

Bacteriocin-Nanoconjugates (Bac10307-AgNPs) Biosynthesized from Lactobacillus acidophilus-Derived Bacteriocins Exhibit Enhanced and Promising Biological Activities

 ,
,  ,
,  ,
,  , , , , ,
, , , , ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial Strains and Growth Conditions
2.2. Extraction and Purification of Bacteriocin from L. acidophilus
2.3. Analysis of Antibacterial Activity of Partially Purified Bacteriocin-Bac10307
2.3.1. Bacterial Strains and Growth Conditions
2.3.2. Agar Well Diffusion Assay
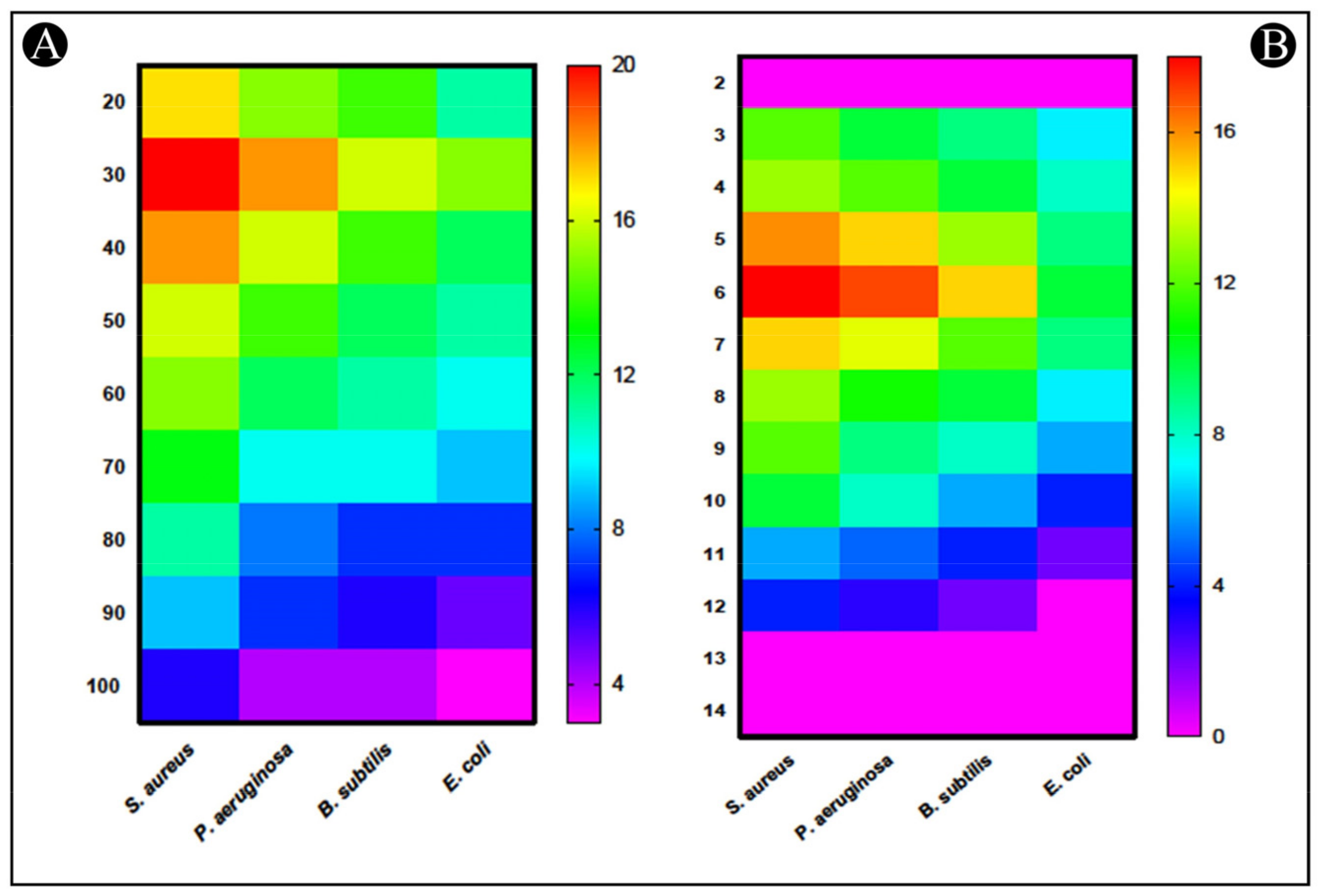
2.4. Assessment of Stability of Bacteriocin-Bac10307
2.4.1. Impact of Temperatures
2.4.2. Impact of pH
2.4.3. Impact of Enzymes
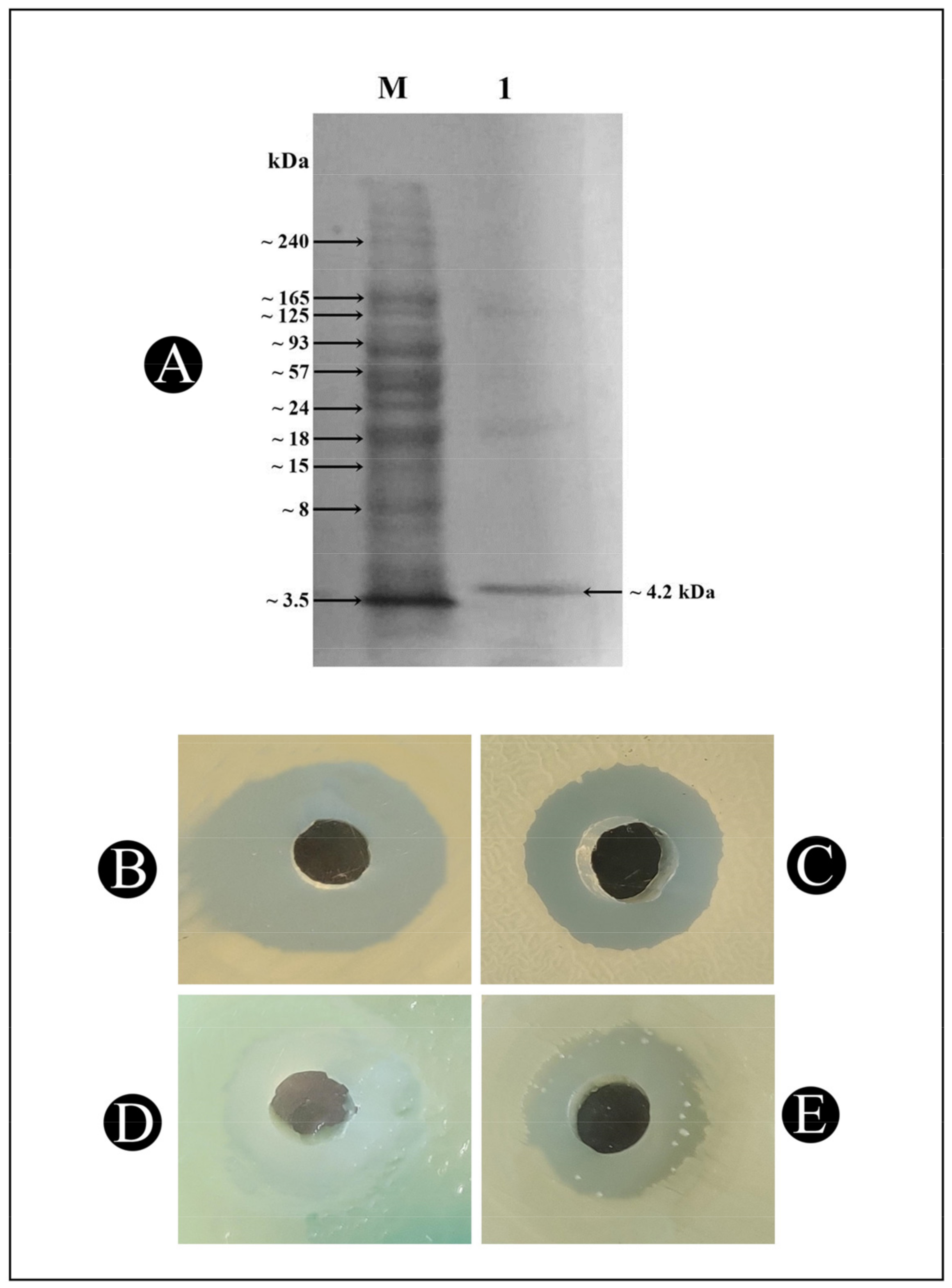
2.5. Molecular Mass Determination of Bacteriocin Bac10307 Using SDS–PAGE
2.6. Synthesis of Bacteriocin-10307-AgNPs
2.7. Characterization of Bac10307-AgNPs
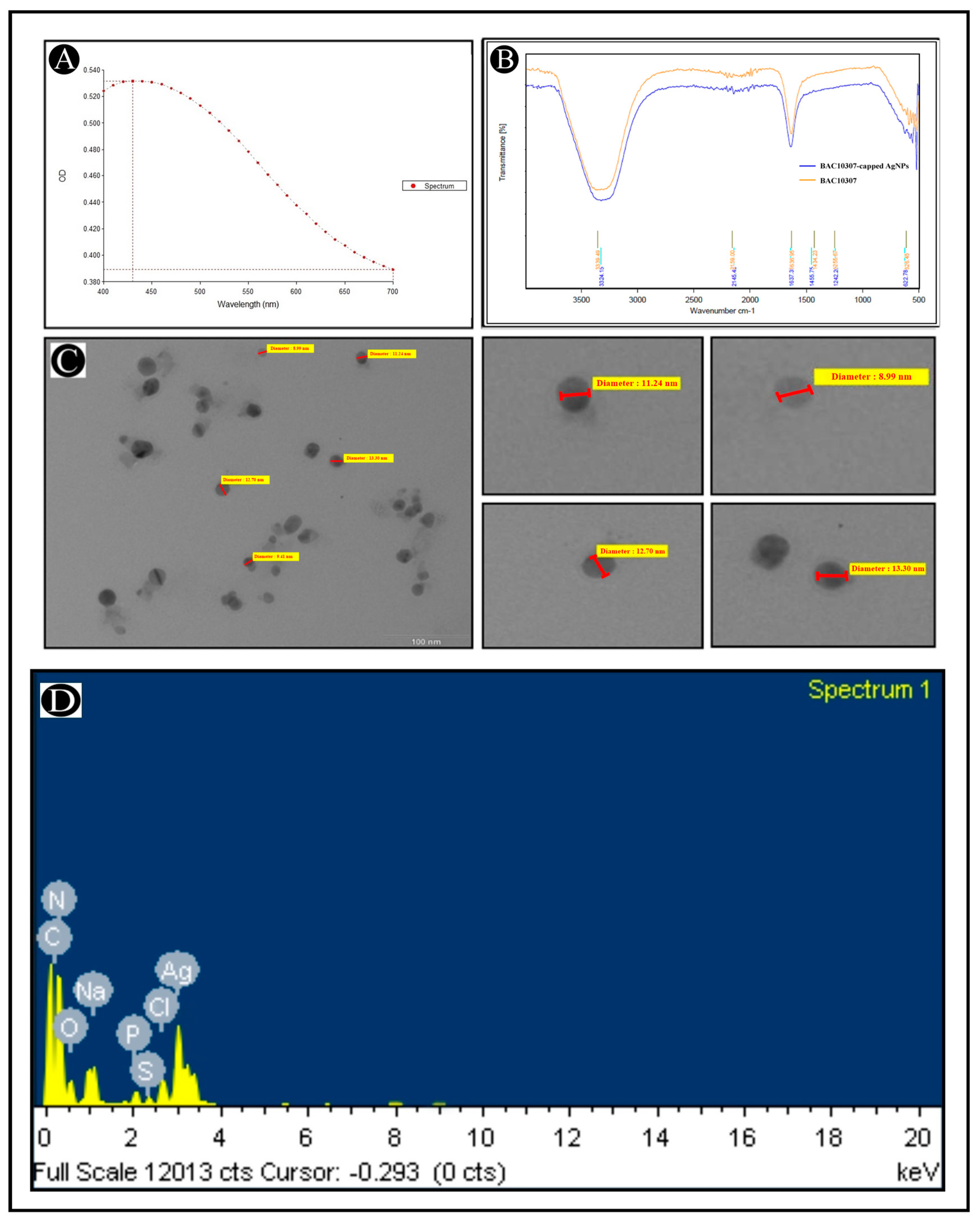
2.7.1. UV–Vis Spectrophotometer
2.7.2. Transmission Electron Microscopy (TEM) and Energy-Dispersive X-Ray (EDX)
2.7.3. Fourier-Transform Infrared (FT-IR) Analysis
2.8. Screening of Antibacterial Activity of Bac10307-AgNPs
2.8.1. Agar Well Diffusion Assay
2.8.2. Assessment of Minimum Inhibitory Concentration (MIC)
2.9. Screening of In Vitro Antioxidant Activity of Bac10307-AgNPs
2.10. Screening of In Vitro Cytotoxicity Assay (MTT Assay) of Bac10307-AgNPs
2.11. Molecular Docking (MD) Assays
2.12. Statistical Analysis
3. Results
3.1. Bacteriocin-Bac10307: Extraction, Purification, and Antibacterial Activity Analysis
3.2. Characterization of Purified Bacteriocin-Bac10307
3.3. Synthesis and Characterization of Bac10307-AgNPs
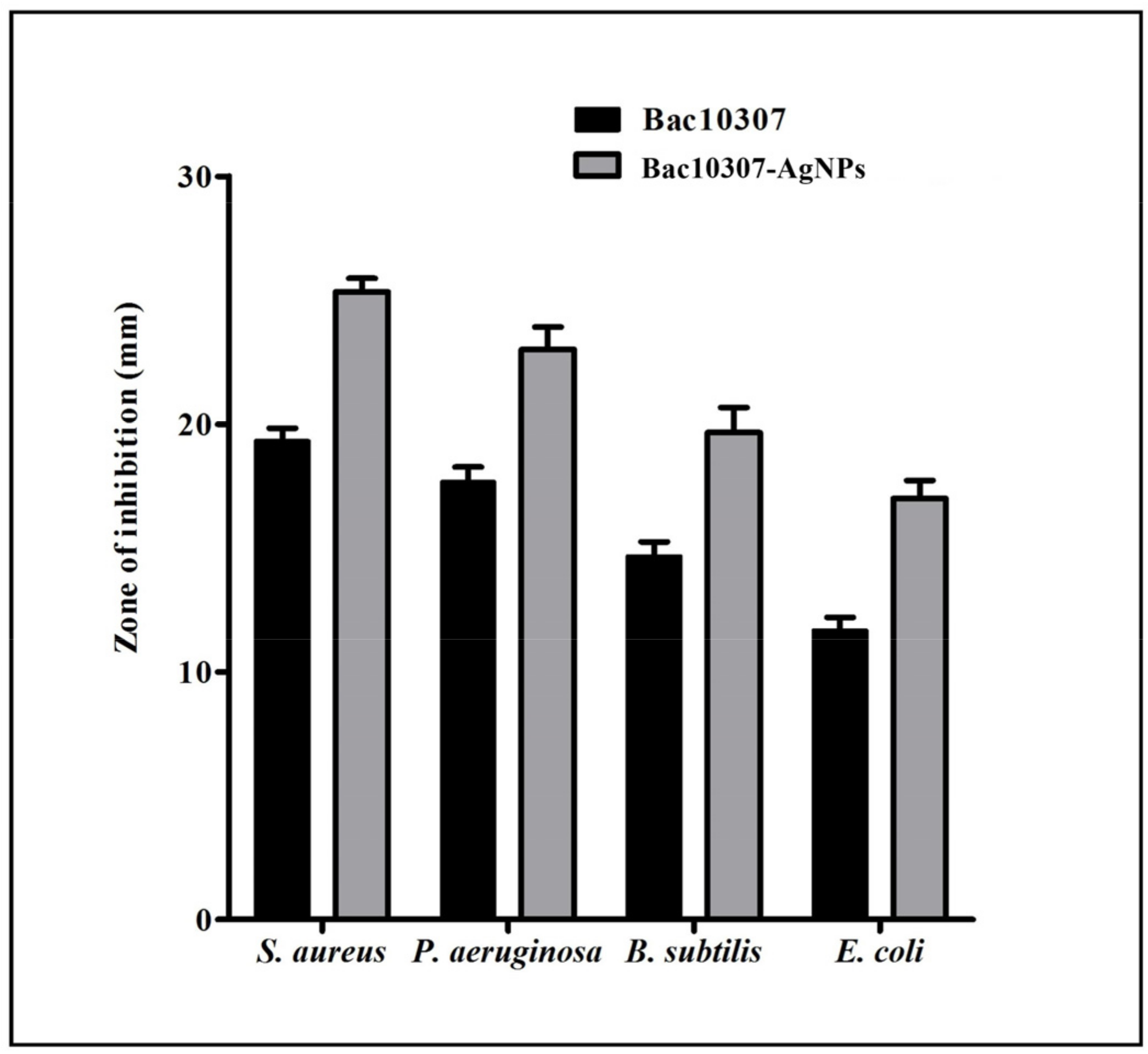
3.4. Antibacterial Activity of Bac10307 and Synthesized Bac10307-AgNPs
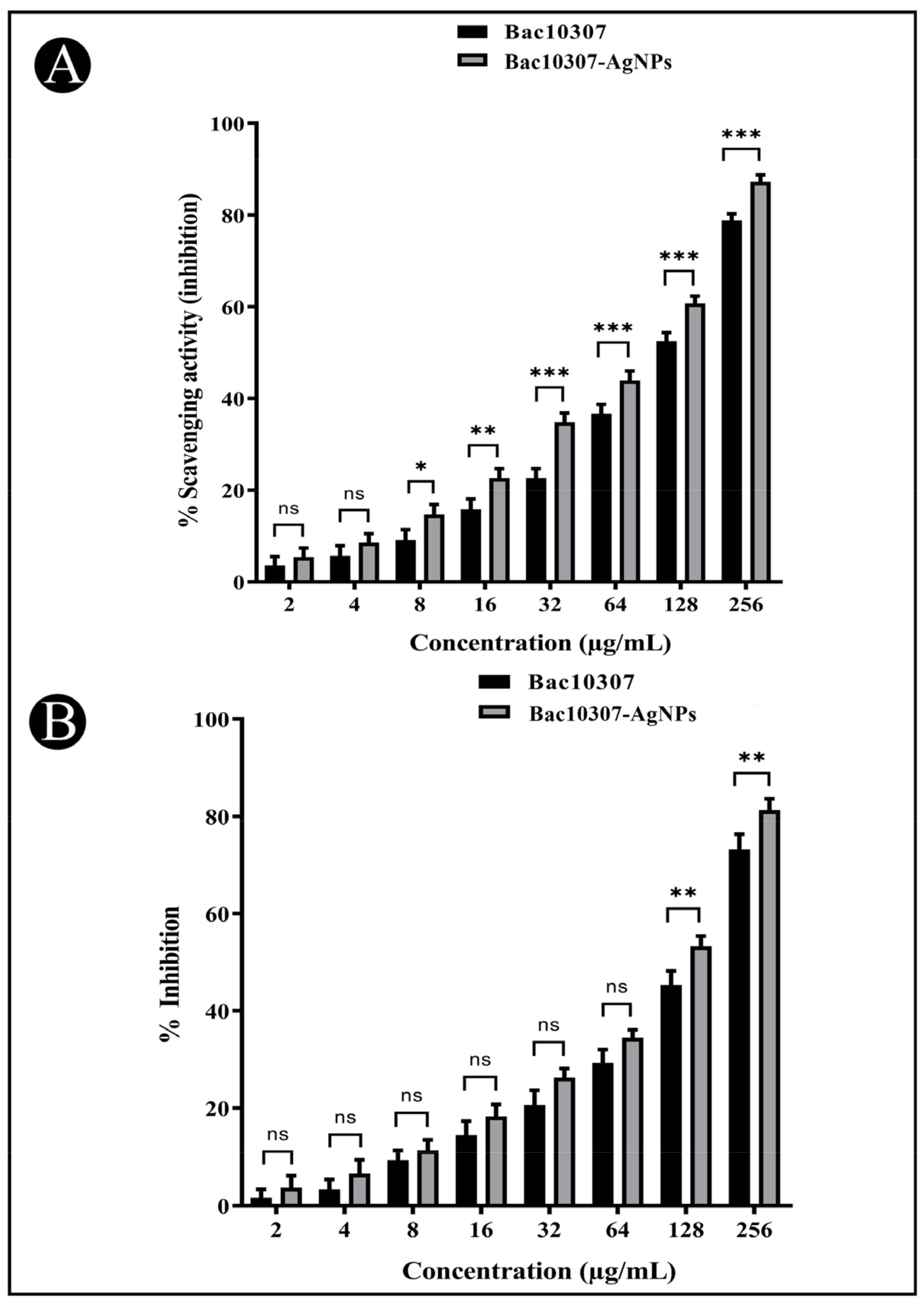
3.5. In Vitro Antioxidant Activity of Synthesized Bac10307-AgNPs
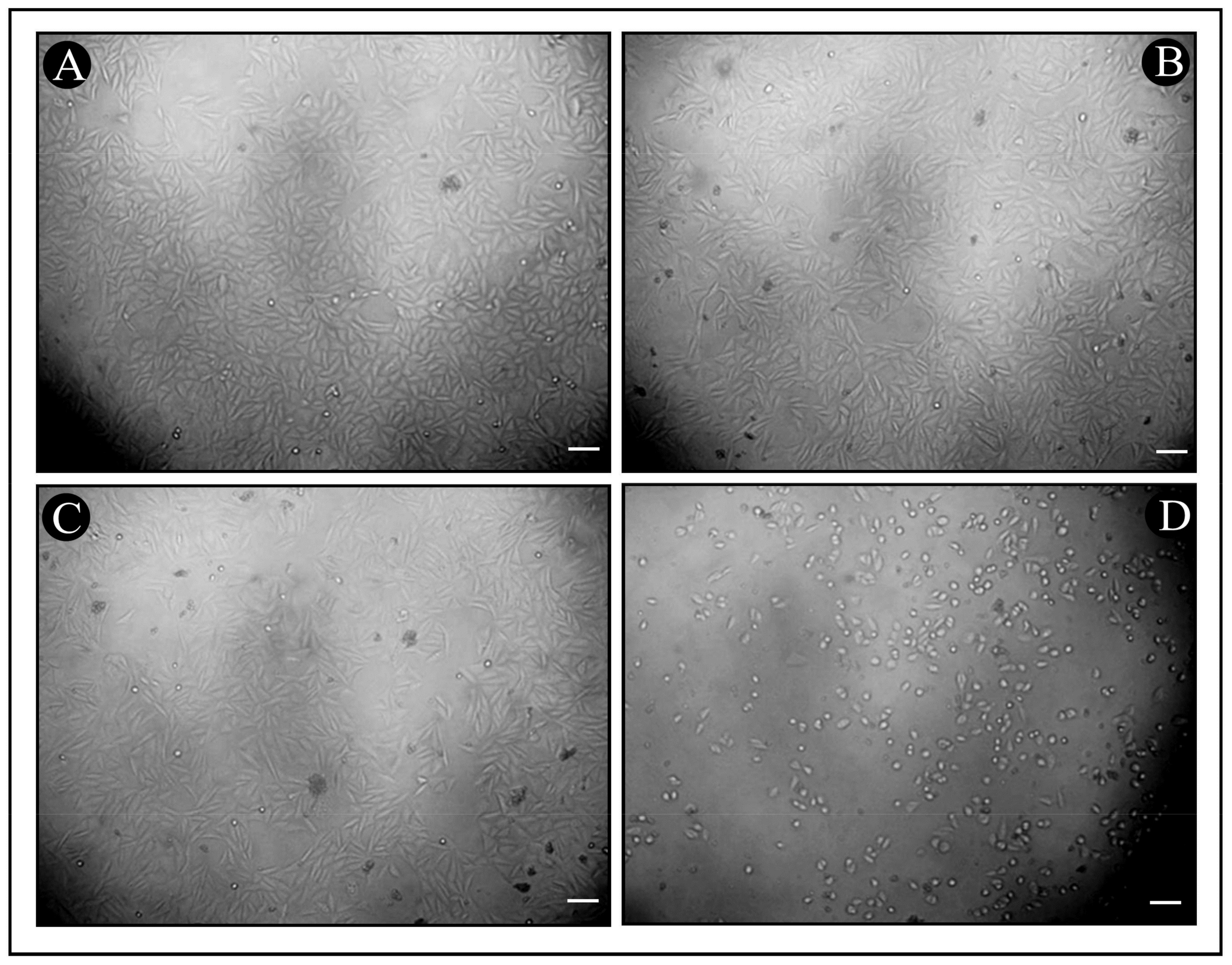
3.6. In Vitro Cytotoxicity of Synthesized Bac10307-AgNPs
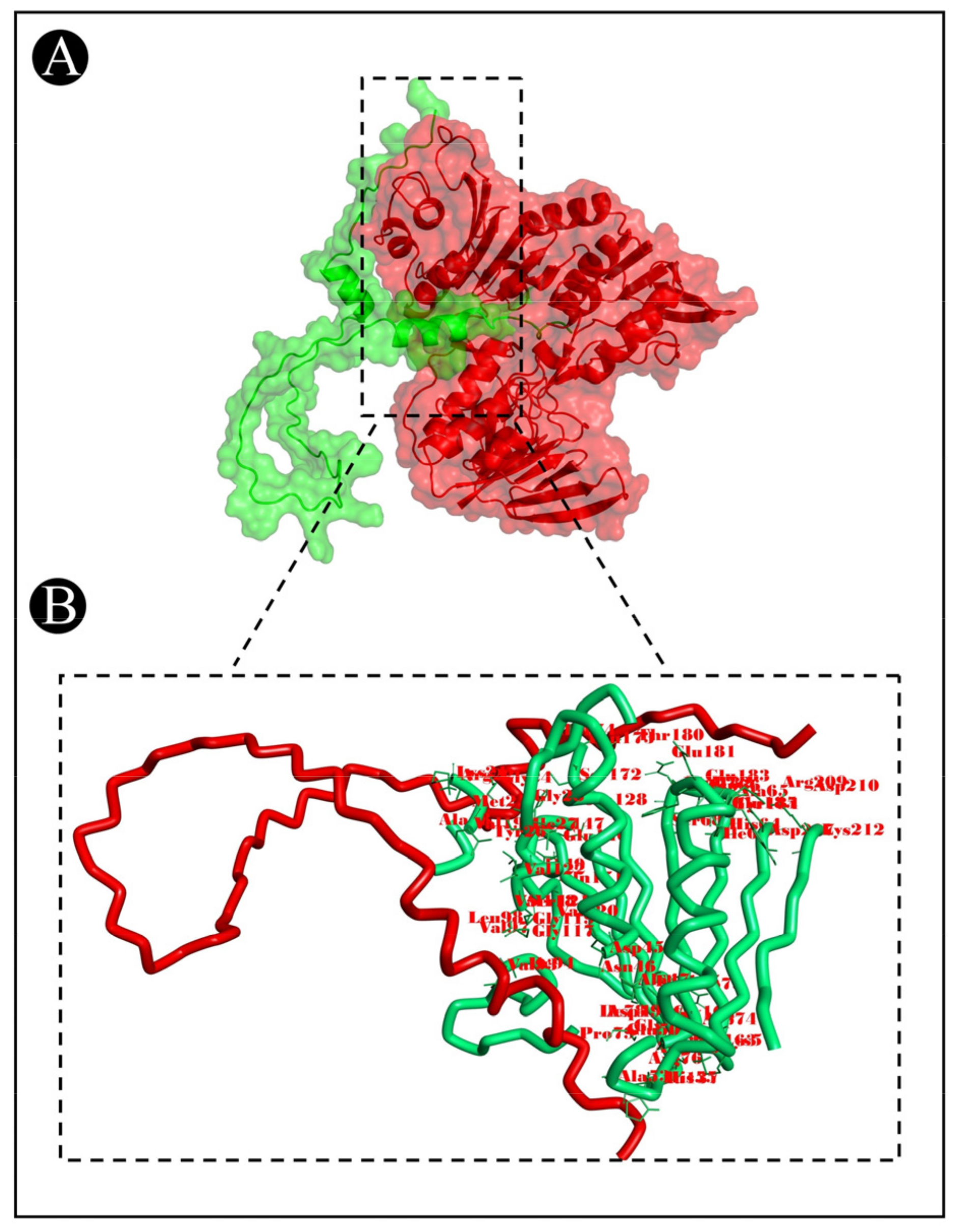
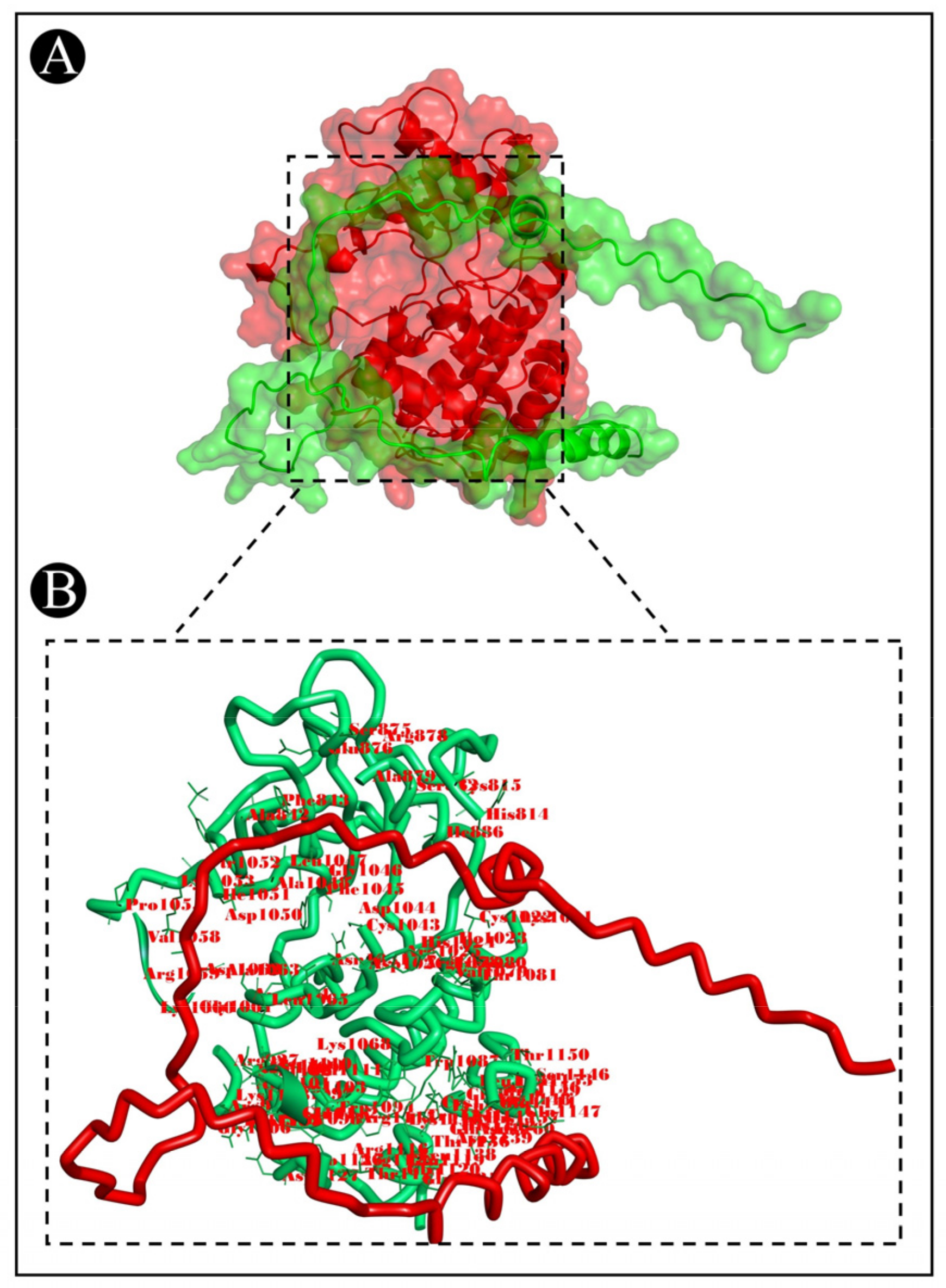
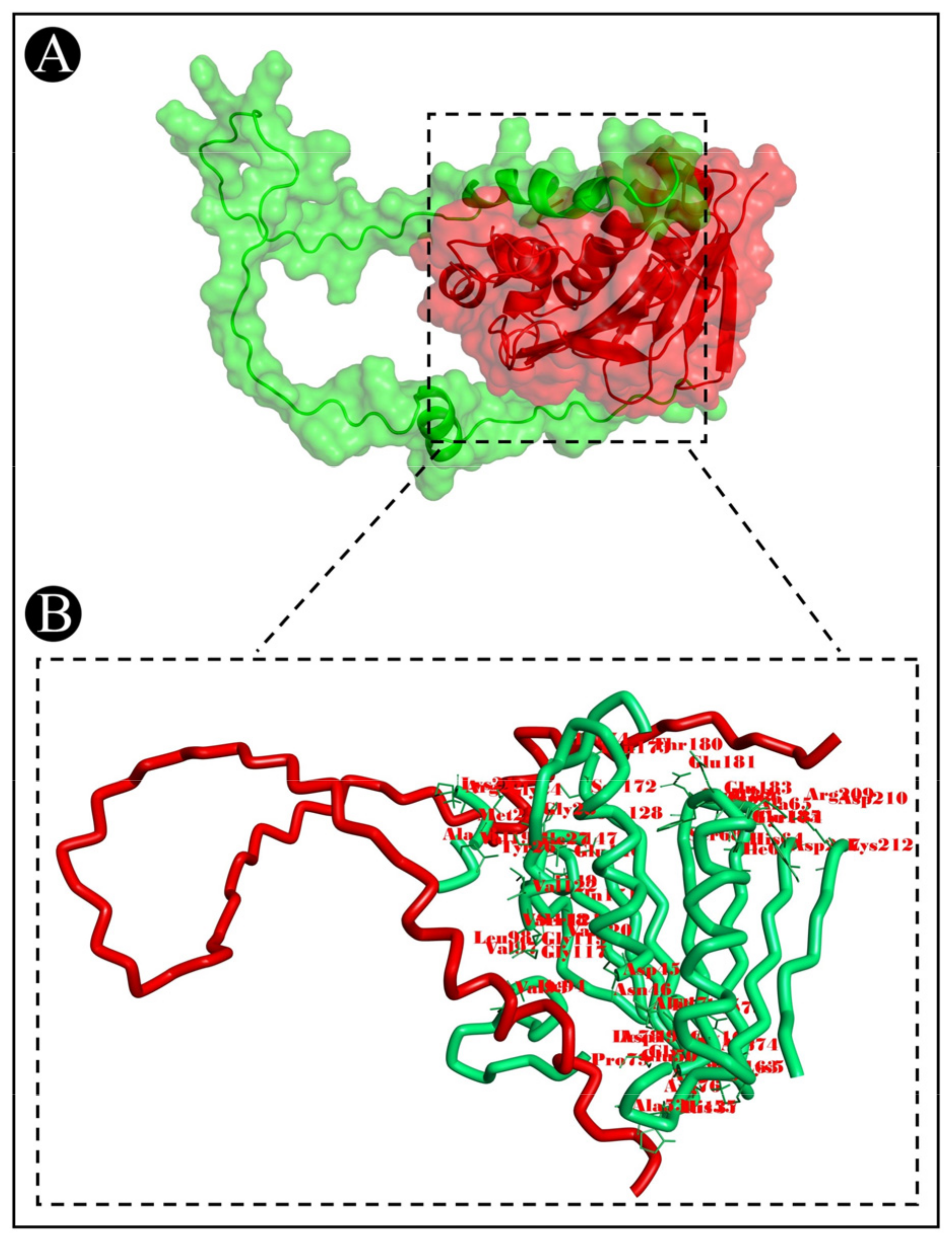
3.7. Molecular Docking Analysis
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Irkitova, A.N.; Matsyura, A.V. Ecological and biological characteristics of Lactobacillus acidophilus. Ukr. J. Ecol. 2017, 7, 214–230. [Google Scholar] [CrossRef] [PubMed]
- Villamil, L.; Reyes, C.; Martínez-Silva, M.A. In vivo and in vitro assessment of Lactobacillus acidophilus as probiotic for tilapia (Oreochromis niloticus, Perciformes: Cichlidae) culture improvement. Aquac. Res. 2014, 45, 1116–1125. [Google Scholar] [CrossRef]
- Remes-Troche, J.M.; Coss-Adame, E.; Valdovinos-Díaz, M.; Gómez-Escudero, O.; Icaza-Chávez, M.E.; Chávez-Barrera, J.A.; Zárate-Mondragón, F.; Velasco, J.A.V.-R.; Aceves-Tavares, G.R.; Lira-Pedrín, M.A.; et al. Lactobacillus acidophilus LB: A useful pharmabiotic for the treatment of digestive disorders. Ther. Adv. Gastroenterol. 2020, 13, 1756284820971201. [Google Scholar] [CrossRef] [PubMed]
- Gaspar, C.; Donders, G.G.; Palmeira-De-Oliveira, R.; Queiroz, J.A.; Tomaz, C.; Martinez-De-Oliveira, J. Bacteriocin production of the probiotic Lactobacillus acidophilus KS400. AMB Express 2018, 8, 153. [Google Scholar] [CrossRef]
- Tang, H.W.; Phapugrangkul, P.; Fauzi, H.M.; Tan, J.S. Lactic Acid Bacteria Bacteriocin, an Antimicrobial Peptide Effective Against Multidrug Resistance: A Comprehensive Review. Int. J. Pept. Res. Ther. 2022, 28, 14. [Google Scholar] [CrossRef]
- Silva, S.P.; Ribeiro, S.C.; Teixeira, J.A.; Silva, C.C. Application of an alginate-based edible coating with bacteriocin-producing Lactococcus strains in fresh cheese preservation. LWT 2022, 153, 112486. [Google Scholar] [CrossRef]
- Cotter, P.; Hill, C.; Ross, R. Bacteriocins: Developing innate immunity for food. Nat. Rev. Genet. 2005, 3, 777–788. [Google Scholar] [CrossRef]
