
Effects of Dimerization, Dendrimerization, and Chirality in p-BthTX-I Peptide Analogs on the Antibacterial Activity and Enzymatic Inhibition of the SARS-CoV-2 PLpro Protein

 , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
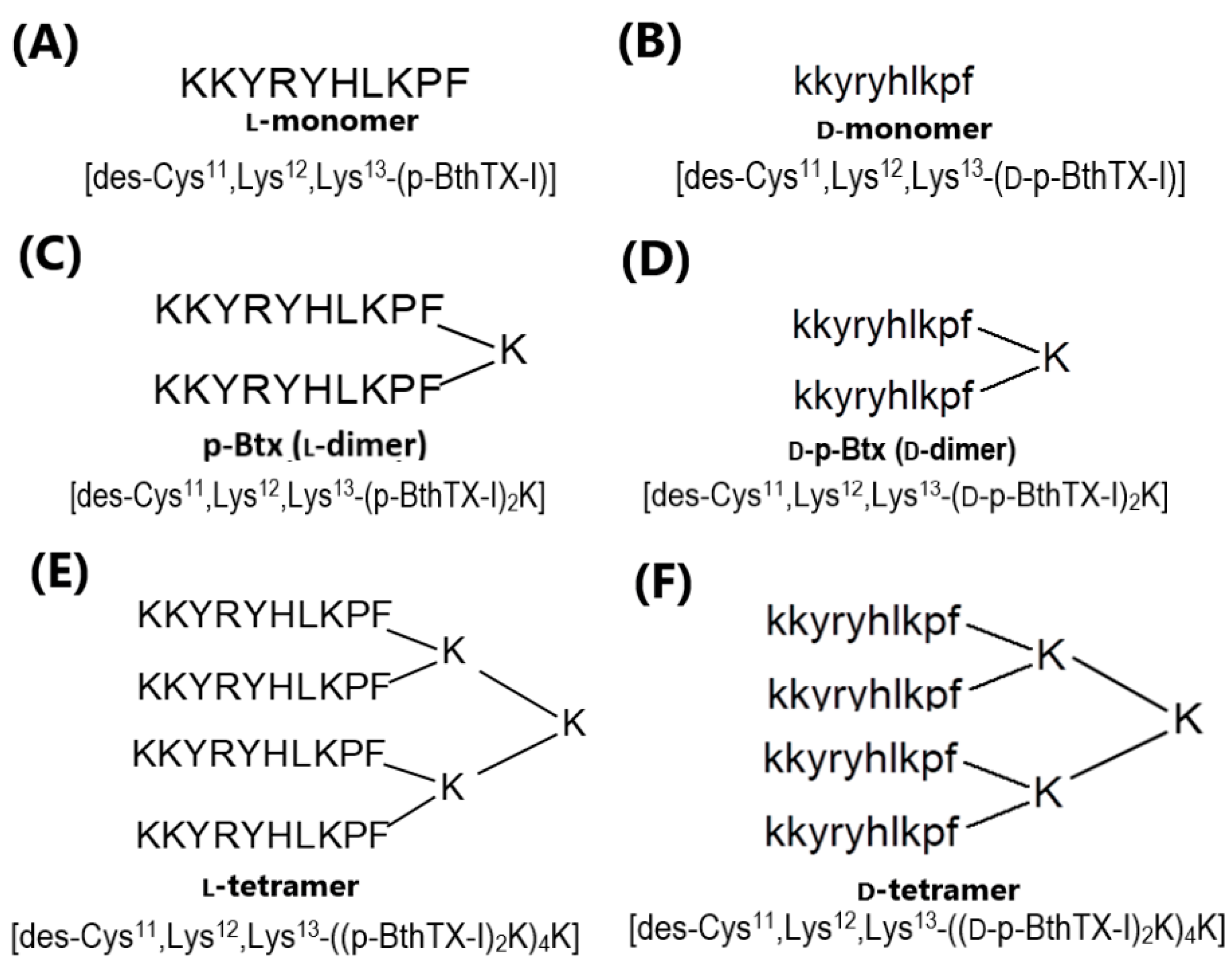
2.1. Peptides Synthesis
2.2. Determination of the Minimum Inhibitory Concentration and Minimum Bactericidal Concentration
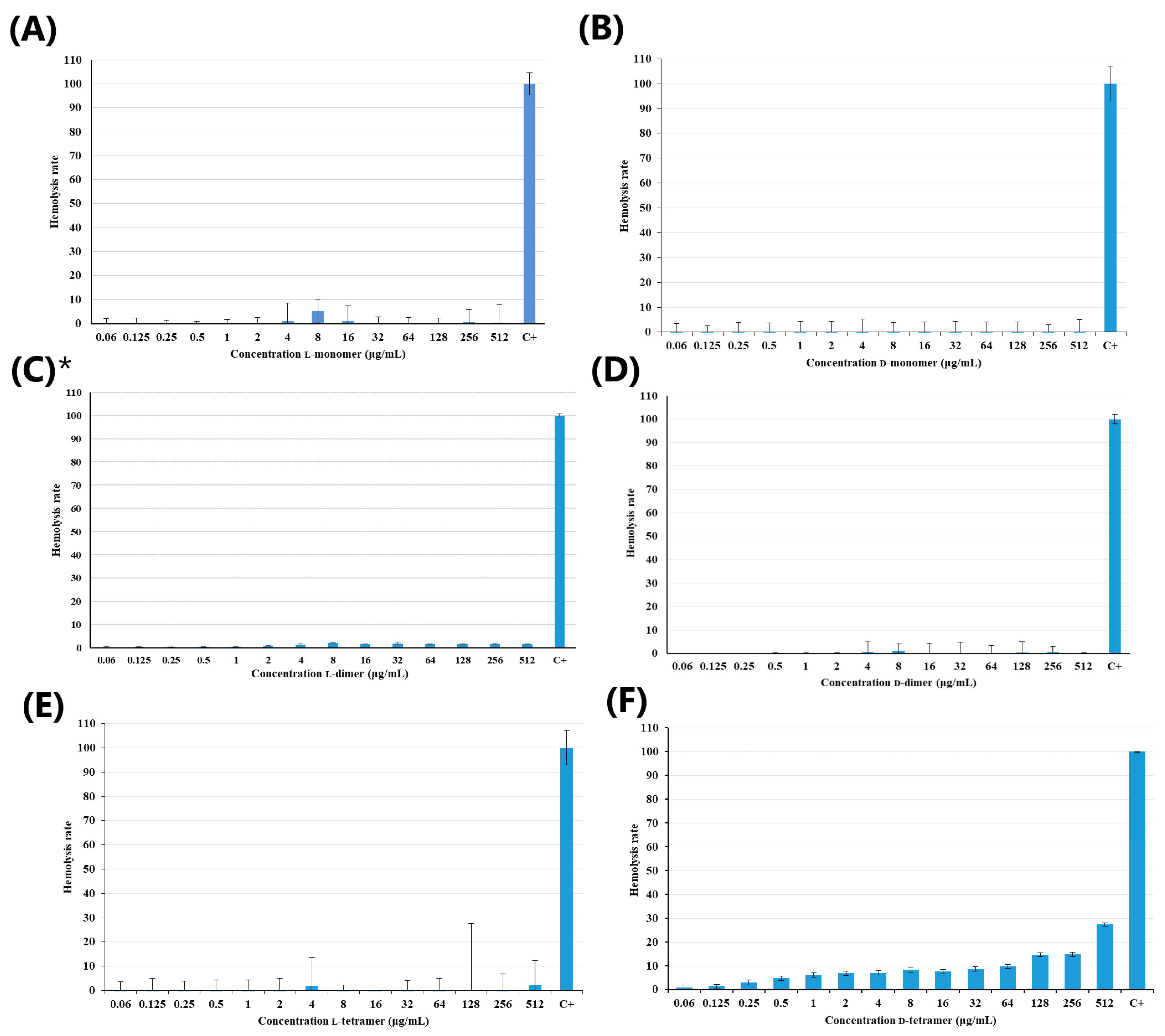
2.3. Hemolysis Assay
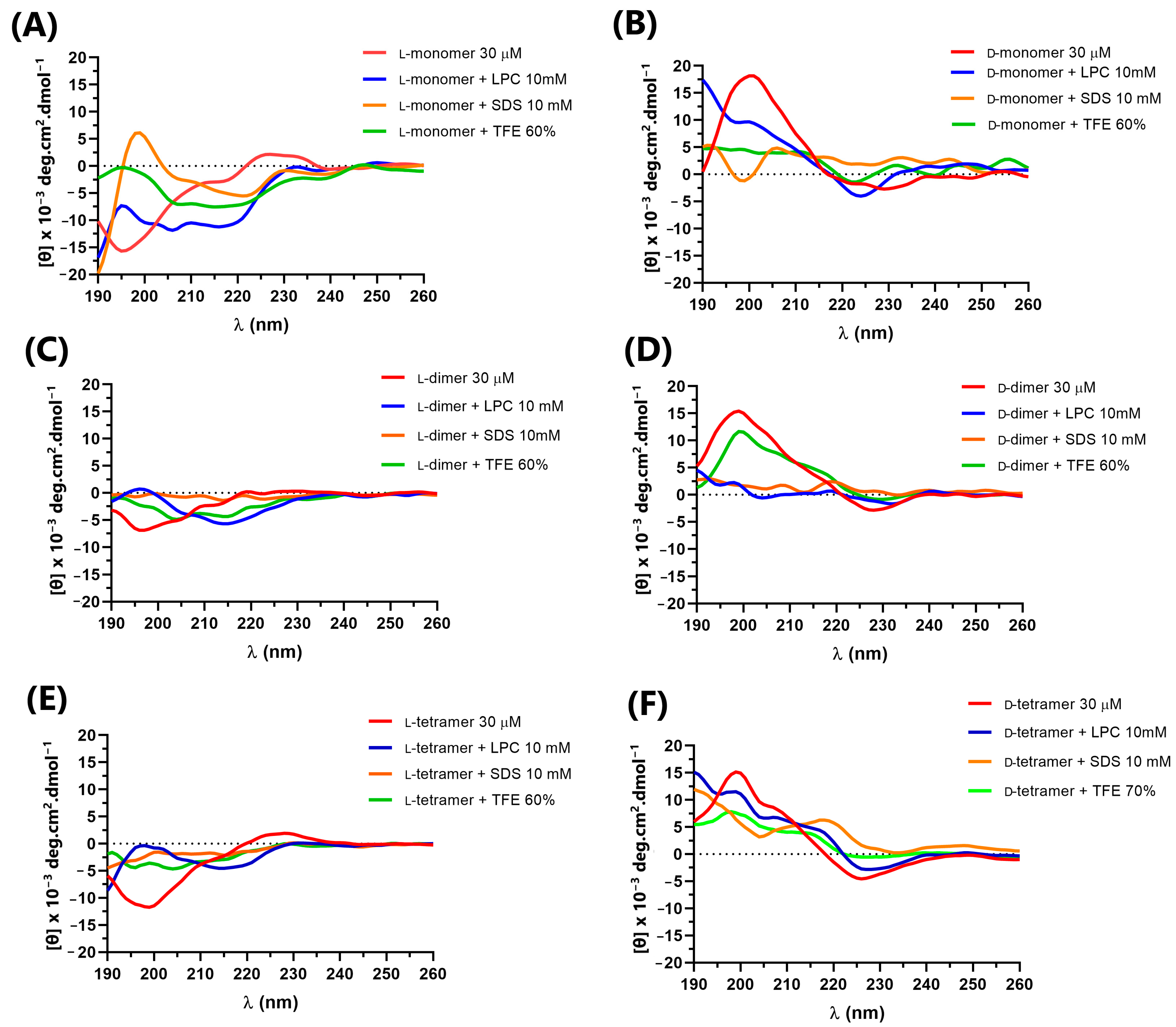
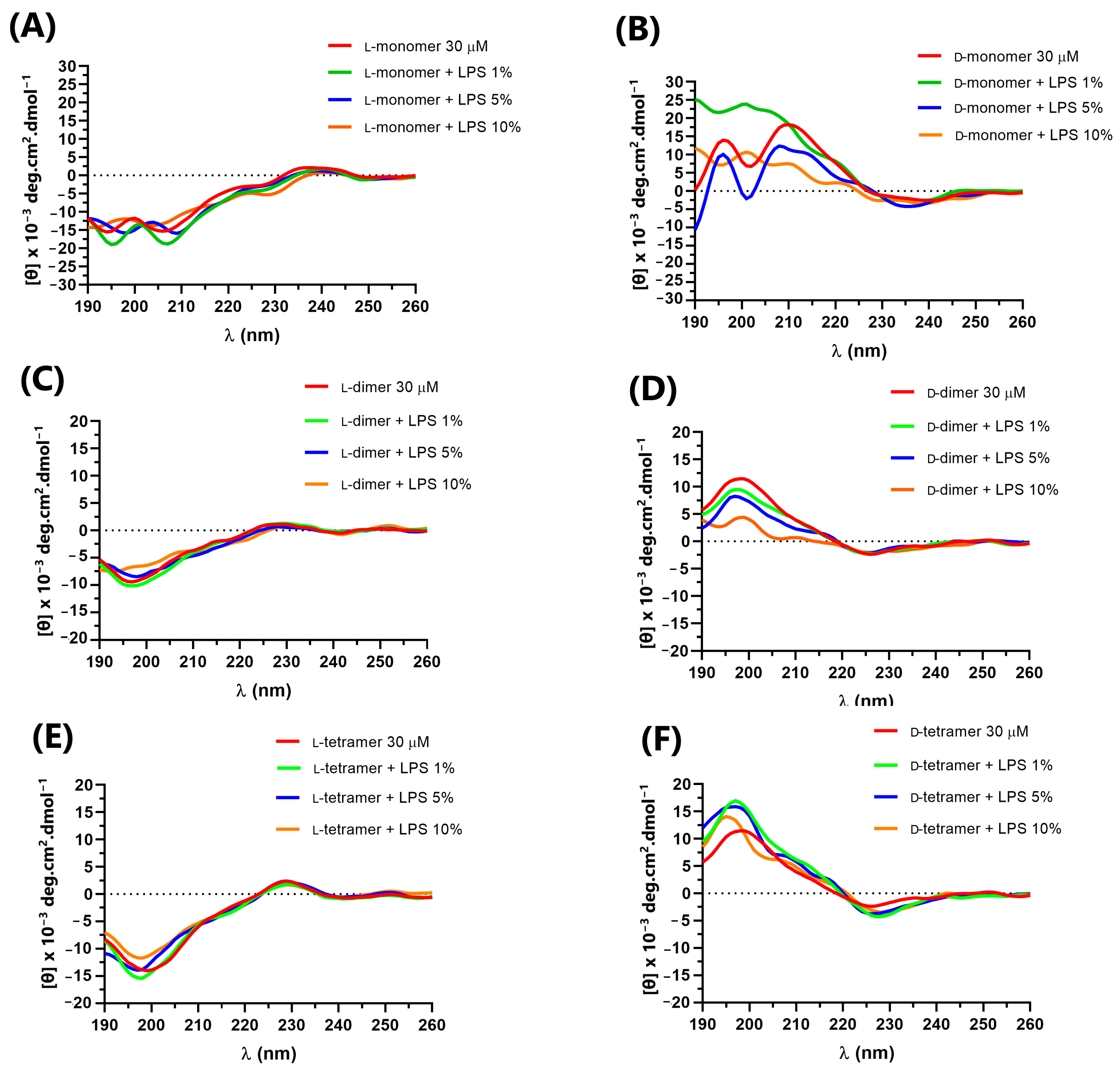
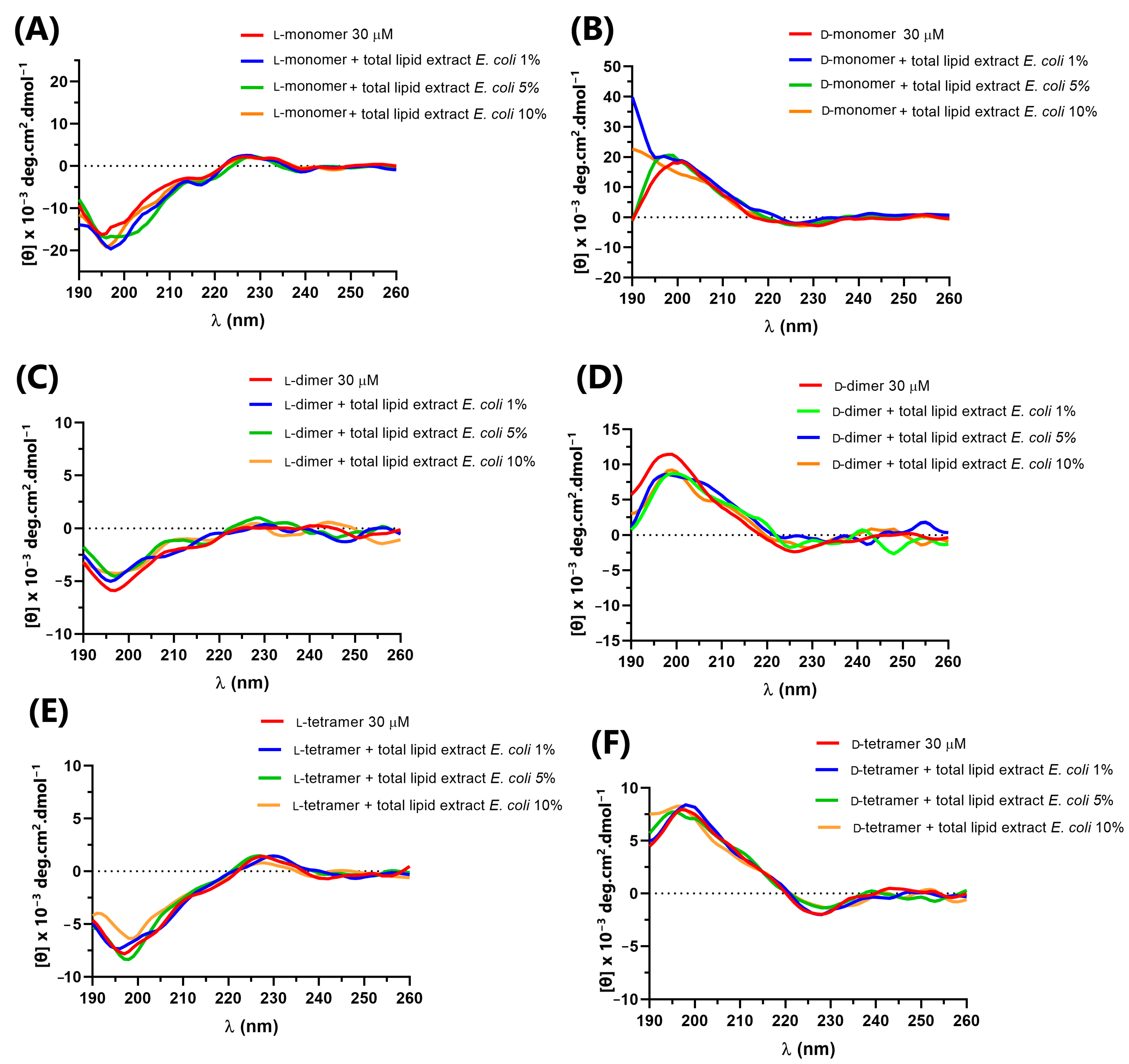
2.4. Circular Dichroism Spectroscopy
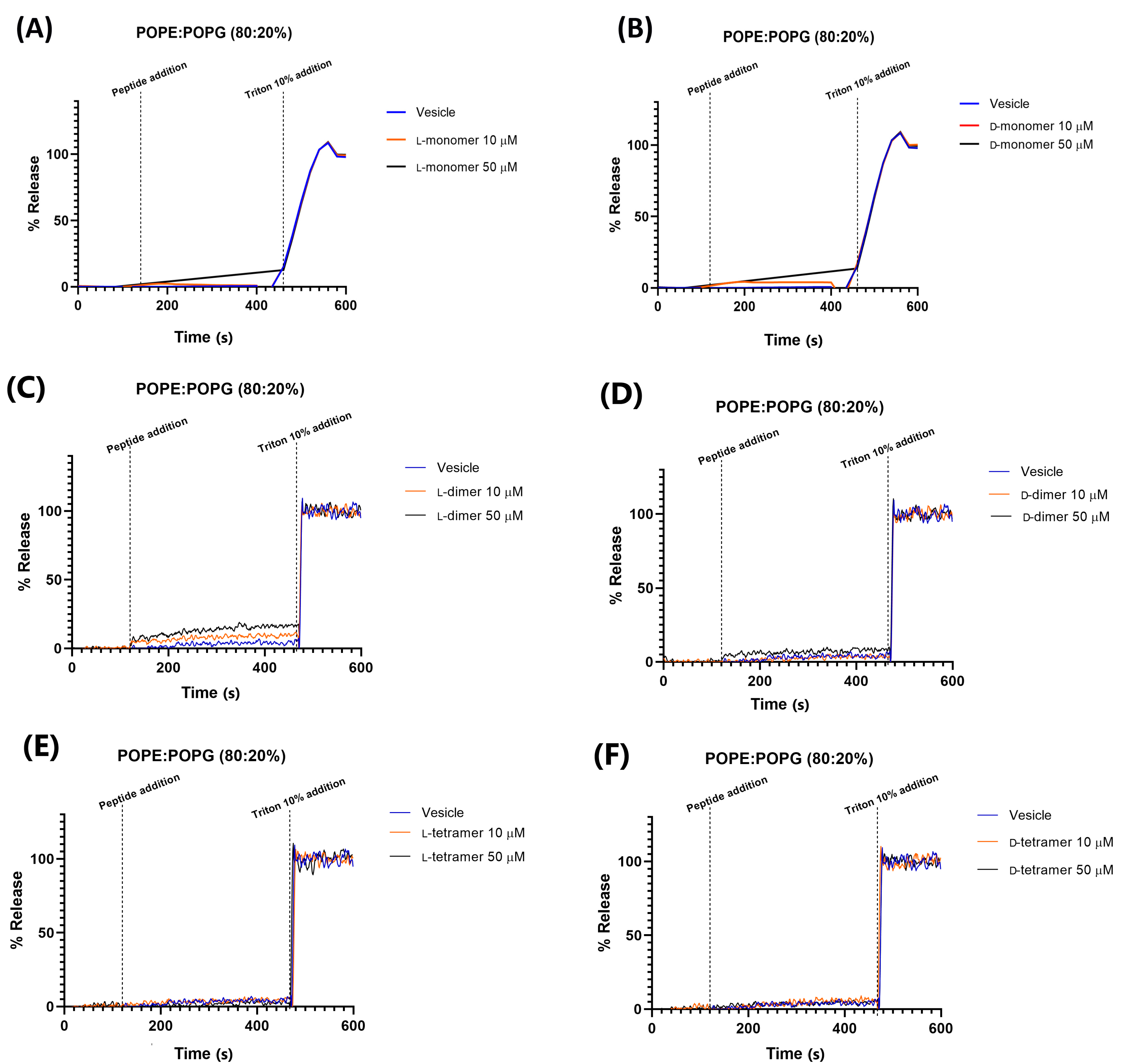
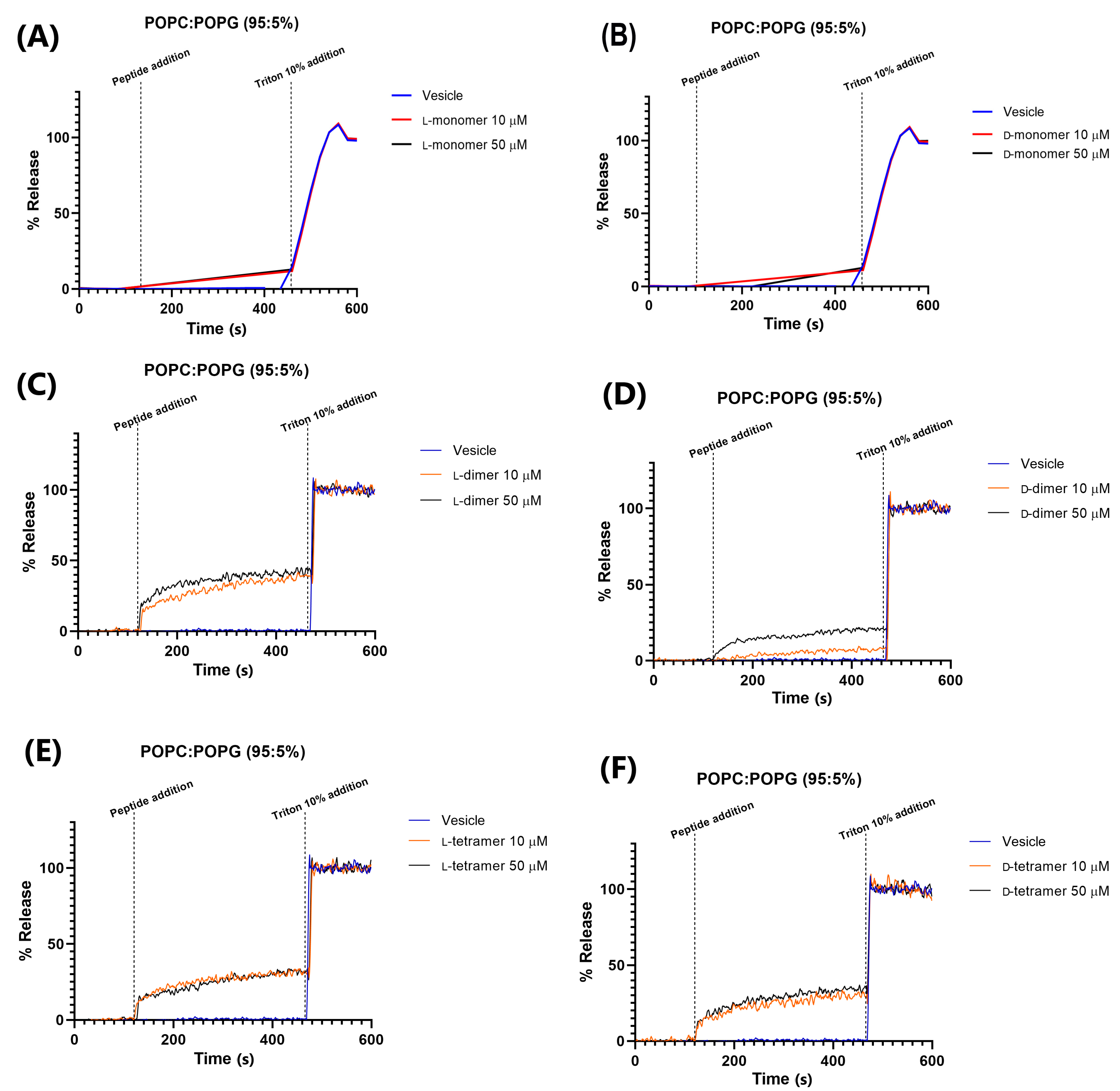
2.5. Permeability in Different Vesicle Compositions
2.6. PLpro Inhibition
3. Results and Discussion
3.1. Peptide Synthesis
3.2. Biological Activity
3.3. Hemolysis Assay
3.4. Circular Dichroism
3.5. Vesicle Permeabilization
3.6. Inhibition of SARS-CoV-2 PLpro Enzymatic Activity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Oliveira, D.M.; Forde, B.M.; Kidd, T.J.; Harris, P.N.; Schembri, M.A.; Beatson, S.A.; Paterson, D.L.; Walker, M.J. Antimicrobial Resistance in ESKAPE Pathogens. Clin. Microbiol. Rev. 2020, 33, e00181-19. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.; Kapley, A. Antibiotic resistance: Global health crisis and metagenomics. Biotechnol. Rep. 2021, 29, e00604. [Google Scholar] [CrossRef] [PubMed]
- Dijksteel, G.S.; Ulrich, M.M.; Middelkoop, E.; Boekema, B.K. Lessons learned from clinical trials using antimicrobial peptides (AMPs). Front. Microbiol. 2021, 12, 616979. [Google Scholar] [CrossRef] [PubMed]
