
Apoptotic Mechanisms of Quercetin in Liver Cancer: Recent Trends and Advancements
 , ,
, ,  , ,
, ,  , ,
, ,
Abstract
:1. Introduction
2. Chemistry of Quercetin
3. Absorption and Metabolism
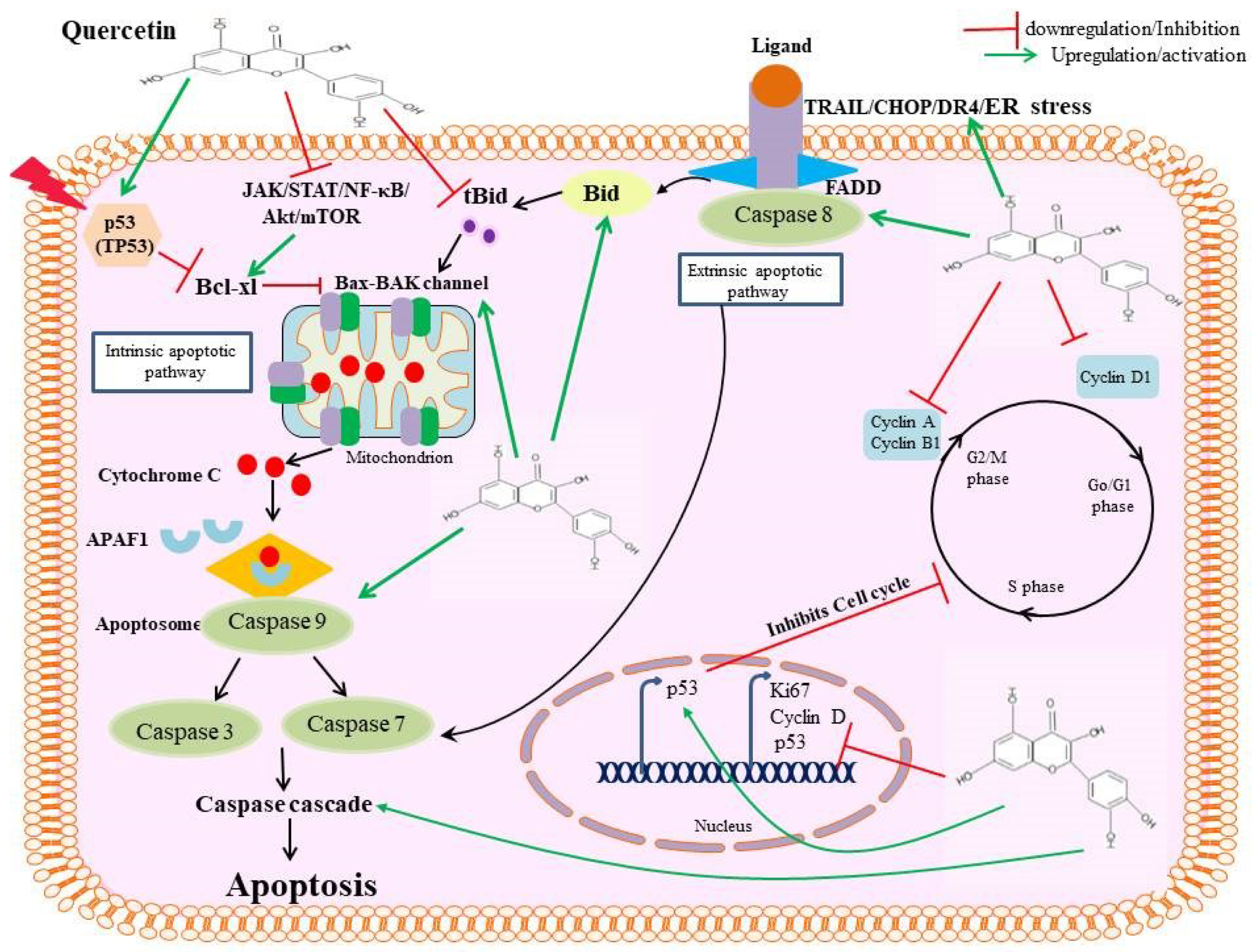
4. Apoptotic Mechanisms of Quercetin
4.1. Activating Caspase Proteases
4.2. Modulating the Bcl2-Bax Pathway
4.3. Targeting the PI3K-Akt-mTOR Pathway
4.4. Targeting JAK-STAT3 Signal Pathway
4.5. Inducing Apoptosis via Autophagy Modulation
5. Synergism of Quercetin in the Liver Cancer
6. Nano Delivery of Quercetin in Liver Cancer
7. Safety Aspects
8. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Agnihotri, A. Liver Cancer Deaths are Expected to Rise by More Than 55% by 2040: Research. Health-Hindustan Times, 10 October 2022. [Google Scholar]
- Baskar, R.; Lee, K.A.; Yeo, R.; Yeoh, K.W. Cancer and radiation therapy: Current advances and future directions. Int. J. Med. Sci. 2012, 9, 193–199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swamy, S.G.; Kameshwar, V.H.; Shubha, P.B.; Looi, C.Y.; Shanmugam, M.K.; Arfuso, F.; Dharmarajan, A.; Sethi, G.; Shivananju, N.S.; Bishayee, A. Targeting multiple oncogenic pathways for the treatment of hepatocellular carcinoma. Target. Oncol. 2017, 12, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Kirtonia, A.; Gala, K.; Fernandes, S.G.; Pandya, G.; Pandey, A.K.; Sethi, G.; Khattar, E.; Garg, M. Repurposing of drugs: An attractive pharmacological strategy for cancer therapeutics. Semin. Cancer Biol. 2021, 68, 258–278. [Google Scholar] [CrossRef]
- Lock, M.I.; Klein, J.; Chung, H.T.; Herman, J.M.; Kim, E.Y.; Small, W.; Mayr, N.A.; Lo, S.S. Strategies to tackle the challenges of external beam radiotherapy for liver tumors. World J. Hepatol. 2017, 9, 645–656. [Google Scholar] [CrossRef]
- Wu, H.; Xie, J.; Pan, Q.; Wang, B.; Hu, D.; Hu, X. Anticancer Agent Shikonin Is an Incompetent Inducer of Cancer Drug Resistance. PLoS ONE 2013, 8, e52706. [Google Scholar] [CrossRef] [PubMed]
- Raghunath, A.; Sundarraj, K.; Arfuso, F.; Sethi, G.; Perumal, E. Dysregulation of Nrf2 in Hepatocellular Carcinoma: Role in Cancer Progression and Chemoresistance. Cancers 2018, 10, 481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nair, A.S.; Shishodia, S.; Ahn, K.S.; Kunnumakkara, A.B.; Sethi, G.; Aggarwal, B.B. Deguelin, an Akt inhibitor, suppresses IkappaBα kinase activation leading to suppression of NF-κB-regulated gene expression, potentiation of apoptosis, and inhibition of cellular invasion. J. Immunol. 2006, 177, 5612–5622. [Google Scholar] [CrossRef] [Green Version]
- Ho, M.-C.; Hasegawa, K.; Chen, X.-P.; Nagano, H.; Lee, Y.-J.; Chau, G.-Y.; Zhou, J.; Wang, C.-C.; Choi, Y.R.; Poon, R.T.-P.; et al. Surgery for Intermediate and Advanced Hepatocellular Carcinoma: A Consensus Report from the 5th Asia-Pacific Primary Liver Cancer Expert Meeting (APPLE 2014). Liver Cancer 2016, 5, 245–256. [Google Scholar] [CrossRef]
- Taefehshokr, S.; Parhizkar, A.; Hayati, S.; Mousapour, M.; Mahmoudpour, A.; Eleid, L.; Rahmanpour, D.; Fattahi, S.; Shabani, H.; Taefehshokr, N. Cancer immunotherapy: Challenges and limitations. Pathol. Res. Pract. 2022, 229, 153723. [Google Scholar] [CrossRef]
- Paskeh, M.D.A.; Ghadyani, F.; Hashemi, M.; Abbaspour, A.; Zabolian, A.; Javanshir, S.; Razzazan, M.; Mirzaei, S.; Entezari, M.; Goharrizi, M.A.S.B.; et al. Biological impact and therapeutic perspective of targeting PI3K/Akt signaling in hepatocellular carcinoma: Promises and Challenges. Pharmacol. Res. 2022, 187, 106553. [Google Scholar] [CrossRef]
- Mirzaei, S.; Saghari, S.; Bassiri, F.; Raesi, R.; Zarrabi, A.; Hushmandi, K.; Sethi, G.; Tergaonkar, V. NF-κB as a regulator of cancer metastasis and therapy response: A focus on epithelial-mesenchymal transition. J. Cell. Physiol. 2022, 237, 2770–2795. [Google Scholar] [CrossRef]
- Sewell, R.D.E.; Rafieian-Kopaei, M. The history and ups and downs of herbal medicines usage. J. Herbmed Pharmacol. 2014, 3, 1–3. [Google Scholar]
- Hsieh, Y.S.; Yang, S.F.; Sethi, G.; Hu, D.N. Natural bioactives in cancer treatment and prevention. Biomed Res. Int. 2015, 2015, 182835. [Google Scholar] [CrossRef] [Green Version]
- Mastron, J.K.; Siveen, K.S.; Sethi, G.; Bishayee, A. Silymarin and hepatocellular carcinoma: A systematic, comprehensive, and critical review. Anticancer Drugs 2015, 26, 475–486. [Google Scholar] [CrossRef] [Green Version]
- Bahmani, M.; Shirzad, H.; Shahinfard, N.; Sheivandi, L.; Rafieian-Kopaei, M. Cancer Phytotherapy: Recent Views on the Role of Antioxidant and Angiogenesis Activities. J. Evid. Based. Complement. Altern. Med. 2017, 22, 299–309. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.F.; Weng, C.J.; Sethi, G.; Hu, D.N. Natural bioactives and phytochemicals serve in cancer treatment and prevention. Evid. Based. Complement. Alternat. Med. 2013, 2013, 698190. [Google Scholar] [CrossRef] [Green Version]
