
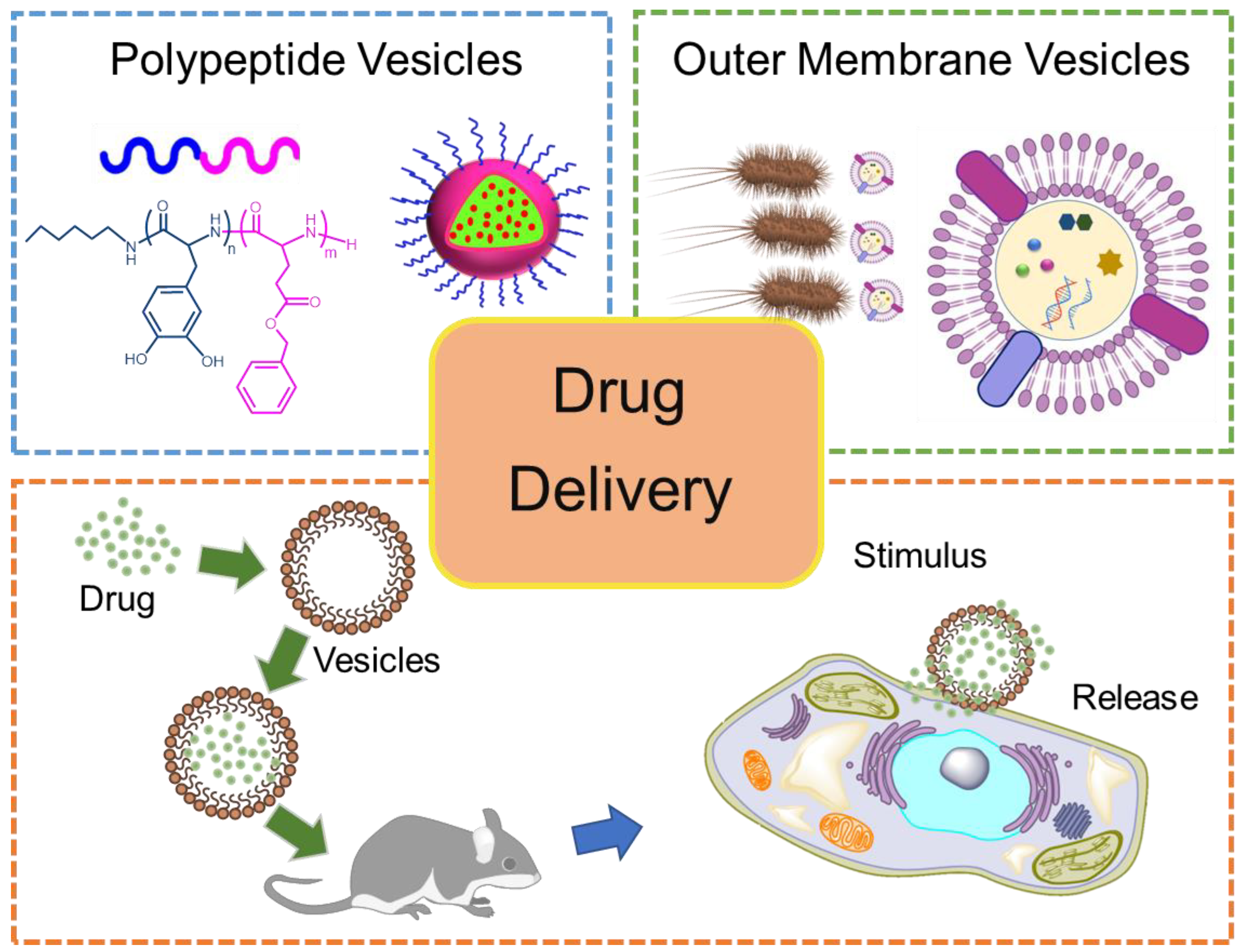
Bio-Inspired Drug Delivery Systems: From Synthetic Polypeptide Vesicles to Outer Membrane Vesicles

Abstract
:1. Introduction
2. Production and Characterization of PVs and OMVs
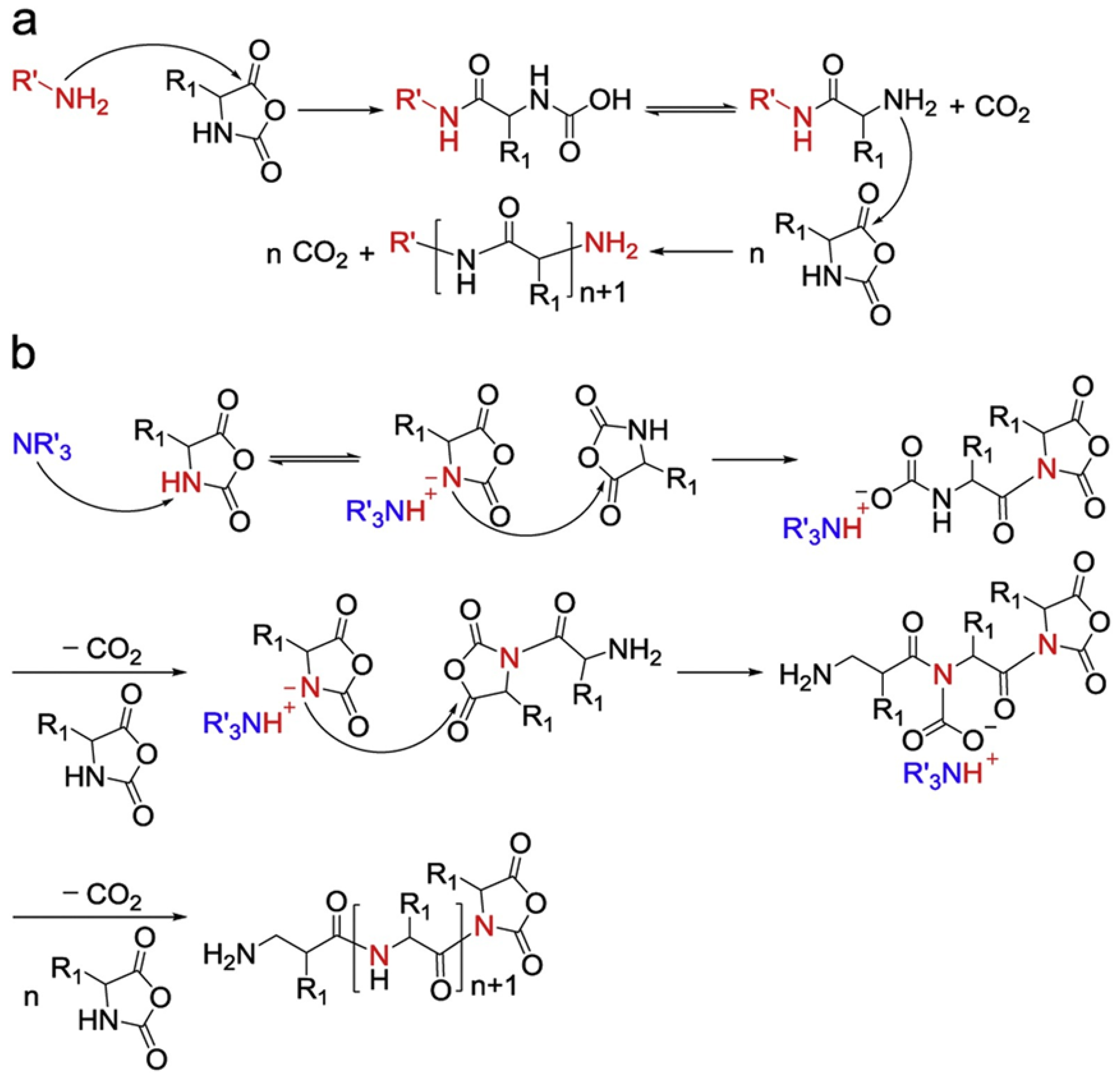
2.1. Production of PVs
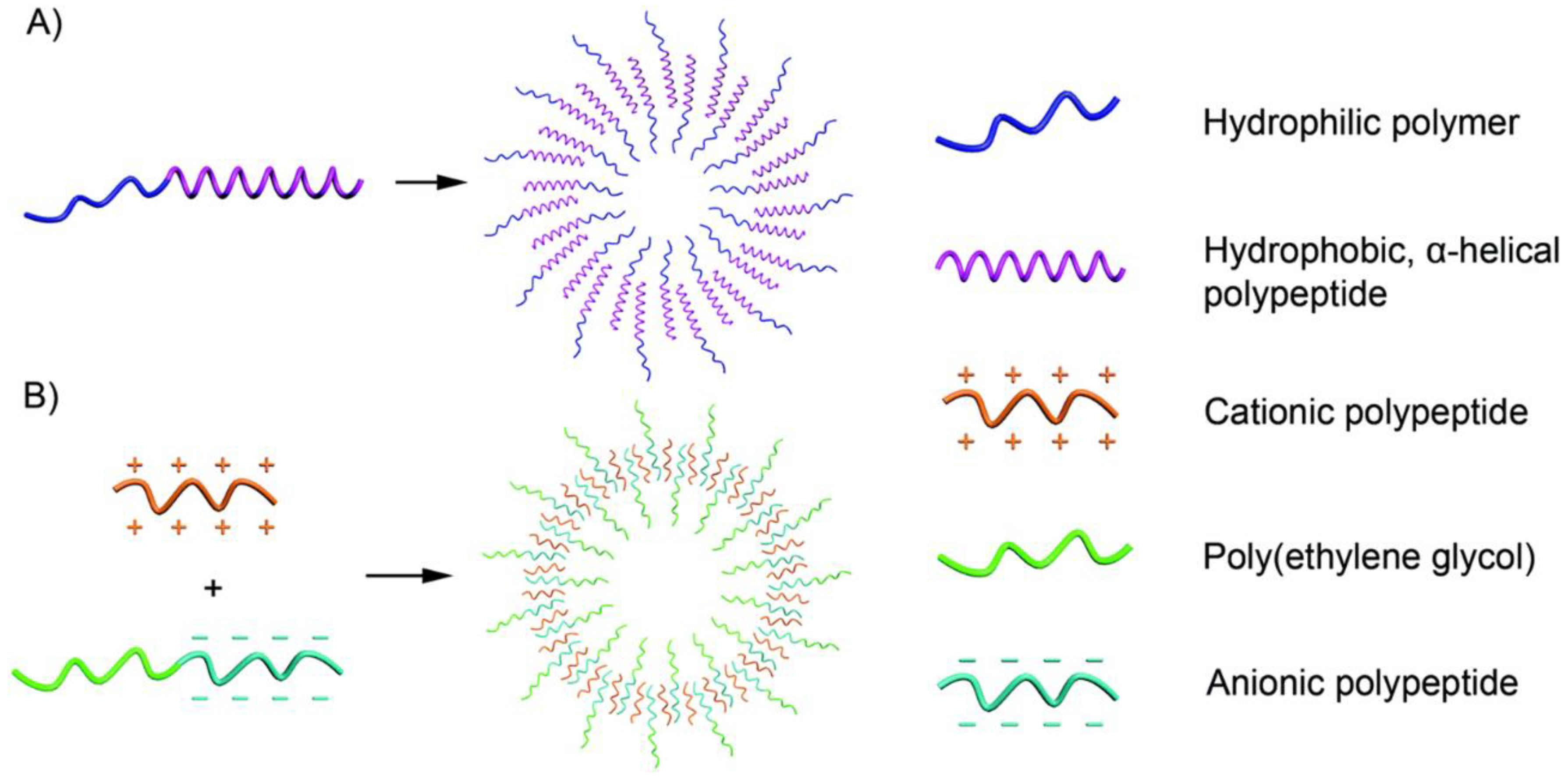
2.1.1. Anisotropic Packing of α-Helical, Hydrophobic Polypeptides
2.1.2. Oppositely Charged Polymers
2.1.3. Others
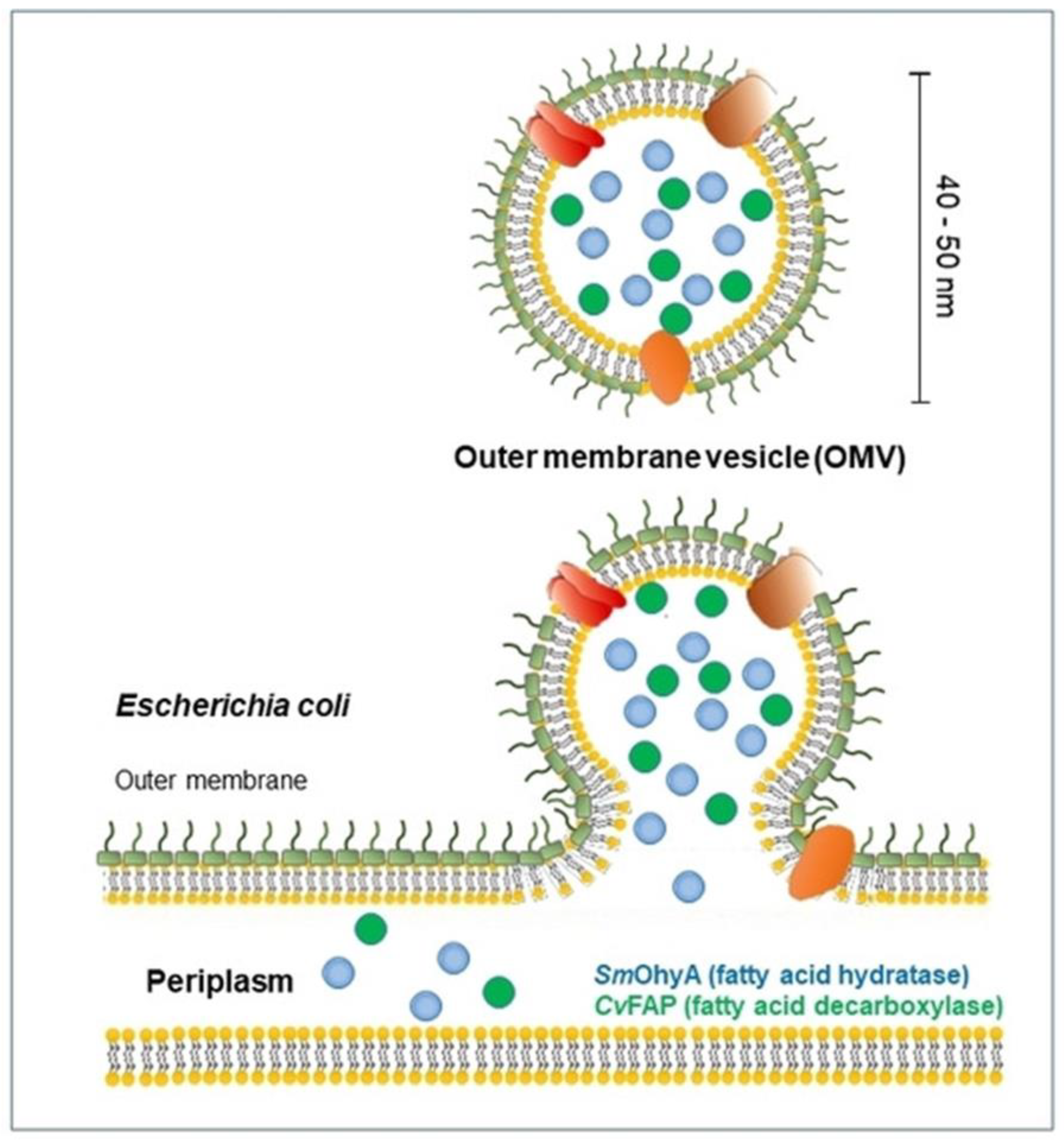
2.2. Production of OMVs
2.2.1. Reduced Lipoprotein Linkages
2.2.2. Peptidoglycan Residues