- Costa, N.C.S.; Piccoli, J.P.; Santos-Filho, N.A.; Clementino, L.C.; Fusco-Almeida, A.M.; De Annunzio, S.R.; Cilli, E.M. Antimicrobial activity of RP-1 peptide conjugate with ferrocene group. PLoS ONE 2020, 15, e0228740. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roque-Borda, C.A.; Silva, P.B.; Rodrigues, M.C.; Filippo, L.D.; Duarte, J.L.; Chorilli, M.; Vicente, E.F.; Garrido, S.S.; Pavan, F.R. Antimicrobial peptides as potential new drugs against WHO list of critical, high, and medium priority bacteria. Pharm. Nanotechnol. Eur. J. Med. Chem. 2022, 241, 114640. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.; Flanagan, T.W.; Kharouf, N.; Bertsch, C.; Mancino, D.; Haikel, Y. Antimicrobial Proteins: Structure, Molecular Action, and Therapeutic Potential. Pharmaceutics 2023, 15, 72. [Google Scholar] [CrossRef]
- Lorenzón, E.N.; Piccoli, J.P.; Santos-Filho, N.A.; Cilli, E.M. Dimerization of antimicrobial peptides: A promising strategy to enhance antimicrobial peptide activity. Protein Pept. Lett. 2019, 26, 98–107. [Google Scholar] [CrossRef]
- Liu, Y.S.; Jingru, T.Z.; Jia, Y.Y.; Bingqing, W.Z. The revitalization of antimicrobial peptides in the resistance era. Pharmacol. Res. 2021, 163, 1043–6618. [Google Scholar] [CrossRef]
- Rezende, S.B.; Oshiro, K.G.; Júnior, N.G.; Franco, O.L.; Cardoso, M.H. Advances on chemically modified antimicrobial peptides for generating peptide antibiotics. Chem. Commun. 2021, 57, 11578–11590. [Google Scholar] [CrossRef]
- Gan, B.G.J.; Rowe, S.M.; Deingruber, T.; Spring, D.R. The multifaceted nature of antimicrobial peptides: Current synthetic chemistry approaches and future directions. Chem. Soc. Rev. 2021, 50, 7820–7880. [Google Scholar] [CrossRef]