- El-Gendy, A.O.; Essam, T.M.; Amin, M.A.; Ahmed, S.H.; Nes, I.F. Clinical Screening for Bacteriocinogenic Enterococcus faecalis Isolated from Intensive Care Unit Inpatient in Egypt. J. Microb. Biochem. Technol. 2013, 4, 161–167. [Google Scholar] [CrossRef] [Green Version]
- Martín, R.; Escobedo, S.; Martín, C.; Crespo, A.; Quiros, L.M.; Suarez, J.E. Chemotherapy, Surface glycosaminoglycans protect eukaryotic cells against membrane-driven peptide bacteriocins. Antimicrob. Agents Chemother. 2015, 59, 677–681. [Google Scholar] [CrossRef] [Green Version]
- Sidhu, P.K.; Nehra, K. Bacteriocin-nanoconjugates as emerging compounds for enhancing antimicrobial activity of bacteriocins. J. King Saud. Univ. 2019, 31, 758–767. [Google Scholar] [CrossRef]
- Ansari, A.; Zohra, R.R.; Tarar, O.M.; Qader, S.A.U.; Aman, A. Screening, purification and characterization of thermostable, protease resistant Bacteriocin active against methicillin resistant Staphylococcus aureus (MRSA). BMC Microbiol. 2018, 18, 192. [Google Scholar] [CrossRef]
- Patil, S.D.; Sharma, R.; Bhattacharyya, T.; Kumar, P.; Gupta, M.; Chaddha, B.S.; Navani, N.K.; Pathania, R. Antibacterial potential of a small peptide from Bacillus sp. RPT-0001 and its capping for green synthesis of silver nanoparticles. J. Microbiol. 2015, 53, 643–652. [Google Scholar] [CrossRef] [PubMed]
- Yang, E.; Fan, L.; Jiang, Y.; Doucette, C.; Fillmore, S. Antimicrobial activity of bacteriocin-producing lactic acid bacteria isolated from cheeses and yogurts. AMB Express 2012, 2, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patel, M.; Siddiqui, A.J.; Hamadou, W.S.; Surti, M.; Awadelkareem, A.M.; Ashraf, S.A.; Alreshidi, M.; Snoussi, M.; Rizvi, S.M.D.; Bardakci, F.; et al. Inhibition of Bacterial Adhesion and Antibiofilm Activities of a Glycolipid Biosurfactant from Lactobacillus rhamnosus with Its Physicochemical and Functional Properties. Antibiotics 2021, 10, 1546. [Google Scholar] [CrossRef]
- Wang, Y.; Shang, N.; Qin, Y.; Zhang, Y.; Zhang, J.; Li, P. The complete genome sequence of Lactobacillus plantarum LPL-1, a novel antibacterial probiotic producing class IIa bacteriocin. J. Biotechnol. 2018, 266, 84–88. [Google Scholar] [CrossRef] [PubMed]
- Elayaraja, S.; Annamalai, N.; Mayavu, P.; Balasubramanian, T. Production, purification and characterization of bacteriocin from Lactobacillus murinus AU06 and its broad antibacterial spectrum. Asian Pac. J. Trop. Biomed. 2014, 4, S305–S311. [Google Scholar] [CrossRef] [Green Version]
- Miao, J.; Guo, H.; Ou, Y.; Liu, G.; Fang, X.; Liao, Z.; Ke, C.; Chen, Y.; Zhao, L.; Cao, Y. Purification and characterization of bacteriocin F1, a novel bacteriocin produced by Lactobacillus paracasei subsp. tolerans FX-6 from Tibetan kefir, a traditional fermented milk from Tibet, China. Food Control 2014, 42, 48–53. [Google Scholar] [CrossRef]
- Zhang, J.; Yang, Y.; Yang, H.; Bu, Y.; Yi, H.; Zhang, L.; Han, X.; Ai, L. Purification and Partial Characterization of Bacteriocin Lac-B23, a Novel Bacteriocin Production by Lactobacillus plantarum J23, Isolated from Chinese Traditional Fermented Milk. Front. Microbiol. 2018, 9, 2165. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.U.; Nayab, H.; Rehman, T.U.; Williamson, M.P.; Haq, K.U.; Shafi, N.; Shafique, F. Characterisation of Bacteriocins Produced by Lactobacillus spp. Isolated from the Traditional Pakistani Yoghurt and Their Antimicrobial Activity against Common Foodborne Pathogens. BioMed Res. Int. 2020, 2020, 8281623. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, P.K.; Nehra, K. Purification and characterization of bacteriocin Bac23 extracted from Lactobacillus plantarum PKLP5 and its interaction with silver nanoparticles for enhanced antimicrobial spectrum against food-borne pathogens. LWT 2021, 139, 110546. [Google Scholar] [CrossRef]
- Adnan, M.; Patel, M.; Reddy, M.N.; Alshammari, E. Formulation, evaluation and bioactive potential of Xylaria primorskensis terpenoid nanoparticles from its major compound xylaranic acid. Sci. Rep. 2018, 8, 1740. [Google Scholar] [CrossRef] [Green Version]
