
Auranofin Targeting the NDM-1 Beta-Lactamase: Computational Insights into the Electronic Configuration and Quasi-Tetrahedral Coordination of Gold Ions


Abstract
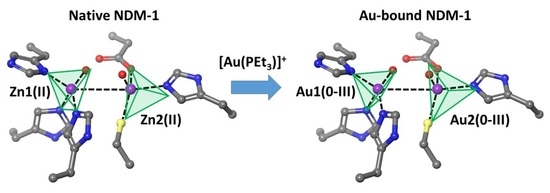
:
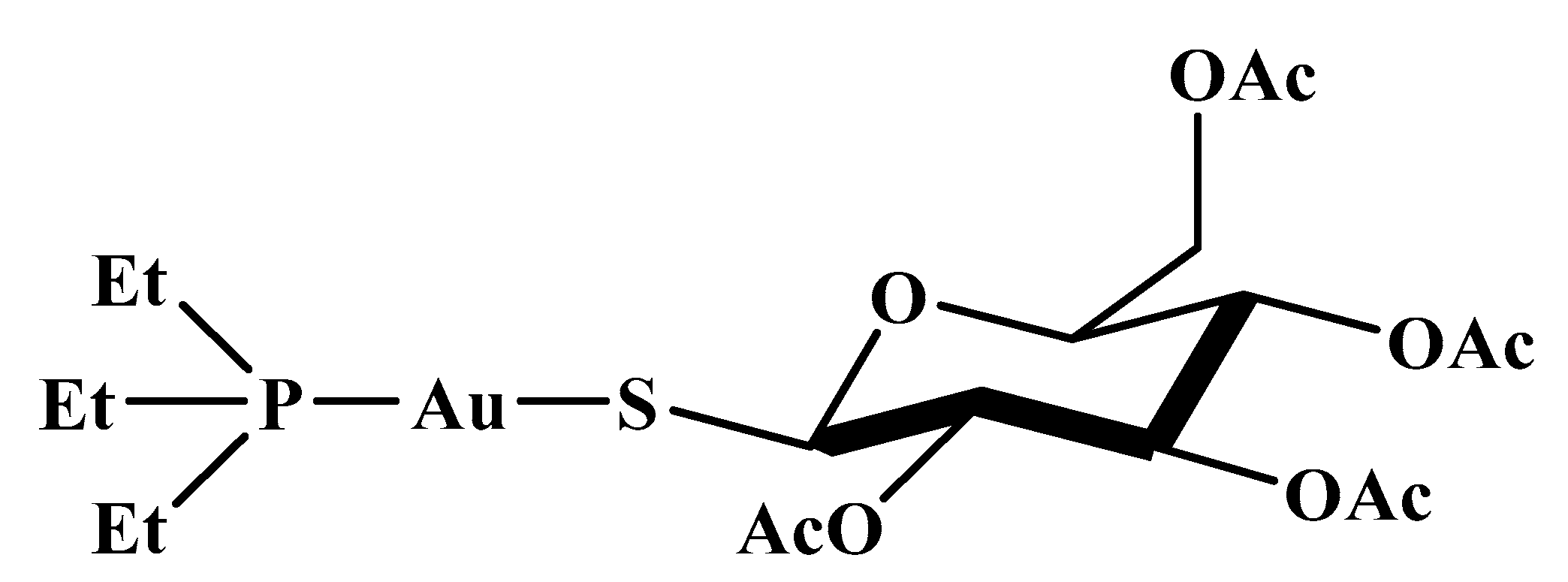
1. Introduction
2. Computational Details
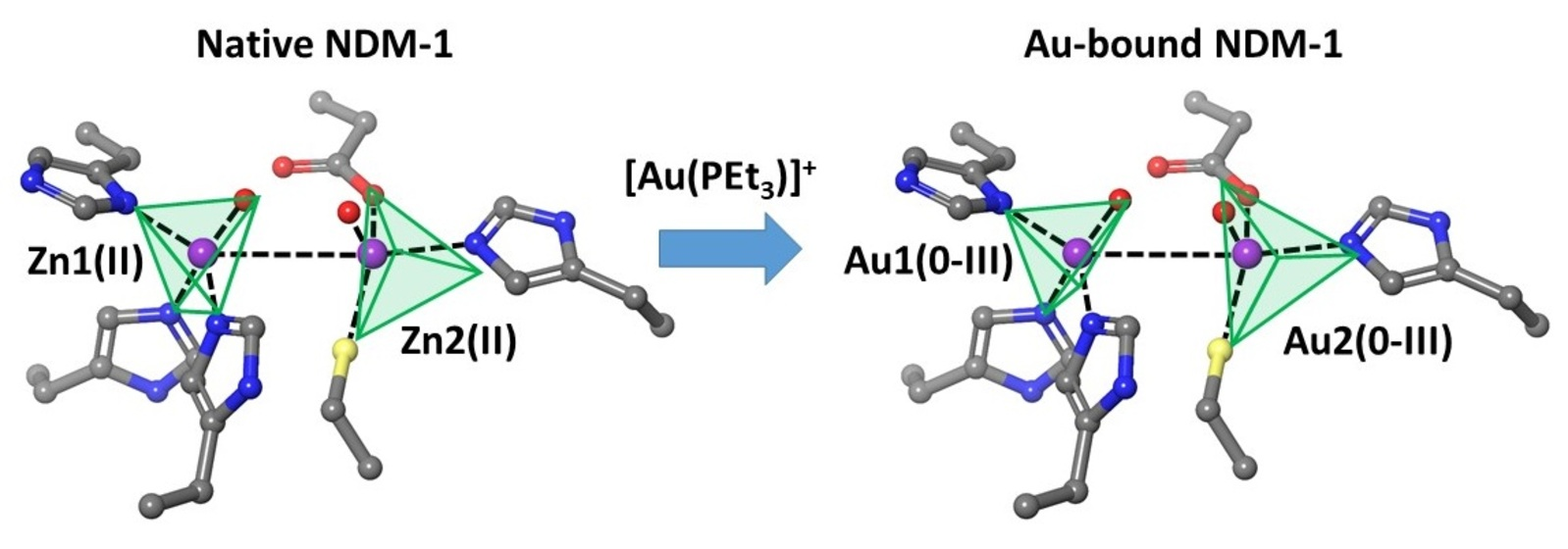
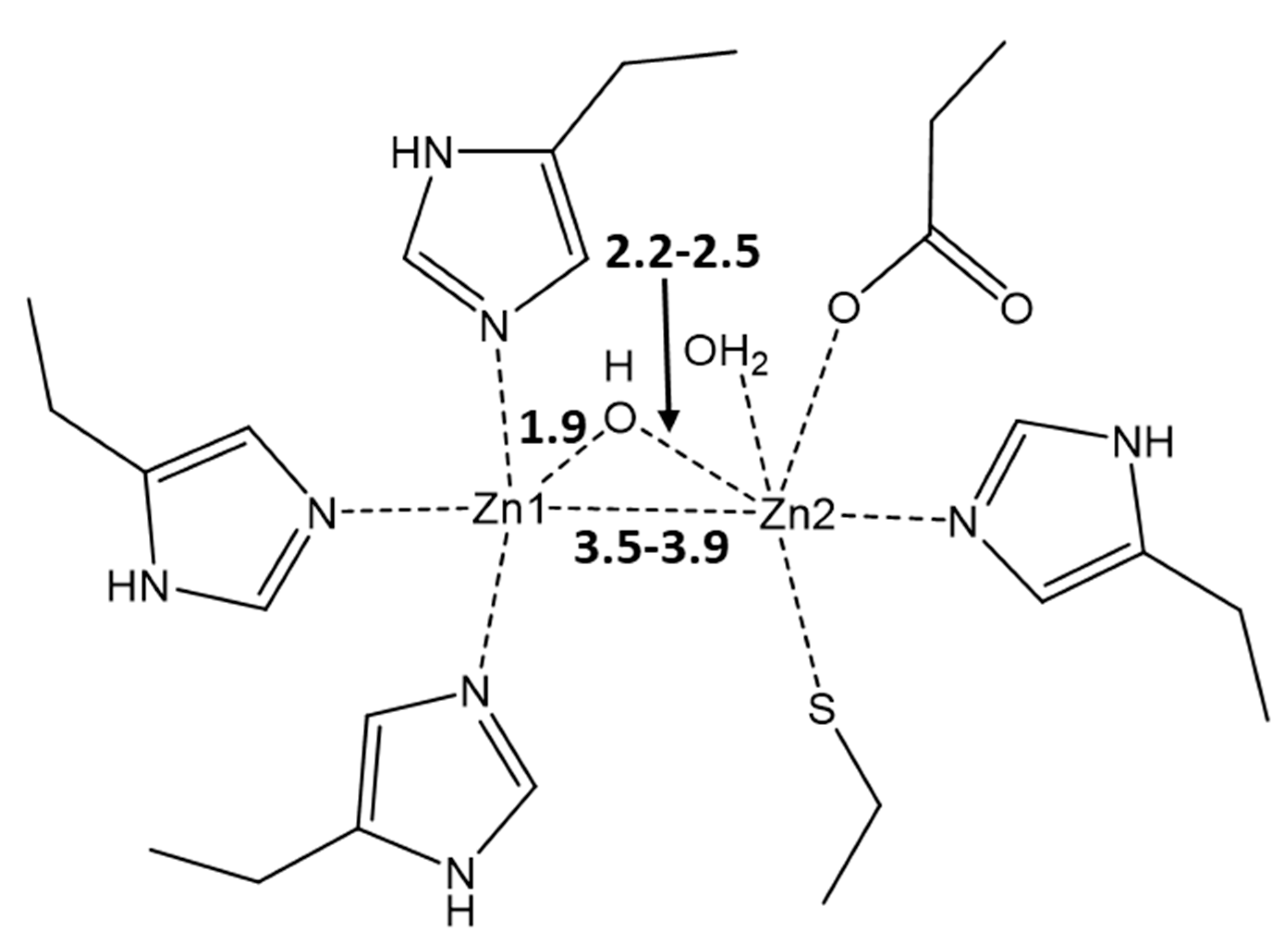
3. Results and Discussion
4. Summary
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaul, G.; Shukla, M.; Dasgupta, A.; Chopra, S. Update on drug-repurposing: Is it useful for tackling antimicrobial resistance? Future Microbiol. 2019, 14, 829–831. [Google Scholar] [CrossRef] [Green Version]
- Gupta, N.; Limbago, B.M.; Patel, J.B.; Kallen, A.J. Carbapenem-resistant Enterobacteriaceae: Epidemiology and prevention. Clin. Infect. Dis. 2011, 53, 60–67. [Google Scholar] [CrossRef] [Green Version]
- Bahr, G.; Gonzalez, L.J.; Vila, A.J. Metallo-β-lactamases in the age of multidrug resistance: From structure and mechanism to evolution, dissemination, and inhibitor design. Chem. Rev. 2021, 121, 7957–8094. [Google Scholar] [CrossRef] [PubMed]
- Bahr, G.; González, L.J.; Vila, A.J. Metallo-β-lactamases and a tug-of-war for the available zinc at the host–pathogen interface. Curr. Opin. Chem. Biol. 2022, 66, 102103. [Google Scholar] [CrossRef]
- Li, H.; Sun, H. A hydroxide lock for metallo-β-lactamases. Nat. Chem. 2022, 14, 6–8. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Hu, L.; Sankaran, B.; Prasad, B.V.; Palzkill, T. Differential active site requirements for NDM-1 β-lactamase hydrolysis of carbapenem versus penicillin and cephalosporin antibiotics. Nat. Commun. 2018, 9, 4524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Seny, D.; Prosperi-Meys, C.; Bebrone, C.; Rossolini, G.M.; Page, M.I.; Noel, P.; Frere, J.M.; Galleni, M. Mutational analysis of the two zinc-binding sites of the Bacillus cereus 569/H/9 metallo-β-lactamase. Biochem. J. 2002, 363, 687–696. [Google Scholar] [CrossRef]