- Lorenzón, E.N.; Cespedes, G.F.; Vicente, E.F.; Nogueira, L.G.; Bauab, T.M.; Cilli, E.M.; Castro, M.S. Effects of Dimerization on the Structure and Biological Activity of Antimicrobial Peptide Ctx-Ha. Antimicrob. Agents Chemother. 2012, 56, 3004–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mirakabad, F.S.T.; Khoramgah, M.S.; Keshavarz, K.; Tabarzad, M.; Ranjbari, J. Peptide dendrimers as valuable biomaterials in medical sciences. Life Sci. 2019, 233, 116754. [Google Scholar] [CrossRef] [PubMed]
- Giuliani, A.R.; Andrea, C. Beyond natural antimicrobial peptides: Multimeric peptides and other peptidomimetic approaches. Cell. Mol. Life Sci. 2011, 68, 2255–2266. [Google Scholar] [CrossRef] [PubMed]
- Santos-Filho, N.A.; Ramos, M.A.; Santos, C.T.; Piccoli, J.P.; Bauab, T.M.; Fusco-Almeida, A.M.; Cilli, E.M. Synthesis and characterization of an antibacterial and non-toxic dimeric peptide derived from the C-terminal region of Bothropstoxin-I. Toxicon 2015, 103, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Santos-Filho, N.A.; Fernandes, R.S.; Sgardioli, B.F.; Ramos, M.A.S.; Piccoli, J.P.; Camargo, I.L.B.C.; Bauab, T.M.; Cilli, E.M. Antibacterial Activity of the Non-Cytotoxic Peptide (p-BthTX-I)2 and Its Serum Degradation Product against Multidrug-Resistant Bacteria. Molecules 2017, 22, 1898. [Google Scholar] [CrossRef] [Green Version]
- Santos-Filho, N.A.; De Freitas, L.M.; Dos Santos, C.T.; Piccoli, J.P.; Fontana, C.R.; Fusco-Almeida, A.M.; Cilli, E.M. Understanding the mechanism of action of peptide (p-BthTX-I)2 derived from C-terminal region of phospholipase A2 (PLA2)-like bothropstoxin-I on Gram-positive and Gram-negative bacteria. Toxicon 2021, 196, 44–55. [Google Scholar] [CrossRef]
- Santos-Filho, N.A.; Righetto, G.M.; Pereira, M.R.; Piccoli, J.P.; Almeida, L.M.T.; Leal, T.C.; Camargo, I.L.B.C.; Cilli, E.M. Effect of C-terminal and N-terminal dimerization and alanine scanning on antibacterial activity of the analogs of the peptide p-BthTX-I. Pept. Sci. 2021, 114, e24243. [Google Scholar] [CrossRef]
