
Extracellular Vesicles for Dental Pulp and Periodontal Regeneration

Abstract
:1. Introduction
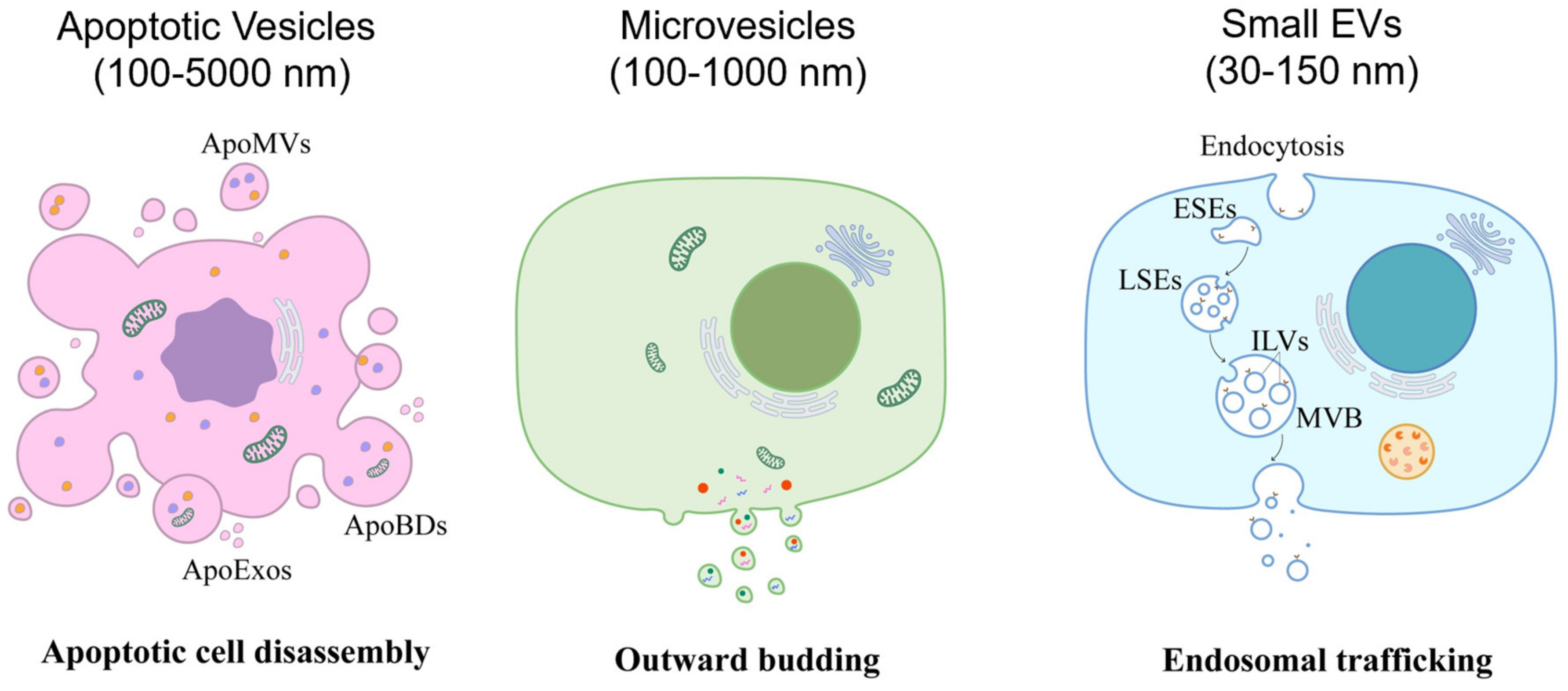
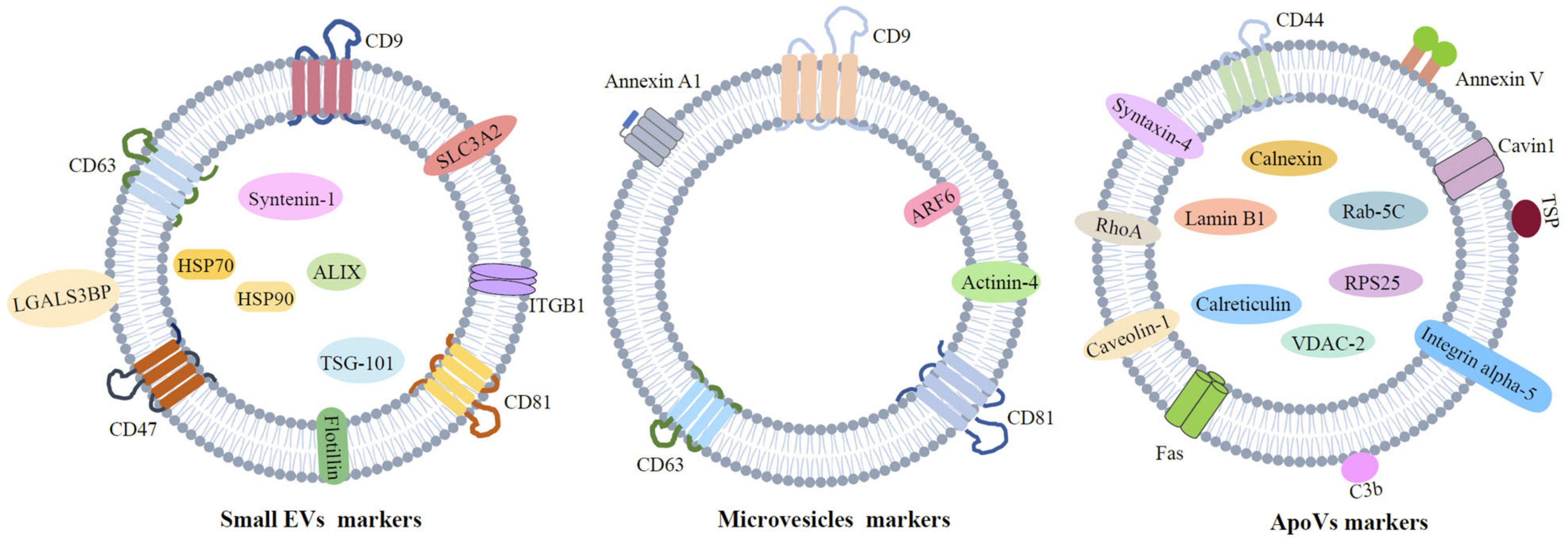
2. Extracellular Vesicles
2.1. Apoptotic Vesicles
2.2. Small EVs
2.3. Microvesicles
3. Mechanisms of EVs in Tissue Regeneration
4. EVs in Pulp Regeneration
4.1. Pulp Vascularization
4.2. Nerve Regeneration and Neural Repair
4.3. Dentin-Pulp Complex Regeneration
4.4. Immunomodulatory Properties in Pulp Regeneration

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Origin of EVs | Types of EVs | Study Models | Key Functions/Potential Molecular Mechanism | References |
|---|---|---|---|---|
| Dental pulp stem cells (DPSCs) | sEVs | In vitro and in vivo (teeth root slices implanted subcutaneously into nude mice dorsum) | Induced lineage-specific differentiation of stem cells/Triggered the P38 MAPK pathway | Huang et al., 2016 [102] |
| Dental pulp cells | sEVs | In vitro | Promoted angiogenesis/Enhanced tubular morphogenesis via p38 MAPK signaling inhibition | Xian et al., 2018 [89] |
| DPSCs | sEVs | In vitro and in vivo (dental pulp capping in SD rat teeth) | Enhanced odontogenesis by switching macrophages toward pro-healing M2 cells/Promoted odontogenesis in DPSCs through BMP2 pathway activation | Zheng et al., 2020 [109] |
| Hertwig’s epithelial root sheath (HERS) cells | sEVs-like vesicles | In vitro and in vivo (transplantation in renal capsule of rat; teeth root slices transplanted subcutaneously into nude mice dorsum) | Triggered regeneration of dental pulp-dentin-like tissue comprised of hard (reparative dentin-like tissue) and soft (blood vessels and neurons) tissue/Endocytosis of sEVs triggered the activation of P38 MAPK pathway | Zhang et al., 2020 [105] |
| DPSCs | EVs | In vitro | Promoted angiogenesis in an injectable hydrogel in vitro | Zhang et al., 2020 [111] |
| Stem cells from apical papilla (SCAPs) | sEVs | In vitro and in vivo (teeth root fragments implanted subcutaneously into nude mice dorsum) | Endocytosed by bone marrow-derived mesenchymal stem cells (BMMSCs) and significantly improved their specific dentinogenesis | Zhuang et al., 2020 [104] |
| Dental pulp cells | sEVs | In vitro | Induced the recruitment and proliferation of human mesenchymal stem cells | Ivica et al., 2020 [112] |
| Stem cells from human exfoliated deciduous teeth (SHED) | sEVs | In vitro and in vivo (teeth root fragments implanted subcutaneously into mice dorsum) | Shuttled miR-26a to promote angiogenesis via TGF- β/SMAD2/3 signaling | Wu et al., 2021 [90] |
| DPSCs | sEVs | In vitro | Lipopolysaccharide (LPS)-stimulated sEVs displayed a better ability on regulating SCs migration and odontogenic differentiation than normal sEVs | Li et al., 2021 [113] |
| DPSCs | sEVs | In vitro | LPS stimulated sEVs (LPS-sEVs) showed better proangiogenic potential of HUVECs compared with control sEVs/The expression of miR-146a-5p, miR-92b-5p, miR-218-5p, miR-23b-5p, miR-2110, miR-27a-5p, and miR-200b-3p was increased in the LPS-sEVs/The expression of miR-223-3p, miR-1246, and miR-494-3p was decreased in the LPS-sEVs | Huang et al., 2021 [114] |
| Dental pulp cells | sEVs | In vitro and in vivo (dental pulp capping in minipig teeth) | sEVs-treated dentin matrix promoted the formation of continuous reparative dentin | Wen et al., 2021 [115] |
| Dental pulp tissue and cells | sEVs | In vitro and in vivo (swine teeth implanted subcutaneously into mice dorsum) | Recruited SCAPs to regenerate connective tissue similar to natural dental pulp | Chen et al., 2022 [96] |
| Platelet | sEVs | In vitro | 5% thrombin-activated platelet-derived sEVs had a high potential to induce dental pulp regeneration | Bagio et al., 2022 [116] |
| Dental pulp cells | EVs | In vitro and in vivo (subcutaneous transplantation in nude mice) | Regulated cellular NFIC level in SCAPs to promote the proliferation, migration of SCAPs, and dentinogenesis | Yang et al., 2022 [117] |
| SHED | ApoVs | In vitro and in vivo (teeth fragments implanted subcutaneously into nude mice dorsum; orthotopic model of beagle dog) | Recruited endogenous endothelial cells (ECs) and facilitated the formation of dental-pulp-like tissue rich in blood vessels/ApoVs-carried mitochondrial Tu translation elongation factor modulated the angiogenic activation of ECs through the transcription factor EB-autophagy pathway | Li et al., 2022 [92] |
5. EVs in Periodontal Regeneration
6. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Doyle, L.M.; Wang, M.Z. Overview of Extracellular Vesicles, Their Origin, Composition, Purpose, and Methods for Exosome Isolation and Analysis. Cells 2019, 8, 727. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yap, T.; Koo, K.; Cheng, L.; Vella, L.J.; Hill, A.F.; Reynolds, E.; Nastri, A.; Cirillo, N.; Seers, C.; McCullough, M. Predicting the Presence of Oral Squamous Cell Carcinoma Using Commonly Dysregulated Microrna in Oral Swirls. Cancer Prev. Res. 2018, 11, 491–502. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wolf, P. The Nature and Significance of Platelet Products in Human Plasma. Br. J. Haematol. 1967, 13, 269–288. [Google Scholar] [CrossRef] [PubMed]
- Théry, C.; Witwer, K.W.; Aikawa, E.; Alcaraz, M.J.; Anderson, J.D.; Andriantsitohaina, R.; Antoniou, A.; Arab, T.; Archer, F.; Atkin-Smith, G.K.; et al. Minimal Information for Studies of Extracellular Vesicles 2018 (Misev2018): A Position Statement of the International Society for Extracellular Vesicles and Update of the Misev2014 Guidelines. J. Extracell. Vesicles 2018, 7, 1535750. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Niel, G.; D’Angelo, G.; Raposo, G. Shedding Light on the Cell Biology of Extracellular Vesicles. Nat. Rev. Mol. Cell Biol. 2018, 19, 213–228. [Google Scholar] [CrossRef] [PubMed]