- Dai, X.; Ahn, K.S.; Kim, C.; Siveen, K.S.; Ong, T.H.; Shanmugam, M.K.; Li, F.; Shi, J.; Kumar, A.P.; Wang, L.Z.; et al. Ascochlorin, an isoprenoid antibiotic inhibits growth and invasion of hepatocellular carcinoma by targeting STAT3 signaling cascade through the induction of PIAS3. Mol. Oncol. 2015, 9, 818–833. [Google Scholar] [CrossRef] [Green Version]
- El-Tablawy, N.H.; Mansour, H.A.; Shaaban, A.E.S.M. Antioxidant activities of some edaphic algae in Egypt. Beni-Suef Univ. J. Basic Appl. Sci. 2020, 9, 1–11. [Google Scholar] [CrossRef]
- Alghazeer, R.; Howell, N.K.; El-Naili, M.B.; Awayn, N. Anticancer and Antioxidant Activities of Some Algae from Western Libyan Coast. Nat. Sci. 2018, 10, 232–246. [Google Scholar] [CrossRef] [Green Version]
- Newman, D.J.; Cragg, G.M. Natural Products as Sources of New Drugs from 1981 to 2014. J. Nat. Prod. 2016, 79, 629–661. [Google Scholar] [CrossRef] [Green Version]
- Vargas, A.J.; Burd, R. Hormesis and synergy: Pathways and mechanisms of quercetin in cancer prevention and management. Nutr. Rev. 2010, 68, 418–428. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Yao, J.; Han, C.; Yang, J.; Chaudhry, M.T.; Wang, S.; Liu, H.; Yin, Y. Quercetin, Inflammation and Immunity. Nutrients 2016, 8, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernández-Palanca, P.; Fondevila, F.; Méndez-Blanco, C.; Tuñón, M.J.; González-Gallego, J.; Mauriz, J.L. Antitumor Effects of Quercetin in Hepatocarcinoma In Vitro and In Vivo Models: A Systematic Review. Nutrients 2019, 11, 2875. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Granado-Serrano, A.B.; Martn, M.Á.; Bravo, L.; Goya, L.; Ramos, S. Quercetin Attenuates TNF-Induced Inflammation in Hepatic Cells by Inhibiting the NF-κB Pathway. Nutr. Cancer 2012, 64, 588–598. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, G.N.; Jang, H.D. Protective Mechanism of Quercetin and Rutin Using Glutathione Metabolism on H2O2-induced Oxidative Stress in HepG2 Cells. Ann. N. Y. Acad. Sci. 2009, 1171, 530–537. [Google Scholar] [CrossRef]
- Casella, M.L.; Parody, J.P.; Ceballos, M.P.; Quiroga, A.D.; Ronco, M.T.; Francés, D.E.; Monti, J.A.; Pisani, G.B.; Carnovale, C.E.; Carrillo, M.C.; et al. Quercetin prevents liver carcinogenesis by inducing cell cycle arrest, decreasing cell proliferation and enhancing apoptosis. Mol. Nutr. Food Res. 2014, 58, 289–300. [Google Scholar] [CrossRef] [Green Version]
- Dai, W.; Gao, Q.; Qiu, J.; Yuan, J.; Wu, G.; Shen, G. Quercetin induces apoptosis and enhances 5-FU therapeutic efficacy in hepatocellular carcinoma. Tumor. Biol. 2016, 37, 6307–6313. [Google Scholar] [CrossRef]
- Salama, Y.A.; El-karef, A.; El Gayyar, A.M.; Abdel-Rahman, N. Beyond its antioxidant properties: Quercetin targets multiple signalling pathways in hepatocellular carcinoma in rats. Life Sci. 2019, 236, 116933. [Google Scholar] [CrossRef]
- Granado-Serrano, A.B.; Martín, M.A.; Bravo, L.; Goya, L.; Ramos, S. Quercetin induces apoptosis via caspase activation, regulation of Bcl-2, and inhibition of PI-3-kinase/Akt and ERK pathways in a human hepatoma cell line (HepG2). J. Nutr. 2006, 136, 2715–2721. [Google Scholar] [CrossRef] [Green Version]
- Brito, A.F.; Ribeiro, M.; Abrantes, A.M.; Mamede, A.C.; Laranjo, M.; Casalta-Lopes, J.E.; Gonçalves, A.C.; Sarmento-Ribeiro, A.B.; Tralhão, J.G.; Botelho, M.F. New Approach for Treatment of Primary Liver Tumors: The Role of Quercetin. Nutr. Cancer 2016, 68, 250–266. [Google Scholar] [CrossRef]
- Wu, L.; Li, J.; Liu, T.; Li, S.; Feng, J.; Yu, Q.; Zhang, J.; Chen, J.; Zhou, Y.; Ji, J.; et al. Quercetin shows anti-tumor effect in hepatocellular carcinoma LM3 cells by abrogating JAK2/STAT3 signaling pathway. Cancer Med. 2019, 8, 4806–4820. [Google Scholar] [CrossRef]
- Reyes-Avendaño, I.; Reyes-Jiménez, E.; González-García, K.; Pérez-Figueroa, D.C.; Baltiérrez-Hoyos, R.; Tapia-Pastrana, G.; Sánchez-Chino, X.M.; Villa-Treviño, S.; Arellanes-Robledo, J.; Vásquez-Garzón, V.R. Quercetin Regulates Key Components of the Cellular Microenvironment during Early Hepatocarcinogenesis. Antioxidants 2022, 11, 358. [Google Scholar] [CrossRef]
- Bentz, A.B. A Review of Quercetin: Chemistry, Antioxident Properties, and Bioavailability. J. Young Investig. 2009. Available online: https://www.jyi.org/2009-april/2017/10/15/a-review-of-quercetin-chemistry-antioxidant-properties-and-bioavailability# (accessed on 30 December 2022).
