
Bio-Inspired Drug Delivery Systems: From Synthetic Polypeptide Vesicles to Outer Membrane Vesicles
by

Yu Zhang
1,
Yiming Lu
1,
Yixin Xu
1,
Zunkang Zhou
2,
Yichong Li
2,
Wei Ling
2 and
Wenliang Song
2,* 1
School of Pharmacy, Shanghai University of Medicine & Health Sciences, Shanghai 201318, China
2
School of Materials and Chemistry, University of Shanghai for Science and Technology, Shanghai 200093, China
*
Author to whom correspondence should be addressed.
Pharmaceutics 2023, 15(2), 368; https://doi.org/10.3390/pharmaceutics15020368
Submission received: 4 December 2022
/
Revised: 10 January 2023
/
Accepted: 19 January 2023
/
Published: 21 January 2023
(This article belongs to the Special Issue Advances of Membrane Vesicles in Drug Delivery Systems)
Abstract
:Nanomedicine is a broad field that focuses on the development of nanocarriers to deliver specific drugs to targeted sites. A synthetic polypeptide is a kind of biomaterial composed of repeating amino acid units that are linked by peptide bonds. The multiplied amphiphilicity segment of the polypeptide could assemble to form polypeptide vesicles (PVs) under suitable conditions. Different from polypeptide vesicles, outer membrane vesicles (OMVs) are spherical buds of the outer membrane filled with periplasmic content, which commonly originate from Gram-negative bacteria. Owing to their biodegradability and excellent biocompatibility, both PVs and OMVs have been utilized as carriers in delivering drugs. In this review, we discuss the recent drug delivery research based on PVs and OMVs. These related topics are presented: (1) a brief introduction to the production methods for PVs and OMVs; (2) a thorough explanation of PV- and OMV-related applications in drug delivery including the vesicle design and biological assessment; (3) finally, we conclude with a discussion on perspectives and future challenges related to the drug delivery systems of PVs and OMVs.
1. Introduction
Pharmacologically active agents (also known as therapeutics or drugs) are used in medicine to control or stop the progression of disease [1,2]. Improved delivery methods and a better understanding of how drug administration affects safety and efficacy have been thoroughly discussed [3,4]. Drug delivery systems (DDSs) are technological systems that comprehensively regulate the distribution of drugs in the organism in terms of space, time, and dose [5,6,7,8,9,10]. The goal is to deliver the right amount of drug to the right place at the right time, thus increasing the efficiency of drug utilization, improving efficacy, reducing costs, and reducing toxic side effects [11,12]. To date, DDS usage has seen widespread integration in synthetic chemistry, materials science, biology, and medical sciences and is becoming increasingly prevalent in clinical practice. However, drug delivery targeting is not easily controlled because the release rate, the targeted sites, and the stability of the carrier and drug are inconsistent in different environments. Understanding the drug delivery characteristics of specific vesicles is a significant basis for their future application.
Polypeptides are crucial, artificially prepared biomaterials [13,14]. Their outstanding biocompatibility, appropriate biodegradability, and biological chain design abilities allow polypeptides to be employed in a variety of applications, such as drug delivery, antibacterial applications, and gene delivery [14,15]. In addition, unique secondary structures of polypeptides, such as α-helices and β-sheets, endow them with interesting self-assembly behavior and biological properties. Generally, three approaches can be applied to the synthesis of polypeptide materials, including solid-phase synthesis, α-amino acid N-carboxyanhydride (NCA) polymerization, and microbial fermentation [16]. Among these, the ring-opening polymerization (ROP) of NCAs represents a facile route to obtain high-molecular-weight polymers with the desired functionality. Due to the variety of ROP chemical and synthetic strategies, a polypeptide not only can be conjugated with multiple polypeptide segments, but it can also be blocked with non-peptidic materials. The peptides mentioned in this review are mainly synthetic polypeptides, which are biomaterials composed of repeating amino acid units linked by peptide bonds. In particular, amphiphilic polypeptides, which are composed of hydrophobic and hydrophilic chains, can easily self-assemble into micelles under suitable conditions. The vesicle is the most interesting assembled morphology of micelles. Further, the designed pH-responsive residues could be used as candidates to load hydrophilic cargos and be released upon appropriate stimulation. A subtle tactic to reduce unintended toxicity and increase effectiveness is the design of smart chemistry methods for programmed release in DDSs. Polymers with a variety of specific functionalities that react to internal or external stimuli and cause payloads to be released at the target site are needed. As a result, controlled release using vesicles, which is otherwise challenging to achieve in delivery systems based on other biopolymers, can be made possible by tuning the built-in molecular structures of polypeptide segments.
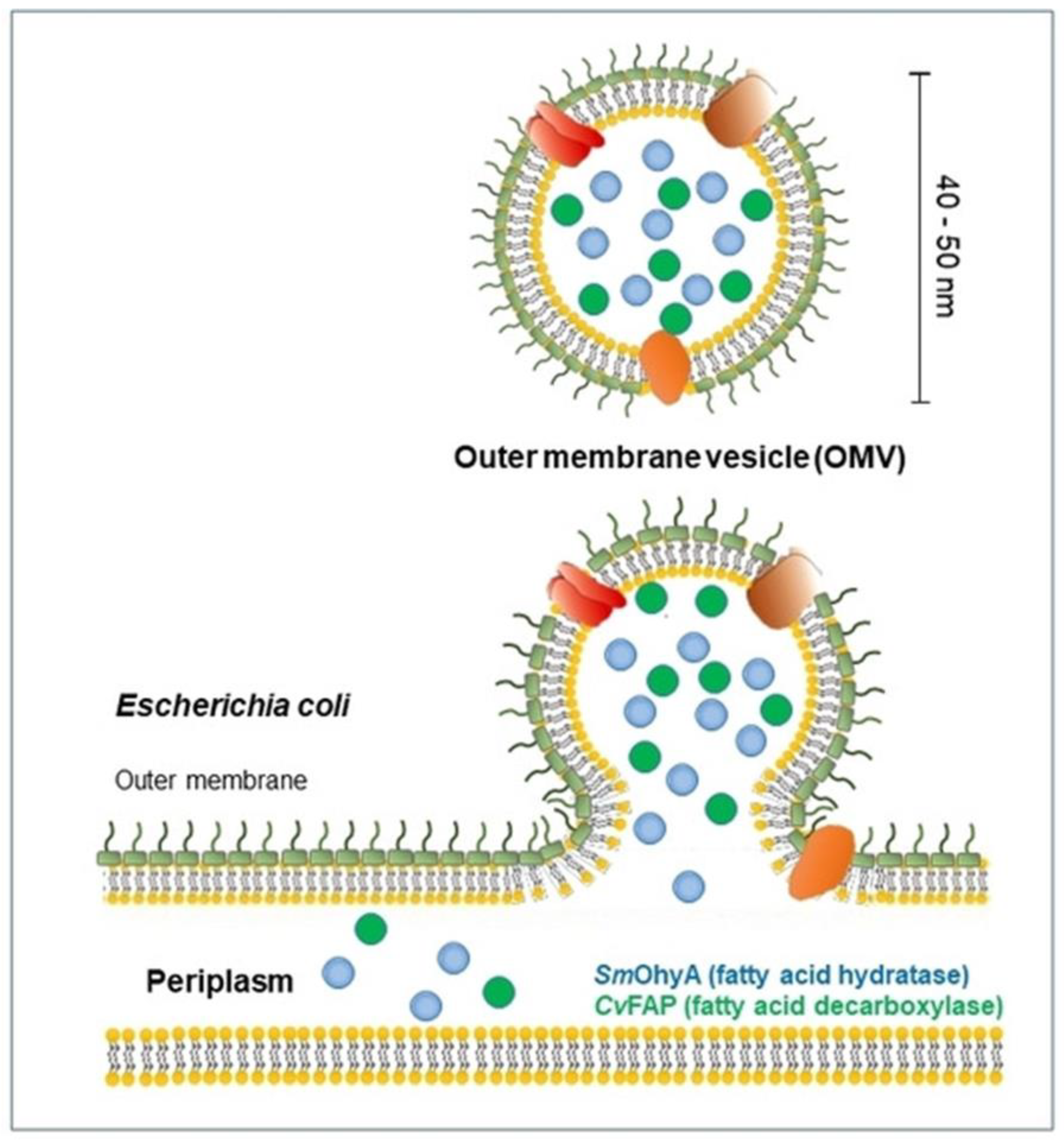
Different from artificially assembled polypeptide vesicles, vesicles derived from the outer membrane of Gram-negative bacteria are called OMVs [17,18,19,20]. OMVs are widely produced, vary in size and composition, and contain soluble periplasmic content. Given the diversity of OMV-producing pathways, species-dependent differences in the envelope architecture, and environmental influences on envelope composition, it is difficult to identify a single molecular or genetic basis for OMV production. OMVs were first reported more than 50 years ago in research on bacterial growth and were described to be able to release vesicles with diameters of 20 to 300 nm [21]. Since the outer membrane of the original bacteria was studied, several bacteria have been reported to be capable of producing OMVs, such as Pseudomonas aeruginosa, Helicobacter pylori, Escherichia coli, Shigella sp., Borrelia burgdorferi, Vibrio sp., Neisseria sp., etc. [22,23,24,25,26]. As they are derived from the membrane, OMVs mainly contain phospholipids, lipopolysaccharides, outer membrane proteins, and periplasm, along with different cytoplasmic components. These vesicles have been connected to several physiological activities because of their composition, including protein transport, nutrition acquisition, cell-to-cell communication, antibacterial activity, toxin delivery, and host immune response regulation. In addition, differently from the clear ROP polymerization and self-assembly mechanism, the mechanism of OMVs remains a mystery. Recently, owing to some gene and biochemistry studies, some clues about this complex process have been reported. In recent years, it has been found that OMVs could be used in antigens for vaccines, drug delivery, bioimaging, nanogenerators, and new adjuvants. Studies have also revealed that OMVs have great potential in the transport of small-molecule drugs. Further, it has been demonstrated how to electroporate medications into the OMV lumen. Finally, passive diffusion can be employed for tiny molecules that are generally hydrophobic and positively charged, so that they can easily interact with lipophilic membranes (Table 1).
In this review, first, we highlight recent achievements in research on artificial PVs and natural OMVs (Figure 1). The production methods of PVs and OMVs are summarized. Then, the comparison of the fabrication of PV- and OMV-based DDSs and especially their biological evaluation are extensively discussed. In the end, the current challenges regarding PVs and OMVs are also presented.
2. Production and Characterization of PVs and OMVs
2.1. Production of PVs
In polypeptide clinical transformation, the precise control of chemical structures and the polydispersity index (PDI) is very important for polymer vesicle formation. In recent decades, continued efforts have been made to produce polypeptides with controlled molecular weight, narrow PDI, peptide segments, and physical–chemical biological effects [13,29]. However, solid-phase peptide synthesis and microbial fermentation have been characterized by low yield, high costs, and limited achievable chain length. Herein, we mainly focus on the polymerization method for obtaining polypeptides via the ROP of α-amino acid NCAs and the vesicles subsequently assembled in solution.