2.2.3. Electrically Charged Lipopolysaccharides (LPSs)
2.2.4. Other Mechanisms
3. Drug Delivery Performance of PVs and OMVs
3.1. Advantages of PVs and OMVs in DDSs
3.2. Techniques for Drug Loading in PVs and OMVs
3.2.1. Drug Loading in PVs
3.2.2. Drug Loading in OMVs
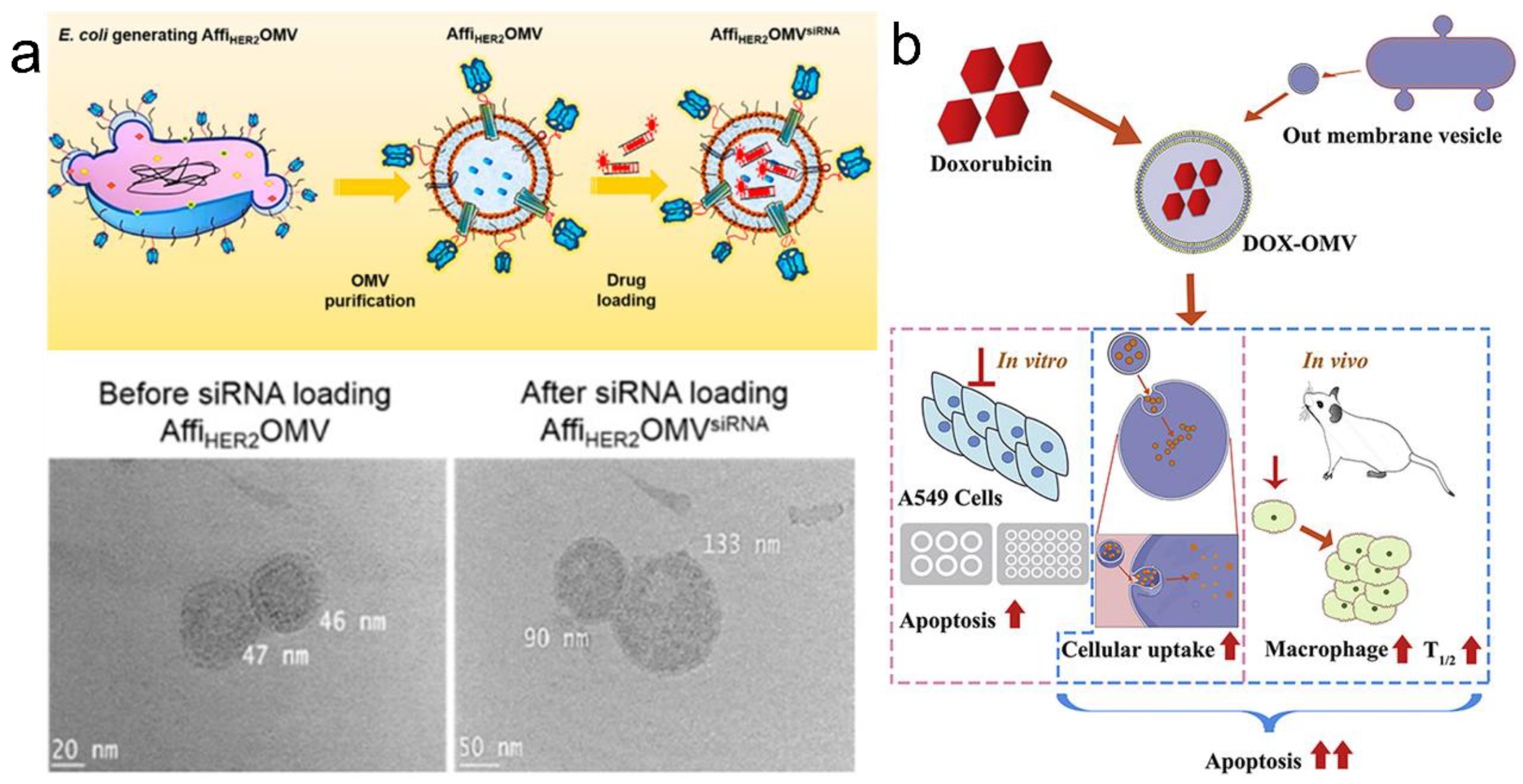
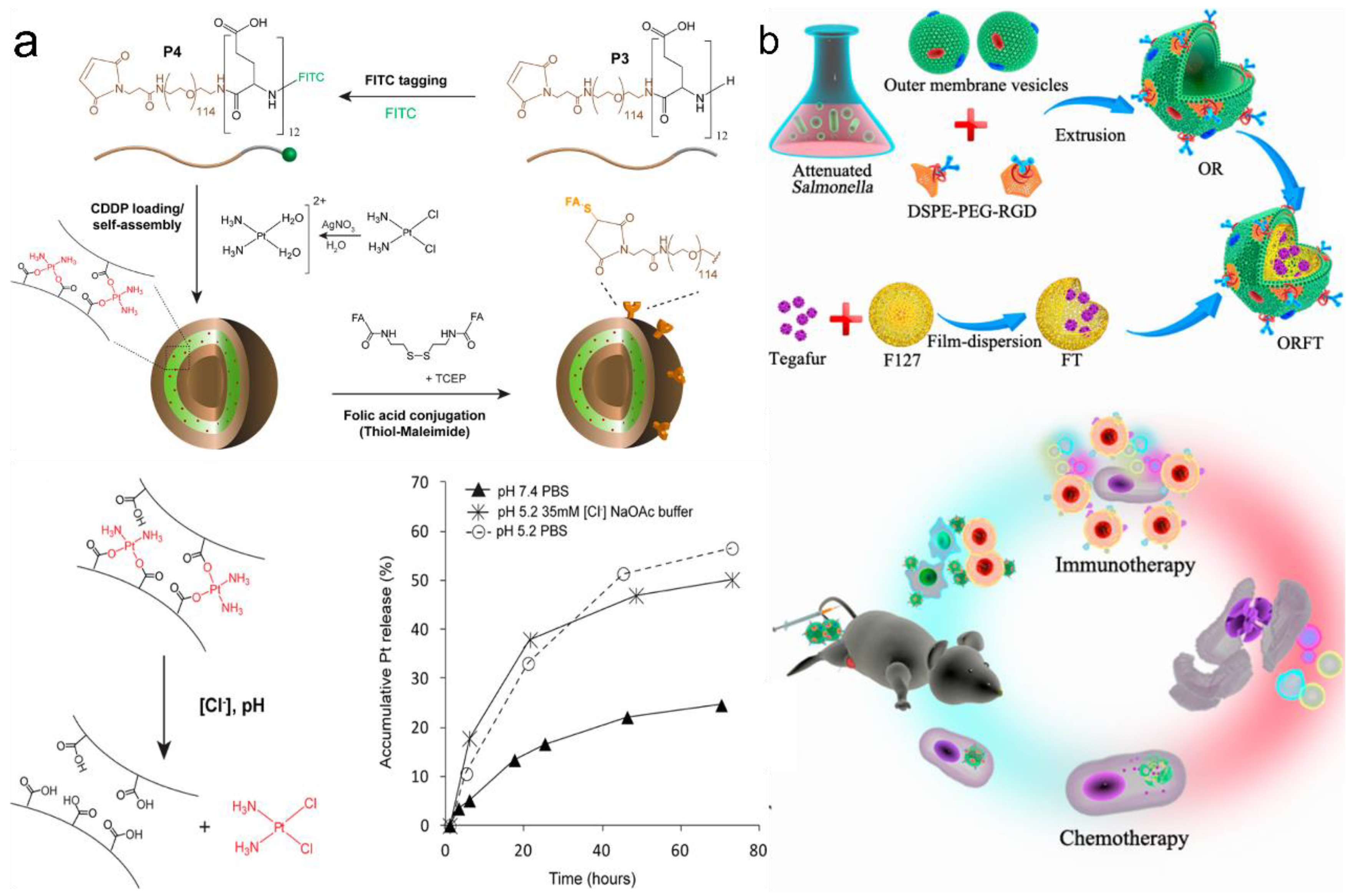
3.3. Targeted Drug Delivery via PVs and OMVs
3.3.1. Targeted Drug Delivery via PVs
3.3.2. Targeted Drug Delivery via OMVs
4. Conclusions and Outlook
- (1)
- Mass production of OMVs and PVs: Currently, the production of OMVs and PVs is still in the laboratory research stage. Although, recently, the production efficiency of polypeptide materials has been rapidly improved, technical solutions for assembling vesicles still lack specific standards, and large-scale production of PVs is not yet possible. OMV sizes are difficult to regulate, and there is significant variance among batches. In addition, low particle programmability is caused by our lack of knowledge regarding the precise OMV assembly mechanism. Enhancing safety and reducing costs are also still problems that need to be addressed.