- Bhaskarachary, K.; Joshi, A.K.R. Natural Bioactive Molecules with Antidiabetic Attributes: Insights into Structure–Activity Relationships. Stud. Nat. Prod. Chem. 2018, 57, 353–388. [Google Scholar] [CrossRef]
- Rossi, M.; Rickles, L.F.; Halpin, W.A. The crystal and molecular structure of quercetin: A biologically active and naturally occurring flavonoid. Bioorg. Chem. 1986, 14, 55–69. [Google Scholar] [CrossRef]
- Simanjuntak, K.; Simanjuntak, J.E.; Rosmalena; Prasasty, V.D. Structure-based drug design of quercetin and its derivatives against HMGB1. Biomed. Pharmacol. J. 2017, 10, 1973–1982. [Google Scholar] [CrossRef]
- Röder, P.V.; Geillinger, K.E.; Zietek, T.S.; Thorens, B.; Koepsell, H.; Daniel, H. The Role of SGLT1 and GLUT2 in Intestinal Glucose Transport and Sensing. PLoS ONE 2014, 9, e89977. [Google Scholar] [CrossRef]
- Hai, Y.; Zhang, Y.; Liang, Y.; Ma, X.; Qi, X.; Xiao, J.; Xue, W.; Luo, Y.; Yue, T. Advance on the absorption, metabolism, and efficacy exertion of quercetin and its important derivatives. Food Front. 2020, 1, 420–434. [Google Scholar] [CrossRef]
- Murota, K.; Terao, J. Antioxidative flavonoid quercetin: Implication of its intestinal absorption and metabolism. Arch. Biochem. Biophys. 2003, 417, 12–17. [Google Scholar] [CrossRef]
- Ulusoy, H.G.; Sanlier, N. A minireview of quercetin: From its metabolism to possible mechanisms of its biological activities. Crit. Rev. Food Sci. Nutr. 2019, 60, 3290–3303. [Google Scholar] [CrossRef]
- Almeida, A.F.; Borge, G.I.A.; Piskula, M.; Tudose, A.; Tudoreanu, L.; Valentová, K.; Williamson, G.; Santos, C.N. Bioavailability of Quercetin in Humans with a Focus on Interindividual Variation. Compr. Rev. Food Sci. Food Saf. 2018, 17, 714–731. [Google Scholar] [CrossRef] [Green Version]
- Ishizawa, K.; Yoshizumi, M.; Kawai, Y.; Terao, J.; Kihira, Y.; Ikeda, Y.; Tomita, S.; Minakuchi, K.; Tsuchiya, K.; Tamaki, T. Pharmacology in Health Food: Metabolism of Quercetin In Vivo and Its Protective Effect Against Arteriosclerosis. J. Pharmacol. Sci. 2011, 115, 466–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, W.; Sun, C.; Mao, L.; Ma, P.; Liu, F.; Yang, J.; Gao, Y. The biological activities, chemical stability, metabolism and delivery systems of quercetin: A review. Trends Food Sci. Technol. 2016, 56, 21–38. [Google Scholar] [CrossRef]
- Asadi, M.; Taghizadeh, S.; Kaviani, E.; Vakili, O.; Taheri-Anganeh, M.; Tahamtan, M.; Savardashtaki, A. Caspase-3: Structure, function, and biotechnological aspects. Biotechnol. Appl. Biochem. 2022, 69, 1633–1645. [Google Scholar] [CrossRef] [PubMed]
- Ha, E.J.; Kim, K.Y.; Kim, C.E.; Jun, D.Y.; Kim, Y.H. Enhancement of Quercetin-Induced Apoptosis by Cotreatment with Autophagy Inhibitor Is Associated with Augmentation of BAK-Dependent Mitochondrial Pathway in Jurkat T Cells. Oxid. Med. Cell. Longev. 2019, 2019, 7989276. [Google Scholar] [CrossRef] [Green Version]
- Tripathi, D.; Gupta, P.K.; Banerjee, S.; Kulkarni, S. Quercetin induces proteolysis of mesenchymal marker vimentin through activation of caspase-3, and decreases cancer stem cell population in human papillary thyroid cancer cell line. Phytomed. Plus 2021, 1, 100108. [Google Scholar] [CrossRef]
- Ji, Y.; Li, L.; Ma, Y.X.; Li, W.T.; Li, L.; Zhu, H.Z.; Wu, M.H.; Zhou, J.R. Quercetin inhibits growth of hepatocellular carcinoma by apoptosis induction in part via autophagy stimulation in mice. J. Nutr. Biochem. 2019, 69, 108–119. [Google Scholar] [CrossRef]
- Ren, K.W.; Li, Y.H.; Wu, G.; Ren, J.Z.; Lu, H.; Bin; Li, Z.M.; Han, X.W. Quercetin nanoparticles display antitumor activity via proliferation inhibition and apoptosis induction in liver cancer cells. Int. J. Oncol. 2017, 50, 1299–1311. [Google Scholar] [CrossRef] [Green Version]
- Abdelmoneem, M.A.; Elnaggar, M.A.; Hammady, R.S.; Kamel, S.M.; Helmy, M.W.; Abdulkader, M.A.; Zaky, A.; Fang, J.Y.; Elkhodairy, K.A.; Elzoghby, A.O. Dual-Targeted Lactoferrin Shell-Oily Core Nanocapsules for Synergistic Targeted/Herbal Therapy of Hepatocellular Carcinoma. ACS Appl. Mater. Interfaces 2019, 11, 26731–26744. [Google Scholar] [CrossRef]
- Wang, G.; Zhang, J.; Liu, L.; Sharma, S.; Dong, Q. Quercetin Potentiates Doxorubicin Mediated Antitumor Effects against Liver Cancer through p53/Bcl-xl. PLoS ONE 2012, 7, e51764. [Google Scholar] [CrossRef] [Green Version]
- Michels, G.; Wätjen, W.; Niering, P.; Steffan, B.; Thi, Q.H.T.; Chovolou, Y.; Kampkötter, A.; Bast, A.; Proksch, P.; Kahl, R. Pro-apoptotic effects of the flavonoid luteolin in rat H4IIE cells. Toxicology 2005, 206, 337–348. [Google Scholar] [CrossRef]
- Jin, Y.K.; Eun, H.K.; Seok, S.P.; Jun, H.L.; Taeg, K.K.; Kyeong, S.C. Quercetin sensitizes human hepatoma cells to TRAIL-induced apoptosis via Sp1-mediated DR5 up-regulation and proteasome-mediated c-FLIPS down-regulation. J. Cell. Biochem. 2008, 105, 1386–1398. [Google Scholar] [CrossRef]
