
Trioleyl Pyridinium, a Cationic Transfection Agent for the Lipofection of Therapeutic Oligonucleotides into Mammalian Cells
 , , , , and
, , , , and 
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Instrumentation
2.2. Preparation of TROPY
2.3. Design and Usage of PPRHs
2.4. Agarose Gel Retardation Assays
2.5. Cell Culture
2.6. Transfection of DNA
2.7. DLS and Z-Potencial
2.8. Cellular Uptake
2.9. Confocal Analyses
2.10. MTT Assays
2.11. Analysis of Survivin Expression
2.11.1. Survivin mRNA Levels
2.11.2. Survivin Protein Levels
2.12. Apoptosis Assays
3. Results and Discussion
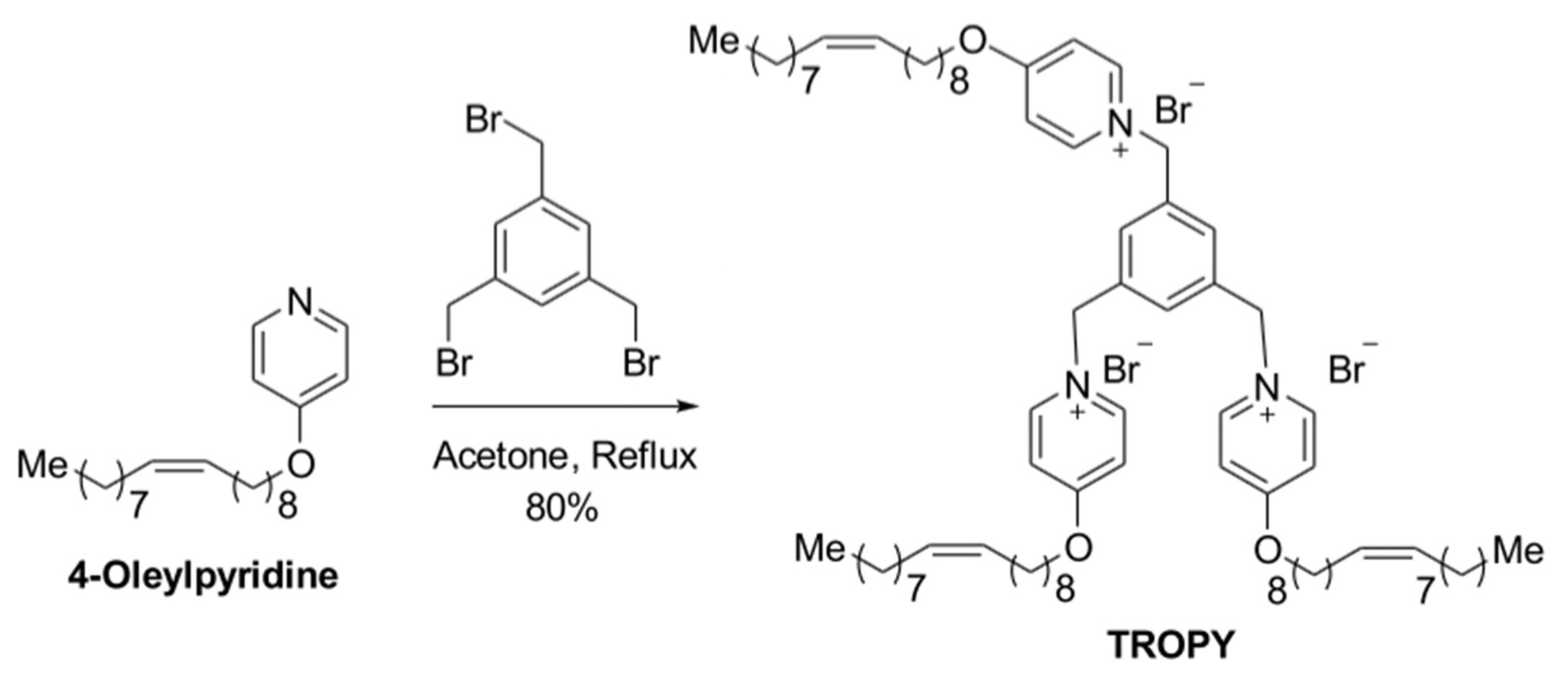
3.1. Synthesis of TROPY
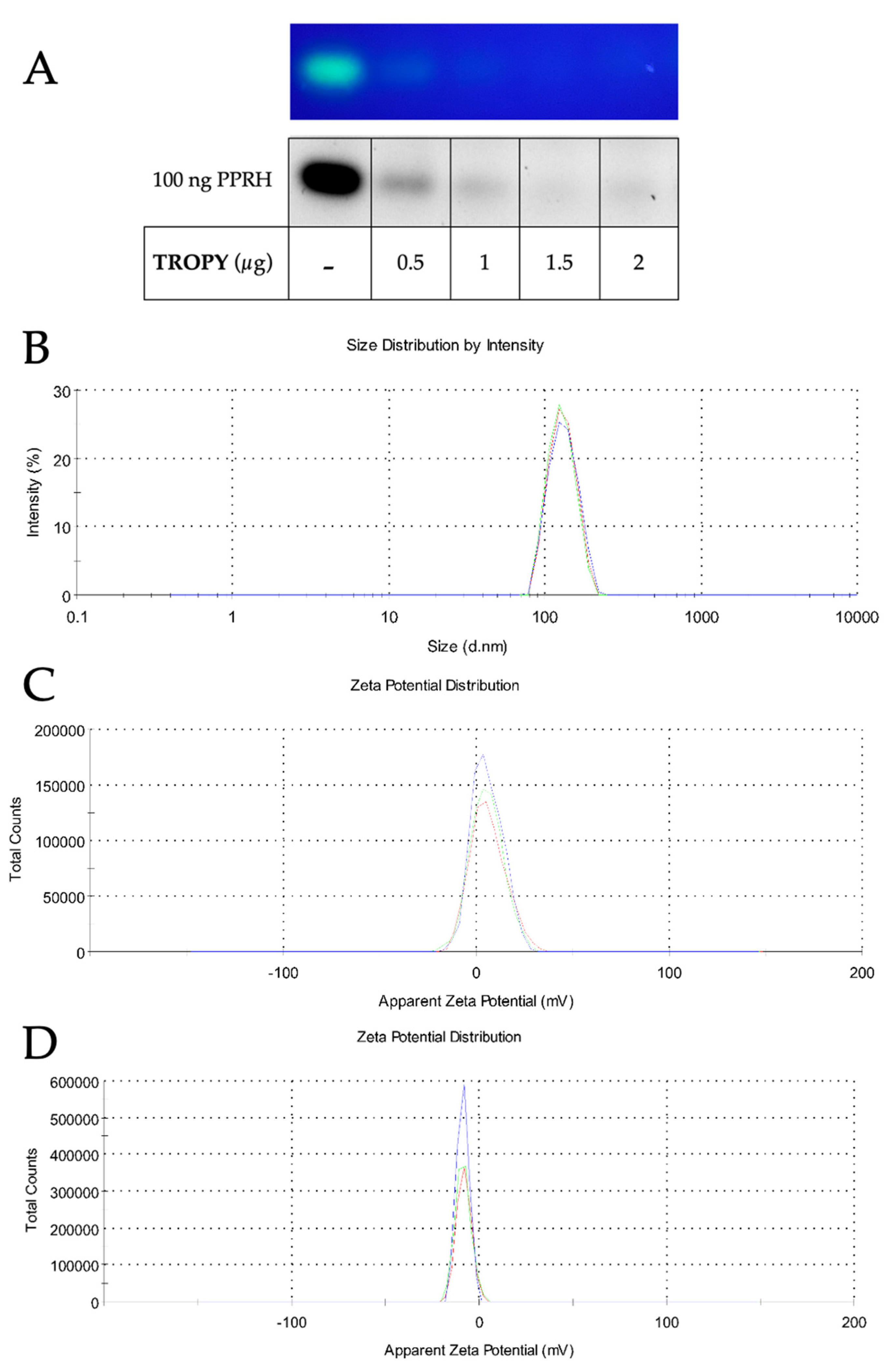
3.2. Characterization of TROPY/PPRH Complexes
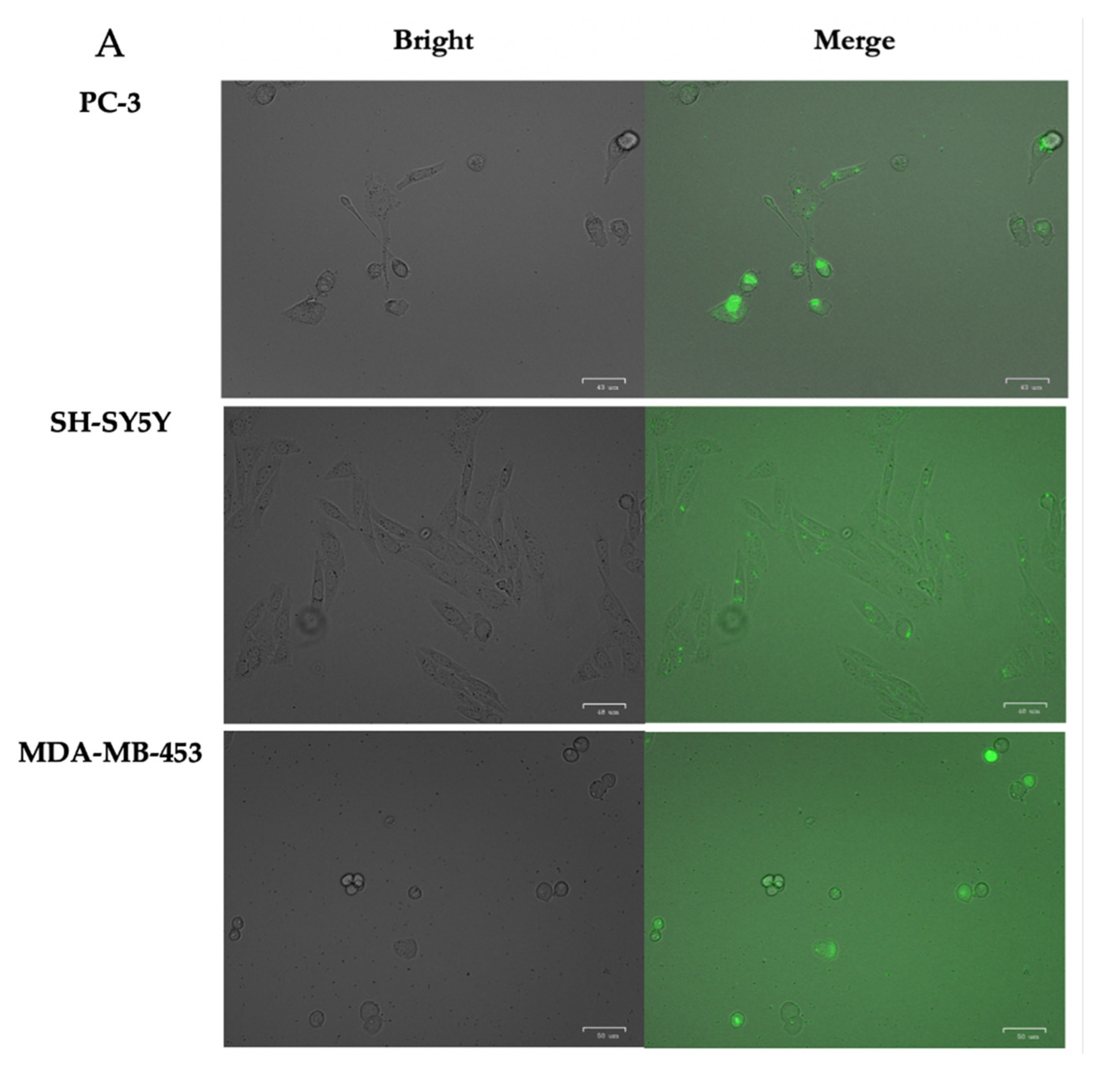
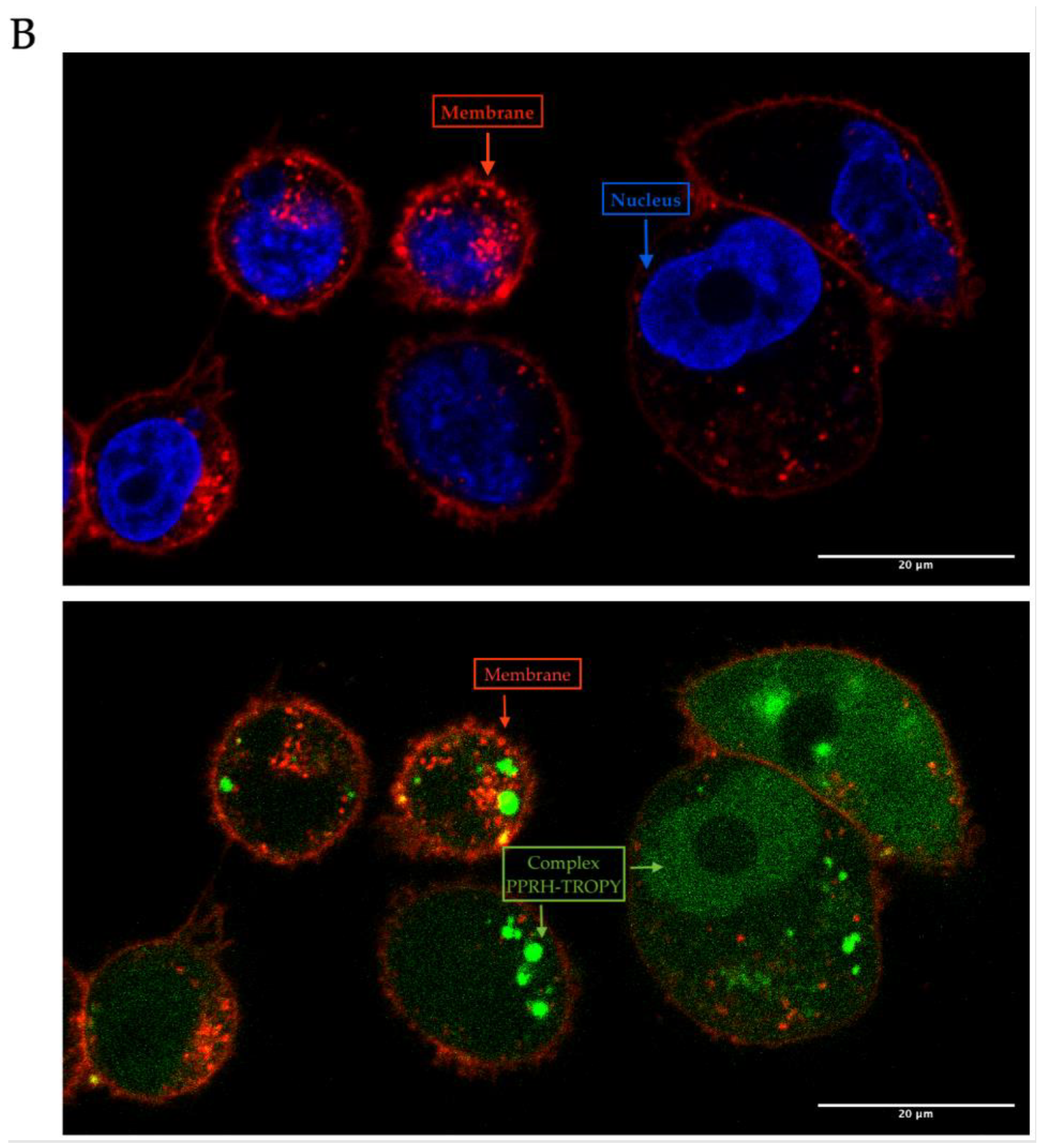
3.3. Internalization of PPRH into Mammalian Cells by Fluorescence and Confocal Microscopy
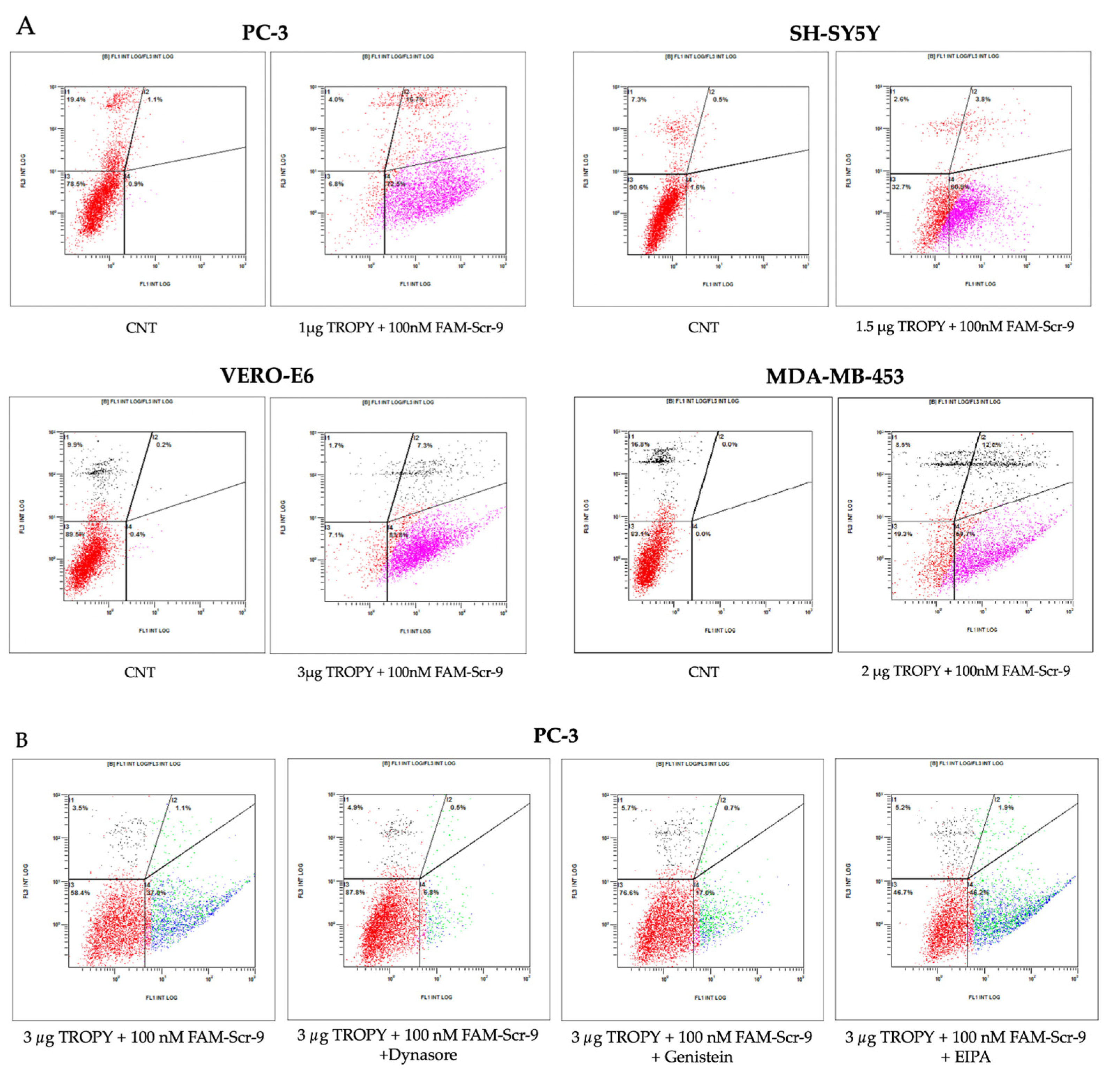
3.4. Cellular Uptake by Flow Cytometry
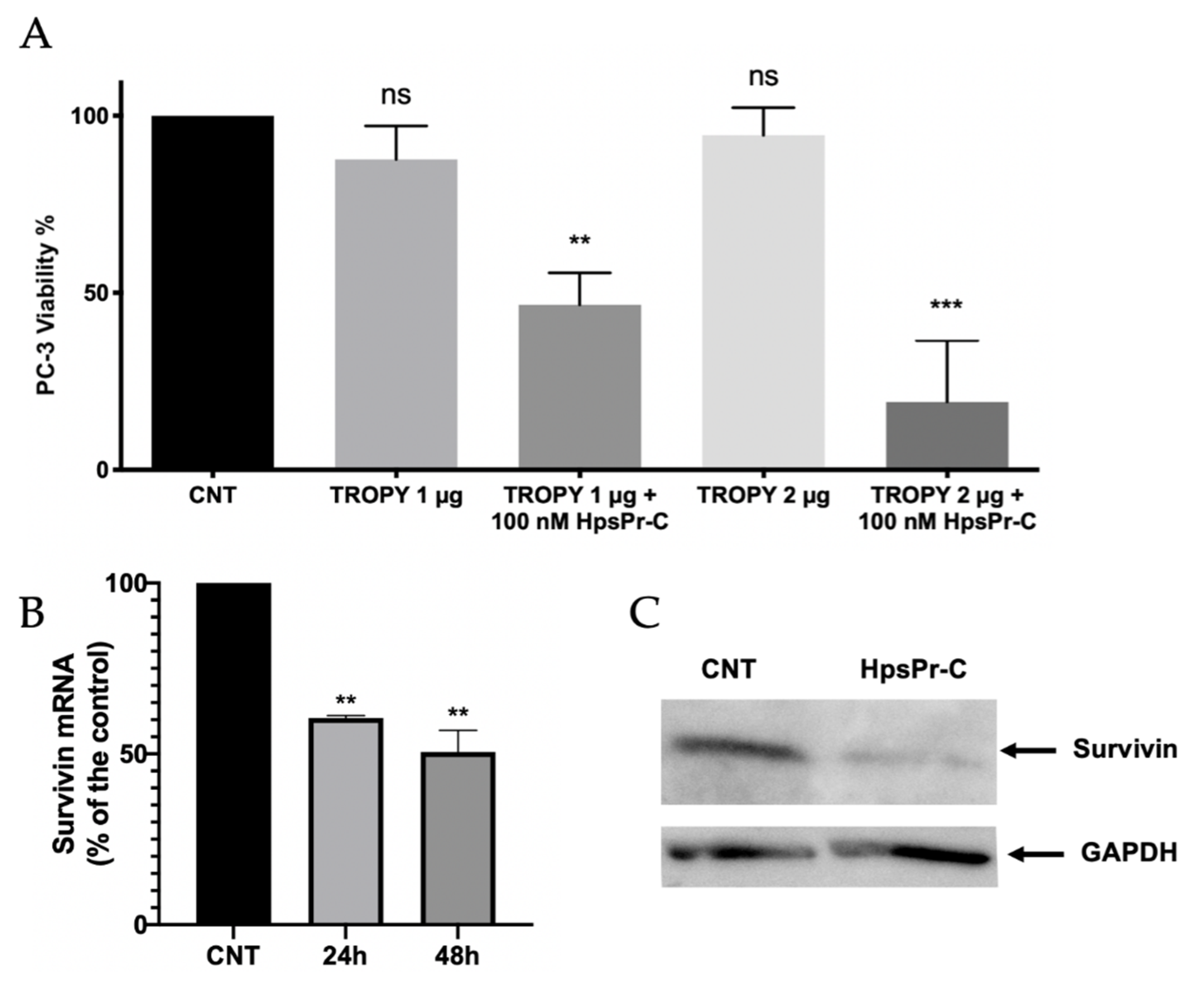
3.5. Effect on Cell Viability and Survivin Expression Caused by PPRH against Survivin Transfected Using TROPY
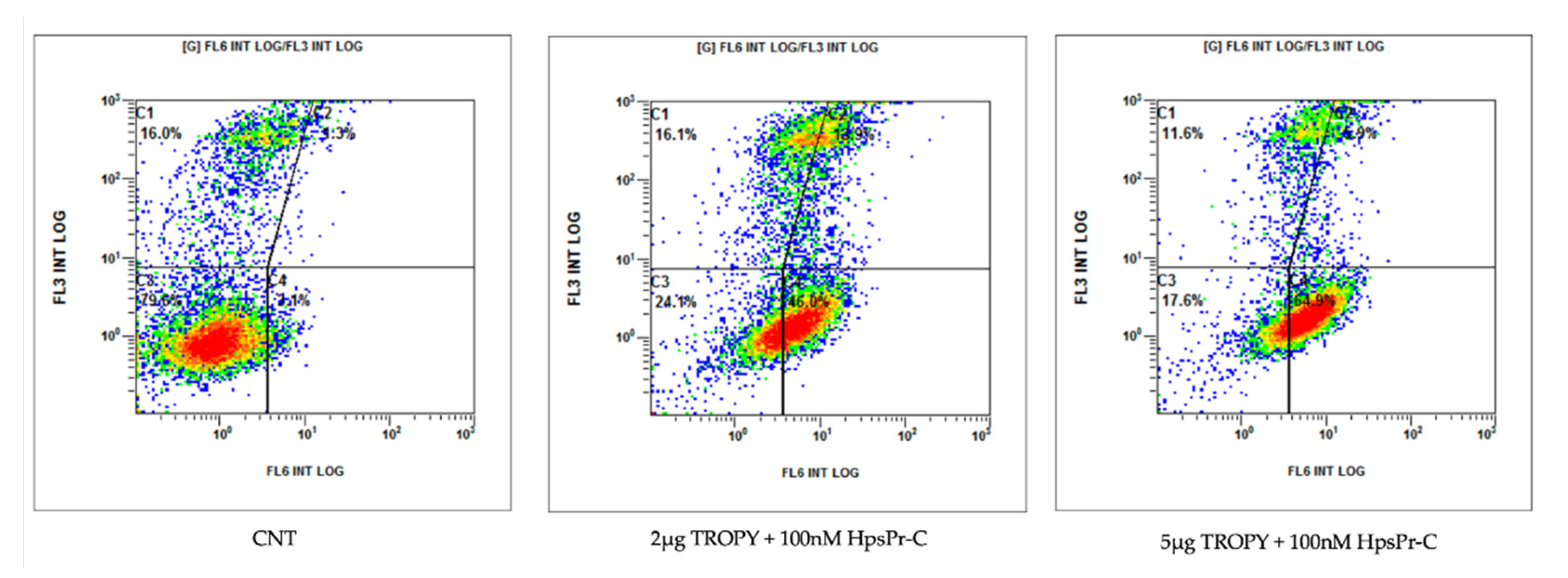
3.6. Effect on Apoptosis Caused by PPRH against Survivin Transfected with TROPY
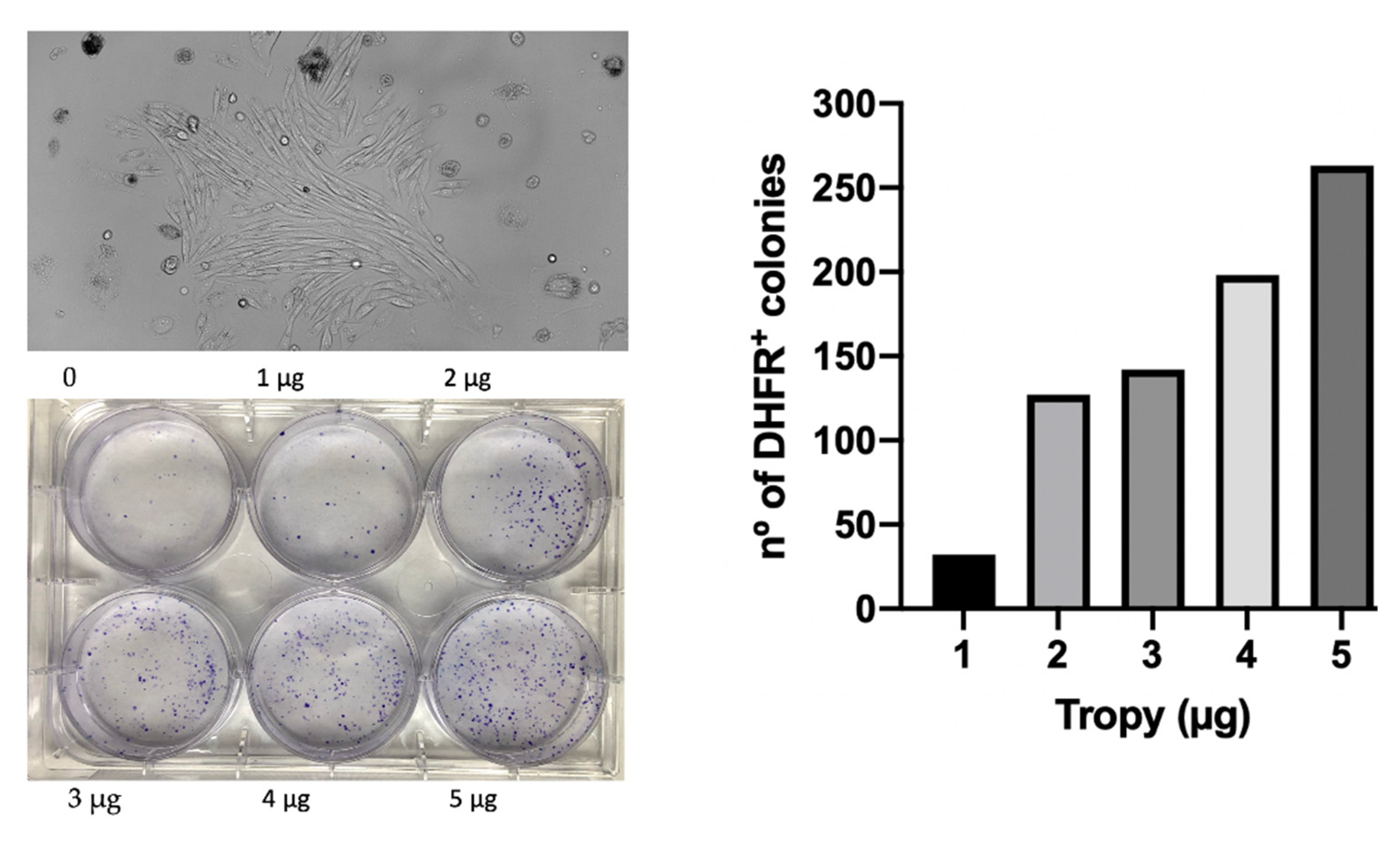
3.7. Transfection of Plasmid DNA into Mammalian Cells
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Amer, M.H. Gene Therapy for Cancer: Present Status and Future Perspective. Mol. Cell. Ther. 2014, 2, 27. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, T.T.; Nielsen, J.E. Antisense Gene Silencing: Therapy for Neurodegenerative Disorders? Genes 2013, 4, 457–484. [Google Scholar] [CrossRef] [Green Version]