- Linciano, P.; Cendron, L.; Gianquinto, E.; Spyrakis, F.; Tondi, D. Ten years with New Delhi metallo-β-lactamase-1 (NDM-1): From structural insights to inhibitor design. ACS Infect. Dis. 2018, 5, 9–34. [Google Scholar] [CrossRef] [PubMed]
- Lisa, M.N.; Palacios, A.R.; Aitha, M.; González, M.M.; Moreno, D.M.; Crowder, M.W.; Bonomo, R.A.; Spencer, J.; Tierney, D.L.; Llarrull, L.I.; et al. A general reaction mechanism for carbapenem hydrolysis by mononuclear and binuclear metallo-β-lactamases. Nat. Commun. 2017, 8, 538. [Google Scholar] [CrossRef] [Green Version]
- Tioni, M.F.; Llarrull, L.I.; Poeylaut-Palena, A.A.; Martí, M.A.; Saggu, M.; Periyannan, G.R.; Mata, E.G.; Bennett, B.; Murgida, D.H.; Vila, A.J. Trapping and characterization of a reaction intermediate in carbapenem hydrolysis by B. cereus metallo-β-lactamase. J. Am. Chem. Soc. 2008, 130, 15852–15863. [Google Scholar] [CrossRef] [Green Version]
- Rasia, R.M.; Vila, A.J. Structural determinants of substrate binding to Bacillus cereus metallo-β-lactamase. J. Biol. Chem. 2004, 279, 26046–26051. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Ma, G.; Zhu, Y.; Zeng, L.; Ahmad, A.; Wang, C.; Pang, B.; Fang, H.; Zhao, L.; Hao, Q. Active-site conformational fluctuations promote the enzymatic activity of NDM-1. Antimicrob. Agents Chemother. 2018, 62, e01579-18. [Google Scholar] [CrossRef] [Green Version]
- King, A.M.; Reid-Yu, S.A.; Wang, W.; King, D.T.; De Pascale, G.; Strynadka, N.C.; Walsh, T.R.; Coombes, B.K.; Wright, G.D. Aspergillomarasmine A overcomes metallo-β-lactamase antibiotic resistance. Nature 2014, 510, 503–506. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolbatov, I.; Storchi, L.; Marrone, A. Structural reshaping of the zinc-finger domain of the SARS-CoV-2 nsp13 protein using bismuth (III) ions: A multilevel computational study. Inorg. Chem. 2022, 61, 15664–15677. [Google Scholar] [CrossRef]
- Sun, H.; Zhang, Q.; Wang, R.; Wang, H.; Wong, Y.T.; Wang, M.; Hao, Q.; Yan, A.; Kao, R.Y.; Ho, P.L.; et al. Resensitizing carbapenem-and colistin-resistant bacteria to antibiotics using auranofin. Nat. Commun. 2020, 11, 5263. [Google Scholar] [CrossRef] [PubMed]