- Awadelkareem, A.M.; Al-Shammari, E.; Elkhalifa, A.O.; Adnan, M.; Siddiqui, A.J.; Patel, M.; Khan, M.I.; Mehmood, K.; Ashfaq, F.; Badraoui, R. Biosynthesized Silver Nanoparticles from Eruca sativa Miller Leaf Extract Exhibits Antibacterial, Antioxidant, Anti-Quorum-Sensing, Antibiofilm, and Anti-Metastatic Activities. Antibiotics 2022, 11, 853. [Google Scholar] [CrossRef]
- Adnan, M.; Siddiqui, A.J.; Ashraf, S.A.; Snoussi, M.; Badraoui, R.; Alreshidi, M.; Elasbali, A.M.; Al-Soud, W.A.; Alharethi, S.H.; Sachidanandan, M. Polyhydroxybutyrate (PHB)-Based Biodegradable Polymer from Agromyces indicus: Enhanced Production, Characterization, and Optimization. Polymers 2022, 14, 3982. [Google Scholar] [CrossRef] [PubMed]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Awadelkareem, A.M.; Al-Shammari, E.; Elkhalifa, A.E.O.; Adnan, M.; Siddiqui, A.J.; Snoussi, M.; Khan, M.I.; Azad, Z.R.A.A.; Patel, M.; Ashraf, S.A. Phytochemical and In Silico ADME/Tox Analysis of Eruca sativa Extract with Antioxidant, Antibacterial and Anticancer Potential against Caco-2 and HCT-116 Colorectal Carcinoma Cell Lines. Molecules 2022, 27, 1409. [Google Scholar] [CrossRef]
- Reddy, M.N.; Adnan, M.; Alreshidi, M.M.; Saeed, M.; Patel, M. Evaluation of Anticancer, Antibacterial and Antioxidant Properties of a Medicinally Treasured Fern Tectaria coadunata with its Phytoconstituents Analysis by HR-LCMS. Anti-Cancer Agents Med. Chem. 2020, 20, 1845–1856. [Google Scholar] [CrossRef] [PubMed]
- Kozakov, D.; Beglov, D.; Bohnuud, T.; Mottarella, S.E.; Xia, B.; Hall, D.R.; Vajda, S. How good is automated protein docking? Proteins Struct. Funct. Bioinform. 2013, 81, 2159–2166. [Google Scholar] [CrossRef] [Green Version]
- Kozakov, D.; Hall, D.R.; Xia, B.; Porter, K.A.; Padhorny, D.; Yueh, C.; Beglov, D.; Vajda, S. The ClusPro web server for protein–protein docking. Nat. Protoc. 2017, 12, 255–278. [Google Scholar] [CrossRef] [PubMed]
- Desta, I.T.; Porter, K.A.; Xia, B.; Kozakov, D.; Vajda, S. Performance and Its Limits in Rigid Body Protein-Protein Docking. Structure 2020, 28, 1071–1081.e3. [Google Scholar] [CrossRef]
- Lagos, R.; Tello, M.; Mercado, G.; García, V.; Monasterio, O. Antibacterial and antitumorigenic properties of microcin E492, a pore-forming bacteriocin. Curr. Pharm. Biotechnol. 2009, 10, 74–85. [Google Scholar] [CrossRef]
- Guzmán-Rodríguez, J.J.; Ochoa-Zarzosa, A.; López-Gómez, R.; López-Meza, J.E. Plant Antimicrobial Peptides as Potential Anticancer Agents. BioMed Res. Int. 2015, 2015, 735087. [Google Scholar] [CrossRef] [Green Version]
- Deslouches, B.; Steckbeck, J.D.; Craigo, J.K.; Doi, Y.; Burns, J.L.; Montelaro, R.C. Engineered Cationic Antimicrobial Peptides To Overcome Multidrug Resistance by ESKAPE Pathogens. Antimicrob. Agents Chemother. 2015, 59, 1329–1333. [Google Scholar] [CrossRef] [Green Version]
- Agrawal, S.; Acharya, D.; Adholeya, A.; Barrow, C.; Deshmukh, S.K. Nonribosomal Peptides from Marine Microbes and Their Antimicrobial and Anticancer Potential. Front. Pharmacol. 2017, 8, 828. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Chen, D.; Wang, L.; Lin, C.; Ma, C.; Xi, X.; Chen, T.; Shaw, C.; Zhou, M. Dermaseptin-PH: A Novel Peptide with Antimicrobial and Anticancer Activities from the Skin Secretion of the South American Orange-Legged Leaf Frog, Pithecopus (Phyllomedusa) hypochondrialis. Molecules 2017, 22, 1805. [Google Scholar] [CrossRef] [PubMed]
- Delves-Brougthon, J. Nisin and its uses as a food preservative. Food Technol. 1990, 44, 100–117. [Google Scholar]
- Bradshaw, J.P. Cationic antimicrobial peptides. BioDrugs 2003, 17, 233–240. [Google Scholar] [CrossRef]
- Schilliager, U.; Geisen, R.; Holzapfel, W.H. Potential of antagonistic microganisms and bacteriocins for the biological preservation of food. Trends Food Sci Tech. 1997, 7, 158–164. [Google Scholar] [CrossRef]