- Winkelsas, A.M.; Fischbeck, K.H. Nucleic Acid Therapeutics in Neurodevelopmental Disease. Curr. Opin. Genet. Dev. 2020, 65, 112–116. [Google Scholar] [CrossRef]
- Walsh, C.E. Gene Therapy Progress and Prospects: Gene Therapy for the Hemophilias. Gene Ther. 2003, 10, 999–1003. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hazan-Halevy, I.; Landesman-Milo, D.; Rosenblum, D.; Mizrahy, S.; Ng, B.D.; Peer, D. Immunomodulation of Hematological Malignancies Using Oligonucleotides Based-Nanomedicines. J. Control. Release 2016, 244, 149–156. [Google Scholar] [CrossRef] [PubMed]
- FDA News Release. FDA Grants Accelerated Approval to First Drug for Duchenne Muscular Dystrophy. Available online: https://www.fda.gov/news-events/press-announcements/fda-grants-accelerated-approval-first-drug-duchenne-muscular-dystrophy (accessed on 22 December 2022).
- FDA. FDA Approves First Drug to Treat Rare Metabolic Disorder. 2020. Available online: https://www.fda.gov/news-events/press-announcements/fda-approves-first-drug-treat-rare-metabolic-disorder (accessed on 22 December 2022).
- Highleyman, L. FDA Approves Fomivirsen, Famciclovir, and Thalidomide. Food and Drug Administration. Available online: https://pubmed.ncbi.nlm.nih.gov/11365993/ (accessed on 22 December 2022).
- Doggrell, S.A. Pegaptanib: The First Antiangiogenic Agent Approved for Neovascular Macular Degeneration. Expert Opin. Pharmacother. 2005, 6, 1421–1423. [Google Scholar] [CrossRef] [PubMed]
- Roberts, T.C.; Langer, R.; Wood, M.J.A. Advances in Oligonucleotide Drug Delivery. Nat. Rev. Drug Discov. 2020, 19, 673–694. [Google Scholar] [CrossRef]
- Noé, V.; Aubets, E.; Félix, A.J.; Ciudad, C.J. Nucleic Acids Therapeutics Using PolyPurine Reverse Hoogsteen Hairpins. Biochem. Pharmacol. 2020, 189, 114371. [Google Scholar] [CrossRef]