- (2)
- Universality of drug loading and delivery targeting: The drug loading mechanism in the process of production and post-treatment is inefficient and is in urgent need of optimization. Given the relevance of various cancer types and given that there are various PV and OMV types, consistent tumor cell targeting standards still need to be achieved and further improved. Individual PVs and OMVs may be effective for certain cancers.
- (3)
- Other specific issues: The immunotoxicity of OMVs is a problem that cannot be ignored in DDSs. Moreover, the artificial production of OMVs also deserves more in-depth study on its potential. In PVs, the deprotection process in the production of specific polypeptides (such as polyhistidine, polylysine, and polycysteine) is tedious. Finally, programmed drug release and the benefits of the unique secondary structure of PVs are also worthy of more in-depth research.
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, J.; Yu, F.; Chen, Y.; Oupický, D. Polymeric drugs: Advances in the development of pharmacologically active polymers. J. Control. Release 2015, 219, 369–382. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gale, G.R.; Morris, C.R.; Atkins, L.M.; Smith, A.B. Binding of an antitumor platinum compound to cells as influenced by physical factors and pharmacologically active agents. Cancer Res. 1973, 33, 813–818. [Google Scholar] [PubMed]
- Xian, C.; Chen, H.; Xiong, F.; Fang, Y.; Huang, H.; Wu, J. Platinum-based chemotherapy via nanocarriers and co-delivery of multiple drugs. Biomater. Sci. 2021, 9, 6023–6036. [Google Scholar] [CrossRef] [PubMed]
- Tabakoglu, S.; Kołbuk, D.; Sajkiewicz, P. Multifluid electrospinning for multi-drug delivery systems: Pros and cons, challenges, and future directions. Biomater. Sci. 2022, 11, 37–61. [Google Scholar] [CrossRef] [PubMed]
- Tibbitt, M.W.; Dahlman, J.E.; Langer, R. Emerging frontiers in drug delivery. J. Am. Chem. Soc. 2016, 138, 704–717. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Gao, J.; Wang, Z. Outer membrane vesicles for vaccination and targeted drug delivery. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2019, 11, e1523. [Google Scholar] [CrossRef] [PubMed]
- Loo, Y.S.; Bose, R.J.; McCarthy, J.R.; Azmi, I.D.M.; Madheswaran, T. Biomimetic bacterial and viral-based nanovesicles for drug delivery, theranostics, and vaccine applications. Drug Discov. Today 2021, 26, 902–915. [Google Scholar] [CrossRef]
- Al-Dossary, A.A.; Isichei, A.C.; Zhang, S.; Li, J.; Errachid, A.; Elaissari, A. Outer membrane vesicles (OMVs) as a platform for vaccination and targeted drug delivery. In Pharmaceutical Nanobiotechnology for Targeted Therapy; Springer: Berlin/Heidelberg, Germany, 2022; pp. 1–25. [Google Scholar]
- Tang, Y.; Varyambath, A.; Ding, Y.; Chen, B.; Huang, X.; Zhang, Y.; Yu, D.-g.; Kim, I.; Song, W. Porous organic polymers for drug delivery: Hierarchical pore structures, variable morphologies, and biological properties. Biomater. Sci. 2022, 10, 5369–5390. [Google Scholar] [CrossRef]
- Wang, Y.; Lu, X.; Wu, X.; Li, Y.; Tang, W.; Yang, C.; Ding, B. Chemically modified DNA nanostructures for drug delivery. Innovation 2022, 3, 100217. [Google Scholar] [CrossRef]
- Song, W.; Zhang, M.; Huang, X.; Chen, B.; Ding, Y.; Zhang, Y.; Yu, D.G.; Kim, I. Smart l-borneol-loaded hierarchical hollow polymer nanospheres with antipollution and antibacterial capabilities. Mater. Today Chem. 2022, 26, 101252. [Google Scholar] [CrossRef]
- Zhou, Y.; Wang, M.; Yan, C.; Liu, H.; Yu, D.-G. Advances in the application of electrospun drug-loaded nanofibers in the treatment of oral ulcers. Biomolecules 2022, 12, 1254. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Liu, R.; Jin, H.; Song, W.; Augustine, R.; Kim, I. Straightforward access to linear and cyclic polypeptides. Commun. Chem. 2018, 1, 40. [Google Scholar] [CrossRef] [Green Version]
- Song, Z.; Han, Z.; Lv, S.; Chen, C.; Chen, L.; Yin, L.; Cheng, J. Synthetic polypeptides: From polymer design to supramolecular assembly and biomedical application. Chem. Soc. Rev. 2017, 46, 6570–6599. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Song, Z.; Wei, S.; Ji, G.; Zheng, X.; Fu, Z.; Cheng, J. Polypeptide-based drug delivery systems for programmed release. Biomaterials 2021, 275, 120913. [Google Scholar] [CrossRef] [PubMed]
- Malonis, R.J.; Lai, J.R.; Vergnolle, O. Peptide-based vaccines: Current progress and future challenges. Chem. Rev. 2019, 120, 3210–3229. [Google Scholar] [CrossRef] [Green Version]
- Li, R.; Liu, Q. Engineered bacterial outer membrane vesicles as multifunctional delivery platforms. Front. Mater. 2020, 7, 202. [Google Scholar] [CrossRef]
- Chen, W.; Wu, Y.; Deng, J.; Yang, Z.; Chen, J.; Tan, Q.; Guo, M.; Jin, Y. Phospholipid-membrane-based nanovesicles acting as vaccines for tumor immunotherapy: Classification, mechanisms and applications. Pharmaceutics 2022, 14, 2446. [Google Scholar] [CrossRef]
- Srivastava, P.; Kim, K.-S. Membrane vesicles derived from gut microbiota and probiotics: Cutting-edge therapeutic approaches for multidrug-resistant superbugs linked to neurological anomalies. Pharmaceutics 2022, 14, 2370. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, L.; Fu, H.; Wei, C.; Jin, Q. Comparative proteomic analysis of outer membrane vesicles from Shigella flexneri under different culture conditions. Biochem. Biophys. Res. Commun. 2014, 453, 696–702. [Google Scholar] [CrossRef]
- Avila-Calderón, E.D.; Ruiz-Palma, M.D.S.; Aguilera-Arreola, M.G.; Velázquez-Guadarrama, N.; Ruiz, E.A.; Gomez-Lunar, Z.; Witonsky, S.; Contreras-Rodríguez, A. Outer membrane vesicles of gram-negative bacteria: An outlook on biogenesis. Front. Microbiol. 2021, 12, 557902. [Google Scholar] [CrossRef]