- Zhang, Z.; Bai, L.; Hou, L.; Deng, H.; Luan, S.; Liu, D.; Huang, M.; Zhao, L. Trends in targeting Bcl-2 anti-apoptotic proteins for cancer treatment. Eur. J. Med. Chem. 2022, 232, 114184. [Google Scholar] [CrossRef]
- Tan, J.; Wang, B.; Zhu, L. Regulation of Survivin and Bcl-2 in HepG2 Cell Apoptosis Induced by Quercetin. Chem. Biodivers. 2009, 6, 1101–1110. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, H.; Wang, Y.; Song, F.; Yuan, Y. Inhibitory effects of quercetin on the progression of liver fibrosis through the regulation of NF-κB/IκBα, p38 MAPK, and Bcl-2/Bax signaling. Int. Immunopharmacol. 2017, 47, 126–133. [Google Scholar] [CrossRef]
- Karim, S.; Burzangi, A.S.; Ahmad, A.; Siddiqui, N.A.; Ibrahim, I.M.; Sharma, P.; Abualsunun, W.A.; Gabr, G.A. PI3K-AKT Pathway Modulation by Thymoquinone Limits Tumor Growth and Glycolytic Metabolism in Colorectal Cancer. Int. J. Mol. Sci. 2022, 23, 2305. [Google Scholar] [CrossRef]
- Wu, H.; Pan, L.; Gao, C.; Xu, H.; Li, Y.; Zhang, L.; Ma, L.; Meng, L.; Sun, X.; Qin, H. Quercetin Inhibits the Proliferation of Glycolysis-Addicted HCC Cells by Reducing Hexokinase 2 and Akt-mTOR Pathway. Molecules 2019, 24, 1993. [Google Scholar] [CrossRef] [Green Version]
- Maurya, A.K.; Vinayak, M. Anticarcinogenic action of quercetin by downregulation of phosphatidylinositol 3-kinase (PI3K) and protein kinase C (PKC) via induction of p53 in hepatocellular carcinoma (HepG2) cell line. Mol. Biol. Rep. 2015, 42, 1419–1429. [Google Scholar] [CrossRef]
- Zhao, X.; Wang, J.; Deng, Y.; Liao, L.; Zhou, M.; Peng, C.; Li, Y. Quercetin as a protective agent for liver diseases: A comprehensive descriptive review of the molecular mechanism. Phyther. Res. 2021, 35, 4727–4747. [Google Scholar] [CrossRef]
- Yang, H.; Xu, S.; Tang, L.; Gong, J.; Fang, H.; Wei, J.; Su, D. Targeting of non-apoptotic cancer cell death mechanisms by quercetin: Implications in cancer therapy. Front. Pharmacol. 2022, 13, 4938. [Google Scholar] [CrossRef]
- Ding, Y.; Chen, X.; Wang, B.; Yu, B.; Ge, J.; Shi, X. Quercetin suppresses the chymotrypsin-like activity of proteasome via inhibition of MEK1/ERK1/2 signaling pathway in hepatocellular carcinoma HepG2 cells. Can. J. Physiol. Pharmacol. 2018, 96, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Chan, J.M.; Zaidi, S.; Love, J.R.; Zhao, J.L.; Setty, M.; Wadosky, K.M.; Gopalan, A.; Choo, Z.N.; Persad, S.; Choi, J.; et al. Lineage plasticity in prostate cancer depends on JAK/STAT inflammatory signaling. Science 2022, 377, 1180–1191. [Google Scholar] [CrossRef] [PubMed]
- Igbe, I.; Shen, X.F.; Jiao, W.; Qiang, Z.; Deng, T.; Li, S.; Liu, W.L.; Liu, H.W.; Zhang, G.L.; Wang, F. Dietary quercetin potentiates the antiproliferative effect of interferon-α in hepatocellular carcinoma cells through activation of JAK/STAT pathway signaling by inhibition of SHP2 phosphatase. Oncotarget 2017, 8, 113734. [Google Scholar] [CrossRef]
- Fan, H.; Tang, H.B.; Shan, L.Q.; Liu, S.C.; Huang, D.G.; Chen, X.; Chen, Z.; Yang, M.; Yin, X.H.; Yang, H.; et al. Quercetin prevents necroptosis of oligodendrocytes by inhibiting macrophages/microglia polarization to M1 phenotype after spinal cord injury in rats. J. Neuroinflamm. 2019, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Seo, H.S.; Ku, J.M.; Choi, H.S.; Choi, Y.K.; Woo, J.K.; Kim, M.; Kim, I.; Na, C.H.; Hur, H.; Jang, B.H.; et al. Quercetin induces caspase-dependent extrinsic apoptosis through inhibition of signal transducer and activator of transcription 3 signaling in HER2-overexpressing BT-474 breast cancer cells. Oncol. Rep. 2016, 36, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zalpoor, H.; Nabi-Afjadi, M.; Forghaniesfidvajani, R.; Tavakol, C.; Farahighasreaboonasr, F.; Pakizeh, F.; Dana, V.G.; Seif, F. Quercetin as a JAK–STAT inhibitor: A potential role in solid tumors and neurodegenerative diseases. Cell. Mol. Biol. Lett. 2022, 27, 1–17. [Google Scholar] [CrossRef]
- Zhao, J.L.; Zhao, J.; Jiao, H.J. Synergistic growth-suppressive effects of quercetin and cisplatin on HepG2 human hepatocellular carcinoma cells. Appl. Biochem. Biotechnol. 2014, 172, 784–791. [Google Scholar] [CrossRef]
- Gao, W.; Wang, X.; Zhou, Y.; Wang, X.; Yu, Y. Autophagy, ferroptosis, pyroptosis, and necroptosis in tumor immunotherapy. Signal Transduct. Target. Ther. 2022, 7, 196. [Google Scholar] [CrossRef]
- Ashrafizadeh, M.; Ahmadi, Z.; Farkhondeh, T.; Samarghandian, S. Autophagy as a molecular target of quercetin underlying its protective effects in human diseases. J. Metab. Dis. 2019, 128, 200–208. [Google Scholar] [CrossRef]
- Patra, S.; Pradhan, B.; Nayak, R.; Behera, C.; Panda, K.C.; Das, S.; Jena, M.; Bhutia, S.K. Apoptosis and autophagy modulating dietary phytochemicals in cancer therapeutics: Current evidences and future perspectives. Phyther. Res. 2021, 35, 4194–4214. [Google Scholar] [CrossRef]
