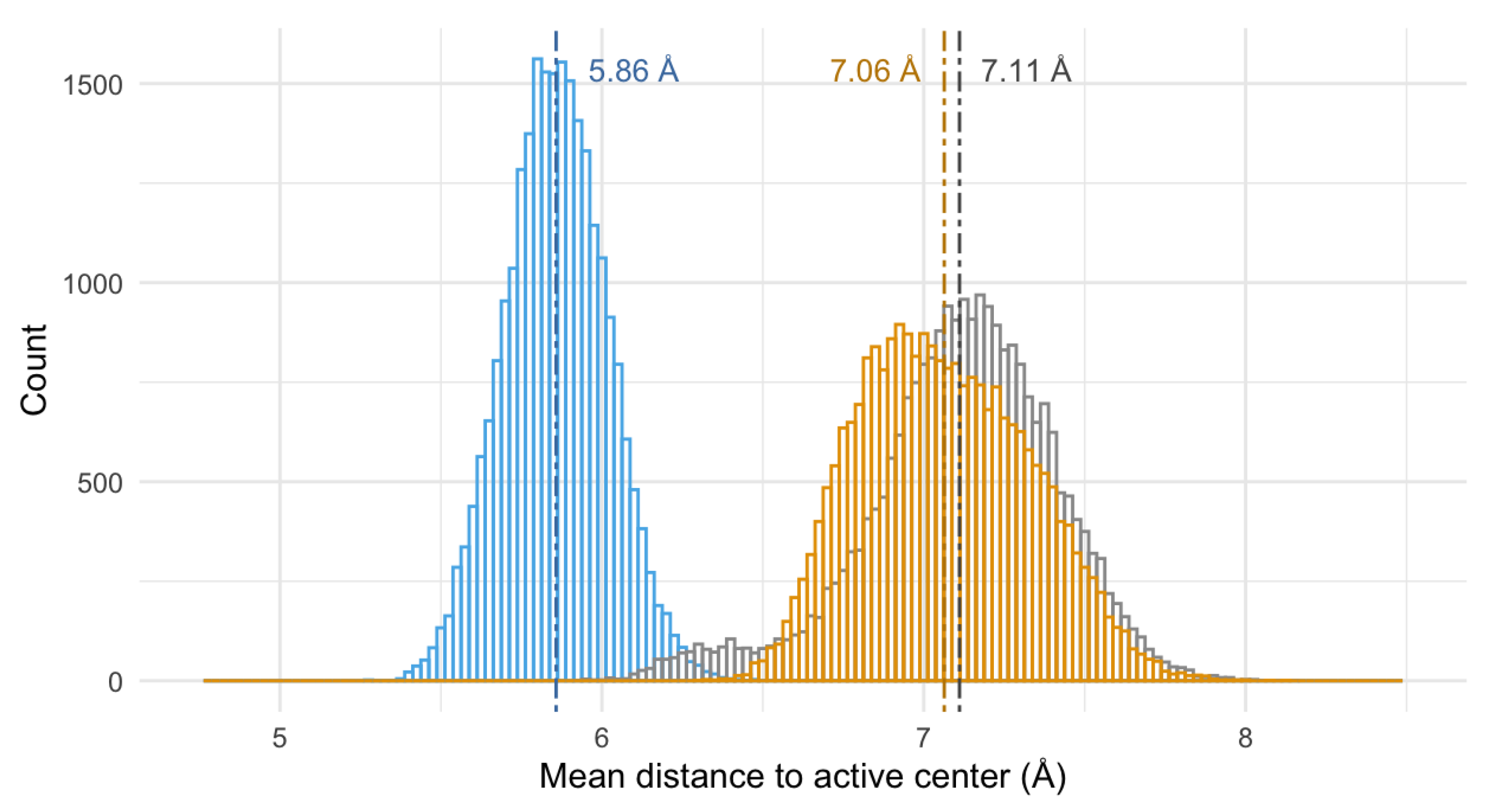
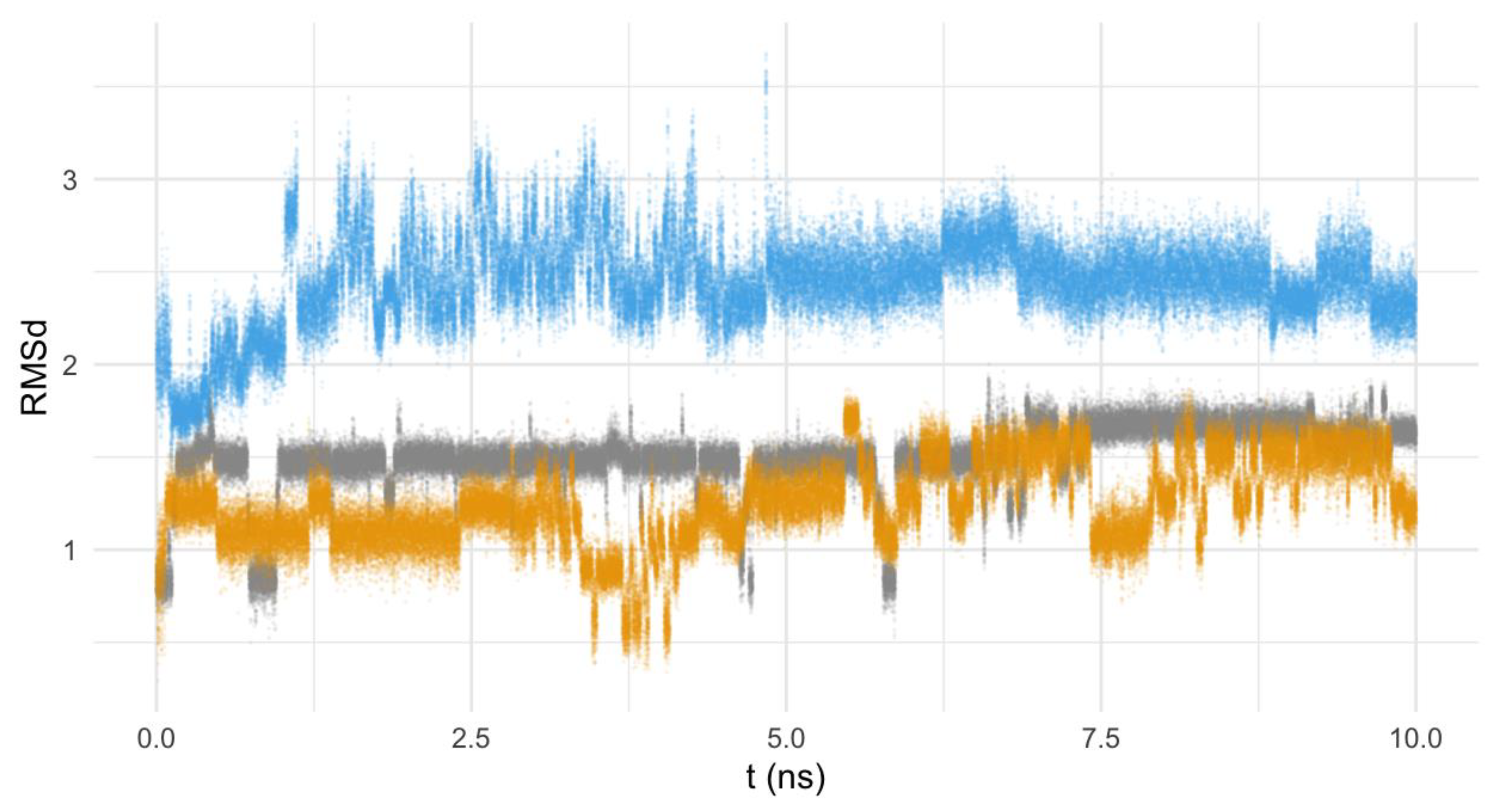
3.6.2. Nature of the Interaction Macrocycle: Fe-SOD
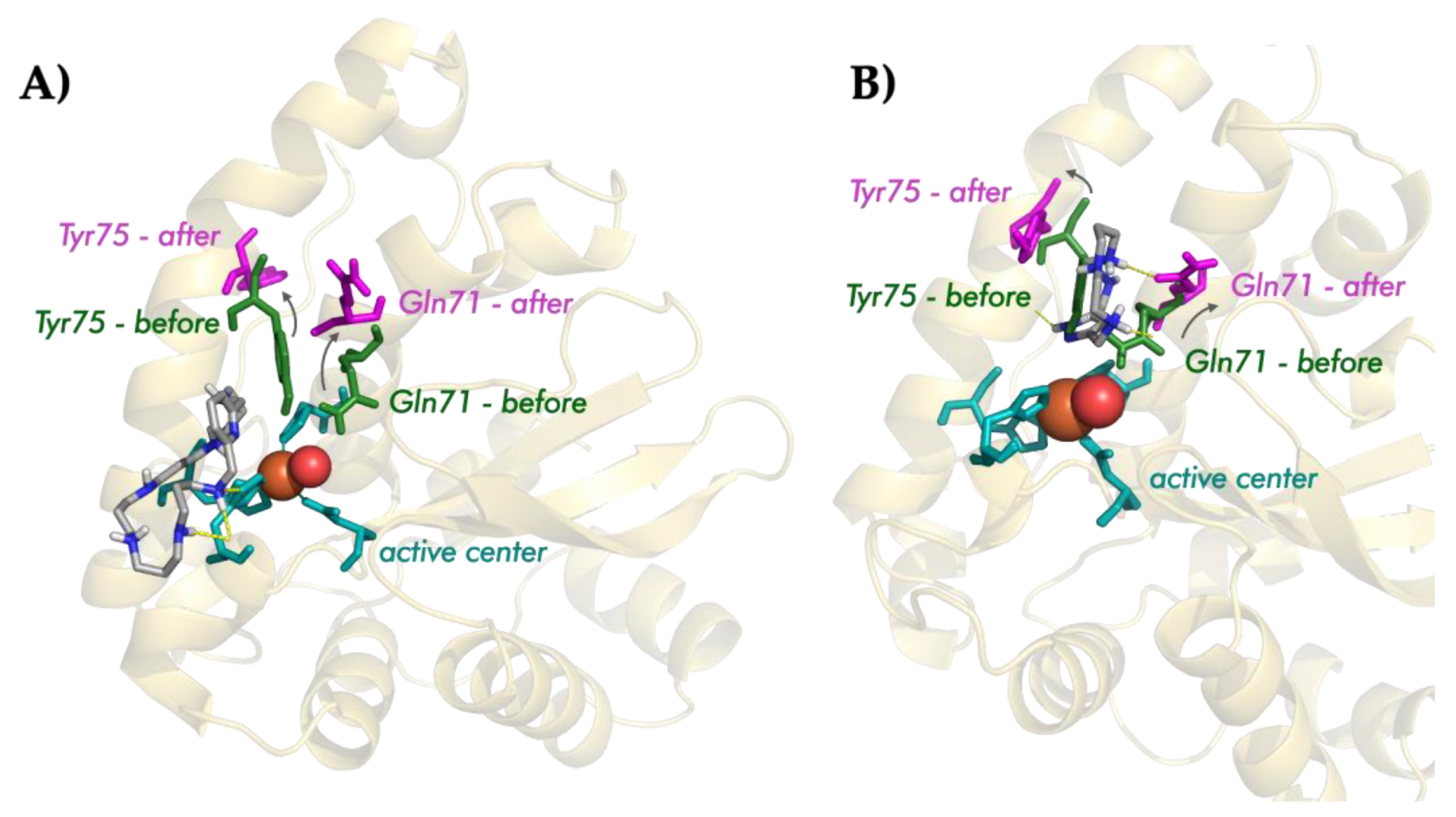
3.6.3. Interaction Mode of the Macrocycles
- (a)
- The three compounds are able to interact with the active center of the enzyme. Indeed, the nature of the interactions is the same in the three cases, i.e., hydrogen and ionic bonds;
- (b)
- The interaction mode differs depending on the structure of the macrocycle: the pyrazole ligands tend to interact with the residues of the hydrogen-bond network, while the pyridine one interacts directly with the histidines of the active center.