- Vilas Boas, L.C.P.; Campos, M.L.; Berlanda, R.L.A.; De Carvalho Neves, N.; Franco, O.L. Antiviral peptides as promising therapeutic drugs. Cell. Mol. Life Sci. 2019, 76, 3525–3542. [Google Scholar] [CrossRef]
- Freire, M.C.; Noske, G.D.; Bitencourt, N.V.; Sanches, P.R.; Santos-Filho, N.A.; Gawriljuk, V.O.; Souza, E.P.; Nogueira, V.H.R.; Godoy, M.O.; Nakamura, A.M.; et al. Non-Toxic Dimeric Peptides Derived from the Bothropstoxin-I Are Potent SARS-CoV-2 and Papain-like Protease Inhibitors. Molecules 2021, 26, 4896. [Google Scholar] [CrossRef]
- Nogueira, R.S.; Salu, B.R.; Nardelli, V.G.; Bonturi, C.R.; Pereira, M.R.; Maffei, F.H.A.; Cilli, E.M.; Oliva, M.L.V. A snake venom-analog peptide that inhibits SARS-CoV-2 and papain-like protease displays antithrombotic activity in mice arterial thrombosis model, without interfering with bleeding time. Thromb. J. 2023, 21, 3–10. [Google Scholar] [CrossRef]
- Merrifield, R.B. Solid Phase Peptide Synthesis. I. The Synthesis of a Tetrapeptide. J. Am. Chem. Soc. 1963, 85, 2149–2154. [Google Scholar] [CrossRef]
- Pankey, G.A.; Sabath, L.D. Clinical relevance of bacteriostatic versus bactericidal mechanisms of action in the treatment of Gram-positive bacterial infections. Clin. Infect. Dis. 2004, 38, 864–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bobone, S.; Stella, L. Selectivity of Antimicrobial Peptides: A Complex Interplay of Multiple Equilibria. Antimicrob. Pept. 2019, 1117, 175–214. [Google Scholar]
- Wang, Y.; Chang, R.Y.K.; Britton, W.J.; Chan, H.K. Advances in the development of antimicrobial peptides and proteins for inhaled therapy. Adv. Drug Deliv. Rev. 2022, 180, 114066. [Google Scholar] [CrossRef] [PubMed]