- Cheng, L.; Hill, A.F. Therapeutically Harnessing Extracellular Vesicles. Nat. Rev. Drug Discov. 2022, 21, 379–399. [Google Scholar] [CrossRef]
- Kalluri, R.; LeBleu, V.S. The Biology, Function, and Biomedical Applications of Exosomes. Science 2020, 367, eaau6977. [Google Scholar] [CrossRef]
- Itoh, Y.; Sasaki, J.I.; Hashimoto, M.; Katata, C.; Hayashi, M.; Imazato, S. Pulp Regeneration by 3-Dimensional Dental Pulp Stem Cell Constructs. J. Dent. Res. 2018, 97, 1137–1143. [Google Scholar] [CrossRef]
- Ricucci, D.; Siqueira, J.F.; Abdelsayed, R.A.; Lio, S.G.; Rôças, I.N. Bacterial Invasion of Pulp Blood Vessels in Teeth with Symptomatic Irreversible Pulpitis. J. Endod. 2021, 47, 1854–1864. [Google Scholar] [CrossRef]
- He, L.; Kim, S.G.; Gong, Q.; Zhong, J.; Wang, S.; Zhou, X.; Ye, L.; Ling, J.; Mao, J.J. Regenerative Endodontics for Adult Patients. J. Endod. 2017, 43, S57–S64. [Google Scholar] [CrossRef]
- Xuan, K.; Li, B.; Guo, H.; Sun, W.; Kou, X.; He, X.; Zhang, Y.; Sun, J.; Liu, A.; Liao, L.; et al. Deciduous Autologous Tooth Stem Cells Regenerate Dental Pulp after Implantation into Injured Teeth. Sci. Transl. Med. 2018, 10, eaaf3227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Matsuzaka, Y.; Yashiro, R. Therapeutic Strategy of Mesenchymal-Stem-Cell-Derived Extracellular Vesicles as Regenerative Medicine. Int. J. Mol. Sci. 2022, 23, 6480. [Google Scholar] [CrossRef] [PubMed]
- Hajishengallis, G. Periodontitis: From Microbial Immune Subversion to Systemic Inflammation. Nat. Rev. Immunol. 2015, 15, 30–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kinane, D.F.; Stathopoulou, P.G.; Papapanou, P.N. Periodontal Diseases. Nat. Rev. Dis. Prim. 2017, 3, 17038. [Google Scholar] [CrossRef] [PubMed]
- Nuñez, J.; Vignoletti, F.; Caffesse, R.G.; Sanz, M. Cellular Therapy in Periodontal Regeneration. Periodontol 2000 2019, 79, 107–116. [Google Scholar] [CrossRef]
- Li, Y.; Duan, X.; Chen, Y.; Liu, B.; Chen, G. Dental Stem Cell-Derived Extracellular Vesicles as Promising Therapeutic Agents in the Treatment of Diseases. Int. J. Oral Sci. 2022, 14, 2. [Google Scholar] [CrossRef]
- He, F.; Li, L.; Fan, R.; Wang, X.; Chen, X.; Xu, Y. Extracellular Vesicles: An Emerging Regenerative Treatment for Oral Disease. Front. Cell Dev. Biol. 2021, 9, 669011. [Google Scholar] [CrossRef]
- Lynch, C.; Panagopoulou, M.; Gregory, C.D. Extracellular Vesicles Arising from Apoptotic Cells in Tumors: Roles in Cancer Pathogenesis and Potential Clinical Applications. Front. Immunol. 2017, 8, 1174. [Google Scholar] [CrossRef] [Green Version]
- Jiang, L.; Paone, S.; Caruso, S.; Atkin-Smith, G.K.; Phan, T.K.; Hulett, M.D.; Poon, I.K.H. Determining the Contents and Cell Origins of Apoptotic Bodies by Flow Cytometry. Sci. Rep. 2017, 7, 14444. [Google Scholar] [CrossRef] [Green Version]
- Jeppesen, D.K.; Fenix, A.M.; Franklin, J.L.; Higginbotham, J.N.; Zhang, Q.; Zimmerman, L.J.; Liebler, D.C.; Ping, J.; Liu, Q.; Evans, R.; et al. Reassessment of Exosome Composition. Cell 2019, 177, 428–445.e18. [Google Scholar] [CrossRef]
- Zhang, X.; Tang, J.; Kou, X.; Huang, W.; Zhu, Y.; Jiang, Y.; Yang, K.; Li, C.; Hao, M.; Qu, Y.; et al. Proteomic Analysis of Msc-Derived Apoptotic Vesicles Identifies Fas Inheritance to Ameliorate Haemophilia a Via Activating Platelet Functions. J. Extracell. Vesicles 2022, 11, e12240. [Google Scholar] [CrossRef]
- Poon, W.; Zhang, Y.-N.; Ouyang, B.; Kingston, B.R.; Wu, J.L.Y.; Wilhelm, S.; Chan, W.C.W. Elimination Pathways of Nanoparticles. ACS Nano 2019, 13, 5785–5798. [Google Scholar] [CrossRef] [PubMed]
- Kerr, J.F.R.; Wyllie, A.H.; Currie, A.R. Apoptosis: A Basic Biological Phenomenon with Wide-Ranging Implications in Tissue Kinetics. Br. J. Cancer 1972, 26, 239–257. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Battistelli, M.; Falcieri, E. Apoptotic Bodies: Particular Extracellular Vesicles Involved in Intercellular Communication. Biology 2020, 9, 21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.; Liu, Y.; Liu, X.; Du, J.; Bhawal, U.K.; Xu, J.; Guo, L.; Liu, Y. Advances in the Therapeutic Effects of Apoptotic Bodies on Systemic Diseases. Int. J. Mol. Sci. 2022, 23, 8202. [Google Scholar] [CrossRef] [PubMed]
- Schiller, M.; Parcina, M.; Heyder, P.; Foermer, S.; Ostrop, J.; Leo, A.; Heeg, K.; Herrmann, M.; Lorenz, H.-M.; Bekeredjian-Ding, I. Induction of Type I Ifn Is a Physiological Immune Reaction to Apoptotic Cell-Derived Membrane Microparticles. J. Immunol. 2012, 189, 1747–1756. [Google Scholar] [CrossRef] [Green Version]
- Lötvall, J.; Hill, A.F.; Hochberg, F.; Buzás, E.I.; Di Vizio, D.; Gardiner, C.; Gho, Y.S.; Kurochkin, I.V.; Mathivanan, S.; Quesenberry, P.; et al. Minimal Experimental Requirements for Definition of Extracellular Vesicles and Their Functions: A Position Statement from the International Society for Extracellular Vesicles. J. Extracell. Vesicles 2014, 3, 26913. [Google Scholar] [CrossRef]
- Dieudé, M.; Bell, C.; Turgeon, J.; Beillevaire, D.; Pomerleau, L.; Yang, B.; Hamelin, K.; Qi, S.; Pallet, N.; Béland, C.; et al. The 20s Proteasome Core, Active within Apoptotic Exosome-Like Vesicles, Induces Autoantibody Production and Accelerates Rejection. Sci. Transl. Med. 2015, 7, 318ra200. [Google Scholar] [CrossRef] [Green Version]