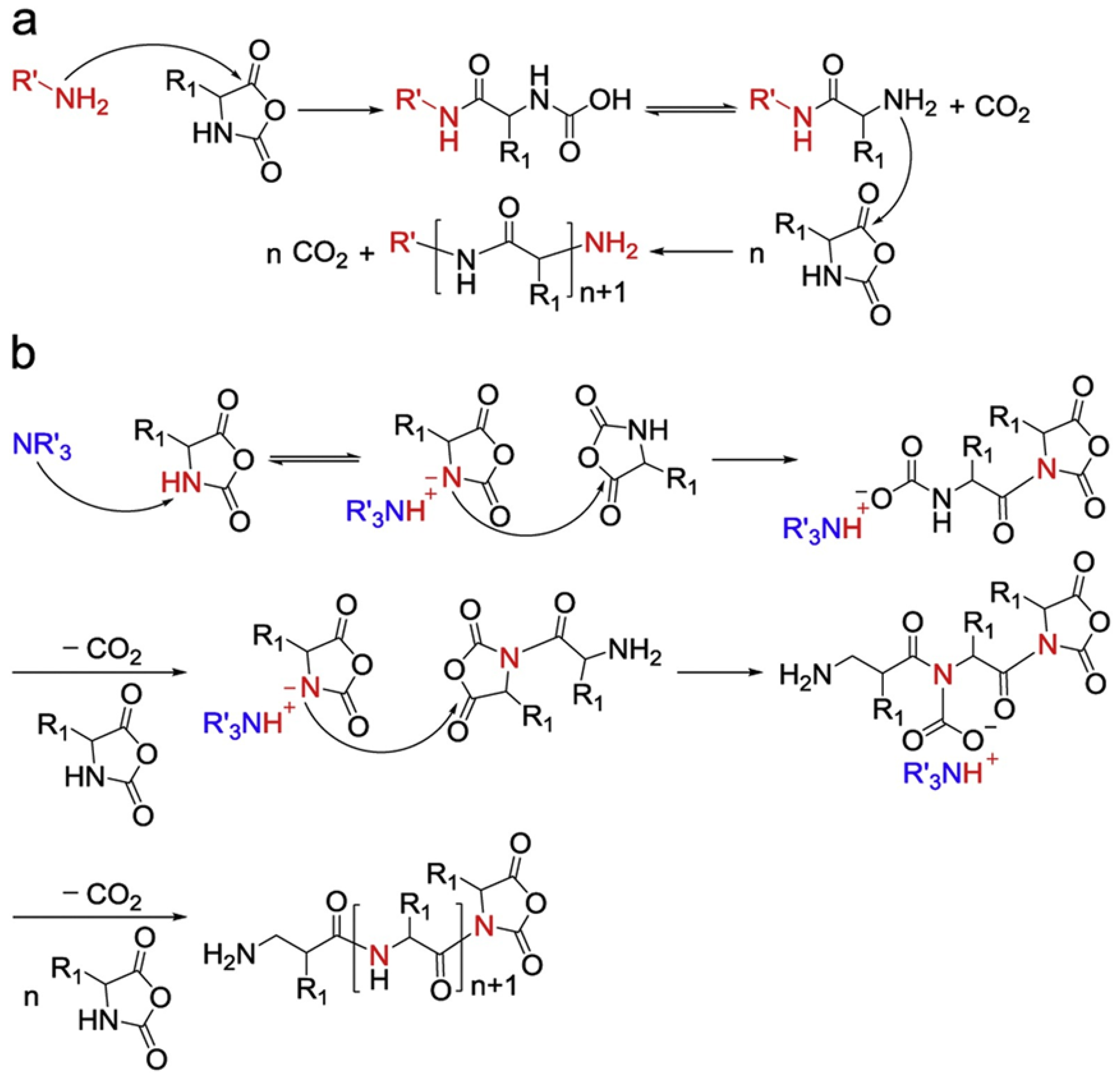
In 1921, Curtius, Wessel, and their coworkers used NCAs to produce polypeptides using ROP methods [30]. The “normal amine mechanism” (NAM) (Figure 2a) and the “active monomer mechanism” (AMM) (Figure 2b) are the two typical routes via which nucleophiles or bases initiate the ROP of NCAs [31]. The NAM not only includes primary and secondary amines but also alcohols and thiols as initiators. Initiation often occurs due to the nucleophilic attack of the carbonyl group of the NCA ring. The intermediate product, carbamic acid, is subsequently formed due to proton transfer and ring opening. Further, the decarboxylation of carbamic acid eliminates CO2 and promotes the propagation of the polymer chain. It has been reported that the NAM can control the molecular weight of polypeptides and the adjustable end groups well. However, due to the side reaction that often occurs along with this process, cyclic polymers and terminated active end groups derived from the impurities in the reactions (water, reaction solvent, or contaminants in NCAs) can be found in the resulting polymers. On the other hand, the AMM mainly involves tertiary amines and alkoxides. It might occur that the resultant NCA anions catch the proton from the nitrogen group of NCAs (3-N). NCA anions can initiate further reactions with other NCAs, as propagation could further involve the attacking of the 5-CO group of other NCAs. In this case, polymerization is untraceable, generating ill-defined polypeptides [32]. It is worth pointing out that these two mechanisms often co-exist in one polymerization process, so the precise control of the reaction conditions is the main issue that needs addressing.
By considering this reported mechanism, various polymerization methods with different initiators, catalysts, and pre-designed NCAs have been reported for polypeptide design over the years. Living polymer methods can greatly enhance the speed of NCA polymerization, and the reaction time can even be minimized to a few minutes. Moreover, “Click chemistry” can be used for the post-conjugation of various groups on the polypeptide chain, thus giving the resulting polypeptide unique stimulus responsiveness. In this method, firstly, various initiators can be used, such as metal-based complexes, amine salts, silane derivatives, etc. Even without any initiators, photo-based chemistry can lead to a polypeptide with tunable molecular weight. Secondly, the catalyst has an important role in controlling the ROP of NCAs. Metallic catalysts and organic catalysts are the two main types of catalysts for controlling the speed of reaction by shortening the propagation time. Metal-based catalysts often result in metal residues on the resulting polypeptide. As organic catalysts do not have this problem, leaving no traces of metal, they offer the most freedom to control the fabrication of polymers that are suitable for various applications, especially in the biomedical area. Additionally, the conditions of high vacuum, low reaction temperatures, N2 flow, and UV can all be applied to polypeptide synthesis and have the ability to accelerate the reaction speed and reduce the side reaction. These improved polymerization technologies are significant, as they give researchers the chance to obtain polypeptide-based polymers with different topologies and interesting structures. Not only can linear, hinged, cyclic, and branched polypeptides be quickly obtained, but also random, alternating, block, and grafted copolypeptides can be obtained by applying sophisticated chemical synthesis skills. These copolymerized polypeptides with different topologies also greatly improve vesicle formation.
PVs are hollow spheres in shape with a bilayer wall of polymers. Commonly, they are formed through the self-assembly of amphiphilic polymers (Figure 3). These vesicles have various advantages over common polymer micelles: (1) the inner aqueous cavity can be loaded with hydrophilic cargo; (2) hydrophobic cargo can be loaded on the membrane of the vesicles; and (3) vesicles can easily disassemble to achieve suitable drug release properties. Thus, stimulus responsiveness is very important in PV formation; for instance, PV nano-assemblies can disassemble or reassemble under the simulation of pH, ROS, enzymes, or infrared light. Thus, in drug delivery, nano-assemblies can easily carry the drug compound to suitable sites and release it with the disassembly of the vesicles.
2.1.1. Anisotropic Packing of α-Helical, Hydrophobic Polypeptides
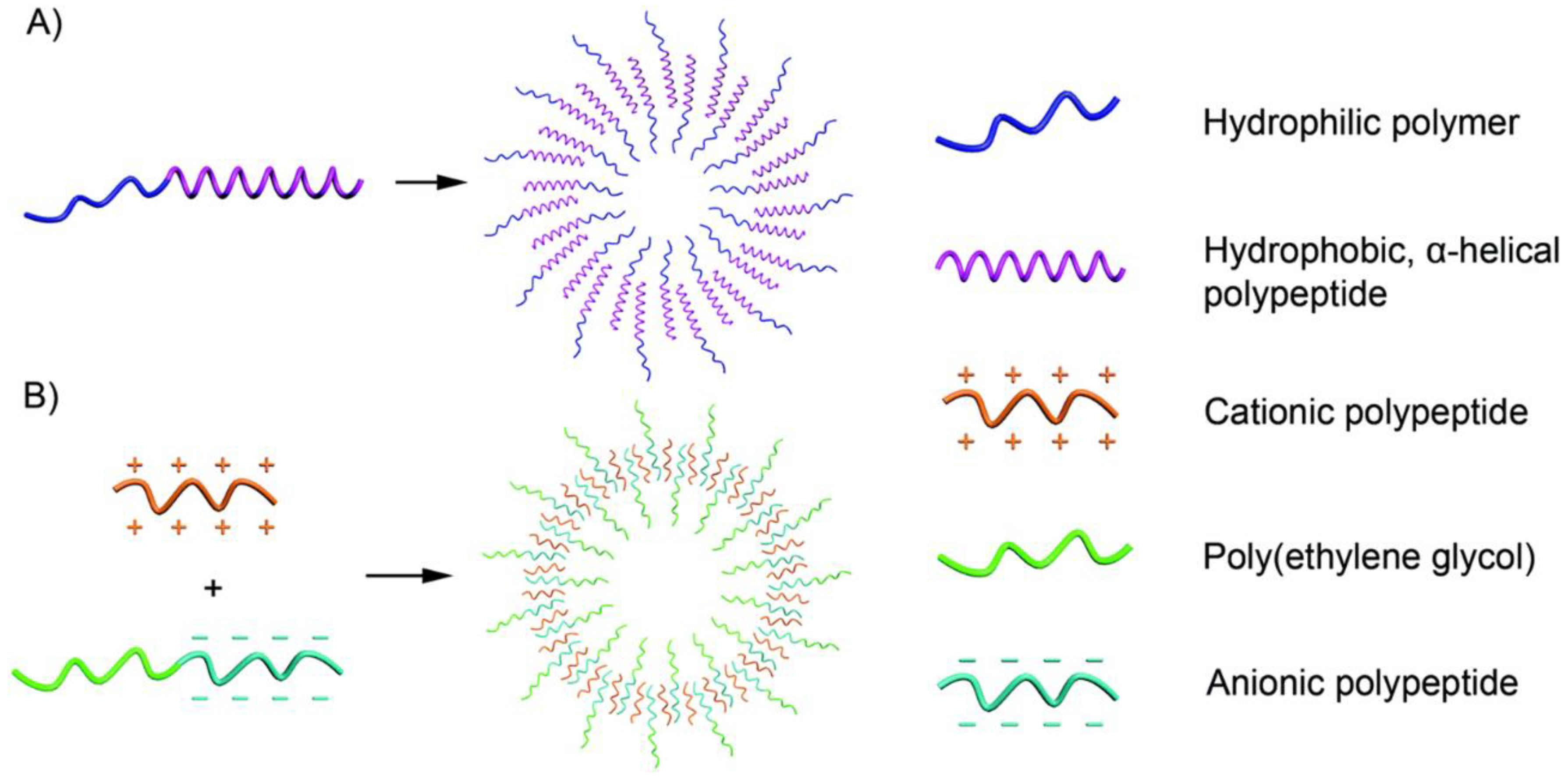
The α-helical conformation can be observed in a variety of polypeptides with rigid rods, such as polyleucine and polyphenylalanine. This is attributed to the hydrophobic polypeptide segment, even at a high hydrophilic-to-hydrophobic ratio. Moreover, research also shows that randomly coiled copolymers can only form micelles, even with finely controlled components. The hydrophobic block becomes the essential component for assembling hydrophobic polypeptides (Figure 3A). For example, as early as 2005, Timothy J. Deming already reported on the charged polypeptides of poly(l-lysine)-b-poly(l-leucine) block copolypeptides, which could be assembled into vesicular assemblies [33]. Afterwards, cationic polypeptides and anionic polypeptides were both reported to be able to form vesicles under suitable conditions [34,35,36]. Polypeptides with carbohydrate moieties can be covalently linked to the side chains of amino acids and form glycopeptides, with this segment as the hydrophilic chain, and polypeptide vesicles can also be formed. Andreas Heise et al. showed that poly(γ-benzyl-l-glutamate) (PBLG)-b-poly (galactosylated propargylglycine) can also form stable vesicles, in which hydrophobic PBLG plays a decisive role [37]. Since hydrophobic polypeptide segments are limited, the hydrophilic chain can be varied. Not only peptide materials but also biodegradable polymers, such as polyethylene glycol (PEG) and hyaluronan (HYA), have been reported in research studies on vesicle preparation.