- Wang, K.; Liu, R.; Li, J.; Mao, J.; Lei, Y.; Wu, J.; Zeng, J.; Zhang, T.; Wu, H.; Chen, L.; et al. Quercetin induces protective autophagy in gastric cancer cells: Involvement of Akt-mTOR- and hypoxia-induced factor 1α-mediated signaling. Autophagy 2011, 7, 966–978. [Google Scholar] [CrossRef] [Green Version]
- Levinson, A.D. Cancer therapy reform. Science 2010, 328, 137. [Google Scholar] [CrossRef] [Green Version]
- Ehrhardt, H.; Häcker, S.; Wittmann, S.; Maurer, M.; Borkhardt, A.; Toloczko, A.; Debatin, K.M.; Fulda, S.; Jeremias, I. Cytotoxic drug-induced, p53-mediated upregulation of caspase-8 in tumor cells. Oncogene 2007, 27, 783–793. [Google Scholar] [CrossRef] [Green Version]
- Ahn, K.S.; Sethi, G.; Aggarwal, B.B. Reversal of chemoresistance and enhancement of apoptosis by statins through down-regulation of the NF-κB pathway. Biochem. Pharmacol. 2008, 75, 907–913. [Google Scholar] [CrossRef] [Green Version]
- Tan, S.M.; Li, F.; Rajendran, P.; Kumar, A.P.; Hui, K.M.; Sethi, G. Identification of beta-escin as a novel inhibitor of signal transducer and activator of transcription 3/Janus-activated kinase 2 signaling pathway that suppresses proliferation and induces apoptosis in human hepatocellular carcinoma cells. J. Pharmacol. Exp. Ther. 2010, 334, 285–293. [Google Scholar] [CrossRef]
- Hisaka, T.; Sakai, H.; Sato, T.; Goto, Y.; Nomura, Y.; Fukutomi, S.; Fujita, F.; Mizobe, T.; Nakashima, O.; Tanigawa, M.; et al. Quercetin Suppresses Proliferation of Liver Cancer Cell Lines In Vitro. Anticancer Res. 2020, 40, 4695–4700. [Google Scholar] [CrossRef]
- Abdu, S.; Juaid, N.; Amin, A.; Moulay, M.; Miled, N. Effects of Sorafenib and Quercetin Alone or in Combination in Treating Hepatocellular Carcinoma: In Vitro and In Vivo Approaches. Molecules 2022, 27, 8082. [Google Scholar] [CrossRef]
- Zou, H.; Zheng, Y.F.; Ge, W.; Wang, S.B.; Mou, X.Z. Synergistic Anti-tumour Effects of Quercetin and Oncolytic Adenovirus expressing TRAIL in Human Hepatocellular Carcinoma. Sci. Rep. 2018, 8, 1–8. [Google Scholar] [CrossRef] [Green Version]
- Aggarwal, V.; Tuli, H.S.; Thakral, F.; Singhal, P.; Aggarwal, D.; Srivastava, S.; Pandey, A.; Sak, K.; Varol, M.; Khan, M.A.; et al. Molecular mechanisms of action of hesperidin in cancer: Recent trends and advancements. Exp. Biol. Med. 2020, 245, 486–497. [Google Scholar] [CrossRef] [Green Version]
- Dehshahri, A.; Ashrafizadeh, M.; Ghasemipour Afshar, E.; Pardakhty, A.; Mandegary, A.; Mohammadinejad, R.; Sethi, G. Topoisomerase inhibitors: Pharmacology and emerging nanoscale delivery systems. Pharmacol. Res. 2020, 151, 104551. [Google Scholar] [CrossRef]
- Vinayak, M.; Maurya, A.K. Quercetin Loaded Nanoparticles in Targeting Cancer: Recent Development. Anticancer Agents Med. Chem. 2019, 19, 1560–1576. [Google Scholar] [CrossRef] [PubMed]
- Davatgaran-Taghipour, Y.; Masoomzadeh, S.; Farzaei, M.H.; Bahramsoltani, R.; Karimi-Soureh, Z.; Rahimi, R.; Abdollahi, M. Polyphenol nanoformulations for cancer therapy: Experimental evidence and clinical perspective. Int. J. Nanomed. 2017, 12, 2689–2702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, T.; Singh, D.; Mahapatra, A.; Mohapatra, P.; Sahoo, S.; Sahoo, S.K. Advancements in clinical translation of flavonoid nanoparticles for cancer treatment. OpenNano 2022, 8, 100074. [Google Scholar] [CrossRef]
- Yadav, N.; Tripathi, A.K.; Parveen, A. PLGA-Quercetin Nano-Formulation Inhibits Cancer Progression via Mitochondrial Dependent Caspase-3,7 and Independent FoxO1 Activation with Concomitant PI3K/AKT Suppression. Pharmaceutics 2022, 14, 1326. [Google Scholar] [CrossRef]
- Ersoz, M.; Erdemir, A.; Derman, S.; Arasoglu, T.; Mansuroglu, B. Quercetin-loaded nanoparticles enhance cytotoxicity and antioxidant activity on C6 glioma cells. Pharm. Dev. Technol. 2020, 25, 757–766. [Google Scholar] [CrossRef]
- Sadhukhan, P.; Kundu, M.; Chatterjee, S.; Ghosh, N.; Manna, P.; Das, J.; Sil, P.C. Targeted delivery of quercetin via pH-responsive zinc oxide nanoparticles for breast cancer therapy. Mater. Sci. Eng. 2019, 100, 129–140. [Google Scholar] [CrossRef]
- Baksi, R.; Singh, D.P.; Borse, S.P.; Rana, R.; Sharma, V.; Nivsarkar, M. In vitro and in vivo anticancer efficacy potential of Quercetin loaded polymeric nanoparticles. Biomed. Pharmacother. 2018, 106, 1513–1526. [Google Scholar] [CrossRef]
- Patra, A.; Satpathy, S.; Shenoy, A.K.; Bush, J.A.; Kazi, M.; Hussain, M.D. Formulation and evaluation of mixed polymeric micelles of quercetin for treatment of breast, ovarian, and multidrug resistant cancers. Int. J. Nanomed. 2018, 13, 2869–2881. [Google Scholar] [CrossRef] [Green Version]