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Burza, S.; Croft, S.L.; Boelaert, M. Leishmaniasis. Lancet 2018, 392, 951–970. [Google Scholar] [CrossRef]
- Leishmaniasis. Available online: https://www.who.int/es/news-room/fact-sheets/detail/leishmaniasis (accessed on 15 November 2022).
- Navarro, P.; Sánchez-Moreno, M.; Marín, C.; García-España, E.; Ramirez-Macías, I.; Olmo, F.; Rosales, M.J.; Gómez-Contreras, F.; Yunta, M.J.R.; Gutiérrez-Sánchez, R. In vitro leishmanicidal activity of pyrazole-containing polyamine macrocycles which inhibit the Fe-SOD enzyme of Leishmania infantum and Leishmania braziliensis species. Parasitology 2014, 141, 1031–1043. [Google Scholar] [CrossRef]
- Marín, C.; Inclán, M.; Ramírez-Macías, I.; Albelda, M.T.; Cañas, R.; Clares, M.P.; Gonzalez-Garcia, J.; Rosales, M.J.; Urbanova, K.; García-España, E.; et al. In vitro antileishmanial activity of aza-scorpiand macrocycles. Inhibition of the antioxidant enzyme iron superoxide dismutase. RSC Adv. 2016, 6, 17446–17455. [Google Scholar] [CrossRef]
- Martín-Montes, A.; Clares, M.P.; Martín-Escolano, R.; Delgado-Pinar, E.; Marín, C.; Verdejo, B.; Martínez-Camarena, A.; Molina-Carreño, D.; García-España, E.; Sánchez-Moreno, M. Heterocyclic Diamines with Leishmanicidal Activity. ACS Infect. Dis. 2021, 7, 3168–3181. [Google Scholar] [CrossRef]
- García, M.P.; García-España, E.; Blasco, S.; Soriano, C.; González-GarcÍa, J.; Verdejo, B.; Inclán, M. Compuestos Macrocíclicos de Tipo Escorpiando y su uso Como Antiparasitarios. España Patent ES2414291B2, 13 February 2014. [Google Scholar]
- Arán, V.; Navarro, P.; Sánchez-Moreno, M.; Marín, C.; Olmo, F.; Ramirez-Macías, I.; García-España, E.; Albelda, M.T. Use of Ester Derivatives of Pyrazole Proton-Ionizable Compounds and the Corresponding Salts Thereof for the Treatment of Chagas Disease and Leishmaniasis. WO 2016/038238, PCT/Es/2015/070658. 17 March 2016. [Google Scholar]
- González, P.; Marín, C.; Rodríguez-González, I.; Hitos, A.B.; Rosales, M.J.; Reina, M.; Díaz, J.G.; González-Coloma, A.; Sánchez-Moreno, M. In vitro activity of C20-diterpenoid alkaloid derivatives in promastigotes and intracellular amastigotes of Leishmania infantum. Int. J. Antimicrob. Agents 2005, 25, 136–141. [Google Scholar] [CrossRef] [PubMed]
- Kirkinezos, I.G.; Moraes, C.T. Reactive oxygen species and mitochondrial diseases. Semin. Cell Dev. Biol. 2001, 12, 449–457. [Google Scholar] [CrossRef] [PubMed]