- Ciudad, C.J.; Rodríguez, L.; Villalobos, X.; Félix, A.J.; Noé, V. Polypurine Reverse Hoogsteen Hairpins as a Gene Silencing Tool for Cancer. Curr. Med. Chem. 2017, 24, 2809–2826. [Google Scholar] [CrossRef]
- Rodríguez, L.; Villalobos, X.; Solé, A.; Lliberós, C.; Ciudad, C.J.; Noé, V. Improved Design of PPRHs for Gene Silencing. Mol. Pharm. 2015, 12, 867–877. [Google Scholar] [CrossRef]
- Aubets, E.; Félix, A.J.; Garavís, M.; Reyes, L.; Aviñó, A.; Eritja, R.; Ciudad, C.J.; Noé, V. Detection of a G-Quadruplex as a Regulatory Element in Thymidylate Synthase for Gene Silencing Using Polypurine Reverse Hoogsteen Hairpins. Int. J. Mol. Sci. 2020, 21, 5028. [Google Scholar] [CrossRef]
- Rodríguez, L.; Villalobos, X.; Dakhel, S.; Padilla, L.; Hervas, R.; Hernández, J.L.; Ciudad, C.J.; Noé, V. Polypurine Reverse Hoogsteen Hairpins as a Gene Therapy Tool against Survivin in Human Prostate Cancer PC3 Cells in Vitro and in Vivo. Biochem. Pharmacol. 2013, 86, 1541–1554. [Google Scholar] [CrossRef]
- Pichon, C.; Billiet, L.; Midoux, P. Chemical Vectors for Gene Delivery: Uptake and Intracellular Trafficking. Curr. Opin. Biotechnol. 2010, 21, 640–645. [Google Scholar] [CrossRef]
- de Almagro, M.C.C.; Coma, S.; Noé, V.; Ciudad, C.J. Polypurine Hairpins Directed against the Template Strand of DNA Knock down the Expression of Mammalian Genes. J. Biol. Chem. 2009, 284, 11579–11589. [Google Scholar] [CrossRef] [Green Version]
- Villalobos, X.; Rodríguez, L.; Prévot, J.; Oleaga, C.; Ciudad, C.J.; Noé, V. Stability and Immunogenicity Properties of the Gene-Silencing Polypurine Reverse Hoogsteen Hairpins. Mol. Pharm. 2014, 11, 254–264. [Google Scholar] [CrossRef]