- Park, S.J.; Kim, J.M.; Kim, J.; Hur, J.; Park, S.; Kim, K.; Shin, H.-J.; Chwae, Y.-J. Molecular Mechanisms of Biogenesis of Apoptotic Exosome-Like Vesicles and Their Roles as Damage-Associated Molecular Patterns. Proc. Natl. Acad. Sci. USA 2018, 115, E11721–E11730. [Google Scholar] [CrossRef] [Green Version]
- Kakarla, R.; Hur, J.; Kim, Y.J.; Kim, J.; Chwae, Y.-J. Apoptotic Cell-Derived Exosomes: Messages from Dying Cells. Exp. Mol. Med. 2020, 52, 1–6. [Google Scholar] [CrossRef]
- Atkin-Smith, G.K.; Tixeira, R.; Paone, S.; Mathivanan, S.; Collins, C.; Liem, M.; Goodall, K.; Ravichandran, K.; Hulett, M.; Poon, I.K. A Novel Mechanism of Generating Extracellular Vesicles During Apoptosis Via a Beads-on-a-String Membrane Structure. Nat. Commun. 2015, 6, 7439. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Chen, X.; Gueydan, C.; Han, J. Plasma Membrane Changes During Programmed Cell Deaths. Cell Res. 2018, 28, 9–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, X.; Lai, Y.; Hua, Z.-C. Apoptosis and Apoptotic Body: Disease Message and Therapeutic Target Potentials. Biosci. Rep. 2019, 39, BSR20180992. [Google Scholar] [CrossRef] [Green Version]
- Caruso, S.; Poon, I.K.H. Apoptotic Cell-Derived Extracellular Vesicles: More Than Just Debris. Front. Immunol. 2018, 9, 1486. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Chen, C.; Liu, D.; Huang, Z.; Li, J.; Liu, H.; Kwok, R.T.K.; Tang, B.; Sui, B.; Zhang, X.; et al. Apoptotic Extracellular Vesicles Are Metabolized Regulators Nurturing the Skin and Hair. Bioact. Mater. 2023, 19, 626–641. [Google Scholar] [CrossRef] [PubMed]
- Kugeratski, F.G.; Hodge, K.; Lilla, S.; McAndrews, K.M.; Zhou, X.; Hwang, R.F.; Zanivan, S.; Kalluri, R. Quantitative Proteomics Identifies the Core Proteome of Exosomes with Syntenin-1 as the Highest Abundant Protein and a Putative Universal Biomarker. Nat. Cell Biol. 2021, 23, 631–641. [Google Scholar] [CrossRef] [PubMed]
- Subiros-Funosas, R.; Mendive-Tapia, L.; Sot, J.; Pound, J.D.; Barth, N.; Varela, Y.; Goñi, F.M.; Paterson, M.; Gregory, C.D.; Albericio, F.; et al. A Trp-Bodipy Cyclic Peptide for Fluorescence Labelling of Apoptotic Bodies. Chem. Commun. 2017, 53, 945–948. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Zhang, Z.; Xia, W.; Cai, J.; Li, Y.; Wu, S. Extracellular Vesicles in Urologic Malignancies-Implementations for Future Cancer Care. Cell Prolif. 2019, 52, e12659. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Liao, L.; Tian, W. Extracellular Vesicles Derived from Apoptotic Cells: An Essential Link between Death and Regeneration. Front. Cell Dev. Biol. 2020, 8, 573511. [Google Scholar] [CrossRef]
- Zheng, C.; Sui, B.; Zhang, X.; Hu, J.; Chen, J.; Liu, J.; Wu, D.; Ye, Q.; Xiang, L.; Qiu, X.; et al. Apoptotic Vesicles Restore Liver Macrophage Homeostasis to Counteract Type 2 Diabetes. J. Extracell. Vesicles 2021, 10, e12109. [Google Scholar] [CrossRef]
- Wang, R.; Hao, M.; Kou, X.; Sui, B.; Sanmillan, M.L.; Zhang, X.; Liu, D.; Tian, J.; Yu, W.; Chen, C.; et al. Apoptotic Vesicles Ameliorate Lupus and Arthritis Via Phosphatidylserine-Mediated Modulation of T Cell Receptor Signaling. Bioact. Mater. 2022, in press. [Google Scholar] [CrossRef]
- Crescitelli, R.; Lässer, C.; Szabó, T.G.; Kittel, A.; Eldh, M.; Dianzani, I.; Buzás, E.I.; Lötvall, J. Distinct Rna Profiles in Subpopulations of Extracellular Vesicles: Apoptotic Bodies, Microvesicles and Exosomes. J. Extracell. Vesicles 2013, 2, 20677. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poon, I.K.H.; Parkes, M.A.F.; Jiang, L.; Atkin-Smith, G.K.; Tixeira, R.; Gregory, C.D.; Ozkocak, D.C.; Rutter, S.F.; Caruso, S.; Santavanond, J.P.; et al. Moving Beyond Size and Phosphatidylserine Exposure: Evidence for a Diversity of Apoptotic Cell-Derived Extracellular Vesicles in Vitro. J. Extracell. Vesicles 2019, 8, 1608786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, D.; Kou, X.; Chen, C.; Liu, S.; Liu, Y.; Yu, W.; Yu, T.; Yang, R.; Wang, R.; Zhou, Y.; et al. Circulating Apoptotic Bodies Maintain Mesenchymal Stem Cell Homeostasis and Ameliorate Osteopenia Via Transferring Multiple Cellular Factors. Cell Res. 2018, 28, 918–933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pavlyukov, M.S.; Yu, H.; Bastola, S.; Minata, M.; Shender, V.O.; Lee, Y.; Zhang, S.; Wang, J.; Komarova, S.; Wang, J.; et al. Apoptotic Cell-Derived Extracellular Vesicles Promote Malignancy of Glioblastoma Via Intercellular Transfer of Splicing Factors. Cancer Cell 2018, 34, 119–135.e10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Halicka, H.; Bedner, E.; Darzynkiewicz, Z. Segregation of Rna and Separate Packaging of DNA and Rna in Apoptotic Bodies During Apoptosis. Exp. Cell Res. 2000, 260, 248–256. [Google Scholar] [CrossRef]
- Choi, D.; Go, G.; Kim, D.; Lee, J.; Park, S.; Di Vizio, D.; Gho, Y.S. Quantitative Proteomic Analysis of Trypsin-Treated Extracellular Vesicles to Identify the Real-Vesicular Proteins. J. Extracell. Vesicles 2020, 9, 1757209. [Google Scholar] [CrossRef]
- Akbari, A.; Jabbari, N.; Sharifi, R.; Ahmadi, M.; Vahhabi, A.; Seyedzadeh, S.J.; Nawaz, M.; Szafert, S.; Mahmoodi, M.; Jabbari, E.; et al. Free and Hydrogel Encapsulated Exosome-Based Therapies in Regenerative Medicine. Life Sci. 2020, 249, 117447. [Google Scholar] [CrossRef]
- Zabeo, D.; Cvjetkovic, A.; Lässer, C.; Schorb, M.; Lötvall, J.; Höög, J.L. Exosomes Purified from a Single Cell Type Have Diverse Morphology. J. Extracell. Vesicles 2017, 6, 1329476. [Google Scholar] [CrossRef] [Green Version]