- Davidson, P.M.; Sofos, J.N.; Branen, A.L. Antimicrobials in Food; CRC Press: Boca Raton, FL, USA, 2005. [Google Scholar]
- Riley, M.A.; Wertz, J.E. Bacteriocins: Evolution, Ecology, and Application. Annu. Rev. Microbiol. 2002, 56, 117–137. [Google Scholar] [CrossRef] [Green Version]
- Saraniya, A.; Jeevaratnam, K. Optimization of nutritional and non-nutritional factors involved for production of antimicrobial compounds from Lactobacillus pentosus SJ65 using response surface methodology. Braz. J. Microbiol. 2014, 45, 81–88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Salmaso, S.; Elvassore, N.; Bertucco, A.; Lante, A.; Caliceti, P. Nisin-loaded poly-l-lactide nano-particles produced by CO2 anti-solvent precipitation for sustained antimicrobial activity. Int. J. Pharm. 2004, 287, 163–173. [Google Scholar] [CrossRef]
- De Giani, A.; Bovio, F.; Forcella, M.; Fusi, P.; Sello, G.; Di Gennaro, P. Identification of a bacteriocin-like compound from Lactobacillus plantarum with antimicrobial activity and effects on normal and cancerogenic human intestinal cells. AMB Express 2019, 9, 88. [Google Scholar] [CrossRef] [PubMed]
- Danilova, T.A.; Adzhieva, A.A.; Danilina, G.A.; Polyakov, N.B.; Soloviev, A.I.; Zhukhovitsky, V.G. Antimicrobial Activity of Supernatant of Lactobacillus plantarum against Pathogenic Microorganisms. Bull. Exp. Biol. Med. 2019, 167, 751–754. [Google Scholar] [CrossRef]
- Abo-Amer, A.E. Characterization of a bacteriocin-like inhibitory substance produced by Lactobacillus plantarum isolated from Egyptian home-made yogurt. Sci. Asia. 2007, 33, 313–319. [Google Scholar] [CrossRef]
- Djadouni, F.; Kihal, M. Characterization and determination of the factors affecting anti-listerial bacteriocins from Lactobacillus plantarum and Pediococcus pentosaceus isolated from dairy milk products. Afr. J. Food Sci. 2013, 7, 35–44. [Google Scholar]
- Moghaddam, M.Z.; Sattari, M.; Mobarez, A.M.; Doctorzadeh, F. Inhibitory effect of yogurt Lactobacilli bacteriocins on growth and verotoxins production of enterohemorrhgic Escherichia coli O157: H7. Pak. J. Biol. Sci. 2006, 9, 2112–2116. [Google Scholar] [CrossRef] [Green Version]
- Milioni, C.; Martínez, B.; Degl’Innocenti, S.; Turchi, B.; Fratini, F.; Cerri, D.; Fischetti, R. A novel bacteriocin produced by Lactobacillus plantarum LpU4 as a valuable candidate for biopreservation in artisanal raw milk cheese. Dairy Sci. Technol. 2015, 95, 479–494. [Google Scholar] [CrossRef]
- Song, J.Y.; Kim, B.S. Rapid biological synthesis of silver nanoparticles using plant leaf extracts. Bioproc. Biosyst. Eng. 2009, 32, 79–84. [Google Scholar] [CrossRef]
- Hata, T.; Tanaka, R.; Ohmomo, S. Isolation and characterization of plantaricin ASM1: A new bacteriocin produced by Lactobacillus plantarum A-1. Int. J. Food Microbiol. 2010, 137, 94–99. [Google Scholar] [CrossRef]
- Atrih, A.; Rekhif, N.; Moir, A.; Lebrihi, A.; Lefebvre, G. Mode of action, purification and amino acid sequence of plantaricin C19, an anti-Listeria bacteriocin produced by Lactobacillus plantarum C19. Int. J. Food Microbiol. 2001, 68, 93–104. [Google Scholar] [CrossRef] [PubMed]
- Hu, M.; Zhao, H.; Zhang, C.; Yu, J.; Lu, Z. Purification and Characterization of Plantaricin 163, a Novel Bacteriocin Produced by Lactobacillus plantarum 163 Isolated from Traditional Chinese Fermented Vegetables. J. Agric. Food Chem. 2013, 61, 11676–11682. [Google Scholar] [CrossRef]
- Chen, Y.-S.; Wang, Y.-C.; Chow, Y.-S.; Yanagida, F.; Liao, C.-C.; Chiu, C.-M. Purification and characterization of plantaricin Y, a novel bacteriocin produced by Lactobacillus plantarum 510. Arch. Microbiol. 2014, 196, 193–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Qin, Y.; Zhang, Y.; Wu, R.; Li, P. Antibacterial mechanism of plantaricin LPL-1, a novel class IIa bacteriocin against Listeria monocytogenes. Food Control 2019, 97, 87–93. [Google Scholar] [CrossRef]
- Mills, S.; Ross, R.; Hill, C. Bacteriocins and bacteriophage; a narrow-minded approach to food and gut microbiology. FEMS Microbiol. Rev. 2017, 41, S129–S153. [Google Scholar] [CrossRef] [PubMed]