- Roder, C.; Thomson, M.J. Auranofin: Repurposing an old drug for a golden new age. Drugs RD 2015, 15, 13–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tolbatov, I.; Marrone, A.; Coletti, C.; Re, N. Computational studies of Au (I) and Au (III) anticancer metallodrugs: A survey. Molecules 2021, 26, 7600. [Google Scholar] [CrossRef]
- Paciotti, R.; Tolbatov, I.; Marrone, A.; Storchi, L.; Re, N.; Coletti, C. Computational investigations of bioinorganic complexes: The case of calcium, gold and platinum ions. In AIP Conference Proceedings; AIP Publishing LLC: Melville, NY, USA, 2019; Volume 2186, p. 030011. [Google Scholar] [CrossRef]
- Hathaway, B.J.; Wilkinson, G.; Gillard, R.D.; McCleverty, J.A. Comprehensive Coordination Chemistry. In The Synthesis, Reactions, Properties and Applications of Coordination Compounds; Pergamon Press: Oxford, UK, 1987; pp. 533–774. [Google Scholar]
- Gimeno, M.C.; Laguna, A. Three-and four-coordinate gold (I) complexes. Chem. Rev. 1997, 97, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Khan, M.N.I.; King, C.; Heinrich, D.D.; Fackler, J.P., Jr.; Porter, L.C. Syntheses and crystal structures (No gold-hydrogen interactions) of gold phosphine luminescent complexes, [Au2(dppm)2][BH3CN]2 and [Au2(dppm)2(I)][Au(CN)2] and [Au2(dppm)2(S2CNEt2)][BH3CN]. Inorg. Chem. 1989, 28, 2150–2154. [Google Scholar] [CrossRef]
- Davila, R.M.; Elduque, A.; Grant, T.; Staples, R.J.; Fackler, J.P., Jr. Synthesis and characterization of dinuclear gold (I) ring and open-ring complexes containing saturated and unsaturated dithiol bridging ligands and phosphine or bis (diphosphine) donor ligands. Crystal structures of [Au2(µ-S(CH2)3S)(µ-dppm)], [Au2(µ-MNT)(PPh3)2], [Au2(µ-S2C6H4)(PPh3)2], and [Au4(µ-S2C6H3CH3)2(PEt3)2]. Inorg. Chem. 1993, 32, 1749–1755. [Google Scholar] [CrossRef]
- Yamashita, M. Auranofin: Past to present, and repurposing. Int. Immunopharmacol. 2021, 101, 108272. [Google Scholar] [CrossRef]
- Palermo, G.; Spinello, A.; Saha, A.; Magistrato, A. Frontiers of metal-coordinating drug design. Expert Opin. Drug Discov. 2021, 16, 497–511. [Google Scholar] [CrossRef] [PubMed]