2.1.2. Oppositely Charged Polymers
Amphiphilic polypeptides can be used to form vesicles using the traditional self-assembly methods introduced by Eisenberg [38] (Figure 3B). First, a solvent dissolves the amphiphilic polypeptides; after that, a solvent specific to one segment is added to the system to induce the formation of vesicles. On the other hand, Kataoka’s group further showed that oppositely charged polypeptides can be assembled into vesicles in aqueous solution. Even without any trace of organic solvent, water can induce vesicle formation.
2.1.3. Others
Recently, reports have also shown that poly(ε-caprolactone)-block-poly(lysine-stat-phenylalanine) [PCL-b-P(Lys-stat-Phe)] and poly(ε-caprolactone)-block-poly(tryptophan)-block-poly(lysine-stat-phenylalanine) might also be able to assemble into vesicles, where PCLs are the resulting hydrophobic membranes [39,40]. Specifically, these are dual corona vesicles, which are induced using solvent-switch methods. In addition, poly (L-glutamic acid) have good solubility at neutral pH; in fact, it was reported that poly (trimethylene carbonate)-b-poly(l-glutamic acid) materials could also assemble into vesicles with altered sizes of 60 nm and 130 nm at pH values of 7.4 and 10.5, respectively [41].
2.2. Production of OMVs
Cells create and release membrane-bound material, which is sometimes referred to as membrane vesicles, microvesicles, exosomes, tolerasomes, agrosomes, and virus-like particles, in all domains of life (Figure 4). The first time OMVs were observed dates to the 1960s, when Chatterjee and Das studied the cell-wall structures of Vibrio cholera (V. cholera) in vitro. Moreover, K. W. Knox et al. firstly found changes in the lysine-limited culture of Escherichia coli, characterized by cell growth with surrounding “globules” and “hollow spheres” [42]. Recently, the biosynthesis field has achieved great advances, and the physiological roles of OMVs have also been found. Moreover, related reports have shown that released vesicles contain lipids, proteins, DNA, RNA, and lipopolysaccharides [43]. In general, OMVs are vesicle structures ranging from about 20 to 300 nm and are produced by almost all Gram-negative bacteria, though a similar phenomenon has also been detected in Gram-positive bacteria. The production of OMVs is still unclear, and why all bacterial cargo can be detected in OMVs remains unknown. However, a growing body of biological evidence is being collected to explain these complex processes. The ability of OMVs to transport a significant amount of the biological materials present in the parent bacterium to distant sites in the host, thus facilitating bacterial communication, the transmission of virulence factors, and the maintenance of bacterial communities, is ultimately due to their capacity to carry cargo [44]. To date, although the mechanism of OMV production remains unclear, three models of OMV biogenesis have been proposed.
2.2.1. Reduced Lipoprotein Linkages
Hypervesiculation is often observed in the case of reduced crosslinking between the outer membrane and the underlying peptidoglycan layer. The reduced number of linkages can lead to the bulging of the outer membrane, thus resulting in the production of vesicles. Dick Hoekstra already showed that vesicles may be easily formed in areas with a low number of outer membrane–murein linkages [45]. Similar results can also be found in the reports by Meta J. Kuehn and Maria Kaparakis-Liaskos [46,47]. Both Acinetobacter baumannii and Escherichia coli have similar OMV production tendencies with reduced crosslinking.
2.2.2. Peptidoglycan Residues
According to this hypothesis, there are locations where the concentration of peptidoglycans is greater during the production of the peptidoglycan layer, which causes protrusions in the outer membrane, signaling the start of vesicle formation. OMVs are released when peptidoglycan residue accumulation causes the outer membrane to bulge even more. For example, Meta J. Kuehn et al. reported that periplasmic peptidoglycan components could increase OMV biogenesis [48].
2.2.3. Electrically Charged Lipopolysaccharides (LPSs)
Electrically charged LPSs are the subject of the third and final OMV production model. Positively charged LPSs and neutrally charged LPSs are the two forms of LPSs that Pseudomonas aeruginosa produce. OMVs are predominantly made of negatively charged LPSs when produced by Pseudomonas aeruginosa under oxidative stress. Due to the repelling effect of the negatively charged outer membrane, an increase in negatively charged LPSs within the cell envelope might facilitate the release of OMVs [49].
2.2.4. Other Mechanisms
OMV biogenesis appears to be a diverse multi-factorial process stimulated or regulated by a variety of pathways/mechanisms that may function simultaneously. For example, the environment has been found to have an important role in OMV production. Kuehn et al. reported that OMV production is a stress-responsive process and that temperature could have an effect on Serratia marcescens and E.coli [48,50]. Wael Elhenawy et al. reported in the Salmonella enterica serovar Typhimurium model that a lipid is necessary for OMV biogenesis [51], that is, deacylation happens within a multifactorial procedure, including planned modification of the outer membrane. Lynne Turnbull et al. used super-resolution microscopy to observe the OMV production process in Pseudomonas aeruginosa biofilms [52]. The release of cytosolic material in Pseudomonas aeruginosa biofilms was found to result from the explosive cell lysis of a subpopulation of cells. Explosive cell lysis also generates broken membrane fragments that form OMVs quickly. Sandro Roier et al. showed that VacJ/Yrb ABC (ATP-binding cassette) is also involved in OMV formation [53]. VacJ/Yrb ABC is a phospholipid transporter. The accumulation of phospholipids in the outer leaflet of the outer membrane causes this general mechanism of OMV generation.
3. Drug Delivery Performance of PVs and OMVs
Recently, it has been found that both PVs and OMVs can be employed in a variety of bio-applications, such as drug delivery [15,29,54,55,56], vaccination [57,58,59], cancer therapy [60,61,62], bioimaging [63,64], biosensing [65,66], and antibacterial applications [39,66,67,68,69,70,71,72]. PVs are generally obtained with amphiphilic block copolymers; thus, hydrophobic membranes can very easily carry hydrophobic drugs. In addition, delivering biomolecules from the parent bacterium to specific distant sites is one of the important functions of OMVs in nature. Further, the stimulus-responsive ability of PVs and OMVs offers the best way to release the loaded drug. Under specific stimuli, such as physical (heat or light), chemical (pH and reducing agents), and biomarker (e.g., ATP or RNA) agents, PVs and OMVs can also respond in a certain way to achieve targeted drug release. Both PVs and OMVs have natural advantages in terms of drug delivery (Table 2).
3.1. Advantages of PVs and OMVs in DDSs
Due to multiple advantages, both PVs and OMVs can be used as platforms in DDSs [15]. The first advantage is the suitable size. PVs can be assembled in vesicles of suitable size by changing the nanoprecipitation conditions. The solvent and injection rate are the main impact factors when water is added to the polymer solvent. Furthermore, pH, polymer concentration, and temperature are important factors in the design of the size of vesicles. Moreover, the size can be altered from several nanometers to micrometers. OMVs commonly have sizes of 50 nm~250 nm. The size of both PVs and OMVs can allow them to pass through the tumor cell membrane owing to the enhanced permeability and retention effect [27]. Secondly, they are biocompatible and biodegradable. Due to the fact that they are derived from natural compounds, both PVs and OMVs are biocompatible and enzyme biodegradable. For example, Sofroniew and Deming showed that amphiphilic polypeptides can be used for the sustainable release of cargo; these polypeptides were detected to have no detectable toxicity to normal cells and could be fully degraded over several weeks [85,86]. OMVs have also been shown to be environmentally inert and biodegradable. Using electron microscopy to analyze OMV morphology, Schulz et al. also discovered that OMVs are biocompatible with differentiated macrophages and epithelial cells [87]. Thirdly, both PVs and OMVs can easily be loaded with the drug compound and protect it [88,89]. Fourthly, both PVs and OMVs can easily deliver drugs to target sites. For example, folic acid can be conjugated with a polypeptide chain and be used as the targeted group [9,90,91,92]. By genetically modifying the parent bacterium, targeting ligands can be added to OMVs. Targeting ligands make it easier for medications to build up at the desired sites. Lastly, PVs can be designed and conjugated with fluorescence groups, such as pyrene or NIR-II, which makes superior fluorescence imaging possible [93,94]. OMVs are produced from bacteria and contain a variety of pathogen-associated molecular patterns; because neutrophils and macrophages can recognize and internalize them, OMVs can also be employed to deliver specific cargoes to these cells [95,96].
Additionally, they each have their own advantages. For example, the chemical synthetic approaches designed so far allow polypeptide-based vesicles to be more easily designed. The stimulus properties of PVs can be engineered with various responsive groups. poly(N-isopropylacrylamide) (PNIPAM) has been found to undergo thermally responsive transformation in water at 32 °C [97]. After conjugating with PNIPAM, PVs become sensitive to temperature, thus achieving selective release. Furthermore, amino acids can be altered depending on the tumor environment. By selecting poly(L-histidine), pH-responsive vehicles were designed based on the closed pKa ≈ 6.0 of the tumor environment [98,99]. In addition, polypeptides can be obtained in just a few minutes depending on the development of the catalysts. Thus, PVs have a bright future in mass production. OMVs trigger an immunological response that is advantageous for the treatment of tumors. This, however, also presents disadvantages, as the immunological response might harm the host if proper measures are not taken to control it.
3.2. Techniques for Drug Loading in PVs and OMVs
3.2.1. Drug Loading in PVs
In PV drug loading, drugs can be divided into hydrophobic drugs, hydrophilic drugs, and metal-based drugs [100,101]. Hydrophobic interaction is the main approach for loading hydrophobic drugs. For example, Sebastien Lecommandoux et al. reported that PEO-b-PBLG diblock and PBLG-b-PEO-b-PBLG triblock copolymers could be loaded with the hydrophobic drug doxorubicin (DOX) [73]. DOX could encapsulate as much as 18 wt.%. Kim et al. showed that poly(ethylene glycol) methyl ether acrylate-block-poly(l-lysine)-block-poly(l-histidine) [p(PEGA)30-b-p(Lys)25-b-p(His)n] can be used for the design of and assembly into the vesicles [74] (Figure 5a). These vesicles, with suitable sizes of 200~372 nm and hydrophobic chains of p(His), were used as a loading site for DOX. The content of DOX was as high as 21 ± 2% and the loading efficiency was 68%.