- Luo, C.L.; Liu, Y.Q.; Wang, P.; Song, C.H.; Wang, K.J.; Dai, L.P.; Zhang, J.Y.; Ye, H. The effect of quercetin nanoparticle on cervical cancer progression by inducing apoptosis, autophagy and anti-proliferation via JAK2 suppression. Biomed. Pharmacother. 2016, 82, 595–605. [Google Scholar] [CrossRef]
- Andres, S.; Pevny, S.; Ziegenhagen, R.; Bakhiya, N.; Schäfer, B.; Hirsch-Ernst, K.I.; Lampen, A. Safety Aspects of the Use of Quercetin as a Dietary Supplement. Mol. Nutr. Food Res. 2018, 62, 1700447. [Google Scholar] [CrossRef]
- Han, M.K.; Barreto, T.A.; Martinez, F.J.; Comstock, A.T.; Sajjan, U.S. Randomised clinical trial to determine the safety of quercetin supplementation in patients with chronic obstructive pulmonary disease. BMJ Open Respir. Res. 2020, 7, 1700447. [Google Scholar] [CrossRef] [Green Version]
- Lu, N.T.; Crespi, C.M.; Liu, N.M.; Vu, J.Q.; Ahmadieh, Y.; Wu, S.; Lin, S.; McClune, A.; Durazo, F.; Saab, S.; et al. A Phase I Dose Escalation Study Demonstrates Quercetin Safety and Explores Potential for Bioflavonoid Antivirals in Patients with Chronic Hepatitis C. Phytother. Res. 2016, 30, 160–168. [Google Scholar] [CrossRef] [Green Version]
- Marsico, M.; Santarsiero, A.; Pappalardo, I.; Convertini, P.; Chiummiento, L.; Sardone, A.; Di Noia, M.A.; Infantino, V.; Todisco, S. Mitochondria-Mediated Apoptosis of HCC Cells Triggered by Knockdown of Glutamate Dehydrogenase 1: Perspective for Its Inhibition through Quercetin and Permethylated Anigopreissin A. Biomedicines 2021, 9, 1664. [Google Scholar] [CrossRef]
- Liu, T.; Li, Z.; Tian, F. Quercetin inhibited the proliferation and invasion of hepatoblastoma cells through facilitating SIRT6-medicated FZD4 silence. Hum. Exp. Toxicol. 2021, 40, S96–S107. [Google Scholar] [CrossRef]
- Choudhury, A.; Mukherjee, S. In silico studies on the comparative characterization of the interactions of SARS-CoV-2 spike glycoprotein with ACE-2 receptor homologs and human TLRs. J. Med. Virol. 2020, 92, 2105–2113. [Google Scholar] [CrossRef]
- Shui, L.; Wang, W.; Xie, M.; Ye, B.; Li, X.; Liu, Y.; Zheng, M. Isoquercitrin induces apoptosis and autophagy in hepatocellular carcinoma cells via AMPK/mTOR/p70S6K signaling pathway. Aging 2020, 12, 24318. [Google Scholar] [CrossRef]
- Ajji, P.K.; Walder, K.; Puri, M. Combination of Balsamin and Flavonoids Induce Apoptotic Effects in Liver and Breast Cancer Cells. Front. Pharmacol. 2020, 11, 574496. [Google Scholar] [CrossRef]
- Jeon, J.S.; Kwon, S.; Ban, K.; Kwon Hong, Y.; Ahn, C.; Sung, J.S.; Choi, I. Regulation of the Intracellular ROS Level Is Critical for the Antiproliferative Effect of Quercetin in the Hepatocellular Carcinoma Cell Line HepG2. Nutr. Cancer 2019, 71, 861. [Google Scholar] [CrossRef]
- Lee, R.H.; Cho, J.H.; Jeon, Y.J.; Bang, W.; Cho, J.J.; Choi, N.J.; Seo, K.S.; Shim, J.H.; Chae, J. Il Quercetin Induces Antiproliferative Activity Against Human Hepatocellular Carcinoma (HepG2) Cells by Suppressing Specificity Protein 1 (Sp1). Drug Dev. Res. 2015, 76, 9–16. [Google Scholar] [CrossRef]
- Huang, G.; Tang, B.; Tang, K.; Dong, X.; Deng, J.; Liao, L.; Liao, Z.; Yang, H.; He, S. Isoquercitrin inhibits the progression of liver cancer in vivo and in vitro via the MAPK signalling pathway. Oncol. Rep. 2014, 31, 2377–2384. [Google Scholar] [CrossRef] [Green Version]
- Sudan, S.; Rupasinghe, H.V. Quercetin-3-O-glucoside induces human DNA topoisomerase II inhibition, cell cycle arrest and apoptosis in hepatocellular carcinoma cells. Anticancer Res. 2014, 34, 1691–1699. [Google Scholar] [PubMed]
- Yuan, Z.P.; Chen, L.J.; Fan, L.Y.; Tang, M.H.; Yang, G.L.; Yang, H.S.; Du, X.B.; Wang, G.Q.; Yao, W.X.; Zhao, Q.M.; et al. Liposomal quercetin efficiently suppresses growth of solid tumors in murine models. Clin. Cancer Res. 2006, 12, 3193–3199. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| S No. | Type of Nano Formulations/ Nanoparticles (Quercetin Based) | Type of Cancer | Study Model (Both In Vitro/In Vivo) | Particle Size (Nanometres (nm)) | Effects | Concentration | References |
|---|---|---|---|---|---|---|---|
| 1 | PLGA nanoparticles | Cervical | HeLa | 89.8 nm | ↑ apoptosis ↑ Caspase-3 and 7 | 1, 10, 25 and 50 µM | [85] |
| 2 | PLGA nanoparticles | Breast | MCF-7 | 89.8 nm | ↑ apoptosis frequency, ↑ Mitochondrial damage in Cancer Cells, | 1, 10, 25 and 50 µM | [85] |
| 3 | PLGA nanoparticles | Breast | DMBA-induced mammary adenocarcinoma SD rats | 89.8 nm | ↓ average number of tumors and prolonged the tumor latency period | 128 mg/kg | [85] |
| 4 | Poly (lactic-co-glycolic acid) nanoparticles | Glioma | C6 | Qu1NP-215.2 nm, Qu2NP-282.3, Qu3NPs-584.5 nm | ↓ cancer cell proliferation, ↑ SOD activity, ↑ GSH levels | 1–100 μg/mL | [86] |
| 5 | Phenylboronic acid (PBA) conjugated Zinc oxide nanoparticles (PBA-zno) | Breast | MCF-7 | 40 nm | ↑ apoptotic frequency, ↑ mitochondrial damage, | 5–50 μg/ml | [87] |