- Martin-Montes, A.; Santivañez-Veliz, M.; Moreno-Viguri, E.; Martín-Escolano, R.; Jiménez-Montes, C.; Lopez-Gonzalez, C.; Marín, C.; Sanmartín, C.; Gutiérrez Sánchez, R.; Sánchez-Moreno, M.; et al. In vitro antileishmanial activity and iron superoxide dismutase inhibition of arylamine Mannich base derivatives. Parasitology 2017, 144, 1783–1790. [Google Scholar] [CrossRef]
- Beyer, W.F.; Fridovich, I. Assaying for superoxide dismutase activity: Some large consequences of minor changes in conditions. Anal. Biochem. 1987, 161, 559–566. [Google Scholar] [CrossRef] [PubMed]
- Fernández-Becerra, C.; Sánchez-Moreno, M.; Osuna, A.; Opperdoes, F.R. Comparative aspects of energy metabolism in plant trypanosomatids. J. Eukaryot. Microbiol. 1997, 44, 523–529. [Google Scholar] [CrossRef]
- Sandes, J.M.; Fontes, A.; Regis-da-Silva, C.G.; de Castro, M.C.A.B.; Lima-Junior, C.G.; Silva, F.P.L.; Vasconcellos, M.L.A.A.; Figueiredo, R.C.B.Q. Trypanosoma cruzi cell death induced by the Morita-Baylis-Hillman adduct 3-hydroxy-2-methylene-3-(4-nitrophenylpropanenitrile). PLoS ONE 2014, 9, e93936. [Google Scholar] [CrossRef] [Green Version]
- Case, D.A.; Betz, R.M.; Cerutti, D.S.; III, T.E.C.; Darden, T.A.; Duke, R.E.; Giese, T.J.; Gohlke, H.; Goetz, A.W.; Homeyer, N.; et al. AMBER 2016; (Version 16); University of California: San Francisco, CA, USA, 2016. [Google Scholar]
- Bachega, J.F.R.; Navarro, M.V.A.S.; Bleicher, L.; Bortoleto-Bugs, R.K.; Dive, D.; Hoffmann, P.; Viscogliosi, E.; Garratt, R.C. Systematic structural studies of iron superoxide dismutases from human parasites and a statistical coupling analysis of metal binding specificity. Proteins Struct. Funct. Bioinform. 2009, 77, 26–37. [Google Scholar] [CrossRef]
- Berman, H.M.; Battistuz, T.; Bhat, T.N.; Bluhm, W.F.; Bourne, P.E.; Burkhardt, K.; Feng, Z.; Gilliland, G.L.; Iype, L.; Jain, S.; et al. The Protein Data Bank. Acta Crystallogr. Sect. D Biol. Crystallogr. 2002, 58, 899–907. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.M.; Wolf, R.M.; Caldwell, J.W.; Kollman, P.A.; Case, D.A. Development and Testing of a General Amber Force Field. J. Comput. Chem. 2004, 25, 1157–1174. [Google Scholar] [CrossRef] [PubMed]
- Maier, J.A.; Martinez, C.; Kasavajhala, K.; Wickstrom, L.; Hauser, K.E.; Simmerling, C. Ff14SB: Improving the Accuracy of Protein Side Chain and Backbone Parameters from Ff99SB. J. Chem. Theory Comput. 2015, 11, 3696–3713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; (Version 4.2.2); R Foundation for Statistical Computing: Vienna, Austria, 2021. [Google Scholar]
- Roe, D.R.; Cheatham III, T.E. PTRAJ and CPPTRAJ: Software for Processing and Analysis of Molecular Synamics Trajectory Data. J. Chem. Theory. Comput. 2013, 9, 3084–3095. [Google Scholar] [CrossRef]