- Greco, I.; Molchanova, N.; Holmedal, E.; Jenssen, H.; Hummel, B.D.; Watts, J.L.; Håkansson, J.; Hansen, P.R.; Svenson, J. Correlation between hemolytic activity, cytotoxicity and systemic in vivo toxicity of synthetic antimicrobial peptides. Sci. Rep. 2020, 10, 13206. [Google Scholar] [CrossRef] [PubMed]
- Neubauer, D.; Jaśkiewicz, M.; Migoń, D.; Bauer, M.; Sikora, K.; Sikorska, E.; Kamysz, E.; Kamysz, W. Retro analog concept: Comparative study on physico-chemical and biological properties of selected antimicrobial peptides. Amino Acids 2017, 49, 1755–1771. [Google Scholar] [CrossRef] [Green Version]
- Pathania, A.R. Circular dichroism and its uses in biomolecular research. E3S Web Conf. 2021, 309, 01095. [Google Scholar]
- Keiderling, T.A. Structure of Condensed Phase Peptides: Insights from Vibrational Circular Dichroism and Raman Optical Activity Techniques. Chem. Rev. 2020, 120, 3381–3419. [Google Scholar] [CrossRef]
- Woody, R.W. Circular dichroism of peptides. In The Peptides; Hruby, V.J., Ed.; Academic Press: New York, NY, USA, 1985; Volume 7, pp. 15–114. [Google Scholar]
- Harada, T.; Moriyama, H. Solid-State Circular Dichroism Spectroscopy. Encyclopedia of Polymer Science and Technology; John Wiley & Sons: Hoboken, NJ, USA, 2002. [Google Scholar]
- Ding, L.; Yang, L.; Weiss, T.M.; Waring, A.J.; Lehrer, R.I.; Huang, H.W. Interaction of Antimicrobial Peptides with Lipopolysaccharides. Biochemistry 2003, 42, 12251–12259. [Google Scholar] [CrossRef] [Green Version]
- Torcato, I.M.; Castanho, M.A.R.B.; Henriques, S.T. The Application of Biophysical Techniques to Study Antimicrobial Peptides. Spectrosc. Int. J. 2012, 27, 541–549. [Google Scholar] [CrossRef] [Green Version]