- Pickering, I.J.; Cheng, Q.; Rengifo, E.M.; Nehzati, S.; Dolgova, N.V.; Kroll, T.; Sokaras, D.; George, G.N.; Arnér, E.S. Direct observation of methylmercury and auranofin binding to selenocysteine in thioredoxin reductase. Inorg. Chem. 2020, 59, 2711–2718. [Google Scholar] [CrossRef]
- Abhishek, S.; Sivadas, S.; Satish, M.; Deeksha, W.; Rajakumara, E. Dynamic basis for auranofin drug recognition by thiol-reductases of human pathogens and intermediate coordinated adduct formation with catalytic cysteine residues. ACS Omega 2019, 4, 9593–9602. [Google Scholar] [CrossRef] [PubMed]
- Dos Santos, H.F. Reactivity of auranofin with S-, Se-and N-containing amino acids. Comput. Theor. Chem. 2014, 1048, 95–101. [Google Scholar] [CrossRef]
- Scoditti, S.; Chiodo, F.; Mazzone, G.; Richeter, S.; Sicilia, E. Porphyrins and metalloporphyrins combined with N-heterocyclic carbene (NHC) gold (I) complexes for photodynamic therapy application: What is the weight of the heavy atom effect? Molecules 2022, 27, 4046. [Google Scholar] [CrossRef] [PubMed]
- Pessoa, J.C.; Santos, M.F.; Correia, I.; Sanna, D.; Sciortino, G.; Garribba, E. Binding of vanadium ions and complexes to proteins and enzymes in aqueous solution. Coord. Chem. Rev. 2021, 449, 214192. [Google Scholar] [CrossRef]
- Ugone, V.; Sanna, D.; Sciortino, G.; Crans, D.C.; Garribba, E. ESI-MS study of the interaction of potential oxidovanadium (IV) drugs and amavadin with model proteins. Inorg. Chem. 2020, 59, 9739–9755. [Google Scholar] [CrossRef]
- Frisch, M.J.; Trucks, G.W.; Schlegel, H.B.; Scuseria, G.E.; Robb, M.A.; Cheeseman, J.R.; Scalmani, G.; Barone, V.; Petersson, G.A.; Nakatsuji, H.; et al. Gaussian 16 Revision C. 01. 2016; Gaussian Inc.: Wallingford, CT, USA, 2016; p. 421. [Google Scholar]
- Chai, J.D.; Head-Gordon, M. Systematic optimization of long-range corrected hybrid density functionals. J. Chem. Phys. 2008, 128, 084106. [Google Scholar] [CrossRef] [PubMed]
- Andrae, D.; Haeussermann, U.; Dolg, M.; Stoll, H.; Preuss, H. Energy-adjusted ab initio pseudopotentials for the second and third row transition elements. Theor. Chim. Acta 1990, 77, 123–141. [Google Scholar] [CrossRef]