| 6 | Polymeric (chitosan) nanoparticles | Lung | A549 | <200 nm | ↑ release, ↑ cytotoxicity to cancer cells | 12.5, 25, 50, 75, 100, 150 and 200 μM | [88] |
| 7 | Polymeric (chitosan) nanoparticles | Breast | MDA MB 468 | <200 nm | ↑ release, ↑ cytotoxicity to cancer cells. | 12.5, 25, 50, 75, 100, 150 and 200 μM | [88] |
| 8 | Polymeric micelles | Ovarian Multidrug resistant Breast | Skov-3 NCI/ADR MCF-7 and MDA-MB-231 | 24.83 to 45.88 nm | ↓ cell viability, ↑ targeted drug release directly into the intracellular environment | 6.25 to 100 μM | [89] |
| 9 | Gold-quercetin into poly (DL-lactide-co-glycolide) nanoparticles | Cervical | Male BALB/c nu/nu nude mice xeno-grafted with Caski cells | -- | ↓ tumor xenograft growth and development, ↑ KI-67, ↑ Bax, ↑ Bad, ↑ Cyto-c, ↑ ↓S6RP | -- | [90] |
| Type of Cancer | Cells | Effects | Mechanistic Insight | Concentration | References |
|---|---|---|---|---|---|
| Hepatocellular | HepG2 | Induces Apoptosis, Cell Cycle arrest | ↓ Proliferation of cancer cells, ↑ cell cycle arrest at S and G1 phase, ↑ necrotic and apoptotic cells, | Quercetin 20 to 220 µM or sorafenib 5 to 40 µM) | [78] |
| Hepatocellular | HepG2 | Induces Apoptosis | ↓ Cell proliferation, ↓ Bcl-2, ↓ mitochondrial mass, ↓ mitochondrial membrane potential, ↑ mitochondrial superoxide anion, ↑Caspases 3/7/9, ↓↑ BAX, | Quercetin and Permethylated Anigopreissin A (PAA) (inhibitors of hGDH1)-1,2,4,8,12 and 16 μM | [94] |
| Hepatoblastoma | HepG2, HuH-6 and HepT1 | Induces Apoptosis, | ↑ SIRT6, upregulation of SIRT6, suppressed cell proliferation and invasion, promoted cell apoptosis, ↓ frizzled 4 (FZD4) transcription, ↓ FZD4 and H3K9ac, ↓ Wnt5a, β-catenin, cyclin D1 and c-Myc | 0, 60 and 120 mM | [95] |
| Hepatocellular | HepG2, Huh-7 (Gemcitabine resistant) | Induces Apoptosis, Cell Cycle arrest | ↓ Proliferation of gemcitabine resistant cancer cells, ↑ apoptosis frequency, ↑ gemcitabine efficacy, ↑ accumulation of HepG2 cells in S phase, ↓ G1 and G2/M phase populations, ↑ p53, ↓ cyclin D1 | Quercetin 0, 10, 25, 50, 100, or 200 μM or 0, 10, 25, 50, 100, and 200 nM Gemcitabine | [95,96] |
| Liver | KIM-1, KYN-1, KYN-2, KYN-3, HAK-1A, HAK-1B, HAK-2, HAK-3, HAK-4, HAK-5, and HAK-6 | Induces Apoptosis, Cell Cycle arrest | ↓ Cell proliferation, ↓ viable cell count, ↑ apoptosis frequency, G0/G1, G2/M and S phase cell cycle arrest | 0–100 μM | [77] |
| Hepatocellular | HepG2 and Huh7 | Induces Apoptosis | ↓ cell viability and colony growth, ↑ apoptotic pathway, ↑ caspases ↑ Bax | 100–500 μM | [97] |
| Hepatocellular | HepG2 | Induces Apoptosis, | ↑ cell viability, ↑ caspase-3 and 8, ↑ loss in cell connections, ↑ cell shrinkage, ↑ cell surface detachment, ↑ cytoplasmic density, ↑ dead cells, ↑ Bax, Bid, Bad, and p53, ↓ Bcl-2 and Bcl-XL, ↑ GRP78 and CHOP | Combination of naringenin, quercetin, and naringin and balsamin IC50 values for Nar, Nir and Qu are 150 mM, 20 mM and 37 mM, respectively, and 25 µg/mL of balsamin | [98] |
| Hepatocellular | SMMC7721 and HepG2 | Induces Apoptosis, Cell Cycle arrest | ↓ Growth of HCC cells, ↑ autophagosomes and autolysosomes, ↑ LC3A/B-II and beclin1, ↓ p62, ↓ phosphorylated AKT, mTOR, p70S6K and 4EBP1, ↑ phosphorylated JNK, ERK1/2 and p38MAPK | IC50’s at 21.0 and 34.0 μM | [48] |
| Hepatocellular | LM3 | Induces Apoptosis, Cell Cycle arrest | ↓ Cancer cell viability, ↑ apoptosis frequency, ↑ cleaved DNA, ↑ cells were arrested in the S and G2/M phases, ↓ G0/G1 phase cells, ↓ vimentin and MMP9, ↓ invasion and migration, ↑ LC3, ↓ p-STAT3 | 0, 20, 40, 60, 80, 100,120, 140, 160, and 200 µmol/L | [32] |
| Hepatocellular | HepG2 | Induces Apoptosis, Cell Cycle arrest | ↓ Proliferation of cancer cells, ↓ intracellular ROS level, ↓cyclin E and SOD1 | -- | [99] |
| Hepatocellular | HepG2 | Induces Apoptosis | ↓ Cell viability, ↑ cell apoptosis, ↓ chymotrypsin-like activity of proteasome, ↑ cleaved caspase-3, ↓ Bcl-2, ↑ p38 MAPK and JNK phosphorylation, ↓ ERK1/2 phosphorylation | 0, 25, 50, and 100 µM | [62] |
| Liver | MHCC97H, Hep3B, HCCLM3 and Bel7402 | Induces Apoptosis, | ↓ Cancer cell proliferation, cell migration and colony formation, ↑ caspases, ↑ cytochrome c, ↓ NF-κB ↓ Akt and ERK1/2, P-27 was expressed highly, ↓ c-Myc, ↓ cyclin-D1, ↓ CDK1, ↓ MMP7, ↓ β-catenin, | Gold-quercetin -poly(DL-lactideco-glycolide) nanoparticles-0,10,20,30,40,50 and 50 µg/mL | [49] |
| Hepatocellular | SMMC-7721, HepG2 and HuH-7 | Induces Apoptosis | ↑ ZD55-TRAIL ↑ caspases and cleaved PARP, ↓ ZD55-TRAIL mediated NF-κB activation, ↑ pro-apoptotic action of ZD55-TRAI, ↓ Bcl-2, ↑ Bax | ZD55-TRAIL adenovirus-1,2,5,10 MOI + Quercetin 5, 10, 25, 50 μM | [79] |
| Hepatocellular | HepG2 and SMCC-7721 | Induces Apoptosis, Cell Cycle arrest | ↓ Cell proliferation, ↑ apoptosis, ↑ Bad and Bax, ↓ Bcl-2 and Survivin, ↑ 5-fluorouracil (5-FU) therapeutic efficacy, ↓ cells in S phases, ↑ cells in the G0/G1 phase | 0.05, 0.1, and 0.15 mmol/L | [28] |