- Lazzari, S.; Moscatelli, D.; Codari, F.; Salmona, M.; Morbidelli, M.; Diomede, L. Colloidal stability of polymeric nanoparticles in biological fluids. J. Nanoparticle Res. 2012, 14, 920. [Google Scholar] [CrossRef] [Green Version]
- Niaz, T.; Shabbir, S.; Noor, T.; Imran, M. Antimicrobial and antibiofilm potential of bacteriocin loaded nano-vesicles functionalized with rhamnolipids against foodborne pathogens. LWT 2019, 116, 108583. [Google Scholar] [CrossRef]
- Kumar, C.G.; Mamidyala, S.K. Extracellular synthesis of silver nanoparticles using culture supernatant of Pseudomonas aeruginosa. Colloids Surf. B Biointerfaces 2011, 84, 462–466. [Google Scholar] [CrossRef]
- Saravana, K.; Annalakshmi, A. Enhancing the antimicrobial activity of nisin by encapsulating on silver nanoparticle synthesized by Bacillus sp. Int. J. Pharma. Biol. Arch. 2012, 3, 406–410. [Google Scholar]
- Gomaa, E.Z. Synergistic Antibacterial Efficiency of Bacteriocin and Silver Nanoparticles Produced by Probiotic Lactobacillus paracasei Against Multidrug Resistant Bacteria. Int. J. Pept. Res. Ther. 2019, 25, 1113–1125. [Google Scholar] [CrossRef]
- Thirumurugan, A.; Ramachandran, S.; Gowri, A.S. Combined effect of bacteriocin with gold nanoparticles against food spoiling bacteria-an approach for food packaging material preparation. Int. Food Res. J. 2013, 20, 1909–1912. [Google Scholar]
- Singh, A.K.; Bai, X.; Amalaradjou, M.A.R.; Bhunia, A.K. Antilisterial and Antibiofilm Activities of Pediocin and LAP Functionalized Gold Nanoparticles. Front. Sustain. Food Syst. 2018, 2, 74. [Google Scholar] [CrossRef] [Green Version]
- Morales-Avila, E.; Ferro-Flores, G.; Ocampo-García, B.E.; López-Téllez, G.; López-Ortega, J.; Rogel-Ayala, D.G.; Sánchez-Padilla, D. Antibacterial Efficacy of Gold and Silver Nanoparticles Functionalized with the Ubiquicidin (29–41) Antimicrobial Peptide. J. Nanomater. 2017, 2017, 5831959. [Google Scholar] [CrossRef] [Green Version]
- Elkhalifa, A.E.O.; Al-Shammari, E.; Alam, M.J.; Alcantara, J.C.; Khan, M.A.; Eltoum, N.E.; Ashraf, S.A. Okra-derived dietary Carotenoid lutein against Breast Cancer, with an Approach towards Developing a Nutraceutical Product: A Meta-analysis Study. J. Pharm. Res. Int. 2021, 33, 135–142. [Google Scholar] [CrossRef]
- Cao, W.; Chen, H.-D.; Yu, Y.-W.; Li, N.; Chen, W.-Q. Changing profiles of cancer burden worldwide and in China: A secondary analysis of the global cancer statistics 2020. Chin. Med. J. 2021, 134, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.-C.; Lin, H.-H.; Hsu, C.-H.; Wang, C.-J.; Chiang, T.-A.; Chen, J.-H. Inhibitory effects of andrographolide on migration and invasion in human non-small cell lung cancer A549 cells via down-regulation of PI3K/Akt signaling pathway. Eur. J. Pharmacol. 2010, 632, 23–32. [Google Scholar] [CrossRef] [PubMed]
- Loza-Mejía, M.A.; Salazar, J.R.; Sánchez-Tejeda, J.F. In Silico Studies on Compounds Derived from Calceolaria: Phenylethanoid Glycosides as Potential Multitarget Inhibitors for the Development of Pesticides. Biomolecules 2018, 8, 121. [Google Scholar] [CrossRef] [Green Version]
- Lee, K.; Kim, D. In-Silico Molecular Binding Prediction for Human Drug Targets Using Deep Neural Multi-Task Learning. Genes 2019, 10, 906. [Google Scholar] [CrossRef] [Green Version]
- Bharathi, A.; Roopan, S.M.; Vasavi, C.S.; Munusami, P.; Gayathri, G.A.; Gayathri, M. In silico molecular docking and in vitro antidiabetic studies of dihydropyrimido [4,5-a] acridin-2-amines. BioMed Res. Int. 2014, 2014, 971569. [Google Scholar] [CrossRef] [Green Version]
- Comeau, S.R.; Gatchell, D.W.; Vajda, S.; Camacho, C.J. ClusPro: A fully automated algorithm for protein-protein docking. Nucleic Acids Res. 2004, 32, W96–W99. [Google Scholar] [CrossRef] [Green Version]