- Félix, A.J.; Ciudad, C.J.; Noé, V. Functional Pharmacogenomics and Toxicity of PolyPurine Reverse Hoogsteen Hairpins Directed against Survivin in Human Cells. Biochem. Pharmacol. 2018, 155, 8–20. [Google Scholar] [CrossRef]
- Nayerossadat, N.; Ali, P.; Maedeh, T. Viral and Nonviral Delivery Systems for Gene Delivery. Adv. Biomed. Res. 2012, 1, 27. [Google Scholar] [CrossRef]
- Kim, J.; Kang, Y.; Tzeng, S.Y.; Green, J.J. Synthesis and Application of Poly(Ethylene Glycol)-Co-Poly(β-Amino Ester) Copolymers for Small Cell Lung Cancer Gene Therapy. Acta Biomater. 2016, 41, 293–301. [Google Scholar] [CrossRef]
- Zhang, J.; Wang, Z.; Lin, W.; Chen, S. Gene Transfection in Complex Media Using PCBMAEE-PCBMA Copolymer with Both Hydrolytic and Zwitterionic Blocks. Biomaterials 2014, 35, 7909–7918. [Google Scholar] [CrossRef]
- Belmadi, N.; Midoux, P.; Loyer, P.; Passirani, C.; Pichon, C.; Le Gall, T.; Jaffres, P.A.; Lehn, P.; Montier, T. Synthetic Vectors for Gene Delivery: An Overview of Their Evolution Depending on Routes of Administration. Biotechnol. J. 2015, 10, 1370–1389. [Google Scholar] [CrossRef]
- Slade, N. Viral Vectors in Gene Therapy. Period. Biol. 2001, 103, 139–143. [Google Scholar]
- Alsaggar, M.; Liu, D. Physical Methods for Gene Transfer. Adv. Genet. 2015, 89, 1–24. [Google Scholar] [CrossRef]
- Pack, D.W.; Hoffman, A.S.; Pun, S.; Stayton, P.S. Design and Development of Polymers for Gene Delivery. Nat. Rev. Drug Discov. 2005, 4, 581–593. [Google Scholar] [CrossRef]
- Florea, B.I.; Meaney, C.; Junginger, H.E.; Borchard, G. Transfection Efficiency and Toxicity of Polyethylenimine in Differentiated Calu-3 and Nondifferentiated COS-1 Cell Cultures. AAPS J. 2002, 4, 12. [Google Scholar] [CrossRef] [PubMed]