- Trams, E.G.; Lauter, C.J.; Salem, N., Jr.; Heine, U. Exfoliation of Membrane Ecto-Enzymes in the Form of Micro-Vesicles. Biochim. Et Biophys. Acta 1981, 645, 63–70. [Google Scholar] [CrossRef]
- Moeinzadeh, L.; Razeghian-Jahromi, I.; Zarei-Behjani, Z.; Bagheri, Z.; Razmkhah, M. Composition, Biogenesis, and Role of Exosomes in Tumor Development. Stem Cells Int. 2022, 2022, 8392509. [Google Scholar] [CrossRef] [PubMed]
- De Souza, W.; Barrias, E.S. Membrane-Bound Extracellular Vesicles Secreted by Parasitic Protozoa: Cellular Structures Involved in the Communication between Cells. Parasitol. Res. 2020, 119, 2005–2023. [Google Scholar] [CrossRef] [PubMed]
- Wollert, T.; Hurley, J.H. Molecular Mechanism of Multivesicular Body Biogenesis by Escrt Complexes. Nature 2010, 464, 864–869. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Verderio, C.; Gabrielli, M.; Giussani, P. Role of Sphingolipids in the Biogenesis and Biological Activity of Extracellular Vesicles. J. Lipid Res. 2018, 59, 1325–1340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baietti, M.F.; Zhang, Z.; Mortier, E.; Melchior, A.; DeGeest, G.; Geeraerts, A.; Ivarsson, Y.; Depoortere, F.; Coomans, C.; Vermeiren, E.; et al. Syndecan-Syntenin-Alix Regulates the Biogenesis of Exosomes. Nat. Cell Biol. 2012, 14, 677–685. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.; Zhang, Q.; Jiang, L. Current Knowledge on Exosome Biogenesis, Cargo-Sorting Mechanism and Therapeutic Implications. Membranes 2022, 12, 498. [Google Scholar] [CrossRef]
- Zhao, H.; Song, S.; Ma, J.; Yan, Z.; Xie, H.; Feng, Y.; Che, S. Cd47 as a Promising Therapeutic Target in Oncology. Front. Immunol. 2022, 13, 757480. [Google Scholar] [CrossRef]
- Sasaki, T.; Brakebusch, C.; Engel, J.; Timpl, R. Mac-2 Binding Protein Is a Cell-Adhesive Protein of the Extracellular Matrix Which Self-Assembles into Ring-Like Structures and Binds Beta1 Integrins, Collagens and Fibronectin. EMBO J. 1998, 17, 1606–1613. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Liu, Y.; Liu, H.; Tang, W.H. Exosomes: Biogenesis, Biologic Function and Clinical Potential. Cell Biosci. 2019, 9, 19. [Google Scholar] [CrossRef]
- Mittelbrunn, M.; Gutiérrez-Vázquez, C.; Villarroya-Beltri, C.; González, S.; Sánchez-Cabo, F.; González, M.A.; Bernad, A.; Sánchez-Madrid, F. Unidirectional Transfer of Microrna-Loaded Exosomes from T Cells to Antigen-Presenting Cells. Nat. Commun. 2011, 2, 282. [Google Scholar] [CrossRef]
- Huang, X.; Yuan, T.; Tschannen, M.; Sun, Z.; Jacob, H.; Du, M.; Liang, M.; Dittmar, R.L.; Liu, Y.; Liang, M.; et al. Characterization of Human Plasma-Derived Exosomal Rnas by Deep Sequencing. BMC Genom. 2013, 14, 319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Subra, C.; Grand, D.; Laulagnier, K.; Stella, A.; Lambeau, G.; Paillasse, M.; De Medina, P.; Monsarrat, B.; Perret, B.; Silvente-Poirot, S.; et al. Exosomes Account for Vesicle-Mediated Transcellular Transport of Activatable Phospholipases and Prostaglandins. J. Lipid Res. 2010, 51, 2105–2120. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hargett, L.A.; Bauer, N.N. On the Origin of Microparticles: From “Platelet Dust” to Mediators of Intercellular Communication. Pulm. Circ. 2013, 3, 329–340. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.; Wang, S.; Xia, H.; Tan, X.; Song, S.; Zhang, S.; Meng, D.; Chen, Q.; Jin, Y. The Potential Applications of Microparticles in the Diagnosis, Treatment, and Prognosis of Lung Cancer. J. Transl. Med. 2022, 20, 404. [Google Scholar] [CrossRef] [PubMed]
- Agrahari, V.; Agrahari, V.; Burnouf, P.-A.; Chew, C.H.; Burnouf, T. Extracellular Microvesicles as New Industrial Therapeutic Frontiers. Trends Biotechnol. 2019, 37, 707–729. [Google Scholar] [CrossRef] [PubMed]
- Ståhl, A.-L.; Johansson, K.; Mossberg, M.; Kahn, R.; Karpman, D. Exosomes and Microvesicles in Normal Physiology, Pathophysiology, and Renal Diseases. Pediatr. Nephrol. 2019, 34, 11–30. [Google Scholar] [CrossRef] [Green Version]
- Hu, Y.; Sun, Y.; Wan, C.; Dai, X.; Wu, S.; Lo, P.-C.; Huang, J.; Lovell, J.F.; Jin, H.; Yang, K. Microparticles: Biogenesis, Characteristics and Intervention Therapy for Cancers in Preclinical and Clinical Research. J. Nanobiotechnol. 2022, 20, 189. [Google Scholar] [CrossRef]
- Mathieu, M.; Névo, N.; Jouve, M.; Valenzuela, J.I.; Maurin, M.; Verweij, F.J.; Palmulli, R.; Lankar, D.; Dingli, F.; Loew, D.; et al. Specificities of Exosome Versus Small Ectosome Secretion Revealed by Live Intracellular Tracking of Cd63 and Cd9. Nat. Commun. 2021, 12, 4389. [Google Scholar] [CrossRef]
- Guan, F.; Xiang, X.; Xie, Y.; Li, H.; Zhang, W.; Shu, Y.; Wang, J.; Qin, W. Simultaneous Metabolomics and Proteomics Analysis of Plasma-Derived Extracellular Vesicles. Anal. Methods Adv. Methods Appl. 2021, 13, 1930–1938. [Google Scholar] [CrossRef]
- Chen, X.; Liang, H.; Zhang, J.; Zen, K.; Zhang, C.-Y. Secreted Micrornas: A New Form of Intercellular Communication. Trends Cell Biol. 2012, 22, 125–132. [Google Scholar] [CrossRef]
- Liang, H.; Yan, X.; Pan, Y.; Wang, Y.; Wang, N.; Li, L.; Liu, Y.; Chen, X.; Zhang, C.-Y.; Gu, H.; et al. Microrna-223 Delivered by Platelet-Derived Microvesicles Promotes Lung Cancer Cell Invasion Via Targeting Tumor Suppressor Epb41l3. Mol. Cancer 2015, 14, 58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Wu, X.; Chandra, S.; Lyon, C.; Ning, B.; Jiang, L.; Fan, J.; Hu, T.Y. Extracellular Vesicles: Emerging Tools as Therapeutic Agent Carriers. Acta Pharm. Sinica. B 2022, 12, 3822–3842. [Google Scholar] [CrossRef] [PubMed]