- Zhou, N.; Luo, Z.; Luo, J.; Fan, X.; Cayabyab, M.; Hiraoka, M.; Huang, Z. Exploring the Stereochemistry of CXCR4-Peptide Recognition and Inhibiting HIV-1 Entry with d-Peptides Derived from Chemokines. J. Biol. Chem. 2002, 277, 17476–17485. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Avitabile, C.; D’andrea, L.D.; Romanelli, A. Circular Dichroism studies on the interactions of antimicrobial peptides with bacterial cells. Sci. Rep. 2014, 4, 4293. [Google Scholar] [CrossRef]
- Wimley, W.C.; Hristova, K. The mechanism of membrane permeabilization by peptides: Still an enigma. Aust. J. Chem. 2019, 73, 96–103. [Google Scholar] [CrossRef] [PubMed]
- Corrêa, J.A.F.; Evangelista, A.G.; De Melo Nazareth, T.; Luciano, F.B. Fundamentals on the molecular mechanism of action of antimicrobial peptides. Materialia 2019, 8, 100494. [Google Scholar] [CrossRef]
- Lopes, S.C.; Ribeiro, C.; Gameiro, P. A New Approach to Counteract Bacteria Resistance: A Comparative Study Between Moxifloxacin and a New Moxifloxacin Derivative in Different Model Systems of Bacterial Membrane. Chem. Biol. Drug Des. 2013, 81, 265–274. [Google Scholar] [CrossRef] [PubMed]
- Crusca, E., Jr.; Rezende, A.A.; Marchetto, R.; Mendes-Giannini, M.J.; Fontes, W.; Castro, M.S.; Cilli, E.M. Influence of N-terminus modifications on the biological activity, membrane interaction, and secondary structure of the antimicrobial peptide hylin-a1. Pept. Sci. 2011, 96, 41–48. [Google Scholar] [CrossRef] [PubMed]
- Gogoi, P.; Shrivastava, S.; Shah, P.; Saxena, S.; Srivastava, S.; Gaur, G.K. Linear and branched forms of short antimicrobial peptide-IRK inhibit growth of multi drug resistant Staphylococcus aureus isolates from mastitic cow milk. Int. J. Pept. Res. Ther. 2021, 27, 2149–2159. [Google Scholar] [CrossRef]
- Ludtke, S.J.; He, K.; Heller, W.T.; Harroun, T.A.; Yang, L.; Huang, H.W. Membrane pores induced by magainin. Biochemistry 1996, 35, 13723–13728. [Google Scholar] [CrossRef]