- García-Garrido, E.; Cordani, M.; Somoza, Á. Modified Gold Nanoparticles to Overcome the Chemoresistance to Gemcitabine in Mutant P53 Cancer Cells. Pharmaceutics 2021, 13, 2067. [Google Scholar] [CrossRef]
- Falsini, S.; Ristori, S. Lipoplexes from Non-Viral Cationic Vectors: DOTAP-DOPE Liposomes and Gemini Micelles. In Methods in Molecular Biology; Humana Press Inc.: Totowa, NJ, USA, 2016; Volume 1445, pp. 33–43. [Google Scholar]
- Leventis, R.; Silvius, J.R. Interactions of Mammalian Cells with Lipid Dispersions Containing Novel Metabolizable Cationic Amphiphiles. BBA-Biomembr. 1990, 1023, 124–132. [Google Scholar] [CrossRef]
- Aubets, E.; Griera, R.; Felix, A.J.; Rigol, G.; Sikorski, C.; Limón, D.; Mastrorosa, C.; Busquets, M.A.; Pérez-García, L.; Noé, V.; et al. Synthesis and Validation of DOPY: A New Gemini Dioleylbispyridinium Based Amphiphile for Nucleic Acid Transfection. Eur. J. Pharm. Biopharm. 2021, 165, 279–292. [Google Scholar] [CrossRef] [PubMed]
- Gaddis, S.S.; Wu, Q.; Thames, H.D.; Digiovanni, J.; Walborg, E.F.; Macleod, M.C.; Vasquez, K.M. A Web-Based Search Engine for Triplex-Forming Oligonucleotide Target Sequences. Oligonucleotides 2006, 16, 196–201. [Google Scholar] [CrossRef] [PubMed]
- Urlaub, G.; Mitchell, P.J.; Kas, E.; Chasin, L.A.; Funanage, V.L.; Myoda, T.T.; Hamlin, J. Effect of Gamma Rays at the Dihydrofolate Reductase Locus: Deletions and Inversions. Somat. Cell Mol. Genet. 1986, 12, 555–566. [Google Scholar] [CrossRef]
- Macia, E.; Ehrlich, M.; Massol, R.; Boucrot, E.; Brunner, C.; Kirchhausen, T. Dynasore, a Cell-Permeable Inhibitor of Dynamin. Dev. Cell 2006, 10, 839–850. [Google Scholar] [CrossRef] [Green Version]