Besides hydrophobic drugs, hydrophilic drugs can be loaded into PVs through electrostatic interactions, and metallic coordination can easily encapsulate metal-based drugs. For instance, Chen’s group developed PEG-b-poly(l-glutamic acid) (PEG-b-PLG) through the ROP of NCAs using PEG-NH2 as the initiator [75]. Having a portable amino group in the sugar moiety, DOX·HCl is a positively charged and amphiphilic drug. The carboxylate of the glutamic acid units is then electrostatically loaded with DOX-HCl, creating mPEG-b-PLG-DOX·HCl through intermolecular hydrophobic stacking. Kazunori Kataoka’s group prepared oxaliplatin and (1,2-diaminocyclohexane)platinum(II) (DACHPt)-loaded PEG-b-PLG copolymers [77] (Figure 5b). This stable polymer metal complex could be formed to induce drug-loaded PVs.
3.2.2. Drug Loading in OMVs
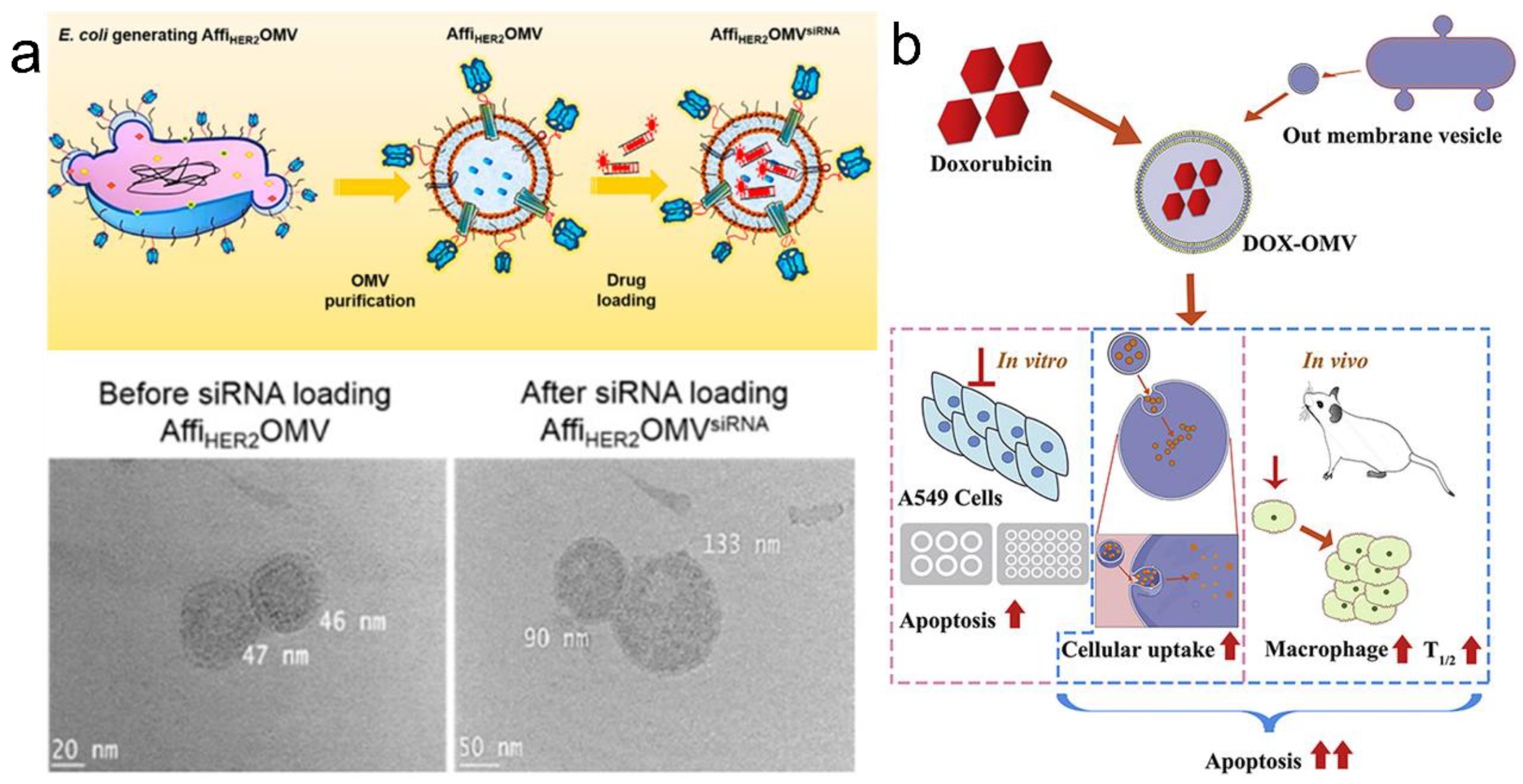
The in vitro and in vivo loading methods are the two main techniques for drug loading in OMVs. The lipid bilayer of OMVs endows them with the potential to be loaded with both hydrophobic and hydrophilic drugs. For example, Sangyong Jon et al. showed that electroporation can be used for loading siRNA into AffiHER2OMVs [55] (Figure 6a). This method was well tolerated by OMVs as evidenced by the integrity of OMVs following siRNA loading and the preserved circular shape. Li Ye’s group also showed that DOX could be loaded into OMVs via passive diffusion [76] (Figure 6b). OMVs from Klebsiella pneumonia were loaded with DOX via passive diffusion in 12 h. DOX fluorescence was observed, which might have concentrated in the nucleus due to the preferential DNA-binding property of DOX. Besides these approaches, incubation, ultrasonication, and extrusion have been reported to be effective methods for in vitro drug loading.
The uniqueness of drug loading in OMVs is that the drug loading process can also be designed during their biogenesis. With such a bioengineering process, it is possible to load cancer treatment drugs. During the generation of OMVs, the incorporation of the drug compound within their parental bacterium is possible. Terry J. Beveridge’s group reported on the treatment of Pseudomonas aeruginosa PAO1 with gentamicin; the released OMVs were found to have the gentamicin [102]. Moreover, gentamicin effects that could arrest cancer cell growth were observed. Ana L. Carvalho et al. showed that OMVs could produce keratinocyte growth factor-2 (KGF-2), a human therapeutic protein, in a stable form [78]. It decreased disease severity when given orally and encouraged intestinal epithelium repair and recovery in animals given colitis-inducing dextran sodium sulfate. Vasilis Ntziachristos’ group also showed that the engineered E. coli could express rate-limiting enzymes in melanin biosynthesis [79]. Thus, the resulting OMVs could kill melanin in vivo. These reports suggest that OMVs could induce cytokine-mediated antitumor activity.
3.3. Targeted Drug Delivery via PVs and OMVs
3.3.1. Targeted Drug Delivery via PVs
After loading the drug compound, its programmed delivery and release at the targeted site are significant. In PVs, changes in the polypeptide assembly behavior and drug–polypeptide interactions are the two main approaches to releasing the drug compound. Since PVs can be easily destroyed due to changes in pH, light, glucose, and reactive oxygen species (ROS) [104] and due to the acidic environment of the tumor, pH responsiveness is the most common stimulus used to release the drug compound. For instance, the sensitivity of poly(l-histidine) could allow the released DOX load to kill cancers [105]. Ding et al. reported that the micelle-to-vesicle transition could be observed on a cholesterol-decorated polypeptide [80]. Due to ROS changes, the conformation of the polypeptide could change from β-sheet to α-helix and further induce on-demand drug release at the targeted sites. The advantage of synthesized polypeptides is that the various environmentally sensitive nonpeptide groups can be conjugated on light polymer chains. Light- and glucose-responsive groups such as phenylboronic acid, spiropyran, and coumarin can change the assembly behavior upon environmental changes such as UV irradiation and insulin delivery systems. For instance, Dong’s group showed that it is possible to first design S-(o-nitrobenzyl)-l-cysteine and then block it with PEG via ROP [81]. The o-nitrobenzyl groups can be leveled off with UV irradiation. Moreover, the release of the drug compound can be controlled using the irradiation time.
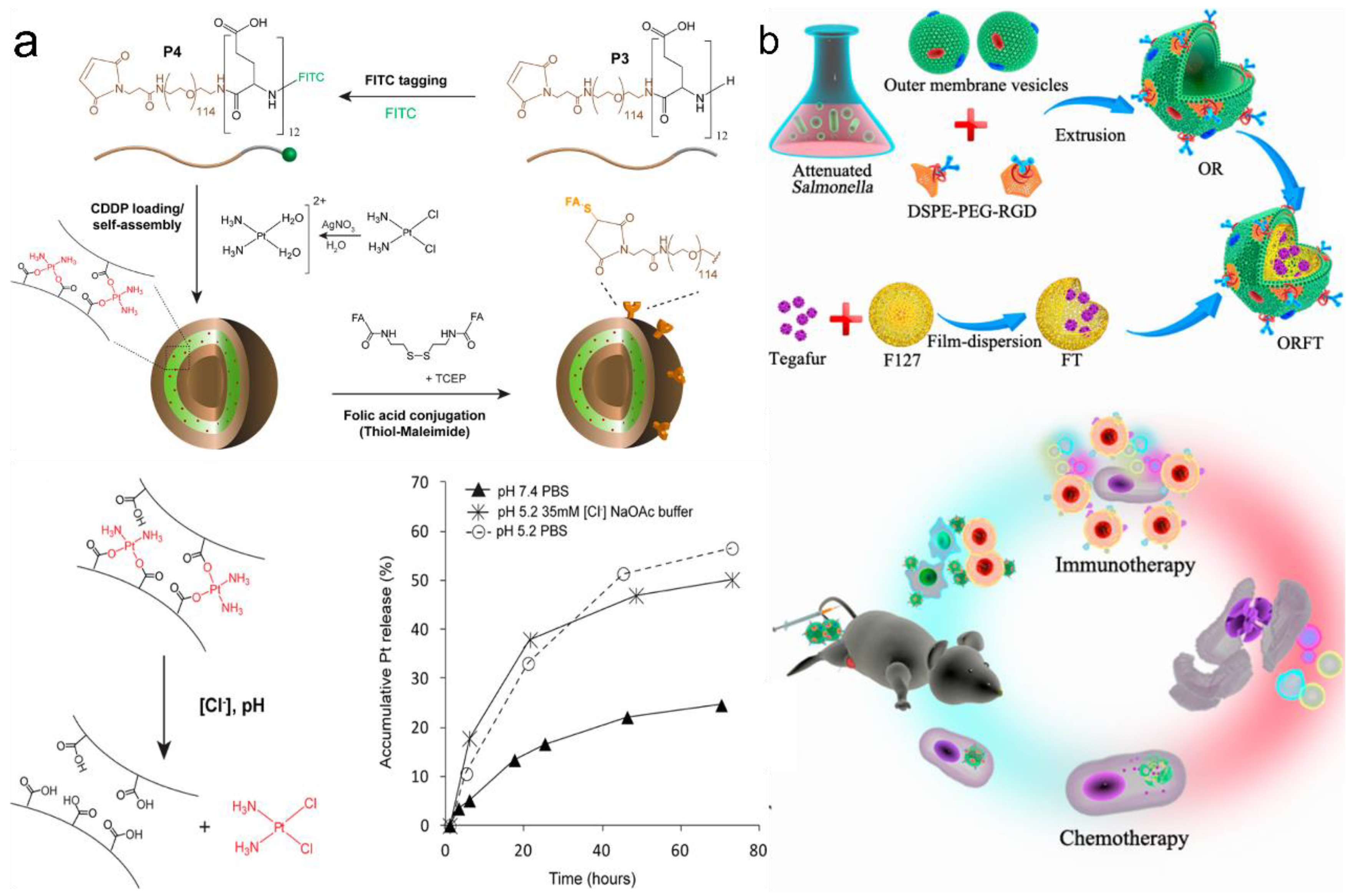
Another important approach is destroying the interactions between the drug compound and the polypeptides. As mentioned above, the drug compound can be covalently or non-covalently conjugated with the polypeptides. Metal complexation and covalently linked drugs bearing polypeptides are mainly used in this release method. Kazunori Kataoka et al. designed hydrazide groups as the linkages between polypeptide chains and the drug DOX [82]. The hydrazide group can be disrupted as the pH changes and further release free DOX. Greg G. Qiao’s group showed that ligand exchange could be used for metal-based drug delivery [103] (Figure 7a). Cisplatin complexed with PEG-B-PLG could be released in the presence of chloride ions. However, it was found that the metal–ligand exchange could only lead to a relatively slow release of the drug compound. This release rate was enhanced when mimicking the endosomal/lysosomal environment (pH 5.2/35 mM [Cl−]).