- Weigend, F.; Ahlrichs, R. Balanced basis sets of split valence, triple zeta valence and quadruple zeta valence quality for H to Rn: Design and assessment of accuracy. Phys. Chem. Chem. Phys. 2005, 7, 3297–3305. [Google Scholar] [CrossRef]
- Tolbatov, I.; Marrone, A. Reactivity of N-heterocyclic carbene half-sandwich Ru-, Os-, Rh-, and Ir-based complexes with cysteine and selenocysteine: A computational study. Inorg. Chem. 2021, 61, 746–754. [Google Scholar] [CrossRef] [PubMed]
- Tolbatov, I.; Marrone, A. Reaction of dirhodium and diruthenium paddlewheel tetraacetate complexes with nucleophilic protein sites: A computational study. Inorg. Chim. Acta 2022, 530, 120684. [Google Scholar] [CrossRef]
- Tolbatov, I.; Marrone, A.; Paciotti, R.; Re, N.; Coletti, C. Multilayered modelling of the metallation of biological targets. In Proceedings of the International Conference on Computational Science and Its Applications, Cagliari, Italy, 13 September 2021; Springer: Cham, Switzerland, 2021; pp. 398–412. [Google Scholar] [CrossRef]
- Tolbatov, I.; Marrone, A. Kinetics of reactions of dirhodium and diruthenium paddlewheel tetraacetate complexes with nucleophilic protein sites: Computational insights. Inorg. Chem. 2022, 61, 16421–16429. [Google Scholar] [CrossRef] [PubMed]
- Tolbatov, I.; Marrone, A. Computational strategies to model the interaction and the reactivity of biologically-relevant transition metal complexes. Inorg. Chim. Acta 2022, 530, 120686. [Google Scholar] [CrossRef]
- Glendening, E.D.; Reed, A.E.; Carpenter, J.E.; Weinhold, F. Gaussian NBO Version 3.1; Gaussian Inc.: Pittsburgh, PA, USA, 2001. [Google Scholar]
- Tomasi, J.; Mennucci, B.; Cancès, E. The IEF version of the PCM solvation method: An overview of a new method addressed to study molecular solutes at the QM ab initio level. J. Mol. Struct. THEOCHEM 1999, 464, 211–226. [Google Scholar] [CrossRef]