- Vercauteren, D.; Vandenbroucke, R.E.; Jones, A.T.; Rejman, J.; Demeester, J.; De Smedt, S.C.; Sanders, N.N.; Braeckmans, K. The Use of Inhibitors to Study Endocytic Pathways of Gene Carriers: Optimization and Pitfalls. Mol. Ther. 2010, 18, 561–569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lin, H.-P.; Singla, B.; Ghoshal, P.; Faulkner, J.L.; Cherian-Shaw, M.; O’Connor, P.M.; She, J.-X.; Belin de Chantemele, E.J.; Csányi, G. Identification of Novel Macropinocytosis Inhibitors Using a Rational Screen of Food and Drug Administration-Approved Drugs. Br. J. Pharmacol. 2018, 175, 3640–3655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubruel, P.; Christiaens, B.; Vanloo, B.; Bracke, K.; Rosseneu, M.; Vandekerckhove, J.; Schacht, E. Physicochemical and Biological Evaluation of Cationic Polymethacrylates as Vectors for Gene Delivery. Eur. J. Pharm. Sci. 2003, 18, 211–220. [Google Scholar] [CrossRef]
- Pajuste, K.; Hyvönen, Z.; Petrichenko, O.; Kaldre, D.; Rucins, M.; Cekavicus, B.; Ose, V.; Skrivele, B.; Gosteva, M.; Morin-Picardat, E.; et al. Gene Delivery Agents Possessing Antiradical Activity: Self-Assembling Cationic Amphiphilic 1,4-Dihydropyridine Derivatives. New J. Chem. 2013, 37, 3062–3075. [Google Scholar] [CrossRef]
- Cai, X.; Ma, S.; Gu, M.; Zu, C.; Qu, W.; Zheng, X. Survivin Regulates the Expression of VEGF-C in Lymphatic Metastasis of Breast Cancer. Diagn. Pathol. 2012, 7, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, G.H.; Joo, Y.E.; Koh, Y.S.; Chung, I.J.; Park, Y.K.; Lee, J.H.; Kim, H.S.; Choi, S.K.; Rew, J.S.; Park, C.S.; et al. Expression of Survivin in Gastric Cancer and Its Relationship with Tumor Angiogenesis. Eur. J. Gastroenterol. Hepatol. 2006, 18, 957–963. [Google Scholar] [CrossRef]