3.3.2. Targeted Drug Delivery via OMVs
Compared with the scalable and chemically designable PVs, the use of OMVs in targeted drug delivery has only been sporadically reported in recent years and is still an emerging area requiring urgent research [28]. Sangyong Jon’s study is recognized as the first report on using OMVs for targeted drug delivery [55]. After the electroporation of the siRNA drug compound (~15%) into OMVs, the anticancer activity of the drug-containing OMVs was found to be able to inhibit cancer cell proliferation. The detailed mechanism was observed with a confocal microscope as follows: the acidic environment could enhance the degradation of OMVs, and free siRNA was gradually detected along with dead cancer cells. Further, an in vivo study also suggested that the EPR effect is the drug force necessary for targeted drug delivery. Recently, Ping et al. showed that Salmonella-based OMVs could be even coated on polymer F127 nanoparticles [83] (Figure 7b). In this case, the drug compound was loaded on polymer micelles, whereas the OMVs only triggered the host immune response to cancer immunotherapy. This OMV-coated type of DDS provides a new combined system for potential targeted drug delivery. In addition, some recently emerged research also showed that bioengineered OMVs could induce specific CD8 T cell response, and this cell could control tumor growth via viral replication [84]. These recent findings open further possibilities for OMV drug delivery studies.
4. Conclusions and Outlook
This review mainly describes PVs and OMVs in drug delivery applications. PVs are obtained with the artificial synthesis of polypeptides, where the NMM and AMM are the two major mechanisms for polypeptide synthesis via the ROP of NCAs. Polymer vesicles are then obtained via self-assembly. The anisotropic packing of α-helical, hydrophobic polypeptides and the use of oppositely charged polymers are methods for the preparation of PVs. Compared with PVs, OMV biogenesis remains a mystery; however, three models, i.e., reduced lipoprotein linkages, peptidoglycan residues, and electrically charged lipopolysaccharides, are recognized as the major influencing factors. In addition, other factors, such as the environment and lipids, are important for OMV biogenesis in certain bacteria. Finally, the similarities and differences between these two vesicle types in drug delivery applications are elaborately compared. Specifically, we compare their applications in DDSs in the following aspects: general advantages, drug loading techniques, and targeted drug delivery performance. Based on state-of-the-art PVs and OMVs, we believe that future research should address the following issues to advance toward further DDS applications:
- (1)
- Mass production of OMVs and PVs: Currently, the production of OMVs and PVs is still in the laboratory research stage. Although, recently, the production efficiency of polypeptide materials has been rapidly improved, technical solutions for assembling vesicles still lack specific standards, and large-scale production of PVs is not yet possible. OMV sizes are difficult to regulate, and there is significant variance among batches. In addition, low particle programmability is caused by our lack of knowledge regarding the precise OMV assembly mechanism. Enhancing safety and reducing costs are also still problems that need to be addressed.
- (2)
- Universality of drug loading and delivery targeting: The drug loading mechanism in the process of production and post-treatment is inefficient and is in urgent need of optimization. Given the relevance of various cancer types and given that there are various PV and OMV types, consistent tumor cell targeting standards still need to be achieved and further improved. Individual PVs and OMVs may be effective for certain cancers.
- (3)
- Other specific issues: The immunotoxicity of OMVs is a problem that cannot be ignored in DDSs. Moreover, the artificial production of OMVs also deserves more in-depth study on its potential. In PVs, the deprotection process in the production of specific polypeptides (such as polyhistidine, polylysine, and polycysteine) is tedious. Finally, programmed drug release and the benefits of the unique secondary structure of PVs are also worthy of more in-depth research.
Author Contributions
Y.Z. and W.S. wrote the original draft; Y.L. (Yiming Lu) and Z.Z. reviewed and edited the manuscript; Y.X., Y.L. (Yichong Li) and W.L. critically reviewed and revised the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
The following financial supports are appreciated: Scientific Research Foundation of the Shanghai University of Medicine and Health Sciences (SSF-22-07-02) and Shanghai Sailing Program (22YF1417700).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Data supporting this publication are available from the corresponding authors.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Li, J.; Yu, F.; Chen, Y.; Oupický, D. Polymeric drugs: Advances in the development of pharmacologically active polymers. J. Control. Release 2015, 219, 369–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gale, G.R.; Morris, C.R.; Atkins, L.M.; Smith, A.B. Binding of an antitumor platinum compound to cells as influenced by physical factors and pharmacologically active agents. Cancer Res. 1973, 33, 813–818. [Google Scholar] [PubMed]
- Xian, C.; Chen, H.; Xiong, F.; Fang, Y.; Huang, H.; Wu, J. Platinum-based chemotherapy via nanocarriers and co-delivery of multiple drugs. Biomater. Sci. 2021, 9, 6023–6036. [Google Scholar] [CrossRef] [PubMed]
- Tabakoglu, S.; Kołbuk, D.; Sajkiewicz, P. Multifluid electrospinning for multi-drug delivery systems: Pros and cons, challenges, and future directions. Biomater. Sci. 2022, 11, 37–61. [Google Scholar] [CrossRef] [PubMed]
- Tibbitt, M.W.; Dahlman, J.E.; Langer, R. Emerging frontiers in drug delivery. J. Am. Chem. Soc. 2016, 138, 704–717. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gao, J.; Wang, Z. Outer membrane vesicles for vaccination and targeted drug delivery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1523. [Google Scholar] [CrossRef] [PubMed]
- Loo, Y.S.; Bose, R.J.; McCarthy, J.R.; Azmi, I.D.M.; Madheswaran, T. Biomimetic bacterial and viral-based nanovesicles for drug delivery, theranostics, and vaccine applications. Drug Discov. Today 2021, 26, 902–915. [Google Scholar] [CrossRef]
- Al-Dossary, A.A.; Isichei, A.C.; Zhang, S.; Li, J.; Errachid, A.; Elaissari, A. Outer membrane vesicles (OMVs) as a platform for vaccination and targeted drug delivery. In Pharmaceutical Nanobiotechnology for Targeted Therapy; Springer: Berlin/Heidelberg, Germany, 2022; pp. 1–25. [Google Scholar]
- Tang, Y.; Varyambath, A.; Ding, Y.; Chen, B.; Huang, X.; Zhang, Y.; Yu, D.-g.; Kim, I.; Song, W. Porous organic polymers for drug delivery: Hierarchical pore structures, variable morphologies, and biological properties. Biomater. Sci. 2022, 10, 5369–5390. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, X.; Wu, X.; Li, Y.; Tang, W.; Yang, C.; Ding, B. Chemically modified DNA nanostructures for drug delivery. Innovation 2022, 3, 100217. [Google Scholar] [CrossRef]
- Song, W.; Zhang, M.; Huang, X.; Chen, B.; Ding, Y.; Zhang, Y.; Yu, D.G.; Kim, I. Smart l-borneol-loaded hierarchical hollow polymer nanospheres with antipollution and antibacterial capabilities. Mater. Today Chem. 2022, 26, 101252. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, M.; Yan, C.; Liu, H.; Yu, D.-G. Advances in the application of electrospun drug-loaded nanofibers in the treatment of oral ulcers. Biomolecules 2022, 12, 1254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, R.; Jin, H.; Song, W.; Augustine, R.; Kim, I. Straightforward access to linear and cyclic polypeptides. Commun. Chem. 2018, 1, 40. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Han, Z.; Lv, S.; Chen, C.; Chen, L.; Yin, L.; Cheng, J. Synthetic polypeptides: From polymer design to supramolecular assembly and biomedical application. Chem. Soc. Rev. 2017, 46, 6570–6599. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, Z.; Wei, S.; Ji, G.; Zheng, X.; Fu, Z.; Cheng, J. Polypeptide-based drug delivery systems for programmed release. Biomaterials 2021, 275, 120913. [Google Scholar] [CrossRef] [PubMed]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-based vaccines: Current progress and future challenges. Chem. Rev. 2019, 120, 3210–3229. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Liu, Q. Engineered bacterial outer membrane vesicles as multifunctional delivery platforms. Front. Mater. 2020, 7, 202. [Google Scholar] [CrossRef]