- Abraham, M.J.; Murtola, T.; Schulz, R.; Páll, S.; Smith, J.C.; Hess, B.; Lindahl, E. GROMACS: High performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 2015, 1, 19–25. [Google Scholar] [CrossRef] [Green Version]
- Raczynska, J.E.; Shabalin, I.G.; Minor, W.; Wlodawer, A.; Jaskolski, M. A close look onto structural models and primary ligands of metallo-β-lactamases. Drug Resist. Updates 2018, 40, 1–2. [Google Scholar] [CrossRef] [PubMed]
- Irwin, M.D.; Abdou, H.E.; Mohamed, A.A.; Fackler, J.P., Jr. Synthesis and X-ray structures of silver and gold guanidinate-like complexes. A Au (ii) complex with a 2.47 Å Au–Au distance. Chem. Commun. 2003, 13, 2882–2883. [Google Scholar] [CrossRef]
- Abdou, H.E.; Mohamed, A.A.; Fackler, J.P. Synthesis and X-ray structures of dinuclear and trinuclear gold (I) and dinuclear gold (II) amidinate complexes. Inorg. Chem. 2005, 44, 166–168. [Google Scholar] [CrossRef]
- Coker, N.L.; Krause Bauer, J.A.; Elder, R.C. Emission energy correlates with inverse of gold− gold distance for various [Au(SCN)2]-salts. J. Am. Chem. Soc. 2004, 126, 12–13. [Google Scholar] [CrossRef] [PubMed]
- Kumar, K.; Stefanczyk, O.; Chorazy, S.; Nakabayashi, K.; Ohkoshi, S.I. Ratiometric and colorimetric optical thermometers using emissive dimeric and trimeric {[Au(SCN)2]−}n moieties generated in d–f heterometallic assemblies. Angew. Chem. 2022, 134, e202201265. [Google Scholar] [CrossRef]
- Mazzei, L.; Massai, L.; Cianci, M.; Messori, L.; Ciurli, S. Medicinal Au (I) compounds targeting urease as prospective antimicrobial agents: Unveiling the structural basis for enzyme inhibition. Dalton Trans. 2021, 50, 14444–14452. [Google Scholar] [CrossRef]
- Johnson, A.; Marzo, I.; Gimeno, M.C. Heterobimetallic propargyl gold complexes with π-bound copper or silver with enhanced anticancer activity. Dalton Trans. 2020, 49, 11736–11742. [Google Scholar] [CrossRef] [PubMed]
- Yamada, S.; Rokusha, Y.; Kawano, R.; Fujisawa, K.; Tsutsumi, O. Mesogenic gold complexes showing aggregation-induced enhancement of phosphorescence in both crystalline and liquid-crystalline phases. Faraday Discuss. 2017, 196, 269–283. [Google Scholar] [CrossRef]