- Xue, L.C.; Dobbs, D.; Honavar, V. HomPPI: A class of sequence homology based protein-protein interface prediction methods. BMC Bioinform. 2011, 12, 244. [Google Scholar] [CrossRef] [Green Version]
- Hao, T.; Peng, W.; Wang, Q.; Wang, B.; Sun, J. Reconstruction and Application of Protein–Protein Interaction Network. Int. J. Mol. Sci. 2016, 17, 907. [Google Scholar] [CrossRef] [Green Version]
- Alfonso, E.E.; Deng, Z.; Boaretto, D.; Hood, B.L.; Vasile, S.; Smith, L.H.; Chambers, J.W.; Chapagain, P.; Leng, F. Novel and Structurally Diversified Bacterial DNA Gyrase Inhibitors Discovered through a Fluorescence-Based High-Throughput Screening Assay. ACS Pharmacol. Transl. Sci. 2022, 5, 932–944. [Google Scholar] [CrossRef] [PubMed]
- Suh, Y.-A.; Arnold, R.S.; Lassegue, B.; Shi, J.; Xu, X.X.; Sorescu, D.; Chung, A.B.; Griendling, K.K.; Lambeth, J.D. Cell transformation by the superoxide-generating oxidase Mox1. Nat. Cell Biol. 1999, 401, 79–82. [Google Scholar] [CrossRef]
- Dupuy, C.; Ohayon, R.; Valent, A.; Noël-Hudson, M.-S.; Dème, D.; Virion, A. Purification of a Novel Flavoprotein Involved in the Thyroid NADPH Oxidase. J. Biol. Chem. 1999, 274, 37265–37269. [Google Scholar] [CrossRef] [Green Version]
- Bedard, K.; Krause, K.-H. The NOX Family of ROS-Generating NADPH Oxidases: Physiology and Pathophysiology. Physiol. Rev. 2007, 87, 245–313. [Google Scholar] [CrossRef] [PubMed]
- Gao, H.-M.; Zhou, H.; Hong, J.-S. NADPH oxidases: Novel therapeutic targets for neurodegenerative diseases. Trends Pharmacol. Sci. 2012, 33, 295–303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simons, M.; Gordon, E.; Claesson-Welsh, L. Mechanisms and regulation of endothelial VEGF receptor signalling. Nat. Rev. Mol. Cell Biol. 2016, 17, 611–625. [Google Scholar] [CrossRef]
- Yu, J.; Zhang, Y.; Leung, L.-H.; Liu, L.; Yang, F.; Yao, X. Efficacy and safety of angiogenesis inhibitors in advanced gastric cancer: A systematic review and meta-analysis. J. Hematol. Oncol. 2016, 9, 111. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Test Organisms | Zone of Inhibition (mm) | MIC (μg/mL) Bac10307-AgNPs | MBC (μg/mL) Bac10307-AgNPs | ||
|---|---|---|---|---|---|
| Bac10307 | AgNPs | Bac10307- AgNPs | |||
| S. aureus | 19.66 | 11.66 | 25.33 | 8 | 16 |
| P. aeruginosa | 17.33 | 10.33 | 23.00 | 8 | 16 |
| B. subtilis | 14.66 | 8.33 | 19.66 | 16 | 32 |
| E. coli | 11.66 | 7.66 | 17.00 | 64 | 128 |
| Proteins | Receptor-Ligand | Interection Type | Distance |
|---|---|---|---|
| 2CDU-Acidocin A | A:LYS78:HZ2–A:ASP282:OD2 | Salt Bridge;Attractive Charge | 1.76041 |
| A:LYS78:HZ3–A:ASP282:OD1 | Salt Bridge;Attractive Charge | 1.75112 | |
| A:CYS133:HN–A:HIS81:NE2 | Conventional Hydrogen Bond | 2.43145 | |
| A:ILE160:HN–A:PHE80:O | Conventional Hydrogen Bond | 2.31372 | |
| A:GLY244:HN–A:HIS81:O | Conventional Hydrogen Bond | 1.88916 | |
| A:ALA300:HN–A:PHE76:O | Conventional Hydrogen Bond | 2.81082 | |
| A:THR301:HN–A:GLY74:O | Conventional Hydrogen Bond | 2.55908 | |
| A:ARG305:HH11–A:TRP71:O | Conventional Hydrogen Bond | 1.81072 | |
| A:ARG305:HH21–A:TRP71:O | Conventional Hydrogen Bond | 1.84157 | |
| A:ARG431:HH12–A:SER3:O | Conventional Hydrogen Bond | 2.70789 | |
| A:PHE433:HN–A:ILE5:O | Conventional Hydrogen Bond | 2.04607 | |
| A:SER3:HN–A:GLU366:OE1 | Conventional Hydrogen Bond | 2.29836 | |
| A:SER7:HG–A:PRO432:O | Conventional Hydrogen Bond | 1.84543 | |
| A:GLN9:HE22–A:ASP422:OD1 | Conventional Hydrogen Bond | 2.12445 | |
| A:THR79:HN–A:PRO298:O | Conventional Hydrogen Bond | 2.73643 | |
| A:THR79:HG1–A:PRO298:O | Conventional Hydrogen Bond | 1.85808 | |
| A:SER41:CB–A:GLY77:O | Conventional Hydrogen Bond | 3.70135 | |
| A:ARG431:CD–A:SER3:O | Carbon Hydrogen Bond | 3.15632 | |
| A:HIS81:CE1–A:LEU241:O | Carbon Hydrogen Bond | 3.13598 | |
| A:LYS187:NZ–A:PHE80 | Pi-Cation | 3.51826 | |
| A:ARG305:NH1–A:TRP71 | Pi-Cation | 4.77402 | |
| A:SER7:CA–A:PHE433 | Pi-Sigma | 3.67277 | |
| A:ILE243:C,O;GLY244:N–A:PHE80 | Amide-Pi Stacked | 3.93853 | |
| A:ALA300–A:LYS78 | Alkyl | 4.9093 | |