- Chen, W.; Wu, Y.; Deng, J.; Yang, Z.; Chen, J.; Tan, Q.; Guo, M.; Jin, Y. Phospholipid-membrane-based nanovesicles acting as vaccines for tumor immunotherapy: Classification, mechanisms and applications. Pharmaceutics 2022, 14, 2446. [Google Scholar] [CrossRef]
- Srivastava, P.; Kim, K.-S. Membrane vesicles derived from gut microbiota and probiotics: Cutting-edge therapeutic approaches for multidrug-resistant superbugs linked to neurological anomalies. Pharmaceutics 2022, 14, 2370. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, L.; Fu, H.; Wei, C.; Jin, Q. Comparative proteomic analysis of outer membrane vesicles from Shigella flexneri under different culture conditions. Biochem. Biophys. Res. Commun. 2014, 453, 696–702. [Google Scholar] [CrossRef]
- Avila-Calderón, E.D.; Ruiz-Palma, M.D.S.; Aguilera-Arreola, M.G.; Velázquez-Guadarrama, N.; Ruiz, E.A.; Gomez-Lunar, Z.; Witonsky, S.; Contreras-Rodríguez, A. Outer membrane vesicles of gram-negative bacteria: An outlook on biogenesis. Front. Microbiol. 2021, 12, 557902. [Google Scholar] [CrossRef]
- Gao, W.; Fang, R.H.; Thamphiwatana, S.; Luk, B.T.; Li, J.; Angsantikul, P.; Zhang, Q.; Hu, C.-M.J.; Zhang, L. Modulating antibacterial immunity via bacterial membrane-coated nanoparticles. Nano Lett. 2015, 15, 1403–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, L.; Praszkier, J.; Hutton, M.L.; Steer, D.; Ramm, G.; Kaparakis-Liaskos, M.; Ferrero, R.L. Increased outer membrane vesicle formation in a Helicobacter pylori tolB mutant. Helicobacter 2015, 20, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-L.; Oh, J.; Singh, S.; Chen, R.; Chen, W. Functional assembly of minicellulosomes on the Saccharomyces cerevisiae cell surface for cellulose hydrolysis and ethanol production. Appl. Environ. Microbiol. 2009, 75, 6087–6093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauman, S.J.; Kuehn, M.J. Purification of outer membrane vesicles from Pseudomonas aeruginosa and their activation of an IL-8 response. Microbes Infect. 2006, 8, 2400–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, D.; Chaudhuri, K. Association of cholera toxin with Vibrio cholerae outer membrane vesicles which are internalized by human intestinal epithelial cells. FEBS Lett. 2011, 585, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef]
- Huang, Y.; Nieh, M.P.; Chen, W.; Lei, Y. Outer membrane vesicles (OMVs) enabled bio-applications: A critical review. Biotechnol. Bioeng. 2022, 119, 34–47. [Google Scholar] [CrossRef]
- Zhang, Y.; Kim, I.; Lu, Y.; Xu, Y.; Yu, D.-G.; Song, W. Intelligent poly(l-histidine)-based nanovehicles for controlled drug delivery. J. Control. Release 2022, 349, 963–982. [Google Scholar] [CrossRef]
- Leuchs, H.; Geiger, W. The anhydrides of alpha-amino-N-carboxylic and alpha-amino acids. Ber. Dtsch. Chem. Ges. 1908, 41, 1721–1726. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, D.; Ding, J.; Chen, X. Controlled synthesis of polypeptides. Chin. Chem. Lett. 2020, 31, 3001–3014. [Google Scholar] [CrossRef]
- Deming, T.J. Polypeptide Materials: New synthetic methods and applications. Adv. Mater. 1997, 9, 299–311. [Google Scholar] [CrossRef]
- Holowka, E.P.; Pochan, D.J.; Deming, T.J. Charged Polypeptide vesicles with controllable diameter. J. Am. Chem. Soc. 2005, 127, 12423–12428. [Google Scholar] [CrossRef] [PubMed]
- Sun, V.Z.; Choe, U.J.; Rodriguez, A.R.; Dai, H.; Deming, T.J.; Kamei, D.T. Transfection of mammalian cells using block copolypeptide vesicles. Macromol. Biosci. 2013, 13, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Choe, U.-J.; Rodriguez, A.R.; Lee, B.S.; Knowles, S.M.; Wu, A.M.; Deming, T.J.; Kamei, D.T. Endocytosis and intracellular trafficking properties of transferrin-conjugated block copolypeptide vesicles. Biomacromolecules 2013, 14, 1458–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.R.; Choe, U.-J.; Kamei, D.T.; Deming, T.J. Blending of diblock and triblock copolypeptide amphiphiles yields cell penetrating vesicles with low toxicity. Macromol. Biosci. 2015, 15, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Bonduelle, C.; Thévenot, J.; Lecommandoux, S.; Heise, A. Biologically active polymersomes from amphiphilic glycopeptides. J. Am. Chem. Soc. 2012, 134, 119–122. [Google Scholar] [CrossRef]
- Yu, K.; Eisenberg, A. Bilayer morphologies of self-assembled crew-cut aggregates of amphiphilic PS-b-PEO diblock copolymers in solution. Macromolecules 1998, 31, 3509–3518. [Google Scholar] [CrossRef]
- Yang, Y.-Y.; Chen, L.-S.; Sun, M.; Wang, C.-Y.; Fan, Z.; Du, J.-Z. Biodegradable polypeptide-based vesicles with intrinsic blue fluorescence for antibacterial visualization. Chin. J. Polym. Sci. 2021, 39, 1412–1420. [Google Scholar] [CrossRef]
- Xi, Y.; Wang, Y.; Gao, J.; Xiao, Y.; Du, J. Dual Corona vesicles with intrinsic antibacterial and enhanced antibiotic delivery capabilities for effective treatment of biofilm-induced periodontitis. ACS Nano 2019, 13, 13645–13657. [Google Scholar] [CrossRef]
- Sanson, C.; Schatz, C.; Le Meins, J.-F.; Soum, A.; Thévenot, J.; Garanger, E.; Lecommandoux, S. A simple method to achieve high doxorubicin loading in biodegradable polymersomes. J. Control. Release 2010, 147, 428–435. [Google Scholar] [CrossRef]
- Knox, K.; Vesk, M.; Work, E. Relation between excreted lipopolysaccharide complexes and surface structures of a lysine-limited culture of Escherichia coli. J. Bacteriol. 1966, 92, 1206–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsui, N.; Tsuchido, T.; Hiramatsu, R.; Fujikawa, S.; Takano, M.; Shibasaki, I. Heat-induced blebbing and vesiculation of the outer membrane of Escherichia coli. J. Bacteriol. 1982, 151, 1523–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.-W.; Baeg, Y.; Jeong, H.-Y.; Lee, J.; Oh, D.-K.; Hollmann, F.; Park, J.-B. Bacterial outer membrane vesicles as nano-scale bioreactors: A fatty acid conversion case study. ChemCatChem 2021, 13, 4080–4086. [Google Scholar] [CrossRef]
- Hoekstra, D.; van der Laan, J.W.; de Leij, L.; Witholt, B. Release of outer membrane fragments from normally growing Escherichia coli. Biochim. Biophys. Acta (BBA) Biomembr. 1976, 455, 889–899. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Rodriguez, D.L.; Kuehn, M.J. NlpI-mediated modulation of outer membrane vesicle production through peptidoglycan dynamics in Escherichia coli. MicrobiologyOpen 2015, 4, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Pathirana, R.D.; Kaparakis-Liaskos, M. Bacterial membrane vesicles: Biogenesis, immune regulation and pathogenesis. Cell. Microbiol. 2016, 18, 1518–1524. [Google Scholar] [CrossRef] [Green Version]
- McBroom, A.J.; Kuehn, M.J. Release of outer membrane vesicles by gram-negative bacteria is a novel envelope stress response. Mol. Microbiol. 2007, 63, 545–558. [Google Scholar] [CrossRef]
- Kadurugamuwa, J.L.; Beveridge, T.J. Virulence factors are released from Pseudomonas aeruginosa in association with membrane vesicles during normal growth and exposure to gentamicin: A novel mechanism of enzyme secretion. J. Bacteriol. 1995, 177, 3998–4008. [Google Scholar] [CrossRef] [Green Version]
- McMahon, K.J.; Castelli, M.E.; Vescovi, E.G.; Feldman, M.F. Biogenesis of outer membrane vesicles in Serratia marcescens is thermoregulated and can be induced by activation of the Rcs phosphorelay system. J. Bacteriol. 2012, 194, 3241–3249. [Google Scholar] [CrossRef] [Green Version]
- Elhenawy, W.; Bording-Jorgensen, M.; Valguarnera, E.; Haurat, M.F.; Wine, E.; Feldman, M.F. LPS remodeling triggers formation of outer membrane vesicles in Salmonella. mBio 2016, 7, e00940-16. [Google Scholar] [CrossRef]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roier, S.; Zingl, F.G.; Cakar, F.; Durakovic, S.; Kohl, P.; Eichmann, T.O.; Klug, L.; Gadermaier, B.; Weinzerl, K.; Prassl, R.; et al. A novel mechanism for the biogenesis of outer membrane vesicles in Gram-negative bacteria. Nat. Commun. 2016, 7, 10515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Song, W.; Lu, Y.; Xu, Y.; Wang, C.; Yu, D.-G.; Kim, I. Recent advances in poly(alpha-L-glutamic acid)-based nanomaterials for drug delivery. Biomolecules 2022, 12, 636. [Google Scholar] [CrossRef] [PubMed]
- Gujrati, V.; Kim, S.; Kim, S.-H.; Min, J.J.; Choy, H.E.; Kim, S.C.; Jon, S. Bioengineered bacterial outer membrane vesicles as cell-specific drug-delivery vehicles for cancer therapy. ACS Nano 2014, 8, 1525–1537. [Google Scholar] [CrossRef]
- Yin, L.; Cheng, J.; Deming, T.J.; Vicent, M.J. Synthetic polypeptides for drug and gene delivery, and tissue engineering. Adv. Drug Delivery Rev. 2021, 178, 113995. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Song, K.; Yen, A.; Peeler, D.J.; Nguyen, D.C.; Olshefsky, A.; Sylvestre, M.; Srinivasan, S.; Stayton, P.S.; Pun, S.H. Well-defined mannosylated polymer for peptide vaccine delivery with enhanced antitumor immunity. Adv. Healthc. Mater. 2022, 11, 2101651. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.L.; Shrimali, P.C.; Clapacs, Z.P.; Files, M.A.; Rudra, J.S. Peptide-based supramolecular vaccine systems. Acta Biomater. 2021, 133, 153–167. [Google Scholar] [CrossRef]