- Gao, W.; Fang, R.H.; Thamphiwatana, S.; Luk, B.T.; Li, J.; Angsantikul, P.; Zhang, Q.; Hu, C.-M.J.; Zhang, L. Modulating antibacterial immunity via bacterial membrane-coated nanoparticles. Nano Lett. 2015, 15, 1403–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, L.; Praszkier, J.; Hutton, M.L.; Steer, D.; Ramm, G.; Kaparakis-Liaskos, M.; Ferrero, R.L. Increased outer membrane vesicle formation in a Helicobacter pylori tolB mutant. Helicobacter 2015, 20, 269–283. [Google Scholar] [CrossRef] [PubMed]
- Tsai, S.-L.; Oh, J.; Singh, S.; Chen, R.; Chen, W. Functional assembly of minicellulosomes on the Saccharomyces cerevisiae cell surface for cellulose hydrolysis and ethanol production. Appl. Environ. Microbiol. 2009, 75, 6087–6093. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bauman, S.J.; Kuehn, M.J. Purification of outer membrane vesicles from Pseudomonas aeruginosa and their activation of an IL-8 response. Microbes Infect. 2006, 8, 2400–2408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chatterjee, D.; Chaudhuri, K. Association of cholera toxin with Vibrio cholerae outer membrane vesicles which are internalized by human intestinal epithelial cells. FEBS Lett. 2011, 585, 1357–1362. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhou, H.; Yang, C.; Wu, Y.; Zhou, X.; Liu, H.; Wang, Y. Bacterial outer membrane vesicles as a platform for biomedical applications: An update. J. Control. Release 2020, 323, 253–268. [Google Scholar] [CrossRef]
- Huang, Y.; Nieh, M.P.; Chen, W.; Lei, Y. Outer membrane vesicles (OMVs) enabled bio-applications: A critical review. Biotechnol. Bioeng. 2022, 119, 34–47. [Google Scholar] [CrossRef]
- Zhang, Y.; Kim, I.; Lu, Y.; Xu, Y.; Yu, D.-G.; Song, W. Intelligent poly(l-histidine)-based nanovehicles for controlled drug delivery. J. Control. Release 2022, 349, 963–982. [Google Scholar] [CrossRef]
- Leuchs, H.; Geiger, W. The anhydrides of alpha-amino-N-carboxylic and alpha-amino acids. Ber. Dtsch. Chem. Ges. 1908, 41, 1721–1726. [Google Scholar] [CrossRef] [Green Version]
- Liu, Y.; Li, D.; Ding, J.; Chen, X. Controlled synthesis of polypeptides. Chin. Chem. Lett. 2020, 31, 3001–3014. [Google Scholar] [CrossRef]
- Deming, T.J. Polypeptide Materials: New synthetic methods and applications. Adv. Mater. 1997, 9, 299–311. [Google Scholar] [CrossRef]
- Holowka, E.P.; Pochan, D.J.; Deming, T.J. Charged Polypeptide vesicles with controllable diameter. J. Am. Chem. Soc. 2005, 127, 12423–12428. [Google Scholar] [CrossRef] [PubMed]
- Sun, V.Z.; Choe, U.J.; Rodriguez, A.R.; Dai, H.; Deming, T.J.; Kamei, D.T. Transfection of mammalian cells using block copolypeptide vesicles. Macromol. Biosci. 2013, 13, 539–550. [Google Scholar] [CrossRef] [PubMed]
- Choe, U.-J.; Rodriguez, A.R.; Lee, B.S.; Knowles, S.M.; Wu, A.M.; Deming, T.J.; Kamei, D.T. Endocytosis and intracellular trafficking properties of transferrin-conjugated block copolypeptide vesicles. Biomacromolecules 2013, 14, 1458–1464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.R.; Choe, U.-J.; Kamei, D.T.; Deming, T.J. Blending of diblock and triblock copolypeptide amphiphiles yields cell penetrating vesicles with low toxicity. Macromol. Biosci. 2015, 15, 90–97. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Bonduelle, C.; Thévenot, J.; Lecommandoux, S.; Heise, A. Biologically active polymersomes from amphiphilic glycopeptides. J. Am. Chem. Soc. 2012, 134, 119–122. [Google Scholar] [CrossRef]
- Yu, K.; Eisenberg, A. Bilayer morphologies of self-assembled crew-cut aggregates of amphiphilic PS-b-PEO diblock copolymers in solution. Macromolecules 1998, 31, 3509–3518. [Google Scholar] [CrossRef]
- Yang, Y.-Y.; Chen, L.-S.; Sun, M.; Wang, C.-Y.; Fan, Z.; Du, J.-Z. Biodegradable polypeptide-based vesicles with intrinsic blue fluorescence for antibacterial visualization. Chin. J. Polym. Sci. 2021, 39, 1412–1420. [Google Scholar] [CrossRef]
- Xi, Y.; Wang, Y.; Gao, J.; Xiao, Y.; Du, J. Dual Corona vesicles with intrinsic antibacterial and enhanced antibiotic delivery capabilities for effective treatment of biofilm-induced periodontitis. ACS Nano 2019, 13, 13645–13657. [Google Scholar] [CrossRef]
- Sanson, C.; Schatz, C.; Le Meins, J.-F.; Soum, A.; Thévenot, J.; Garanger, E.; Lecommandoux, S. A simple method to achieve high doxorubicin loading in biodegradable polymersomes. J. Control. Release 2010, 147, 428–435. [Google Scholar] [CrossRef]
- Knox, K.; Vesk, M.; Work, E. Relation between excreted lipopolysaccharide complexes and surface structures of a lysine-limited culture of Escherichia coli. J. Bacteriol. 1966, 92, 1206–1217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katsui, N.; Tsuchido, T.; Hiramatsu, R.; Fujikawa, S.; Takano, M.; Shibasaki, I. Heat-induced blebbing and vesiculation of the outer membrane of Escherichia coli. J. Bacteriol. 1982, 151, 1523–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, J.-W.; Baeg, Y.; Jeong, H.-Y.; Lee, J.; Oh, D.-K.; Hollmann, F.; Park, J.-B. Bacterial outer membrane vesicles as nano-scale bioreactors: A fatty acid conversion case study. ChemCatChem 2021, 13, 4080–4086. [Google Scholar] [CrossRef]
- Hoekstra, D.; van der Laan, J.W.; de Leij, L.; Witholt, B. Release of outer membrane fragments from normally growing Escherichia coli. Biochim. Biophys. Acta (BBA) Biomembr. 1976, 455, 889–899. [Google Scholar] [CrossRef]
- Schwechheimer, C.; Rodriguez, D.L.; Kuehn, M.J. NlpI-mediated modulation of outer membrane vesicle production through peptidoglycan dynamics in Escherichia coli. MicrobiologyOpen 2015, 4, 375–389. [Google Scholar] [CrossRef] [Green Version]