- Zhu, Y.; Li, J.; Pang, Z. Recent insights for the emerging COVID-19: Drug discovery, therapeutic options and vaccine development. Asian J. Pharm. Sci. 2021, 16, 4–23. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Peptides | S. aureus ATCC 25923 | S. epidermidis ATCC 35984 | E. faecalis ATCC 29212 | E. faecium ATCC 700221 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC μg/mL | MIC µM | MBC μg/mL | Act | MIC μg/mL | MIC µM | MBC μg/mL | Act | MIC μg/mL | MIC µM | MBC μg/mL | Act | MIC μg/mL | MIC µM | MBC μg/mL | Act | |
| L-monomer | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND |
| D-monomer | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND |
| L-dimer | 32 | 11 | 128 | Bc | 8 | 2 | 16 | Bc | 64 | 22 | 128 | Bc | 16 | 5 | >64 | Bs |
| D-dimer | 256 | 89 | >512 | ND | 16 | 5 | 16 | Bc | 512 | 178 | 512 | Bc | 64 | 22 | >256 | Bs |
| L-tetramer | 64 | 10 | 256 | Bc | 16 | 2 | 32 | Bc | 64 | 10 | 128 | Bc | 16 | 2 | 32 | Bc |
| D-tetramer | 64 | 10 | 64 | Bc | 16 | 2 | 16 | Bc | 32 | 5 | 32 | Bc | 16 | 2 | 16 | Bc |
| Peptides | K. pneumoniae ATCC 700603 | E. coli ATCC 25922 | A. baumannii ATCC 19606 | P. aeruginosa ATCC 27853 | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| MIC μg/mL | MIC µM | MBC μg/mL | Act | MIC μg/mL | MIC µM | MBC μg/mL | Act | MIC μg/mL | MIC µM | MBC μg/mL | Act | MIC μg/mL | MIC µM | MBC μg/mL | Act | |
| L-monomer | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND |
| D-monomer | 512 | 371 | 512 | Bc | 256 | 185 | 256 | Bc | >512 | >371 | >512 | ND | >512 | >371 | >512 | ND |
| L-dimer | >128 | >44 | ND | ND | 32 | 11 | 64 | Bc | 256 | 89 | >512 | ND | >512 | >178 | ND | ND |
| D-dimer | 64 | 22 | 128 | Bc | 32 | 11 | 32 | Bc | 16 | 5 | 64 | Bc | 256 | 89 | 512 | Bc |