- van der Pol, L.; Stork, M.; van der Ley, P. Outer membrane vesicles as platform vaccine technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.Y.; Park, H.T.; Dinh, N.T.H.; Choi, S.J.; Lee, J.; Kim, J.H.; Lee, S.-W.; Gho, Y.S. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat. Commun. 2017, 8, 626. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Ma, Y.; Du, C.; Wang, C.; Chen, T.; Wang, Y.; Wang, J.; Yao, Y.; Dong, C.-M. NO-releasing polypeptide nanocomposites reverse cancer multidrug resistance via triple therapies. Acta Biomater. 2021, 123, 335–345. [Google Scholar] [CrossRef]
- Wang, Q.; Jiang, N.; Fu, B.; Huang, F.; Liu, J. Self-assembling peptide-based nanodrug delivery systems. Biomater. Sci. 2019, 7, 4888–4911. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.B.; Gao, Y.J.; Wang, L.; Wang, H. Self-assembled peptide-based nanomaterials for biomedical imaging and therapy. Adv. Mater. 2018, 30, 1703444. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, H.; Feng, J.-J.; Luo, X.; Fang, K.-M.; Wang, Z.-G.; Wang, A.-J. A polypeptide-mediated synthesis of green fluorescent gold nanoclusters for Fe3+ sensing and bioimaging. J. Colloid Interface Sci. 2017, 506, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, D.; Hui, N. A low fouling electrochemical biosensor based on the zwitterionic polypeptide doped conducting polymer PEDOT for breast cancer marker BRCA1 detection. Bioelectrochemistry 2020, 136, 107595. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.J.; Cash, K.J.; Hugenberg, V.; Plaxco, K.W. Peptide Beacons: A new design for polypeptide-based optical biosensors. Bioconjug. Chem. 2007, 18, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, W.; Li, S.; Kim, D.-K.; Kim, J.H.; Kim, J.R.; Kim, I. Facile and scalable synthesis of topologically nanoengineered polypeptides with excellent antimicrobial activities. Chem. Commun. 2020, 56, 356–359. [Google Scholar] [CrossRef]
- Lu, Z.; Wu, Y.; Cong, Z.; Qian, Y.; Wu, X.; Shao, N.; Qiao, Z.; Zhang, H.; She, Y.; Chen, K. Effective and biocompatible antibacterial surfaces via facile synthesis and surface modification of peptide polymers. Bioact. Mater. 2021, 6, 4531–4541. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, K.; Wu, X.; Liu, L.; Zhang, W.; Ding, Y.; Liu, S.; Zhou, M.; Shao, N.; Ji, Z.; et al. Superfast and Water-Insensitive Polymerization on α-amino acid N-carboxyanhydrides to prepare polypeptides using tetraalkylammonium carboxylate as the initiator. Angew. Chem. Int. Ed. 2021, 60, 26063–26071. [Google Scholar] [CrossRef]
- Ding, X.; Duan, S.; Ding, X.; Liu, R.; Xu, F.-J. Versatile antibacterial materials: An emerging arsenal for combatting bacterial pathogens. Adv. Funct. Mater. 2018, 28, 1802140. [Google Scholar] [CrossRef]
- Pizarro-Cerdá, J.; Cossart, P. Bacterial adhesion and entry into host cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef]
- Asadi, A.; Razavi, S.; Talebi, M.; Gholami, M. A review on anti-adhesion therapies of bacterial diseases. Infection 2019, 47, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Kakkar, D.; Mazzaferro, S.; Thevenot, J.; Schatz, C.; Bhatt, A.; Dwarakanath, B.S.; Singh, H.; Mishra, A.K.; Lecommandoux, S. Amphiphilic PEO-b-PBLG diblock and PBLG-b-PEO-b-PBLG triblock copolymer based nanoparticles: Doxorubicin loading and in vitro evaluation. Macromol. Biosci. 2015, 15, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.P.; Uthaman, S.; John, J.V.; Lee, H.R.; Lee, S.J.; Park, H.; Park, I.-K.; Suh, H.; Kim, I. Poly(PEGA)-b-poly(l-lysine)-b-poly(l-histidine) hybrid vesicles for tumoral pH-triggered intracellular delivery of doxorubicin hydrochloride. ACS Appl. Mater. Interfaces 2015, 7, 21770–21779. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Song, W.; Tang, Z.; Lv, S.; Lin, L.; Sun, H.; Li, Q.; Yang, Y.; Hong, H.; Chen, X. Nanoscaled poly(l-glutamic acid)/doxorubicin-amphiphile complex as pH-responsive drug delivery system for effective treatment of nonsmall cell lung cancer. ACS Appl. Mater. Interfaces 2013, 5, 1781–1792. [Google Scholar] [CrossRef]
- Kuerban, K.; Gao, X.; Zhang, H.; Liu, J.; Dong, M.; Wu, L.; Ye, R.; Feng, M.; Ye, L. Doxorubicin-loaded bacterial outer-membrane vesicles exert enhanced anti-tumor efficacy in non-small-cell lung cancer. Acta Pharm. Sin. B 2020, 10, 1534–1548. [Google Scholar] [CrossRef]
- Ahn, J.; Miura, Y.; Yamada, N.; Chida, T.; Liu, X.; Kim, A.; Sato, R.; Tsumura, R.; Koga, Y.; Yasunaga, M.; et al. Antibody fragment-conjugated polymeric micelles incorporating platinum drugs for targeted therapy of pancreatic cancer. Biomaterials 2015, 39, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.L.; Fonseca, S.; Miquel-Clopés, A.; Cross, K.; Kok, K.-S.; Wegmann, U.; Gil-Cardoso, K.; Bentley, E.G.; Al Katy, S.H.; Coombes, J.L. Bioengineering commensal bacteria-derived outer membrane vesicles for delivery of biologics to the gastrointestinal and respiratory tract. J. Extracell. Vesicles 2019, 8, 1632100. [Google Scholar] [CrossRef]
- Gujrati, V.; Prakash, J.; Malekzadeh-Najafabadi, J.; Stiel, A.; Klemm, U.; Mettenleiter, G.; Aichler, M.; Walch, A.; Ntziachristos, V. Bioengineered bacterial vesicles as biological nano-heaters for optoacoustic imaging. Nat. Commun. 2019, 10, 1114. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, R.; Wei, J.; Cheng, C.; Zheng, Y.; Pan, Y.; He, X.; Ding, M.; Tan, H.; Fu, Q. Conformation-directed micelle-to-vesicle transition of cholesterol-decorated polypeptide triggered by oxidation. J. Am. Chem. Soc. 2018, 140, 6604–6610. [Google Scholar] [CrossRef]
- Liu, G.; Dong, C.-M. Photoresponsive Poly(S-(o-nitrobenzyl)-l-cysteine)-b-PEO from a l-cysteine N-carboxyanhydride monomer: Synthesis, self-assembly, and phototriggered drug release. Biomacromolecules 2012, 13, 1573–1583. [Google Scholar] [CrossRef]
- Bae, Y.; Fukushima, S.; Harada, A.; Kataoka, K. Design of Environment-Sensitive Supramolecular assemblies for intracellular drug delivery: Polymeric micelles that are responsive to intracellular pH change. Angew. Chem. Int. Ed. 2003, 42, 4640–4643. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Bai, H.; Wu, W.; Huang, G.; Li, Y.; Wu, M.; Tang, G.; Ping, Y. Bioengineering bacterial vesicle-coated polymeric nanomedicine for enhanced cancer immunotherapy and metastasis prevention. Nano Lett. 2020, 20, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Schetters, S.T.T.; Jong, W.S.P.; Horrevorts, S.K.; Kruijssen, L.J.W.; Engels, S.; Stolk, D.; Daleke-Schermerhorn, M.H.; Garcia-Vallejo, J.; Houben, D.; Unger, W.W.J. Outer membrane vesicles engineered to express membrane-bound antigen program dendritic cells for cross-presentation to CD8+ T cells. Acta Biomater. 2019, 91, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Song, J.; Zhang, S.; Anderson, M.A.; Ao, Y.; Yang, C.-Y.; Deming, T.J.; Sofroniew, M.V. Sustained local delivery of bioactive nerve growth factor in the central nervous system via tunable diblock copolypeptide hydrogel depots. Biomaterials 2012, 33, 9105–9116. [Google Scholar] [CrossRef]
- Zhang, S.; Anderson, M.A.; Ao, Y.; Khakh, B.S.; Fan, J.; Deming, T.J.; Sofroniew, M.V. Tunable diblock copolypeptide hydrogel depots for local delivery of hydrophobic molecules in healthy and injured central nervous system. Biomaterials 2014, 35, 1989–2000. [Google Scholar] [CrossRef] [Green Version]
- Schulz, E.; Goes, A.; Garcia, R.; Panter, F.; Koch, M.; Müller, R.; Fuhrmann, K.; Fuhrmann, G. Biocompatible bacteria-derived vesicles show inherent antimicrobial activity. J. Control. Release 2018, 290, 46–55. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, J.; Zhao, X.; Zhang, Y.; Zhong, Z.; Deng, C. A polymeric IDO inhibitor based on poly(ethylene glycol)-b-poly(l-tyrosine-co-1-methyl-d-tryptophan) enables facile trident cancer immunotherapy. Biomater. Sci. 2022, 10, 5731–5743. [Google Scholar] [CrossRef]
- Mohammadi, M.; Taghavi, S.; Abnous, K.; Taghdisi, S.M.; Ramezani, M.; Alibolandi, M. Hybrid vesicular drug delivery systems for cancer therapeutics. Adv. Funct. Mater. 2018, 28, 1802136. [Google Scholar] [CrossRef]
- Song, W.; Zhang, Y.; Yu, D.-G.; Tran, C.H.; Wang, M.; Varyambath, A.; Kim, J.; Kim, I. Efficient synthesis of folate-conjugated hollow polymeric capsules for accurate drug delivery to cancer cells. Biomacromolecules 2021, 22, 732–742. [Google Scholar] [CrossRef]
- Cui, Y.; Deng, R.; Li, X.; Wang, X.; Jia, Q.; Bertrand, E.; Meguellati, K.; Yang, Y.-W. Temperature-sensitive polypeptide brushes-coated mesoporous silica nanoparticles for dual-responsive drug release. Chin. Chem. Lett. 2019, 30, 2291–2294. [Google Scholar] [CrossRef]