- Sathyanarayana, A.; Siddhant, K.; Yamane, M.; Hisano, K.; Prabusankar, G.; Tsutsumi, O. Tuning the Au–Au interactions by varying the degree of polymerisation in linear polymeric Au (I) N-heterocyclic carbene complexes. J. Mat. Chem. C 2022, 10, 6050–6060. [Google Scholar] [CrossRef]
- Pyykkö, P. Theoretical chemistry of gold. Angew. Chem. Int. Ed. 2004, 43, 4412–4456. [Google Scholar] [CrossRef] [PubMed]
- Herrera, R.P.; Gimeno, M.C. Main avenues in gold coordination chemistry. Chem. Rev. 2021, 121, 8311–8363. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
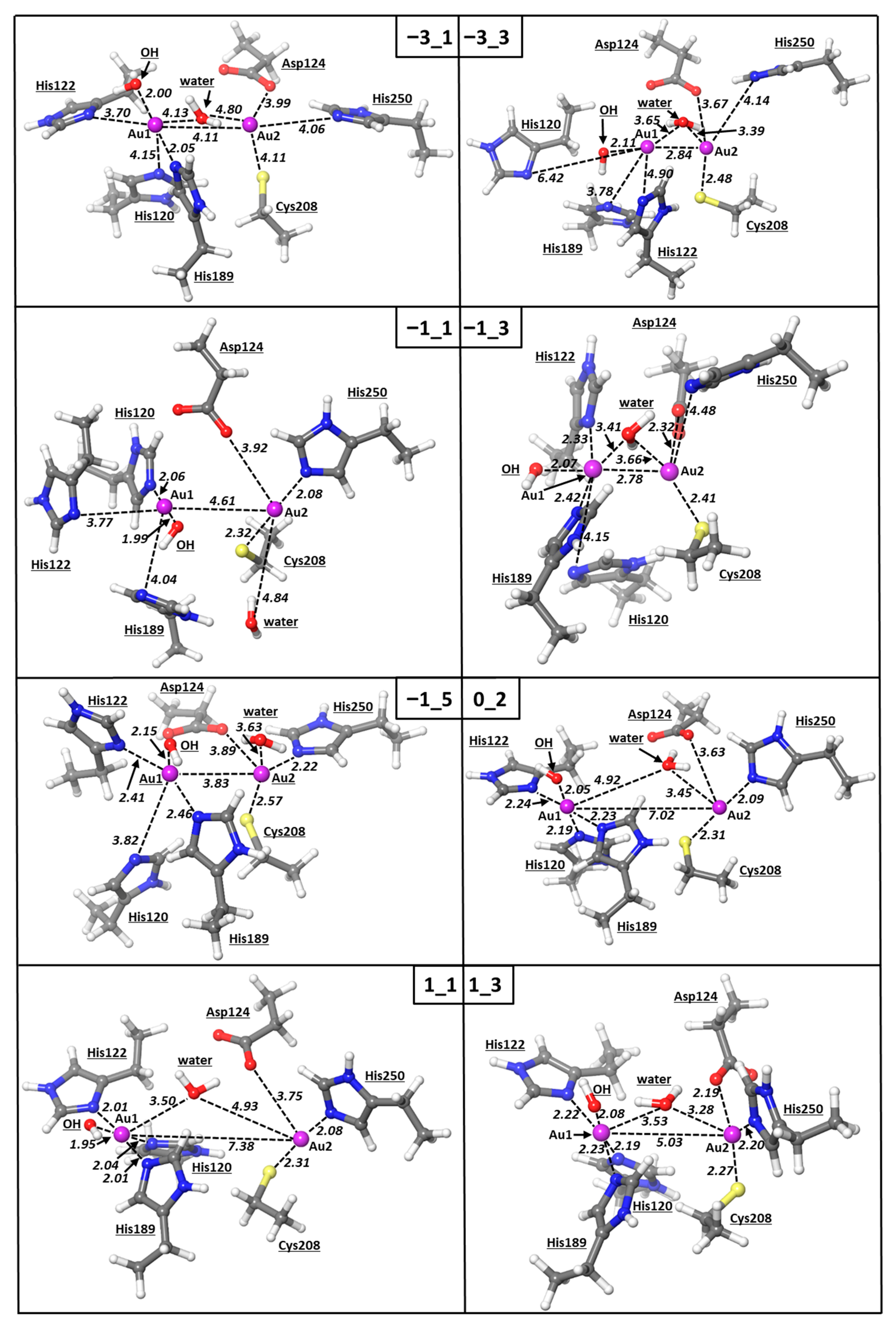{kind=link}
| Name of the Model | A | B | C | D | E | F | G | H |
|---|---|---|---|---|---|---|---|---|
| Charge_Multiplicity | -3_1 | -3_3 | -1_1 | -1_3 | -1_5 | 0_2 | 1_1 | 1_3 |
| Au-Au configuration | s2d10–d10 | s1d10–s1d10 | d10–d10 | s1d9–d10 | s1d9–s1d9 | d10–d9 s1d10–d8 | d10–d8 | d9–d9 |
| Au-Au oxidation state | Au(0)-Au(0) | Au(0)-Au(0) | Au(I)-Au(I) | Au(I)-Au(I) | Au(I)-Au(I) | Au(I)-Au(II) Au(0)-Au(III) | Au(I)-Au(III) | Au(II)-Au(II) |
| Distance | A | B | C | D | E | F | G | H | Exp * | |
|---|---|---|---|---|---|---|---|---|---|---|
| Au1 | Au2 | 4.11 | 2.84 | 4.61/3.55 | 2.78/2.62 | 3.83/5.96 | 7.02 | 7.38 | 5.03/4.30 | 3.76 |
| Au1 | N(His122) | 3.70 | 4.90 | 3.77/3.54 | 2.33/2.39 | 2.41/2.45 | 2.24 | 2.01 | 2.22/2.29 | 2.00 |
| N(His189) | 2.05 | 3.78 | 4.04/4.49 | 2.42/2.35 | 2.46/2.41 | 2.23 | 2.01 | 2.23/2.26 | 2.05 | |
| N(His120) | 4.15 | 6.42 | 2.06/2.04 | 4.15/3.96 | 3.82/3.70 | 2.19 | 2.04 | 2.19/2.16 | 2.50 | |
| O(OH) | 2.00 | 2.11 | 1.99/2.03 | 2.07/2.10 | 2.15/2.11 | 2.05 | 1.95 | 2.08/2.06 | 1.95 | |
| O(H2O) | 4.13 | 3.65 | 6.03/3.47 | 3.41/4.58 | 3.80/7.04 | 4.92 | 3.5 | 3.53/4.41 | 3.57 | |
| Au2 | S(Cys208) | 4.11 | 2.48 | 2.32/2.31 | 2.41/2.52 | 2.57/2.57 | 2.31 | 2.31 | 2.27/2.55 | 2.47 |
| N(His250) | 4.06 | 4.14 | 2.08/2.08 | 5.25/2.31 | 2.22/2.19 | 2.09 | 2.08 | 2.20/2.06 | 2.27 | |
| O(Asp124) | 3.99 | 3.67 | 3.92/3.66 | 2.32/3.61 | 3.89/3.55 | 3.63 | 3.75 | 2.19/2.09 | 2.23 | |