| L-tetramer | 32 | 5 | >128 | Bs | 32 | 5 | >128 | Bs | 32 | 5 | 64 | Bc | 32 | 5 | 64 | Bc |
| D-tetramer | 32 | 5 | 32 | Bc | 32 | 5 | >128 | Bs | 32 | 5 | 32 | Bc | 32 | 5 | 64 | Bc |
| Peptide | Inhibition (%) at 10 µM | IC50 μM |
|---|---|---|
| L-monomer | 93.30 ± 0.00 | 2.1 ± 0.1 |
| D-monomer | 85.00 ± 3.00 | 3.0 ± 1 |
| L-dimer * | 98.05 ± 1.62 | 2.4 ± 0.1 |
| D-dimer * | 96.35 ± 0.91 | 1.30 ± 0.03 |
| L-tetramer * | 98.40 ± 0.00 | 1.40 ± 0.02 |
| D-tetramer | 94.00 ± 3.00 | 2.7 ± 0.5 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bitencourt, N.V.; Righetto, G.M.; Camargo, I.L.B.C.; de Godoy, M.O.; Guido, R.V.C.; Oliva, G.; Santos-Filho, N.A.; Cilli, E.M. Effects of Dimerization, Dendrimerization, and Chirality in p-BthTX-I Peptide Analogs on the Antibacterial Activity and Enzymatic Inhibition of the SARS-CoV-2 PLpro Protein. Pharmaceutics 2023, 15, 436. https://doi.org/10.3390/pharmaceutics15020436
Bitencourt NV, Righetto GM, Camargo ILBC, de Godoy MO, Guido RVC, Oliva G, Santos-Filho NA, Cilli EM. Effects of Dimerization, Dendrimerization, and Chirality in p-BthTX-I Peptide Analogs on the Antibacterial Activity and Enzymatic Inhibition of the SARS-CoV-2 PLpro Protein. Pharmaceutics. 2023; 15(2):436. https://doi.org/10.3390/pharmaceutics15020436
Chicago/Turabian StyleBitencourt, Natália Vitória, Gabriela Marinho Righetto, Ilana Lopes Baratella Cunha Camargo, Mariana Ortiz de Godoy, Rafael Victorio Carvalho Guido, Glaucius Oliva, Norival Alves Santos-Filho, and Eduardo Maffud Cilli. 2023. "Effects of Dimerization, Dendrimerization, and Chirality in p-BthTX-I Peptide Analogs on the Antibacterial Activity and Enzymatic Inhibition of the SARS-CoV-2 PLpro Protein" Pharmaceutics 15, no. 2: 436. https://doi.org/10.3390/pharmaceutics15020436