- Nagelkerke, A.; Ojansivu, M.; Van Der Koog, L.; Whittaker, T.E.; Cunnane, E.M.; Silva, A.M.; Dekker, N.; Stevens, M.M. Extracellular Vesicles for Tissue Repair and Regeneration: Evidence, Challenges and Opportunities. Adv. Drug Deliv. Rev. 2021, 175, 113775. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Lin, L.; Zou, R.; Wen, C.; Wang, Z.; Lin, F. Msc-Derived Exosomes Promote Proliferation and Inhibit Apoptosis of Chondrocytes Via Lncrna-Klf3-As1/Mir-206/Git1 Axis in Osteoarthritis. Cell Cycle 2018, 17, 2411–2422. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, L.; Liu, G.; Wu, X. Exosomes Derived from Umbilical Cord Mesenchymal Stem Cells in Mechanical Environment Show Improved Osteochondral Activity Via Upregulation of Lncrna H19. J. Orthop. Transl. 2021, 26, 111–120. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Liu, D.; Chen, C.; Hamamura, K.; Moshaverinia, A.; Yang, R.; Liu, Y.; Jin, Y.; Shi, S. MSC Transplantation Improves Osteopenia Via Epigenetic Regulation of Notch Signaling in Lupus. Cell Metab. 2015, 22, 606–618. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiu, G.; Zheng, G.; Ge, M.; Wang, J.; Huang, R.; Shu, Q.; Xu, J. Functional Proteins of Mesenchymal Stem Cell-Derived Extracellular Vesicles. Stem Cell Res. Ther. 2019, 10, 359. [Google Scholar] [CrossRef] [Green Version]
- Pan, Z.; Sun, W.; Chen, Y.; Tang, H.; Lin, W.; Chen, J.; Chen, C. Extracellular Vesicles in Tissue Engineering: Biology and Engineered Strategy. Adv. Healthc. Mater. 2022, 11, e2201384. [Google Scholar] [CrossRef]
- Zhang, Y.; Xie, Y.; Hao, Z.; Zhou, P.; Wang, P.; Fang, S.; Li, L.; Xu, S.; Xia, Y. Umbilical Mesenchymal Stem Cell-Derived Exosome-Encapsulated Hydrogels Accelerate Bone Repair by Enhancing Angiogenesis. ACS Appl. Mater. Interfaces 2021, 13, 18472–18487. [Google Scholar] [CrossRef]
- Sun, S.-J.; Wei, R.; Li, F.; Liao, S.-Y.; Tse, H.-F. Mesenchymal Stromal Cell-Derived Exosomes in Cardiac Regeneration and Repair. Stem Cell Rep. 2021, 16, 1662–1673. [Google Scholar] [CrossRef]
- Anderson, J.D.; Johansson, H.J.; Graham, C.S.; Vesterlund, M.; Pham, M.T.; Bramlett, C.S.; Montgomery, E.N.; Mellema, M.S.; Bardini, R.L.; Contreras, Z.; et al. Comprehensive Proteomic Analysis of Mesenchymal Stem Cell Exosomes Reveals Modulation of Angiogenesis Via Nuclear Factor-Kappab Signaling. Stem Cells 2016, 34, 601–613. [Google Scholar] [CrossRef] [Green Version]
- Zhang, K.; Liu, L.; Shi, K.; Zhang, K.; Zheng, C.; Jin, Y. Extracellular Vesicles for Immunomodulation in Tissue Regeneration. Tissue Eng. Part C Methods 2022, 28, 393–404. [Google Scholar] [CrossRef] [PubMed]
- Murray, P.J.; Allen, J.E.; Biswas, S.K.; Fisher, E.A.; Gilroy, D.W.; Goerdt, S.; Gordon, S.; Hamilton, J.A.; Ivashkiv, L.B.; Lawrence, T.; et al. Macrophage Activation and Polarization: Nomenclature and Experimental Guidelines. Immunity 2014, 41, 14–20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Huang, R.; Xu, Q.; Zheng, G.; Qiu, G.; Ge, M.; Shu, Q.; Xu, J. Mesenchymal Stem Cell-Derived Extracellular Vesicles Alleviate Acute Lung Injury Via Transfer of Mir-27a-3p. Crit. Care Med. 2020, 48, e599–e610. [Google Scholar] [CrossRef] [PubMed]
- Duan, S.; Wang, F.; Cao, J.; Wang, C. Exosomes Derived from Microrna-146a-5p-Enriched Bone Marrow Mesenchymal Stem Cells Alleviate Intracerebral Hemorrhage by Inhibiting Neuronal Apoptosis and Microglial M1 Polarization. Drug Des. Dev. Ther. 2020, 14, 3143–3158. [Google Scholar] [CrossRef] [PubMed]
- Kou, X.; Xu, X.; Chen, C.; Sanmillan, M.L.; Cai, T.; Zhou, Y.; Giraudo, C.; Le, A.; Shi, S. The Fas/Fap-1/Cav-1 Complex Regulates Il-1ra Secretion in Mesenchymal Stem Cells to Accelerate Wound Healing. Sci. Transl. Med. 2018, 10, eaai8524. [Google Scholar] [CrossRef] [Green Version]
- Rombouts, C.; Giraud, T.; Jeanneau, C.; About, I. Pulp Vascularization During Tooth Development, Regeneration, and Therapy. J. Dent. Res. 2017, 96, 137–144. [Google Scholar] [CrossRef]
- Yu, S.; Chen, H.; Gao, B. Potential Therapeutic Effects of Exosomes in Regenerative Endodontics. Arch. Oral Biol. 2020, 120, 104946. [Google Scholar] [CrossRef]
- Xian, X.; Gong, Q.; Li, C.; Guo, B.; Jiang, H. Exosomes with Highly Angiogenic Potential for Possible Use in Pulp Regeneration. J. Endod. 2018, 44, 751–758. [Google Scholar] [CrossRef]
- Wu, M.; Liu, X.; Li, Z.; Huang, X.; Guo, H.; Guo, X.; Yang, X.; Li, B.; Xuan, K.; Jin, Y. Shed Aggregate Exosomes Shuttled Mir-26a Promote Angiogenesis in Pulp Regeneration Via Tgf-Beta/Smad2/3 Signalling. Cell Prolif. 2021, 54, e13074. [Google Scholar] [CrossRef]
- György, B.; Szabó, T.G.; Pásztói, M.; Pál, Z.; Misják, P.; Aradi, B.; László, V.; Pállinger, É.; Pap, E.; Kittel, Á.; et al. Membrane Vesicles, Current State-of-the-Art: Emerging Role of Extracellular Vesicles. Cell. Mol. Life Sci. 2011, 68, 2667–2688. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Z.; Wu, M.; Liu, S.; Liu, X.; Huan, Y.; Ye, Q.; Yang, X.; Guo, H.; Liu, A.; Huang, X.; et al. Apoptotic Vesicles Activate Autophagy in Recipient Cells to Induce Angiogenesis and Dental Pulp Regeneration. Mol. Ther. 2022, 30, 3193–3208. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.; Li, X.; Yin, Y.; He, X.-T.; An, Y.; Tian, B.-M.; Hong, Y.-L.; Wu, L.-A.; Chen, F.-M. The Proangiogenic Effects of Extracellular Vesicles Secreted by Dental Pulp Stem Cells Derived from Periodontally Compromised Teeth. Stem Cell Res. Ther. 2020, 11, 110. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Shuai, Y.; Zhou, F.; Yin, J.; Hu, J.; Guo, S.; Wang, Y.; Liu, W. Pdlscs Regulate Angiogenesis of Periodontal Ligaments Via Vegf Transferred by Exosomes in Periodontitis. Int. J. Med. Sci. 2020, 17, 558–567. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.-J.; Xie, J.; Lin, X.; Ou, M.-H.; Zhou, J.; Wei, X.-L. The Role of Small Extracellular Vesicles Derived from Lipopolysaccharide-Preconditioned Human Dental Pulp Stem Cells in Dental Pulp Regeneration. J. Endod. 2021, 47, 961–969. [Google Scholar] [CrossRef]