| O(OH) | 5.20 | 4.93 | 4.64/3.73 | 4.84/4.72 | 4.74/7.58 | 7.93 | 8.83 | 5.45/4.34 | 3.60 | |
| O(H2O) | 5.20 | 3.39 | 4.84/3.70 | 3.66/3.38 | 3.63/3.35 | 3.45 | 4.93 | 3.28/3.35 | 2.43 | |
| RMSD | 1.51 | 1.80 | 1.41/0.95 | 1.42/0.89 | 0.85/1.84 | 1.77 | 2.12 | 0.74/0.48 | ||
| Atom | B | D | E | F | H |
|---|---|---|---|---|---|
| Au1 | 0.41 | 0.86 | 1.32 | 0.57 | 0.60 |
| N(His122) | 0.00 | 0.14 | 0.14 | 0.12 | 0.13 |
| N(His189) | 0.00 | 0.13 | 0.15 | 0.12 | 0.12 |
| N(His120) | 0.00 | 0.00 | 0.00 | 0.06 | 0.08 |
| Au2 | 0.96 | 0.32 | 1.09 | 0.00 | 0.44 |
| S(Cys208) | 0.35 | 0.16 | 0.67 | 0.00 | 0.41 |
| N(His250) | 0.01 | 0.00 | 0.19 | 0.00 | 0.15 |
| O(Asp124) | 0.01 | 0.05 | 0.01 | 0.00 | 0.06 |
| O(OH) | 0.20 | 0.31 | 0.38 | 0.18 | 0.17 |
| O(water) | 0.03 | 0.03 | 0.02 | 0.00 | 0.00 |
| Residue | Gold-Bound | Zinc-Bound | |
|---|---|---|---|
| Chain A | Chain B | ||
| His120 | 172.0 | 157.0 | 169.1 |
| His122 | 140.8 | 136.9 | 162.3 |
| Asp124 | 149.2 | 134.2 | 141.7 |
| His189 | 169.7 | 155.0 | 173.5 |
| Cys208 | 127.5 | 131.9 | 117.7 |
| His250 | 225.0 | 214.4 | 243.5 |
| M1 | 0.0 | 0.0 | 0.0 |
| M2 | 0.0 | 0.0 | 0.0 |
| OH | 10.0 | - | 0.0 |
| H2O | 20.1 | 13.4 | 20.1 |
| total SAS | 1004.3 | 942.8 | 1027.9 |
| per atom SAS | 17.6 | 16.3 | 18.4 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tolbatov, I.; Marrone, A. Auranofin Targeting the NDM-1 Beta-Lactamase: Computational Insights into the Electronic Configuration and Quasi-Tetrahedral Coordination of Gold Ions. Pharmaceutics 2023, 15, 985. https://doi.org/10.3390/pharmaceutics15030985
Tolbatov I, Marrone A. Auranofin Targeting the NDM-1 Beta-Lactamase: Computational Insights into the Electronic Configuration and Quasi-Tetrahedral Coordination of Gold Ions. Pharmaceutics. 2023; 15(3):985. https://doi.org/10.3390/pharmaceutics15030985
Chicago/Turabian StyleTolbatov, Iogann, and Alessandro Marrone. 2023. "Auranofin Targeting the NDM-1 Beta-Lactamase: Computational Insights into the Electronic Configuration and Quasi-Tetrahedral Coordination of Gold Ions" Pharmaceutics 15, no. 3: 985. https://doi.org/10.3390/pharmaceutics15030985