- Schrodinger LLC. The PyMOL Molecular Graphics System; (Version 1.8); Schrodinger LLC: New York, NY, USA, 2015. [Google Scholar]
- Richman, J.E.; Atkins, T.J.; Oetle, W.F. Organic Synthesis; J. Wiley & Sons: New York, NY, USA, 1998; Volume 6, p. 652. [Google Scholar]
- Díaz, P.; Basallote, M.G.; Máñez, M.A.; García-España, E.; Gil, L.; Latorre, J.; Soriano, C.; Verdejo, B.; Luis, S.V. Thermodynamic and kinetic studies on the Cu2+ coordination chemistry of a novel binucleating pyridinophane ligand. Dalton Trans. 2003, 1186–1193. [Google Scholar] [CrossRef]
- Aguilar, J.; Basallote, M.G.; Gil, L.; Hernández, J.C.; Máñez, M.A.; García-España, E.; Soriano, C.; Verdejo, B. Stability and kinetics of the acid-promoted decomposition of Cu(II) complexes with hexaazacyclophanes: Kinetic studies as a probe to detect changes in the coordination mode of the macrocycles. Dalton Trans. 2004, 94–103. [Google Scholar] [CrossRef]
- Basallote, M.G.; Doménech, A.; Ferrer, A.; García-España, E.; Llinares, J.M.; Mañez, M.A.; Soriano, C.; Verdejo, B. Synthesis and Cu(II) coordination of two new hexaamines containing alternated propylenic and ethylenic chains: Kinetic studies on pH-driven metal ion slippage movements. Inorg. Chim. Acta 2006, 359, 2004–2014. [Google Scholar] [CrossRef]
- Belda, R.; Blasco, S.; Verdejo, B.; Jiménez, H.R.; Doménech-Carbó, A.; Soriano, C.; Latorre, J.; Terencio, C.; García-España, E. Homo- and heterobinuclear Cu2+ and Zn2+ complexes of abiotic cyclic hexaazapyridinocyclophanes as SOD mimics. Dalton Trans. 2013, 42, 11194–11204. [Google Scholar] [CrossRef]
- Verdejo, B.; Basallote, M.G.; Ferrer, A.; Mañez, M.A.; Hernández, J.C.; Chadim, M.; Hodacová, J.; Llinares, J.M.; Soriano, C.; García-España, E. Equilibrium and kinetic properties of Cu(II) cyclophane complexes: The effect of changes in the macrocyclic cavity caused by changes in the substitution at the aromatic ring. Eur. J. Inorg. Chem. 2008, 2008, 1497–1507. [Google Scholar] [CrossRef]
- Algarra, A.G.; Basallote, M.G.; Belda, R.; Blasco, S.; Castillo, C.E.; Llinares, J.M.; García-España, E.; Gil, L.; Máñez, M.A.; Soriano, C.; et al. Synthesis, protonation and Cu(II) complexes of two novel isomeric pentaazacyclophane ligands: Potentiometric, DFT, kinetic and AMP recognition studies. Eur. J. Inorg. Chem. 2009, 2009, 62–75. [Google Scholar] [CrossRef]
- Belda, R.; Pitarch-Jarque, J.; Soriano, C.; Llinares, J.M.; Blasco, S.; Ferrando-Soria, J.; García-España, E. Intermolecular binding modes in a novel [1+1] condensation 1H-pyrazole azamacrocycle: A solution and solid state study with evidence for CO2 fixation. Inorg. Chem. 2013, 52, 10795–10803. [Google Scholar] [CrossRef]