- Chen, Y.; Ma, Y.; Yang, X.; Chen, J.; Yang, B.; Tian, W. The Application of Pulp Tissue Derived-Exosomes in Pulp Regeneration: A Novel Cell-Homing Approach. Int. J. Nanomed. 2022, 17, 465–476. [Google Scholar] [CrossRef] [PubMed]
- Sui, B.; Chen, C.; Kou, X.; Li, B.; Xuan, K.; Shi, S.; Jin, Y. Pulp Stem Cell-Mediated Functional Pulp Regeneration. J. Dent. Res. 2019, 98, 27–35. [Google Scholar] [CrossRef]
- Wang, D.; Lyu, Y.; Yang, Y.; Zhang, S.; Chen, G.; Pan, J.; Tian, W. Schwann Cell-Derived Evs Facilitate Dental Pulp Regeneration through Endogenous Stem Cell Recruitment Via Sdf-1/Cxcr4 Axis. Acta Biomater. 2022, 140, 610–624. [Google Scholar] [CrossRef]
- Couve, E.; Schmachtenberg, O. Schwann Cell Responses and Plasticity in Different Dental Pulp Scenarios. Front. Cell. Neurosci. 2018, 12, 299. [Google Scholar] [CrossRef]
- Xie, Z.; Shen, Z.; Zhan, P.; Yang, J.; Huang, Q.; Huang, S.; Chen, L.; Lin, Z. Functional Dental Pulp Regeneration: Basic Research and Clinical Translation. Int. J. Mol. Sci. 2021, 22, 8991. [Google Scholar] [CrossRef]
- Ivica, A.; Zehnder, M.; Weber, F.E. Therapeutic Potential of Mesenchymal Stem Cell-Derived Extracellular Vesicles in Regenerative Endodontics. Eur. Cells Mater. 2021, 41, 233–244. [Google Scholar] [CrossRef] [PubMed]
- Huang, C.-C.; Narayanan, R.; Alapati, S.; Ravindran, S. Exosomes as Biomimetic Tools for Stem Cell Differentiation: Applications in Dental Pulp Tissue Regeneration. Biomaterials 2016, 111, 103–115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, X.; Zhong, Y.; Kong, Y.; Chen, Y.; Feng, J.; Zheng, J. Lineage-Specific Exosomes Promote the Odontogenic Differentiation of Human Dental Pulp Stem Cells (Dpscs) through Tgfβ1/Smads Signaling Pathway Via Transfer of Micrornas. Stem Cell Res. Ther. 2019, 10, 170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuang, X.; Ji, L.; Jiang, H.; Liu, Y.; Liu, X.; Bi, J.; Zhao, W.; Ding, Z.; Chen, X. Exosomes Derived from Stem Cells from the Apical Papilla Promote Dentine-Pulp Complex Regeneration by Inducing Specific Dentinogenesis. Stem Cells Int. 2020, 2020, 5816723. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Yang, Y.; Jia, S.; Chen, H.; Duan, Y.; Li, X.; Wang, S.; Wang, T.; Lyu, Y.; Chen, G.; et al. Exosome-Like Vesicles Derived from Hertwig’s Epithelial Root Sheath Cells Promote the Regeneration of Dentin-Pulp Tissue. Theranostics 2020, 10, 5914–5931. [Google Scholar] [CrossRef]
- Kou, M.; Huang, L.; Yang, J.; Chiang, Z.; Chen, S.; Liu, J.; Guo, L.; Zhang, X.; Zhou, X.; Xu, X.; et al. Mesenchymal Stem Cell-Derived Extracellular Vesicles for Immunomodulation and Regeneration: A Next Generation Therapeutic Tool? Cell Death Dis. 2022, 13, 580. [Google Scholar] [CrossRef]
- Sun, J.; Wang, Z.; Liu, P.; Hu, Y.; Li, T.; Yang, J.; Gao, P.; Xu, Q. Exosomes Derived from Human Gingival Mesenchymal Stem Cells Attenuate the Inflammatory Response in Periodontal Ligament Stem Cells. Front. Chem. 2022, 10, 863364. [Google Scholar] [CrossRef]
- Park, H.C.; Quan, H.; Zhu, T.; Kim, Y.; Kim, B.; Yang, H.-C. The Effects of M1 and M2 Macrophages on Odontogenic Differentiation of Human Dental Pulp Cells. J. Endod. 2017, 43, 596–601. [Google Scholar] [CrossRef]
- Zheng, J.; Kong, Y.; Hu, X.; Li, Z.; Li, Y.; Zhong, Y.; Wei, X.; Ling, J. Microrna-Enriched Small Extracellular Vesicles Possess Odonto-Immunomodulatory Properties for Modulating the Immune Response of Macrophages and Promoting Odontogenesis. Stem Cell Res. Ther. 2020, 11, 517. [Google Scholar] [CrossRef]
- Yu, S.; Chen, X.; Liu, Y.; Zhuang, X.Y.; Wang, A.C.; Liu, X.M.; Zhu, S. Exosomes Derived from Stem Cells from the Apical Papilla Alleviate Inflammation in Rat Pulpitis by Upregulating Regulatory T Cells. Int. Endod. J. 2022, 55, 517–530. [Google Scholar] [CrossRef]
- Zhang, S.; Thiebes, A.L.; Kreimendahl, F.; Rüetten, S.; Buhl, E.M.; Wolf, M.; Jockenhoevel, S.; Apel, C. Extracellular Vesicles-Loaded Fibrin Gel Supports Rapid Neovascularization for Dental Pulp Regeneration. Int. J. Mol. Sci. 2020, 21, 4226. [Google Scholar] [CrossRef] [PubMed]
- Ivica, A.; Ghayor, C.; Zehnder, M.; Valdec, S.; Weber, F.E. Pulp-Derived Exosomes in a Fibrin-Based Regenerative Root Filling Material. J. Clin. Med. 2020, 9, 491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Ju, Y.; Liu, S.; Fu, Y.; Zhao, S. Exosomes Derived from Lipopolysaccharide-Preconditioned Human Dental Pulp Stem Cells Regulate Schwann Cell Migration and Differentiation. Connect. Tissue Res. 2021, 62, 277–286. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Qiu, W.; Pan, Y.; Li, J.; Chen, Z.; Zhang, K.; Luo, Y.; Wu, B.; Xu, W. Exosomes from Lps-Stimulated Hdpscs Activated the Angiogenic Potential of Huvecs in Vitro. Stem Cells Int. 2021, 2021, 6685307. [Google Scholar] [CrossRef] [PubMed]
- Wen, B.; Huang, Y.; Qiu, T.; Huo, F.; Xie, L.; Liao, L.; Tian, W.; Guo, W. Reparative Dentin Formation by Dentin Matrix Proteins and Small Extracellular Vesicles. J. Endod. 2021, 47, 253–262. [Google Scholar] [CrossRef]