- Pathirana, R.D.; Kaparakis-Liaskos, M. Bacterial membrane vesicles: Biogenesis, immune regulation and pathogenesis. Cell. Microbiol. 2016, 18, 1518–1524. [Google Scholar] [CrossRef] [Green Version]
- McBroom, A.J.; Kuehn, M.J. Release of outer membrane vesicles by gram-negative bacteria is a novel envelope stress response. Mol. Microbiol. 2007, 63, 545–558. [Google Scholar] [CrossRef]
- Kadurugamuwa, J.L.; Beveridge, T.J. Virulence factors are released from Pseudomonas aeruginosa in association with membrane vesicles during normal growth and exposure to gentamicin: A novel mechanism of enzyme secretion. J. Bacteriol. 1995, 177, 3998–4008. [Google Scholar] [CrossRef] [Green Version]
- McMahon, K.J.; Castelli, M.E.; Vescovi, E.G.; Feldman, M.F. Biogenesis of outer membrane vesicles in Serratia marcescens is thermoregulated and can be induced by activation of the Rcs phosphorelay system. J. Bacteriol. 2012, 194, 3241–3249. [Google Scholar] [CrossRef] [Green Version]
- Elhenawy, W.; Bording-Jorgensen, M.; Valguarnera, E.; Haurat, M.F.; Wine, E.; Feldman, M.F. LPS remodeling triggers formation of outer membrane vesicles in Salmonella. mBio 2016, 7, e00940-16. [Google Scholar] [CrossRef]
- Turnbull, L.; Toyofuku, M.; Hynen, A.L.; Kurosawa, M.; Pessi, G.; Petty, N.K.; Osvath, S.R.; Cárcamo-Oyarce, G.; Gloag, E.S.; Shimoni, R.; et al. Explosive cell lysis as a mechanism for the biogenesis of bacterial membrane vesicles and biofilms. Nat. Commun. 2016, 7, 11220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roier, S.; Zingl, F.G.; Cakar, F.; Durakovic, S.; Kohl, P.; Eichmann, T.O.; Klug, L.; Gadermaier, B.; Weinzerl, K.; Prassl, R.; et al. A novel mechanism for the biogenesis of outer membrane vesicles in Gram-negative bacteria. Nat. Commun. 2016, 7, 10515. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Song, W.; Lu, Y.; Xu, Y.; Wang, C.; Yu, D.-G.; Kim, I. Recent advances in poly(alpha-L-glutamic acid)-based nanomaterials for drug delivery. Biomolecules 2022, 12, 636. [Google Scholar] [CrossRef] [PubMed]
- Gujrati, V.; Kim, S.; Kim, S.-H.; Min, J.J.; Choy, H.E.; Kim, S.C.; Jon, S. Bioengineered bacterial outer membrane vesicles as cell-specific drug-delivery vehicles for cancer therapy. ACS Nano 2014, 8, 1525–1537. [Google Scholar] [CrossRef]
- Yin, L.; Cheng, J.; Deming, T.J.; Vicent, M.J. Synthetic polypeptides for drug and gene delivery, and tissue engineering. Adv. Drug Delivery Rev. 2021, 178, 113995. [Google Scholar] [CrossRef] [PubMed]
- Lv, S.; Song, K.; Yen, A.; Peeler, D.J.; Nguyen, D.C.; Olshefsky, A.; Sylvestre, M.; Srinivasan, S.; Stayton, P.S.; Pun, S.H. Well-defined mannosylated polymer for peptide vaccine delivery with enhanced antitumor immunity. Adv. Healthc. Mater. 2022, 11, 2101651. [Google Scholar] [CrossRef] [PubMed]
- O’Neill, C.L.; Shrimali, P.C.; Clapacs, Z.P.; Files, M.A.; Rudra, J.S. Peptide-based supramolecular vaccine systems. Acta Biomater. 2021, 133, 153–167. [Google Scholar] [CrossRef]
- van der Pol, L.; Stork, M.; van der Ley, P. Outer membrane vesicles as platform vaccine technology. Biotechnol. J. 2015, 10, 1689–1706. [Google Scholar] [CrossRef] [Green Version]
- Kim, O.Y.; Park, H.T.; Dinh, N.T.H.; Choi, S.J.; Lee, J.; Kim, J.H.; Lee, S.-W.; Gho, Y.S. Bacterial outer membrane vesicles suppress tumor by interferon-γ-mediated antitumor response. Nat. Commun. 2017, 8, 626. [Google Scholar] [CrossRef] [Green Version]
- Ding, Y.; Ma, Y.; Du, C.; Wang, C.; Chen, T.; Wang, Y.; Wang, J.; Yao, Y.; Dong, C.-M. NO-releasing polypeptide nanocomposites reverse cancer multidrug resistance via triple therapies. Acta Biomater. 2021, 123, 335–345. [Google Scholar] [CrossRef]
- Wang, Q.; Jiang, N.; Fu, B.; Huang, F.; Liu, J. Self-assembling peptide-based nanodrug delivery systems. Biomater. Sci. 2019, 7, 4888–4911. [Google Scholar] [CrossRef] [PubMed]
- Qi, G.B.; Gao, Y.J.; Wang, L.; Wang, H. Self-assembled peptide-based nanomaterials for biomedical imaging and therapy. Adv. Mater. 2018, 30, 1703444. [Google Scholar] [CrossRef] [PubMed]
- Li, H.; Huang, H.; Feng, J.-J.; Luo, X.; Fang, K.-M.; Wang, Z.-G.; Wang, A.-J. A polypeptide-mediated synthesis of green fluorescent gold nanoclusters for Fe3+ sensing and bioimaging. J. Colloid Interface Sci. 2017, 506, 386–392. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Wang, D.; Hui, N. A low fouling electrochemical biosensor based on the zwitterionic polypeptide doped conducting polymer PEDOT for breast cancer marker BRCA1 detection. Bioelectrochemistry 2020, 136, 107595. [Google Scholar] [CrossRef] [PubMed]
- Oh, K.J.; Cash, K.J.; Hugenberg, V.; Plaxco, K.W. Peptide Beacons: A new design for polypeptide-based optical biosensors. Bioconjug. Chem. 2007, 18, 607–609. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Song, W.; Li, S.; Kim, D.-K.; Kim, J.H.; Kim, J.R.; Kim, I. Facile and scalable synthesis of topologically nanoengineered polypeptides with excellent antimicrobial activities. Chem. Commun. 2020, 56, 356–359. [Google Scholar] [CrossRef]
- Lu, Z.; Wu, Y.; Cong, Z.; Qian, Y.; Wu, X.; Shao, N.; Qiao, Z.; Zhang, H.; She, Y.; Chen, K. Effective and biocompatible antibacterial surfaces via facile synthesis and surface modification of peptide polymers. Bioact. Mater. 2021, 6, 4531–4541. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.; Chen, K.; Wu, X.; Liu, L.; Zhang, W.; Ding, Y.; Liu, S.; Zhou, M.; Shao, N.; Ji, Z.; et al. Superfast and Water-Insensitive Polymerization on α-amino acid N-carboxyanhydrides to prepare polypeptides using tetraalkylammonium carboxylate as the initiator. Angew. Chem. Int. Ed. 2021, 60, 26063–26071. [Google Scholar] [CrossRef]
- Ding, X.; Duan, S.; Ding, X.; Liu, R.; Xu, F.-J. Versatile antibacterial materials: An emerging arsenal for combatting bacterial pathogens. Adv. Funct. Mater. 2018, 28, 1802140. [Google Scholar] [CrossRef]