- Miyachi, K.; Sasaki, K.; Onodera, S.; Taguchi, T.; Nagamachi, M.; Kaneko, H.; Sunagawa, M. Correlation between Survivin MRNA Expression and Lymph Node Metastasis in Gastric Cancer. Gastric Cancer 2003, 6, 217–224. [Google Scholar] [CrossRef] [Green Version]
- Zhang, M.; Coen, J.J.; Suzuki, Y.; Siedow, M.R.; Niemierko, A.; Khor, L.Y.; Pollack, A.; Zhang, Y.; Zietman, A.L.; Shipley, W.U.; et al. Survivin Is a Potential Mediator of Prostate Cancer Metastasis. Int. J. Radiat. Oncol. Biol. Phys. 2010, 78, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Azuhata, T.; Scott, D.; Takamizawa, S.; Wen, J.; Davidoff, A.; Fukuzawa, M.; Sandler, A. The Inhibitor of Apoptosis Protein Survivin Is Associated with High-Risk Behavior of Neuroblastoma. J. Pediatr. Surg. 2001, 36, 1785–1791. [Google Scholar] [CrossRef]
- Ito, R.; Asami, S.; Motohashi, S.; Ootsuka, S.; Yamaguchi, Y.; Chin, M.; Shichino, H.; Yoshida, Y.; Nemoto, N.; Mugishima, H.; et al. Significance of Survivin MRNA Expression in Prognosis of Neuroblastoma. Biol. Pharm. Bull. 2005, 28, 565–568. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Ye, Y.; Yang, D.; Lin, J. Survivin and Vascular Endothelial Growth Factor Are Associated with Spontaneous Pulmonary Metastasis of Osteosarcoma: Development of an Orthotopic Mouse Model. Oncol. Lett. 2014, 8, 2577–2580. [Google Scholar] [CrossRef] [Green Version]
- Ciudad, C.J.J.; Urlaub, G.; Chasin, L.A.A. Deletion Analysis of the Chinese Hamster Dihydrofolate Reductase Gene Promoter. J. Biol. Chem. 1988, 263, 16274–16282. [Google Scholar] [CrossRef] [PubMed]
- Vieira, J.; Messing, J. Production of Single-Stranded Plasmid DNA. Methods Enzymol. 1987, 153, 3–11. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | PPRH | Sequence 5′–3′ | Location |