- Bagio, D.A.; Julianto, I.; Margono, A.; Suprastiwi, E. Analysis of Thrombin-Activated Platelet-Derived Exosome (T-Apde) Potential for Dental Pulp Regeneration: In-Vitro Study. Eur. J. Dent. 2022. [Google Scholar] [CrossRef]
- Yang, S.; Liu, Q.; Chen, S.; Zhang, F.; Li, Y.; Fan, W.; Mai, L.; He, H.; Huang, F. Extracellular Vesicles Delivering Nuclear Factor I/C for Hard Tissue Engineering: Treatment of Apical Periodontitis and Dentin Regeneration. J. Tissue Eng. 2022, 13, 20417314221084095. [Google Scholar] [CrossRef]
- Reynolds, M.A.; Kao, R.T.; Camargo, P.M.; Caton, J.G.; Clem, D.S.; Fiorellini, J.P.; Geisinger, M.L.; Mills, M.P.; Nares, S.; Nevins, M.L. Periodontal Regeneration—Intrabony Defects: A Consensus Report from the Aap Regeneration Workshop. J. Periodontol. 2015, 86, S105–S107. [Google Scholar] [CrossRef] [Green Version]
- Tassi, S.A.; Sergio, N.Z.; Misawa, M.Y.O.; Villar, C.C. Efficacy of Stem Cells on Periodontal Regeneration: Systematic Review of Pre-Clinical Studies. J. Periodontal Res. 2017, 52, 793–812. [Google Scholar] [CrossRef]
- Chu, C.; Wei, S.; Wang, Y.; Wang, Y.; Man, Y.; Qu, Y. Extracellular Vesicle and Mesenchymal Stem Cells in Bone Regeneration: Recent Progress and Perspectives. J. Biomed. Mater. Res. Part A 2019, 107, 243–250. [Google Scholar] [CrossRef]
- Chew, J.R.J.; Chuah, S.J.; Teo, K.Y.W.; Zhang, S.; Lai, R.C.; Fu, J.H.; Lim, L.P.; Lim, S.K.; Toh, W.S. Mesenchymal Stem Cell Exosomes Enhance Periodontal Ligament Cell Functions and Promote Periodontal Regeneration. Acta Biomater. 2019, 89, 252–264. [Google Scholar] [CrossRef]
- Trubiani, O.; Pizzicannella, J.; Caputi, S.; Marchisio, M.; Mazzon, E.; Paganelli, R.; Paganelli, A.; Diomede, F. Periodontal Ligament Stem Cells: Current Knowledge and Future Perspectives. Stem Cells Dev. 2019, 28, 995–1003. [Google Scholar] [CrossRef]
- Lei, F.; Li, M.; Lin, T.; Zhou, H.; Wang, F.; Su, X. Treatment of Inflammatory Bone Loss in Periodontitis by Stem Cell-Derived Exosomes. Acta Biomater. 2022, 141, 333–343. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Rao, N.; Jiang, H.; Dai, Y.; Yang, S.; Yang, H.; Hu, J. Small Extracellular Vesicles from Dental Follicle Stem Cells Provide Biochemical Cues for Periodontal Tissue Regeneration. Stem Cell Res. Ther. 2022, 13, 92. [Google Scholar] [CrossRef] [PubMed]
- Wei, J.; Song, Y.; Du, Z.; Yu, F.; Zhang, Y.; Jiang, N.; Ge, X. Exosomes Derived from Human Exfoliated Deciduous Teeth Ameliorate Adult Bone Loss in Mice through Promoting Osteogenesis. J. Mol. Histol. 2020, 51, 455–466. [Google Scholar] [CrossRef] [PubMed]
- Luo, L.; Avery, S.J.; Waddington, R.J. Exploring a Chemotactic Role for Evs from Progenitor Cell Populations of Human Exfoliated Deciduous Teeth for Promoting Migration of Naïve Bmscs in Bone Repair Process. Stem Cells Int. 2021, 2021, 6681771. [Google Scholar] [CrossRef]
- Zarubova, J.; Hasani-Sadrabadi, M.M.; Dashtimoghadam, E.; Zhang, X.; Ansari, S.; Li, S.; Moshaverinia, A. Engineered Delivery of Dental Stem-Cell-Derived Extracellular Vesicles for Periodontal Tissue Regeneration. Adv. Healthc. Mater. 2022, 11, e2102593. [Google Scholar] [CrossRef] [PubMed]
- Shen, Z.; Kuang, S.; Zhang, Y.; Yang, M.; Qin, W.; Shi, X.; Lin, Z. Chitosan Hydrogel Incorporated with Dental Pulp Stem Cell-Derived Exosomes Alleviates Periodontitis in Mice Via a Macrophage-Dependent Mechanism. Bioact. Mater. 2020, 5, 1113–1126. [Google Scholar] [CrossRef]
- Liu, L.; Guo, S.; Shi, M.W.; Liu, Q.; Huo, F.; Wu, Y.; Tian, W. Bone Marrow Mesenchymal Stem Cell-Derived Small Extracellular Vesicles Promote Periodontal Regeneration. Tissue Eng. Part A 2021, 27, 962–976. [Google Scholar] [CrossRef]
- Nakao, Y.; Fukuda, T.; Zhang, Q.; Sanui, T.; Shinjo, T.; Kou, X.; Chen, C.; Liu, D.; Watanabe, Y.; Hayashi, C.; et al. Exosomes from Tnf-Alpha-Treated Human Gingiva-Derived Mscs Enhance M2 Macrophage Polarization and Inhibit Periodontal Bone Loss. Acta Biomater. 2021, 122, 306–324. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.; Huang, C.-C.; Gajendrareddy, P.; Lu, Y.; Shirazi, S.; Ravindran, S.; Cooper, L.F. Extracellular Vesicles from Tnfα Preconditioned Mscs: Effects on Immunomodulation and Bone Regeneration. Front. Immunol. 2022, 13, 878194. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Guo, S.; Liu, L.; Liu, Q.; Huo, F.; Ding, Y.; Tian, W. Small Extracellular Vesicles from Lipopolysaccharide-Preconditioned Dental Follicle Cells Promote Periodontal Regeneration in an Inflammatory Microenvironment. ACS Biomater. Sci. Eng. 2020, 6, 5797–5810. [Google Scholar] [CrossRef] [PubMed]
- Mohammed, E.; Khalil, E.; Sabry, D. Effect of Adipose-Derived Stem Cells and Their Exo as Adjunctive Therapy to Nonsurgical Periodontal Treatment: A Histologic and Histomorphometric Study in Rats. Biomolecules 2018, 8, 167. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yi, G.; Zhang, S.; Ma, Y.; Yang, X.; Huo, F.; Chen, Y.; Yang, B.; Tian, W. Matrix Vesicles from Dental Follicle Cells Improve Alveolar Bone Regeneration Via Activation of the Plc/Pkc/Mapk Pathway. Stem Cell Res. Ther. 2022, 13, 41. [Google Scholar] [CrossRef]
- Klyachko, N.L.; Arzt, C.J.; Li, S.M.; Gololobova, O.A.; Batrakova, E.V. Extracellular Vesicle-Based Therapeutics: Preclinical and Clinical Investigations. Pharmaceutics 2020, 12, 1171. [Google Scholar] [CrossRef]
- Gandham, S.; Su, X.; Wood, J.; Nocera, A.L.; Alli, S.C.; Milane, L.; Zimmerman, A.; Amiji, M.; Ivanov, A.R. Technologies and Standardization in Research on Extracellular Vesicles. Trends Biotechnol. 2020, 38, 1066–1098. [Google Scholar] [CrossRef]