- Lin, B.; Wu, J.; Wang, Y.; Sun, S.; Yuan, Y.; Tao, X.; Lv, R. Peptide functionalized upconversion/NIR II luminescent nanoparticles for targeted imaging and therapy of oral squamous cell carcinoma. Biomater. Sci. 2021, 9, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, C.; Ruan, Z.; Xu, P.; Yang, X.; Yuan, P.; Wang, Q.; Yan, L. Polypeptide-conjugated second near-infrared organic fluorophore for image-guided photothermal therapy. ACS Nano 2019, 13, 3691–3702. [Google Scholar] [CrossRef] [PubMed]
- Duhamel, J.; Kanagalingam, S.; O’Brie, T.J.; Ingratta, M.W. Side-chain dynamics of an α-helical polypeptide monitored by fluorescence. J. Am. Chem. Soc. 2003, 125, 12810–12822. [Google Scholar] [CrossRef]
- Tan, K.; Li, R.; Huang, X.; Liu, Q. Outer membrane vesicles: Current status and future direction of these novel vaccine adjuvants. Front. Microbiol. 2018, 9, 783. [Google Scholar] [CrossRef] [Green Version]
- Bielig, H.; Rompikuntal, P.; Dongre, M.; Zurek, B.; Lindmark, B.; Ramstedt, M.; Wai, S.; Kufer, T. NOD-like receptor activation by outer membrane vesicles from Vibrio cholerae non-O1 non-O139 strains is modulated by the quorum-sensing regulator HapR. Infect. Immun. 2011, 79, 1418–1427. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.P.; Jeong, Y.I.; John, J.V.; Chung, C.-W.; Kang, D.H.; Selvaraj, M.; Suh, H.; Kim, I. Dual stimuli-responsive poly(N-isopropylacrylamide)-b-poly(l-histidine) chimeric materials for the controlled delivery of doxorubicin into liver carcinoma. Biomacromolecules 2013, 14, 1434–1443. [Google Scholar] [CrossRef]
- Liu, J.; Huang, Y.; Kumar, A.; Tan, A.; Jin, S.; Mozhi, A.; Liang, X.-J. pH-Sensitive nano-systems for drug delivery in cancer therapy. Biotechnol. Adv. 2014, 32, 693–710. [Google Scholar] [CrossRef] [PubMed]
- Bazban-Shotorbani, S.; Hasani-Sadrabadi, M.M.; Karkhaneh, A.; Serpooshan, V.; Jacob, K.I.; Moshaverinia, A.; Mahmoudi, M. Revisiting structure-property relationship of pH-responsive polymers for drug delivery applications. J. Control. Release 2017, 253, 46–63. [Google Scholar] [CrossRef]
- He, T.; Li, D.; Yang, Y.; Ding, J.; Jin, F.; Zhuang, X.; Chen, X. Mesomeric configuration makes polyleucine micelle an optimal nanocarrier. Biomater. Sci. 2016, 4, 814–818. [Google Scholar] [CrossRef]
- Zhang, Y.; Uthaman, S.; Song, W.; Eom, K.H.; Jeon, S.H.; Huh, K.M.; Babu, A.; Park, I.-K.; Kim, I. Multistimuli-responsive polymeric vesicles for accelerated drug release in chemo-photothermal therapy. ACS Biomater. Sci. Eng. 2020, 6, 5012–5023. [Google Scholar] [CrossRef]
- Allan, N.D.; Beveridge, T.J. Gentamicin delivery to burkholderia cepacia group IIIa strains via membrane vesicles from pseudomonas aeruginosa PAO1. Antimicrob. Agents Chemother. 2003, 47, 2962–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirbin, S.J.; Ladewig, K.; Fu, Q.; Klimak, M.; Zhang, X.; Duan, W.; Qiao, G.G. Cisplatin-induced formation of biocompatible and biodegradable polypeptide-based vesicles for targeted anticancer drug delivery. Biomacromolecules 2015, 16, 2463–2474. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Gupta, M.K.; Bang, J.B.; Bae, H.; Sung, H.J. Current progress in reactive oxygen species (ROS)-responsive materials for biomedical applications. Adv. Healthc. Mater. 2013, 2, 908–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.P.; Jeong, Y.-I.; Choi, E.; Chung, C.-W.; Kang, D.H.; Oh, S.-O.; Suh, H.; Kim, I. Biocompatible poly(2-hydroxyethyl methacrylate)-b-poly(L-histidine) hybrid materials for pH-sensitive intracellular anticancer drug delivery. Adv. Funct. Mater. 2012, 22, 1058–1068. [Google Scholar] [CrossRef]
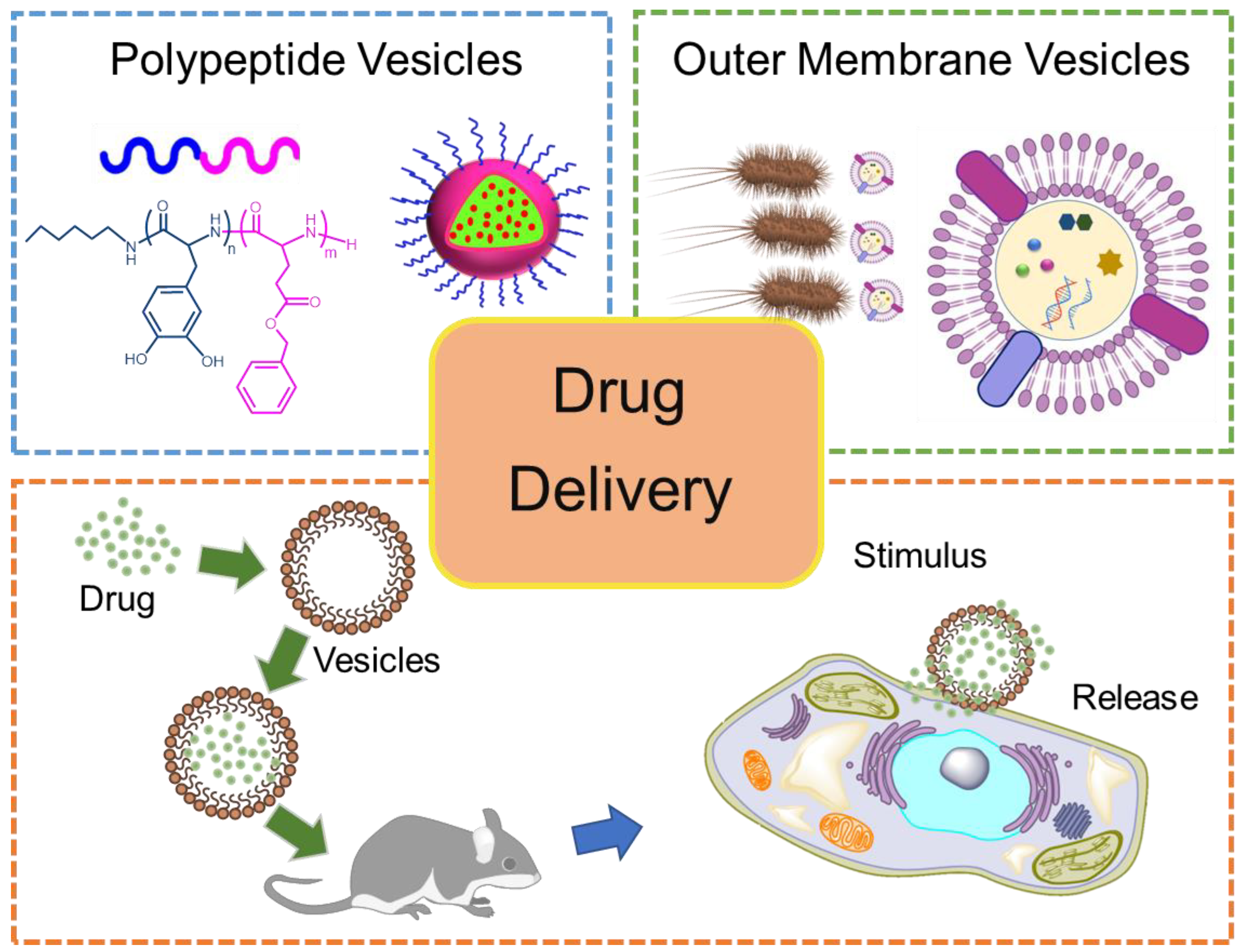
Figure 1.
Schematic illustration of PVs and OMVs, along with their drug delivery properties.

Figure 2.
Schematic illustration of polypeptide formation mechanism, (a) normal amine mechanism and (b) active monomer mechanism. Reprinted with permission from Ref. [32]. Copyright: 1997, John Wiley & Sons, Inc.
Figure 2.
Schematic illustration of polypeptide formation mechanism, (a) normal amine mechanism and (b) active monomer mechanism. Reprinted with permission from Ref. [32]. Copyright: 1997, John Wiley & Sons, Inc.

Figure 3.
Schematic illustration of polypeptide vesicle formation. (A) a-helical polypeptide segments and (B) PVs from two polymers containing charged polypeptide segments with opposite charges. Reprinted with permission from Ref. [14]. Copyright: 2017, The Royal Society of Chemistry.
Figure 3.
Schematic illustration of polypeptide vesicle formation. (A) a-helical polypeptide segments and (B) PVs from two polymers containing charged polypeptide segments with opposite charges. Reprinted with permission from Ref. [14]. Copyright: 2017, The Royal Society of Chemistry.

Figure 4.
Biogenesis of OMV production in bacteria. Reprinted with permission from Ref. [44]. Copyright: 2021, John Wiley & Sons, Inc.
Figure 4.
Biogenesis of OMV production in bacteria. Reprinted with permission from Ref. [44]. Copyright: 2021, John Wiley & Sons, Inc.

Figure 5.
Drug loading in PVs. (a) p(PEGA)30-b-p(Lys)25-b-p(His)n assembled into vesicles with DOX loading. Reprinted with permission from Ref. [74]. Copyright: 2015, American Chemical Society. (b) DACHPt loading into PEG-b-PLG copolymers. Reprinted with permission from Ref. [77]. Copyright: 2015, Elsevier.
Figure 5.
Drug loading in PVs. (a) p(PEGA)30-b-p(Lys)25-b-p(His)n assembled into vesicles with DOX loading. Reprinted with permission from Ref. [74]. Copyright: 2015, American Chemical Society. (b) DACHPt loading into PEG-b-PLG copolymers. Reprinted with permission from Ref. [77]. Copyright: 2015, Elsevier.

Figure 6.
Drug loading in OMVs. (a) Loading of siRNA into AffiHER2OMVs. Reprinted with permission from Ref. [55]. Copyright: 2014, American Chemical Society. (b) DOX can be loaded into OMVs via passive diffusion. Reprinted with permission from Ref. [76]. Copyright: 2020, Elsevier.

Figure 7.
Targeted drug delivery via PVs and OMVs. (a) Drug release ability of cisplatin complexed with the PEG-b-PLG. Reprinted with permission from Ref. [103]. Copyright: 2015, American Chemical Society. (b) Schematic diagram of bioengineered OMV-coated polymeric nanomedicine. Reprinted with permission from Ref. [83]. Copyright: 2020, American Chemical Society.
Figure 7.
Targeted drug delivery via PVs and OMVs. (a) Drug release ability of cisplatin complexed with the PEG-b-PLG. Reprinted with permission from Ref. [103]. Copyright: 2015, American Chemical Society. (b) Schematic diagram of bioengineered OMV-coated polymeric nanomedicine. Reprinted with permission from Ref. [83]. Copyright: 2020, American Chemical Society.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Comparison of characteristics of PVs and OMVs.
| Classification | PVs | OMVs | Reference |
|---|---|---|---|
| Origin | Synthetic polymers | Gram-negative bacteria | [13,17] |
| Production | Solution self-assembly | Biogenesis | [15,21] |
| Size | Several nanometers~micrometers | 20~300 nm | [14,27] |
| Biomedical applications | Antibacterial | Bacterial vaccines | [14,28] |
| Mineralization | Adjuvants | ||
| Drug delivery | Cancer immunotherapy | ||
| Imaging and sensing | Drug delivery | ||
| Therapeutic | Anti-bacteria adhesion |
Table 2.
Comparison of drug loading and releasing by PVs and OMVs.
| DDS | PVs | References | OMVs | References |
|---|---|---|---|---|
| Drug loading | Hydrophobic interaction | [73,74] | Electroporation | [55] |
| Electrostatic interactions | [75] | Passive diffusion | [76] | |
| Metal coordination | [77] | Biogenesis | [78,79] | |
| Drug releasing | Change peptide assembly behavior | [80,81] | Degradation of OMVs | [55] |
| Change drug–polypeptide interaction | [82] | Coating on polymer | [83] | |
| Bioengineering | [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Zhang, Y.; Lu, Y.; Xu, Y.; Zhou, Z.; Li, Y.; Ling, W.; Song, W. Bio-Inspired Drug Delivery Systems: From Synthetic Polypeptide Vesicles to Outer Membrane Vesicles. Pharmaceutics 2023, 15, 368. https://doi.org/10.3390/pharmaceutics15020368
AMA Style
Zhang Y, Lu Y, Xu Y, Zhou Z, Li Y, Ling W, Song W. Bio-Inspired Drug Delivery Systems: From Synthetic Polypeptide Vesicles to Outer Membrane Vesicles. Pharmaceutics. 2023; 15(2):368. https://doi.org/10.3390/pharmaceutics15020368
Chicago/Turabian StyleZhang, Yu, Yiming Lu, Yixin Xu, Zunkang Zhou, Yichong Li, Wei Ling, and Wenliang Song. 2023. "Bio-Inspired Drug Delivery Systems: From Synthetic Polypeptide Vesicles to Outer Membrane Vesicles" Pharmaceutics 15, no. 2: 368. https://doi.org/10.3390/pharmaceutics15020368
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.