- Lopera, A.; Gil-Martínez, A.; Pitarch-Jarque, J.; Verdejo, B.; Blasco, S.; Clares, M.P.; Jiménez, H.R.; García-España, E. Influence of the chain length and metal: Ligand ratio on the self-organization processes of Cu2+ complexes of [1+1] 1H-pyrazole azamacrocycles. Dalton Trans. 2020, 49, 8614–8624. [Google Scholar] [CrossRef] [PubMed]
- Belda, R.; García-España, E.; Morris, G.A.; Steed, J.W.; Aguilar, J.A. Guanosine-5′-monophosphate polyamine hybrid hydrogels: Enhanced gel strength probed by z-spectroscopy. Chem. Eur. J. 2017, 23, 7755–7760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nwaka, S.; Ramirez, B.; Brun, R.; Maes, L.; Douglas, F.; Ridley, R. Advancing drug innovation for neglected diseases—Criteria for lead progression. PLoS Neglect. Trop. Dis. 2009, 3, e440. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound Codes | Compound IC50 (μM) | Macrophage Macrophage Toxicity IC50 (µM) | Selectivity Index | ||
|---|---|---|---|---|---|
| Promastigote Forms | Amastigote Forms | Promastigote Forms | Amastigote Forms | ||
| Glucantime | 50.1 ± 2.3 | 84.6 ± 4.3 | 175.8 ± 10.6 | 3.5 | 2 |
| Py3223 | 191.9 ± 5.8 | nd | 165.3 ± 4.9 | 1 (0) | nd |
| Py33233 | 484.0 ± 10.7 | nd | 1292.3 ± 77.5 | 3 (1) | nd |
| Py32323 | 90.6 ± 3.8 | nd | 85.4 ± 4.6 | 1 (0) | nd |
| Py32223 | 93.4 ± 3.7 | nd | 57.7 ± 1.9 | 1 (0) | nd |
| Py33333 | 8.51 ± 0.6 | 1.7 ± 0.2 | 1500.0 ± 75.3 | 176 (44) | 882 (441) |
| mB323 | 95.6 ± 4.8 | nd | 428.5 ± 22.6 | 4 (1) | nd |
| mB333 | 86.1 ± 3.7 | nd | 915.9 ± 51.6 | 11 (3) | nd |
| mB232 | 205.1 ± 30.7 | nd | 667.1 ± 88.5 | 3 (1) | nd |
| pB232 | 367.2 ± 10.5 | nd | 719.1 ± 20.5 | 2 (1) | nd |
| PZ333 | 70.0 ± 0.7 | nd | 984.9 ± 72.8 | 14 (4) | nd |
| PZ232 | 13.0 ± 0.4 | 4.9 ± 0.4 | 673.8 ± 66.3 | 52 (13) | 138 (69) |
| mB3223 | 63.4 ± 2.4 | nd | 1450.7 ± 89.7 | 23 (6) | nd |
| Py333 | 30.4 ± 1.4 | nd | 309.9 ± 43.8 | 10 (3) | nd |
| Py34 | 56.7 ± 2.6 | nd | 568.9 ± 45.8 | 10 (3) | nd |
| Compound Codes | Compound IC50 (μM) | Macrophage Toxicity IC50 (µM) | Selectivity Index | ||
|---|---|---|---|---|---|
| Promastigote Forms | Amastigote Forms | Promastigote Forms | Amastigote Forms | ||
| Glucantime | 51.4 ± 2.5 | 103.3 | 175.8 ± 10.6 | 3 | 1.7 |
| Py3223 | 290.7 ± 3.8 | nd | 165.3 ± 4.9 | 1 (0) | nd |
| Py33233 | 507.1 ± 8.3 | nd | 1292.3 ± 77.5 | 3 (1) | nd |
| Py32323 | 1634.3 ± 76.4 | nd | 85.4 ± 4.6 | 0 (0) | nd |
| Py32223 | 441.7 ± 5.8 | nd | 57.7 ± 1.9 | 0 (0) | nd |
| Py33333 | 36.5 ± 1.1 | nd | 1500.0 ± 75.3 | 41 (14) | nd |
| mB323 | 2.3 ± 0.5 | 17.1 ± 1.1 | 428.5 ± 22.6 | 186 (62) | 25 (13) |