| A:ILE438–A:ILE5 | Alkyl | 4.68217 | |
| A:LYS10–A:MET420 | Alkyl | 4.61562 | |
| A:PHE367–A:ILE2 | Pi-Alkyl | 5.07639 | |
| A:TRP71–A:VAL304 | Pi-Alkyl | 5.08256 | |
| A:TRP71–A:ARG305 | Pi-Alkyl | 4.70863 | |
| A:TRP71–A:ARG305 | Pi-Alkyl | 4.06273 | |
| A:TRP71–A:ARG308 | Pi-Alkyl | 5.17719 | |
| A:PHE76–A:LEU330 | Pi-Alkyl | 4.80412 | |
| A:HIS81–A:PRO117 | Pi-Alkyl | 4.38714 | |
| A:HIS81–A:LEU132 | Pi-Alkyl | 5.41817 | |
| 6f86-Acidocin A | A:MET1:N–A:ASP210:OD2 | Attractive Charge | 5.4936 |
| A:ARG22:HH11–A:LEU59:O | Conventional Hydrogen Bond | 1.77015 | |
| A:ARG22:HH21–A:SER58:OG | Conventional Hydrogen Bond | 1.72382 | |
| A:TYR26:HH–A:ASP64:OD2 | Conventional Hydrogen Bond | 1.97086 | |
| A:ASN46:HD22–A:ALA72:O | Conventional Hydrogen Bond | 1.98784 | |
| A:ASN46:HD22–A:THR73:O | Conventional Hydrogen Bond | 2.63274 | |
| A:ARG76:HH12–A:GLY77:O | Conventional Hydrogen Bond | 1.8161 | |
| A:ARG136:HH11–A:PHE76:O | Conventional Hydrogen Bond | 2.53417 | |
| A:ARG136:HH11–A:LYS78:O | Conventional Hydrogen Bond | 2.17681 | |
| A:ARG136:HH21–A:LYS78:O | Conventional Hydrogen Bond | 1.72262 | |
| A:THR180:HG1–A:SER7:OG | Conventional Hydrogen Bond | 1.87972 | |
| A:SER3:HN–A:ARG209:O | Conventional Hydrogen Bond | 2.36379 | |
| A:SER3:HG–A:ARG209:O | Conventional Hydrogen Bond | 1.86756 | |
| A:GLN9:HE21–A:GLU174:OE2 | Conventional Hydrogen Bond | 2.01259 | |
| A:LYS10:HN–A:GLN128:OE1 | Conventional Hydrogen Bond | 2.17849 | |
| A:ALA18:HN–A:ASP14:O | Conventional Hydrogen Bond | 1.93332 | |
| A:ALA18:HN–A:LYS15:O | Conventional Hydrogen Bond | 2.65481 | |
| A:SER21:HN–A:ALA18:O | Conventional Hydrogen Bond | 2.35624 | |
| A:GLY23:HN–A:ALA18:O | Conventional Hydrogen Bond | 1.89654 | |
| A:LYS24:HZ3–A:TYR26:OH | Conventional Hydrogen Bond | 1.63416 | |
| A:TYR26:HN–A:LYS24:O | Conventional Hydrogen Bond | 2.39531 | |
| A:LYS62:HZ1–A:TYR26:O | Conventional Hydrogen Bond | 2.61388 | |
| A:LYS62:HZ2–A:TYR26:O | Conventional Hydrogen Bond | 2.68718 | |
| A:LYS62:HZ3–A:MET25:O | Conventional Hydrogen Bond | 1.66846 | |
| A:LEU68:HN–A:LEU98:O | Conventional Hydrogen Bond | 2.64427 | |
| A:THR73:HG1–A:ASP49:OD1 | Conventional Hydrogen Bond | 1.98276 | |
| A:THR79:HG1–A:ARG76:O | Conventional Hydrogen Bond | 1.81635 | |
| A:ARG76:CD–A:GLY77:O | Carbon Hydrogen Bond | 3.35609 | |
| A:MET4:CA–A:GLU181:OE2 | Carbon Hydrogen Bond | 3.26235 | |
| A:TRP71:CD1–A:VAL97:O | Carbon Hydrogen Bond | 3.75115 | |
| A:GLU50:OE1–A:PHE76 | Pi-Anion | 3.4452 | |
| A:ALA18–A:ILE61 | Alkyl | 4.4781 | |
| A:VAL118–A:LEU65 | Alkyl | 4.41069 | |
| A:LYS62–A:MET25 | Alkyl | 5.37497 | |
| A:ALA72–A:VAL120 | Alkyl | 5.07682 | |
| A:ALA75–A:PRO79 | Alkyl | 5.06784 | |
| A:ALA75–A:ILE94 | Alkyl | 4.19361 | |
| A:TYR26–A:LYS62 | Pi-Alkyl | 4.48685 | |
| A:TYR26–A:LYS24 | Pi-Alkyl | 4.99044 | |
| A:TRP71–A:VAL97 | Pi-Alkyl | 4.62974 | |
| A:TRP71–A:VAL97 | Pi-Alkyl | 5.0319 | |
| A:PHE76–A:ILE78 | Pi-Alkyl | 5.21439 | |
| A:PHE80–A:ARG76 | Pi-Alkyl | 4.69087 | |
| 2OH4-Acidocin A | A:ARG1078:HH11–A:ASP14:OD1 | Salt Bridge;Attractive Charge | 1.77815 |
| A:ARG1122:HH22–A:ASP64:OD2 | Salt Bridge;Attractive Charge | 1.82447 | |
| A:LYS24:HZ1–A:ASP1044:OD1 | Salt Bridge;Attractive Charge | 2.0004 | |
| A:LYS24:HZ3–A:ASP1044:OD1 | Salt Bridge;Attractive Charge | 2.03938 | |
| A:ARG55:HH21–A:GLU1112:OE2 | Salt Bridge;Attractive Charge | 1.89818 | |
| A:LYS56:HZ1–A:GLU1111:OE1 | Salt Bridge;Attractive Charge | 1.69962 | |
| A:LYS56:HZ3–A:GLU1111:OE2 | Salt Bridge;Attractive Charge | 1.80997 | |
| A:LYS78:HZ3–A:GLU1156:OE2 | Salt Bridge;Attractive Charge | 1.71364 | |
| A:ARG1078:NH2–A:ASP14:OD2 | Attractive Charge | 2.75179 | |
| A:ARG1122:NH1–A:ASP64:OD1 | Attractive Charge | 4.71658 | |
| A:ARG55:NH1–A:ASP1110:OD2 | Attractive Charge | 5.55384 | |
| A:ARG55:NH1–A:GLU1112:OE1 | Attractive Charge | 2.72487 | |
| A:ARG878:HH11–A:TYR27:OH | Conventional Hydrogen Bond | 1.773 | |
| A:ARG927:HH11–A:SER38:O | Conventional Hydrogen Bond | 3.0835 | |