- Momen-Heravi, F. Isolation of Extracellular Vesicles by Ultracentrifugation. Methods Mol. Biol. 2017, 1660, 25–32. [Google Scholar] [CrossRef]
- McNamara, R.P.; Caro-Vegas, C.P.; Costantini, L.M.; Landis, J.T.; Griffith, J.D.; Damania, B.A.; Dittmer, D.P. Large-Scale, Cross-Flow Based Isolation of Highly Pure and Endocytosis-Competent Extracellular Vesicles. J. Extracell. Vesicles 2018, 7, 1541396. [Google Scholar] [CrossRef] [Green Version]
- Buschmann, D.; Mussack, V.; Byrd, J.B. Separation, Characterization, and Standardization of Extracellular Vesicles for Drug Delivery Applications. Adv. Drug Deliv. Rev. 2021, 174, 348–368. [Google Scholar] [CrossRef]
- Guo, S.-C.; Tao, S.-C.; Dawn, H. Microfluidics-Based on-a-Chip Systems for Isolating and Analysing Extracellular Vesicles. J. Extracell. Vesicles 2018, 7, 1508271. [Google Scholar] [CrossRef]
- Chen, M.; Lin, S.; Zhou, C.; Cui, D.; Haick, H.; Tang, N. From Conventional to Microfluidic: Progress in Extracellular Vesicle Separation and Individual Characterization. Adv. Healthc. Mater. 2022, e2202437. [Google Scholar] [CrossRef] [PubMed]
- Goss, D.M.; Vasilescu, S.A.; Sacks, G.; Gardner, D.K.; Warkiani, M.E. Microfluidics Facilitating the Use of Small Extracellular Vesicles in Innovative Approaches to Male Infertility. Nat. Rev. Urol. 2022, 1–30. [Google Scholar] [CrossRef]
- Lui, P.P.Y.; Leung, Y.T. Practical Considerations for Translating Mesenchymal Stromal Cell-Derived Extracellular Vesicles from Bench to Bed. Pharmaceutics 2022, 14, 1684. [Google Scholar] [CrossRef] [PubMed]
- Sharma, A. Role of Stem Cell Derived Exosomes in Tumor Biology. Int. J. Cancer 2018, 142, 1086–1092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Milano, G.; Biemmi, V.; Lazzarini, E.; Balbi, C.; Ciullo, A.; Bolis, S.; Ameri, P.; Di Silvestre, D.; Mauri, P.; Barile, L.; et al. Intravenous Administration of Cardiac Progenitor Cell-Derived Exosomes Protects against Doxorubicin/Trastuzumab-Induced Cardiac Toxicity. Cardiovasc. Res. 2020, 116, 383–392. [Google Scholar] [CrossRef] [PubMed]




| Origin of EVs | Types of EVs | Study Models | Key Functions/Potential Molecular Mechanism | References |
|---|---|---|---|---|
| Adipose-derived stem cells | sEVs | In vitro and in vivo (ligature-induced periodontitis in rat) | Induced highly organized structures formation that was comparable to normal healthy periodontal tissue | Mohammed et al., 2018 [133] |
| Mesenchymal stem cells (MSCs) | sEVs | In vitro and in vivo (periodontal intrabony defect in rat) | Increased periodontal ligament (PDL) cell migration and proliferation through CD73-mediated adenosine receptor activation of pro-survival AKT and ERK signaling | Chew et al., 2019 [121] |
| Dental follicle cells (DFCs) | sEVs | In vitro and in vivo (ligature-induced periodontitis in rat) | Inhibited osteoclast formation and promoted periodontal regeneration via OPG/RANK/RANKL signaling pathway | Shi et al., 2020 [132] |
| DPSCs | sEVs | In vitro and in vivo (ligature-induced periodontitis in mouse) | Modulated macrophages from a pro-inflammatory phenotype to an anti-inflammatory phenotype in the periodontium of mice with periodontitis, which could be associated with miR-1246 | Shen et al., 2020 [128] |
| SHED | sEVs | In vitro and in vivo (ligature-induced periodontitis +Porphyromonas gingivalis inoculation in mouse) | Directly promoted BMSCs osteogenesis, differentiation, and bone formation | Wei et al., 2020 [125] |
| BMMSCs | sEVs | In vitro and in vivo (ligature-induced periodontitis in rat) | Promoted the regeneration of periodontal tissues/Regulated the inflammatory immune response via the OPG–RANKL–RANK signaling pathway | Liu et al., 2021 [129] |
| Gingival mesenchymal stem cells (GMSCs) | sEVs | In vitro and in vivo (ligature-induced periodontitis in mouse) | Induced the resolution of inflammation and prevented bone loss in the periodontal tissue/Inhibited osteoclastogenesis via the Wnt5a-mediated RANKL pathway | Nakao et al., 2021 [130] |
| Periodontal ligament stem cells (PDLSCs) | sEVs | In vitro and in vivo (surgically created periodontal defect and ligature-induced periodontitis +LPS inoculation in rat) | Accelerated bone formation in the defect of alveolar bone in rat models of periodontitis/Recovered the osteogenic differentiation capacity of inflammatory PDLSCs via suppressing the over-activation of canonical Wnt signaling | Lei et al., 2022 [123] |
| Dental follicle stem cells (DFSCs) | sEVs | In vitro and in vivo (surgically created periodontal defect in rat) | Promoted periodontal tissue regeneration/Enhanced the proliferation, migration, and osteogenic differentiation of PDLSCs through the p38 MAPK signaling pathway | Ma et al., 2022 [124] |
| DFCs | MVs | In vitro and in vivo (surgically created periodontal defect in rat) | Strengthened alveolar bone regeneration through activating PLC/PKC/MAPK pathways | Yi et al., 2022 [134] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lai, H.; Li, J.; Kou, X.; Mao, X.; Zhao, W.; Ma, L. Extracellular Vesicles for Dental Pulp and Periodontal Regeneration. Pharmaceutics 2023, 15, 282. https://doi.org/10.3390/pharmaceutics15010282
Lai H, Li J, Kou X, Mao X, Zhao W, Ma L. Extracellular Vesicles for Dental Pulp and Periodontal Regeneration. Pharmaceutics. 2023; 15(1):282. https://doi.org/10.3390/pharmaceutics15010282
Chicago/Turabian StyleLai, Hongbin, Jiaqi Li, Xiaoxing Kou, Xueli Mao, Wei Zhao, and Lan Ma. 2023. "Extracellular Vesicles for Dental Pulp and Periodontal Regeneration" Pharmaceutics 15, no. 1: 282. https://doi.org/10.3390/pharmaceutics15010282