- Pizarro-Cerdá, J.; Cossart, P. Bacterial adhesion and entry into host cells. Cell 2006, 124, 715–727. [Google Scholar] [CrossRef]
- Asadi, A.; Razavi, S.; Talebi, M.; Gholami, M. A review on anti-adhesion therapies of bacterial diseases. Infection 2019, 47, 13–23. [Google Scholar] [CrossRef] [PubMed]
- Kakkar, D.; Mazzaferro, S.; Thevenot, J.; Schatz, C.; Bhatt, A.; Dwarakanath, B.S.; Singh, H.; Mishra, A.K.; Lecommandoux, S. Amphiphilic PEO-b-PBLG diblock and PBLG-b-PEO-b-PBLG triblock copolymer based nanoparticles: Doxorubicin loading and in vitro evaluation. Macromol. Biosci. 2015, 15, 124–137. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.P.; Uthaman, S.; John, J.V.; Lee, H.R.; Lee, S.J.; Park, H.; Park, I.-K.; Suh, H.; Kim, I. Poly(PEGA)-b-poly(l-lysine)-b-poly(l-histidine) hybrid vesicles for tumoral pH-triggered intracellular delivery of doxorubicin hydrochloride. ACS Appl. Mater. Interfaces 2015, 7, 21770–21779. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Song, W.; Tang, Z.; Lv, S.; Lin, L.; Sun, H.; Li, Q.; Yang, Y.; Hong, H.; Chen, X. Nanoscaled poly(l-glutamic acid)/doxorubicin-amphiphile complex as pH-responsive drug delivery system for effective treatment of nonsmall cell lung cancer. ACS Appl. Mater. Interfaces 2013, 5, 1781–1792. [Google Scholar] [CrossRef]
- Kuerban, K.; Gao, X.; Zhang, H.; Liu, J.; Dong, M.; Wu, L.; Ye, R.; Feng, M.; Ye, L. Doxorubicin-loaded bacterial outer-membrane vesicles exert enhanced anti-tumor efficacy in non-small-cell lung cancer. Acta Pharm. Sin. B 2020, 10, 1534–1548. [Google Scholar] [CrossRef]
- Ahn, J.; Miura, Y.; Yamada, N.; Chida, T.; Liu, X.; Kim, A.; Sato, R.; Tsumura, R.; Koga, Y.; Yasunaga, M.; et al. Antibody fragment-conjugated polymeric micelles incorporating platinum drugs for targeted therapy of pancreatic cancer. Biomaterials 2015, 39, 23–30. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, A.L.; Fonseca, S.; Miquel-Clopés, A.; Cross, K.; Kok, K.-S.; Wegmann, U.; Gil-Cardoso, K.; Bentley, E.G.; Al Katy, S.H.; Coombes, J.L. Bioengineering commensal bacteria-derived outer membrane vesicles for delivery of biologics to the gastrointestinal and respiratory tract. J. Extracell. Vesicles 2019, 8, 1632100. [Google Scholar] [CrossRef]
- Gujrati, V.; Prakash, J.; Malekzadeh-Najafabadi, J.; Stiel, A.; Klemm, U.; Mettenleiter, G.; Aichler, M.; Walch, A.; Ntziachristos, V. Bioengineered bacterial vesicles as biological nano-heaters for optoacoustic imaging. Nat. Commun. 2019, 10, 1114. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, R.; Wei, J.; Cheng, C.; Zheng, Y.; Pan, Y.; He, X.; Ding, M.; Tan, H.; Fu, Q. Conformation-directed micelle-to-vesicle transition of cholesterol-decorated polypeptide triggered by oxidation. J. Am. Chem. Soc. 2018, 140, 6604–6610. [Google Scholar] [CrossRef]
- Liu, G.; Dong, C.-M. Photoresponsive Poly(S-(o-nitrobenzyl)-l-cysteine)-b-PEO from a l-cysteine N-carboxyanhydride monomer: Synthesis, self-assembly, and phototriggered drug release. Biomacromolecules 2012, 13, 1573–1583. [Google Scholar] [CrossRef]
- Bae, Y.; Fukushima, S.; Harada, A.; Kataoka, K. Design of Environment-Sensitive Supramolecular assemblies for intracellular drug delivery: Polymeric micelles that are responsive to intracellular pH change. Angew. Chem. Int. Ed. 2003, 42, 4640–4643. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Bai, H.; Wu, W.; Huang, G.; Li, Y.; Wu, M.; Tang, G.; Ping, Y. Bioengineering bacterial vesicle-coated polymeric nanomedicine for enhanced cancer immunotherapy and metastasis prevention. Nano Lett. 2020, 20, 11–21. [Google Scholar] [CrossRef] [PubMed]
- Schetters, S.T.T.; Jong, W.S.P.; Horrevorts, S.K.; Kruijssen, L.J.W.; Engels, S.; Stolk, D.; Daleke-Schermerhorn, M.H.; Garcia-Vallejo, J.; Houben, D.; Unger, W.W.J. Outer membrane vesicles engineered to express membrane-bound antigen program dendritic cells for cross-presentation to CD8+ T cells. Acta Biomater. 2019, 91, 248–257. [Google Scholar] [CrossRef] [PubMed]
- Song, B.; Song, J.; Zhang, S.; Anderson, M.A.; Ao, Y.; Yang, C.-Y.; Deming, T.J.; Sofroniew, M.V. Sustained local delivery of bioactive nerve growth factor in the central nervous system via tunable diblock copolypeptide hydrogel depots. Biomaterials 2012, 33, 9105–9116. [Google Scholar] [CrossRef]
- Zhang, S.; Anderson, M.A.; Ao, Y.; Khakh, B.S.; Fan, J.; Deming, T.J.; Sofroniew, M.V. Tunable diblock copolypeptide hydrogel depots for local delivery of hydrophobic molecules in healthy and injured central nervous system. Biomaterials 2014, 35, 1989–2000. [Google Scholar] [CrossRef] [Green Version]
- Schulz, E.; Goes, A.; Garcia, R.; Panter, F.; Koch, M.; Müller, R.; Fuhrmann, K.; Fuhrmann, G. Biocompatible bacteria-derived vesicles show inherent antimicrobial activity. J. Control. Release 2018, 290, 46–55. [Google Scholar] [CrossRef]
- Liu, Y.; Xie, J.; Zhao, X.; Zhang, Y.; Zhong, Z.; Deng, C. A polymeric IDO inhibitor based on poly(ethylene glycol)-b-poly(l-tyrosine-co-1-methyl-d-tryptophan) enables facile trident cancer immunotherapy. Biomater. Sci. 2022, 10, 5731–5743. [Google Scholar] [CrossRef]
- Mohammadi, M.; Taghavi, S.; Abnous, K.; Taghdisi, S.M.; Ramezani, M.; Alibolandi, M. Hybrid vesicular drug delivery systems for cancer therapeutics. Adv. Funct. Mater. 2018, 28, 1802136. [Google Scholar] [CrossRef]
- Song, W.; Zhang, Y.; Yu, D.-G.; Tran, C.H.; Wang, M.; Varyambath, A.; Kim, J.; Kim, I. Efficient synthesis of folate-conjugated hollow polymeric capsules for accurate drug delivery to cancer cells. Biomacromolecules 2021, 22, 732–742. [Google Scholar] [CrossRef]