| Hepatocellular | HepG2 | Apoptosis | ↓ Cell growth, ↑ apoptosis, ↑ nuclear condensation and fragmentation, ↓ Sp1 and Sp1 regulatory protein, ↑ p27, p21, ↑ Bax, ↑ caspases and cleaved PARP | 10–100 µM | [100] |
| Liver | HepG2 and Hep3B | Induces Apoptosis, Cell Cycle arrest | ↑ Apoptosis, ↑ caspase-3, -8 and -9, ↓ phosphorylation of ERK and p38MAPK, ↑ phosphorylation JNK, ↓ PKC, entering the S and the G2/M phases gradually decreased, while most cells were blocked in the G1 phase | 0, 100, 200, 400 and 800 µM | [101] |
| Hepatocellular | HepG2 | Induces Apoptosis | ↓ Cell proliferation, blockade of the cell cycle in the S-phase, ↓ DNA topoisomerase II, ↑ DNA fragmentation, ↑ caspase-3, ↑ apoptosis frequency | 1, 10, 50, 100, 150 and 200 μM | [102] |
| Hepatoma | HepG2 | Induces Apoptosis | ↓Cell viability, ↑ ROS generation, ↑ caspase-3 and -9, ↑↓ caspase-8, ↓ Bcl-xL:Bcl-xS ratio, ↑ translocation of Bax to the mitochondrial membrane, ↓ Akt and ERK | 0–100 μM | [30] |
| Hepatoma | H22, LL/2 | Induces Apoptosis | ↓ cell proliferation, ↑ apoptotic cell (sub-G1 cells) | 0,5,10 and 15 μg | [103] |
| Type of Cancer | Animal Models | Mechanisms | Dosage | Duration | References |
|---|---|---|---|---|---|
| Hepatocellular | Chemically induced HCC rat model with injection of Diethylnitrosamine @200 mg/kg | ↓ Liver enzymes—aminotransferase (ALT), ↓ aspartate aminotransferase (AST), ↓ alkaline phosphatase (ALP), ↓ total proteins (TP) and conjugated bilirubin (direct bilirubin), ↓ C-reactive protein (CRP), ↓ interleukin 6 (IL-6), ↓ lactate dehydrogenase (LDH), ↓ PIVKA-II and AFP, ↓ Ki-67 cells, ↓ TNFa, VEGF, p53 and NF-κB expression | Quercetin 50 mg/kg + sorafenib 7.5 mg/kg | 13 weeks | [78] |
| Hepatoblastoma | BALB/c nude mice bearing HepG2 cells (1 × 107) cells | ↓ Tumors grew slower and size, ↓ weighed, ↑ SIRT6, ↓ FZD4 | 10 mg/kg | 28 days | [78] |
| Hepatocellular | BALB/c nude mice bearing SMMC7721 cells (2 × 106) cells | ↑ Autophagosomes and autolysosomes, ↓ AKT/mTOR, ↑ MAPK, ↑cleaved caspase-3, ↑ BAX, ↓ Bcl-2, ↑ LC3A/B, ↓ p62, ↑ necrosis | 60 mg/kg | 10 days | [48] |
| Hepatocellular | Sprague Dawley rats (TAA Induced) | ↓ Caspase-3, ↓↑ caspase-8, ↑ p53, ↓ cyclin D1 and Ki-67 | TAA -200 mg/kg + Quercetin 100 mg/kg | 21 days | [29] |
| Hepatocellular | Nude mice bearing LM3 cells | ↓ Tumor volume, ↑ necrosis, ↑ TUNEL-positive cells, ↓ PCNA, ↑ Bax | 100 mg/kg | 21 days | [32] |
| Hepatocellular | BALB/c nu/nu nude mice bearing MHCC97H cells (1 × 107 cells) | ↓ Tumor volumes, ↓ AP-2β and COX2 levels, ↑ TUNEL levels, ↓ cleaved caspase-9, ↓ cleaved caspase-3, ↓ cytoplasm Cyto-c, ↓ phosphorylated IKKα, IκBα and NF-κB | 30, 40 and 50 mg/kg Quercetin nanoparticles | 35 days | [49] |
| Hepatocellular | BALB/C nude mice bearing HuH-cells | ↓ Tumor volume, ↑ survival rate | ZD55-TRAIL was injected intra-tumorally at 1 × 109 plaque-forming units + Quercetin 150 mg/kg | 49 days | [79] |
| Hepatocellular | BALB/c nude mice | ↑ 5-fluorouracil (5-FU) therapeutic efficacy, ↑ apoptosis, ↓ tumor growth | Quercetin 40 mg/kg and 5-fluorouracil 30 mg/kg | 23 days | [28] |
| Liver | Wistar rats (two-phase model of hepato carcinogenesis) | ↓ Number and volume of preneoplastic lesions, ↑ apoptosis, ↓ proliferative index, ↓ cell percentages ↓ M phase, ↓ cyclin D1, ↓ cyclin A, ↓ cyclin B, ↓ cyclin-dependent kinase 1, ↑ peroxisome proliferator, ↑ caspase-3 activity, ↑ Bax/Bcl-2, ↑ cytosolic cytochrome c | 10 and 20 mg/kg | 42 days | [27] |
| Hepatoma | C57BL/6N mice bearing LL/2 Lewis lung cancer (1 × 106) | ↓ Tumor growth, ↓ tumor volume, ↓ HSP70, ↑ apoptosis rate | Liposomal Quercetin -50 mg/kg | 15 days | [103] |
| Hepatoma | BALB/c mice bearing H22 tumor models (5 × 105) | ↓ Tumor growth, ↓ tumor volume, ↓ HSP70, ↑ apoptosis rate | Liposomal Quercetin -50 mg/kg | 15 days | [103] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sethi, G.; Rath, P.; Chauhan, A.; Ranjan, A.; Choudhary, R.; Ramniwas, S.; Sak, K.; Aggarwal, D.; Rani, I.; Tuli, H.S. Apoptotic Mechanisms of Quercetin in Liver Cancer: Recent Trends and Advancements. Pharmaceutics 2023, 15, 712. https://doi.org/10.3390/pharmaceutics15020712
Sethi G, Rath P, Chauhan A, Ranjan A, Choudhary R, Ramniwas S, Sak K, Aggarwal D, Rani I, Tuli HS. Apoptotic Mechanisms of Quercetin in Liver Cancer: Recent Trends and Advancements. Pharmaceutics. 2023; 15(2):712. https://doi.org/10.3390/pharmaceutics15020712
Chicago/Turabian StyleSethi, Gautam, Prangya Rath, Abhishek Chauhan, Anuj Ranjan, Renuka Choudhary, Seema Ramniwas, Katrin Sak, Diwakar Aggarwal, Isha Rani, and Hardeep Singh Tuli. 2023. "Apoptotic Mechanisms of Quercetin in Liver Cancer: Recent Trends and Advancements" Pharmaceutics 15, no. 2: 712. https://doi.org/10.3390/pharmaceutics15020712