|---|---|---|---|
| Survivin | HpsPr-C | AGGGGAGGGATGGAGTGCAG T T T AGGGGAGGGATGGAGTGCAG T T | Promoter |
| Survivin | FAM-HpsPr-C | [6FAM]AGGGGAGGGATGGAGTGCAG T T T AGGGGAGGGATGGAGTGCAG T T | Promoter |
| Scr-9 | FAM-HpScr9 | AAGAAGAAGAAGAGAAGAA T T T AAGAAGAAGAAGAGAAGAA T T | - |
| Cell Line | TROPY | DOTAP 10 µM | |||
|---|---|---|---|---|---|
| µg | Transfected Cells (%) | X-Mean | Transfected Cells (%) | X-Mean | |
| PC-3 | 1.0 | 89.2 | 40.2 | 69.4 | 64.4 |
| SH-SY5Y | 1.5 | 64.7 | 8.76 | n.d | n.d |
| MDA-MB-453 | 2.0 | 71.1 | 33.8 | n.d | n.d |
| HepG2 | 2.0 | 37.1 | 70.8 | 7.6 | 33.0 |
| SKBR-3 | 2.0 | 86.1 | 63.5 | 47.6 | 35.0 |
| VERO-E6 | 3.0 | 91.1 | 62.2 | 69.2 | 9.3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Delgado, A.; Griera, R.; Llor, N.; López-Aguilar, E.; Busquets, M.A.; Noé, V.; Ciudad, C.J. Trioleyl Pyridinium, a Cationic Transfection Agent for the Lipofection of Therapeutic Oligonucleotides into Mammalian Cells. Pharmaceutics 2023, 15, 420. https://doi.org/10.3390/pharmaceutics15020420
Delgado A, Griera R, Llor N, López-Aguilar E, Busquets MA, Noé V, Ciudad CJ. Trioleyl Pyridinium, a Cationic Transfection Agent for the Lipofection of Therapeutic Oligonucleotides into Mammalian Cells. Pharmaceutics. 2023; 15(2):420. https://doi.org/10.3390/pharmaceutics15020420
Chicago/Turabian StyleDelgado, Ana, Rosa Griera, Núria Llor, Ester López-Aguilar, Maria Antònia Busquets, Véronique Noé, and Carlos J. Ciudad. 2023. "Trioleyl Pyridinium, a Cationic Transfection Agent for the Lipofection of Therapeutic Oligonucleotides into Mammalian Cells" Pharmaceutics 15, no. 2: 420. https://doi.org/10.3390/pharmaceutics15020420