| A:ARG927:HH21–A:SER38:O | Conventional Hydrogen Bond | 1.74776 | |
| A:ILE1023:HN–A:SER21:O | Conventional Hydrogen Bond | 1.98064 | |
| A:ARG1025:HH11–A:SER21:O | Conventional Hydrogen Bond | 1.79986 | |
| A:ARG1025:HH21–A:SER21:OG | Conventional Hydrogen Bond | 1.96334 | |
| A:ARG1025:HH21–A:GLY22:O | Conventional Hydrogen Bond | 2.07187 | |
| A:LYS1060:HZ2–A:LYS36:O | Conventional Hydrogen Bond | 1.63582 | |
| A:GLY1061:HN–A:THR35:O | Conventional Hydrogen Bond | 2.45869 | |
| A:ALA1063:HN–A:THR35:OG1 | Conventional Hydrogen Bond | 2.01064 | |
| A:LYS1108:HZ1–A:ILE49:O | Conventional Hydrogen Bond | 2.21003 | |
| A:LYS1108:HZ3–A:GLY51:O | Conventional Hydrogen Bond | 1.71447 | |
| A:ARG1115:HH12–A:LYS56:O | Conventional Hydrogen Bond | 1.81567 | |
| A:ARG1116:HH11–A:GLN57:OE1 | Conventional Hydrogen Bond | 1.89055 | |
| A:ARG1116:HH21–A:GLN57:OE1 | Conventional Hydrogen Bond | 1.9005 | |
| A:ARG1116:HH22–A:SER58:O | Conventional Hydrogen Bond | 2.01369 | |
| A:ARG1122:HE–A:ASP64:OD1 | Conventional Hydrogen Bond | 2.45807 | |
| A:GLY1143:HN–A:ASP64:OD2 | Conventional Hydrogen Bond | 2.04712 | |
| A:LYS24:HZ1–A:HIS1024:NE2 | Conventional Hydrogen Bond | 2.30731 | |
| A:LYS24:HZ2–A:HIS1024:O | Conventional Hydrogen Bond | 1.64274 | |
| A:HIS33:HN–A:ASP1050:O | Conventional Hydrogen Bond | 2.89181 | |
| A:THR35:HG1–A:ARG1059:O | Conventional Hydrogen Bond | 2.54504 | |
| A:LYS36:HZ1–A:VAL1107:O | Conventional Hydrogen Bond | 1.69429 | |
| A:ARG55:HN–A:GLU1112:OE1 | Conventional Hydrogen Bond | 2.65589 | |
| A:LYS56:HN–A:GLU1112:OE1 | Conventional Hydrogen Bond | 2.47201 | |
| A:GLN57:HN–A:GLU1112:OE1 | Conventional Hydrogen Bond | 1.99761 | |
| A:SER58:HG–A:GLU1119:OE2 | Conventional Hydrogen Bond | 1.94121 | |
| A:LEU59:HN–A:GLU1119:OE1 | Conventional Hydrogen Bond | 2.04219 | |
| A:ILE61:HN–A:GLU1119:O | Conventional Hydrogen Bond | 1.97082 | |
| A:GLN63:HE21–A:GLY1120:O | Conventional Hydrogen Bond | 2.03591 | |
| A:LEU68:HN–A:ASP1139:OD2 | Conventional Hydrogen Bond | 2.48216 | |
| A:LYS78:HZ1–A:GLN1163:OE1 | Conventional Hydrogen Bond | 1.98774 | |
| A:LYS78:HZ2–A:ASN1160:OD1 | Conventional Hydrogen Bond | 1.69053 | |
| A:SER58:CA–A:GLU1119:OE1 | Carbon Hydrogen Bond | 2.9455 | |
| A:LYS24:NZ–A:HIS1024 | Pi-Cation | 3.6145 | |
| A:ASP64:OD2–A:HIS1142 | Pi-Anion | 4.79084 | |
| A:VAL1058:CG1–A:HIS33 | Pi-Sigma | 3.82332 | |
| A:LYS1021–A:ILE20 | Alkyl | 4.85838 | |
| A:ALA1101–A:LEU39 | Alkyl | 4.05593 | |
| A:PRO1149–A:LEU68 | Alkyl | 5.33682 | |
| A:HIS1142–A:LEU65 | Pi-Alkyl | 5.30386 | |
| A:TYR27–A:ALA879 | Pi-Alkyl | 4.63121 | |
| A:TRP40–A:PRO1105 | Pi-Alkyl | 4.91592 | |
| A:TRP71–A:PRO1149 | Pi-Alkyl | 4.42062 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Siddiqui, A.J.; Patel, M.; Adnan, M.; Jahan, S.; Saxena, J.; Alshahrani, M.M.; Abdelgadir, A.; Bardakci, F.; Sachidanandan, M.; Badraoui, R.; et al. Bacteriocin-Nanoconjugates (Bac10307-AgNPs) Biosynthesized from Lactobacillus acidophilus-Derived Bacteriocins Exhibit Enhanced and Promising Biological Activities. Pharmaceutics 2023, 15, 403. https://doi.org/10.3390/pharmaceutics15020403
Siddiqui AJ, Patel M, Adnan M, Jahan S, Saxena J, Alshahrani MM, Abdelgadir A, Bardakci F, Sachidanandan M, Badraoui R, et al. Bacteriocin-Nanoconjugates (Bac10307-AgNPs) Biosynthesized from Lactobacillus acidophilus-Derived Bacteriocins Exhibit Enhanced and Promising Biological Activities. Pharmaceutics. 2023; 15(2):403. https://doi.org/10.3390/pharmaceutics15020403
Chicago/Turabian StyleSiddiqui, Arif Jamal, Mitesh Patel, Mohd Adnan, Sadaf Jahan, Juhi Saxena, Mohammed Merae Alshahrani, Abdelmushin Abdelgadir, Fevzi Bardakci, Manojkumar Sachidanandan, Riadh Badraoui, and et al. 2023. "Bacteriocin-Nanoconjugates (Bac10307-AgNPs) Biosynthesized from Lactobacillus acidophilus-Derived Bacteriocins Exhibit Enhanced and Promising Biological Activities" Pharmaceutics 15, no. 2: 403. https://doi.org/10.3390/pharmaceutics15020403