| mB333 | 209.6 ± 5.3 | nd | 915.9 ± 51.6 | 4 (1) | nd |
| mB232 | 91.7 ± 3.7 | nd | 667.1 ± 88.5 | 7 (2) | nd |
| pB232 | 27.2 ± 0.9 | nd | 719.1 ± 20.5 | 26 (9) | nd |
| PZ333 | 15.7 ± 0.5 | nd | 984.9 ± 72.8 | 63 (21) | nd |
| PZ232 | 5.8 ± 0.2 | 1.6 ± 0.6 | 673.8 ± 66.3 | 116 (39) | 421 (211) |
| mB3223 | 15.6 ± 0.7 | nd | 1450.7 ± 89.7 | 93 (31) | nd |
| Py333 | 26.2 ± 1.1 | nd | 309.9 ± 43.8 | 12 (4) | nd |
| Py34 | 29.2 ± 0.8 | nd | 568.9 ± 45.8 | 19 (6) | nd |
| Compound Codes | Compound IC50 (μM) | Macrophage Toxicity IC50 (µM) | Selectivity Index | ||
|---|---|---|---|---|---|
| Promastigote Forms | Amastigote Forms | Promastigote Forms | Amastigote Forms | ||
| Glucantime | 23.7 ± 0.8 | 76.4 ± 4.1 | 175.8 ± 10.6 | 7 | 2 |
| Py3223 | 52.5 ± 2.1 | nd | 165.3 ± 4.9 | 3 (0) | nd |
| Py33233 | 583.0 ± 6.9 | nd | 1292.3 ± 77.5 | 2 (0) | nd |
| Py32323 | 130.2 ± 9.5 | nd | 85.4 ± 4.6 | 1 (0) | nd |
| Py32223 | 1000.0 ± 78.4 | nd | 57.7 ± 1.9 | 0 (0) | nd |
| Py33333 | 70.5 ± 5.2 | nd | 1500.0 ± 75.3 | 21 (3) | nd |
| mB323 | 158.1 ± 7.3 | nd | 428.5 ± 22.6 | 3 (0) | nd |
| mB333 | 105.6 ± 7.5 | nd | 915.9 ± 51.6 | 9 (1) | nd |
| mB232 | 171.2 ± 10.4 | nd | 667.1 ± 88.5 | 4 (1) | nd |
| pB232 | 742.9 ± 23.6 | nd | 719.1 ± 20.5 | 1 (0) | nd |
| PZ333 | 1.4 ± 0.3 | 1.7 ± 0.5 | 984.9 ± 72.8 | 704 (101) | 579 (290) |
| PZ232 | 25.0 ± 0.8 | nd | 673.8 ± 66.3 | 27 (4) | nd |
| mB3223 | 126.5 ± 3.8 | nd | 1450.7 ± 89.7 | 11 (2) | nd |
| Py333 | 0.2 ± 0.0 | 16.8 ± 0.5 | 309.9 ± 43.8 | 1549 (221) | 18 (9) |
| Py34 | 28.4 ± 0.6 | nd | 568.9 ± 45.8 | 20 (3) | nd |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martín-Montes, Á.; Martínez-Camarena, Á.; Lopera, A.; Bonastre-Sabater, I.; Clares, M.P.; Verdejo, B.; García-España, E.; Marín, C. The Bioactivity of Xylene, Pyridine, and Pyrazole Aza Macrocycles against Three Representative Leishmania Species. Pharmaceutics 2023, 15, 992. https://doi.org/10.3390/pharmaceutics15030992
Martín-Montes Á, Martínez-Camarena Á, Lopera A, Bonastre-Sabater I, Clares MP, Verdejo B, García-España E, Marín C. The Bioactivity of Xylene, Pyridine, and Pyrazole Aza Macrocycles against Three Representative Leishmania Species. Pharmaceutics. 2023; 15(3):992. https://doi.org/10.3390/pharmaceutics15030992
Chicago/Turabian StyleMartín-Montes, Álvaro, Álvaro Martínez-Camarena, Alberto Lopera, Irene Bonastre-Sabater, M. Paz Clares, Begoña Verdejo, Enrique García-España, and Clotilde Marín. 2023. "The Bioactivity of Xylene, Pyridine, and Pyrazole Aza Macrocycles against Three Representative Leishmania Species" Pharmaceutics 15, no. 3: 992. https://doi.org/10.3390/pharmaceutics15030992