- Cui, Y.; Deng, R.; Li, X.; Wang, X.; Jia, Q.; Bertrand, E.; Meguellati, K.; Yang, Y.-W. Temperature-sensitive polypeptide brushes-coated mesoporous silica nanoparticles for dual-responsive drug release. Chin. Chem. Lett. 2019, 30, 2291–2294. [Google Scholar] [CrossRef]
- Lin, B.; Wu, J.; Wang, Y.; Sun, S.; Yuan, Y.; Tao, X.; Lv, R. Peptide functionalized upconversion/NIR II luminescent nanoparticles for targeted imaging and therapy of oral squamous cell carcinoma. Biomater. Sci. 2021, 9, 1000–1007. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Li, C.; Ruan, Z.; Xu, P.; Yang, X.; Yuan, P.; Wang, Q.; Yan, L. Polypeptide-conjugated second near-infrared organic fluorophore for image-guided photothermal therapy. ACS Nano 2019, 13, 3691–3702. [Google Scholar] [CrossRef] [PubMed]
- Duhamel, J.; Kanagalingam, S.; O’Brie, T.J.; Ingratta, M.W. Side-chain dynamics of an α-helical polypeptide monitored by fluorescence. J. Am. Chem. Soc. 2003, 125, 12810–12822. [Google Scholar] [CrossRef]
- Tan, K.; Li, R.; Huang, X.; Liu, Q. Outer membrane vesicles: Current status and future direction of these novel vaccine adjuvants. Front. Microbiol. 2018, 9, 783. [Google Scholar] [CrossRef] [Green Version]
- Bielig, H.; Rompikuntal, P.; Dongre, M.; Zurek, B.; Lindmark, B.; Ramstedt, M.; Wai, S.; Kufer, T. NOD-like receptor activation by outer membrane vesicles from Vibrio cholerae non-O1 non-O139 strains is modulated by the quorum-sensing regulator HapR. Infect. Immun. 2011, 79, 1418–1427. [Google Scholar] [CrossRef] [Green Version]
- Johnson, R.P.; Jeong, Y.I.; John, J.V.; Chung, C.-W.; Kang, D.H.; Selvaraj, M.; Suh, H.; Kim, I. Dual stimuli-responsive poly(N-isopropylacrylamide)-b-poly(l-histidine) chimeric materials for the controlled delivery of doxorubicin into liver carcinoma. Biomacromolecules 2013, 14, 1434–1443. [Google Scholar] [CrossRef]
- Liu, J.; Huang, Y.; Kumar, A.; Tan, A.; Jin, S.; Mozhi, A.; Liang, X.-J. pH-Sensitive nano-systems for drug delivery in cancer therapy. Biotechnol. Adv. 2014, 32, 693–710. [Google Scholar] [CrossRef] [PubMed]
- Bazban-Shotorbani, S.; Hasani-Sadrabadi, M.M.; Karkhaneh, A.; Serpooshan, V.; Jacob, K.I.; Moshaverinia, A.; Mahmoudi, M. Revisiting structure-property relationship of pH-responsive polymers for drug delivery applications. J. Control. Release 2017, 253, 46–63. [Google Scholar] [CrossRef]
- He, T.; Li, D.; Yang, Y.; Ding, J.; Jin, F.; Zhuang, X.; Chen, X. Mesomeric configuration makes polyleucine micelle an optimal nanocarrier. Biomater. Sci. 2016, 4, 814–818. [Google Scholar] [CrossRef]
- Zhang, Y.; Uthaman, S.; Song, W.; Eom, K.H.; Jeon, S.H.; Huh, K.M.; Babu, A.; Park, I.-K.; Kim, I. Multistimuli-responsive polymeric vesicles for accelerated drug release in chemo-photothermal therapy. ACS Biomater. Sci. Eng. 2020, 6, 5012–5023. [Google Scholar] [CrossRef]
- Allan, N.D.; Beveridge, T.J. Gentamicin delivery to burkholderia cepacia group IIIa strains via membrane vesicles from pseudomonas aeruginosa PAO1. Antimicrob. Agents Chemother. 2003, 47, 2962–2965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shirbin, S.J.; Ladewig, K.; Fu, Q.; Klimak, M.; Zhang, X.; Duan, W.; Qiao, G.G. Cisplatin-induced formation of biocompatible and biodegradable polypeptide-based vesicles for targeted anticancer drug delivery. Biomacromolecules 2015, 16, 2463–2474. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Gupta, M.K.; Bang, J.B.; Bae, H.; Sung, H.J. Current progress in reactive oxygen species (ROS)-responsive materials for biomedical applications. Adv. Healthc. Mater. 2013, 2, 908–915. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, R.P.; Jeong, Y.-I.; Choi, E.; Chung, C.-W.; Kang, D.H.; Oh, S.-O.; Suh, H.; Kim, I. Biocompatible poly(2-hydroxyethyl methacrylate)-b-poly(L-histidine) hybrid materials for pH-sensitive intracellular anticancer drug delivery. Adv. Funct. Mater. 2012, 22, 1058–1068. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Classification | PVs | OMVs | Reference |
|---|---|---|---|
| Origin | Synthetic polymers | Gram-negative bacteria | [13,17] |
| Production | Solution self-assembly | Biogenesis | [15,21] |
| Size | Several nanometers~micrometers | 20~300 nm | [14,27] |
| Biomedical applications | Antibacterial | Bacterial vaccines | [14,28] |
| Mineralization | Adjuvants | ||
| Drug delivery | Cancer immunotherapy | ||
| Imaging and sensing | Drug delivery | ||
| Therapeutic | Anti-bacteria adhesion |
| DDS | PVs | References | OMVs | References |
|---|---|---|---|---|
| Drug loading | Hydrophobic interaction | [73,74] | Electroporation | [55] |
| Electrostatic interactions | [75] | Passive diffusion | [76] | |
| Metal coordination | [77] | Biogenesis | [78,79] | |
| Drug releasing | Change peptide assembly behavior | [80,81] | Degradation of OMVs | [55] |
| Change drug–polypeptide interaction | [82] | Coating on polymer | [83] | |
| Bioengineering | [84] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Y.; Lu, Y.; Xu, Y.; Zhou, Z.; Li, Y.; Ling, W.; Song, W. Bio-Inspired Drug Delivery Systems: From Synthetic Polypeptide Vesicles to Outer Membrane Vesicles. Pharmaceutics 2023, 15, 368. https://doi.org/10.3390/pharmaceutics15020368
Zhang Y, Lu Y, Xu Y, Zhou Z, Li Y, Ling W, Song W. Bio-Inspired Drug Delivery Systems: From Synthetic Polypeptide Vesicles to Outer Membrane Vesicles. Pharmaceutics. 2023; 15(2):368. https://doi.org/10.3390/pharmaceutics15020368
Chicago/Turabian StyleZhang, Yu, Yiming Lu, Yixin Xu, Zunkang Zhou, Yichong Li, Wei Ling, and Wenliang Song. 2023. "Bio-Inspired Drug Delivery Systems: From Synthetic Polypeptide Vesicles to Outer Membrane Vesicles" Pharmaceutics 15, no. 2: 368. https://doi.org/10.3390/pharmaceutics15020368