
Cell-Penetrating Dabcyl-Containing Tetraarginines with Backbone Aromatics as Uptake Enhancers
 , , ,
, , ,  , and
, and
Abstract
:1. Introduction
2. Results
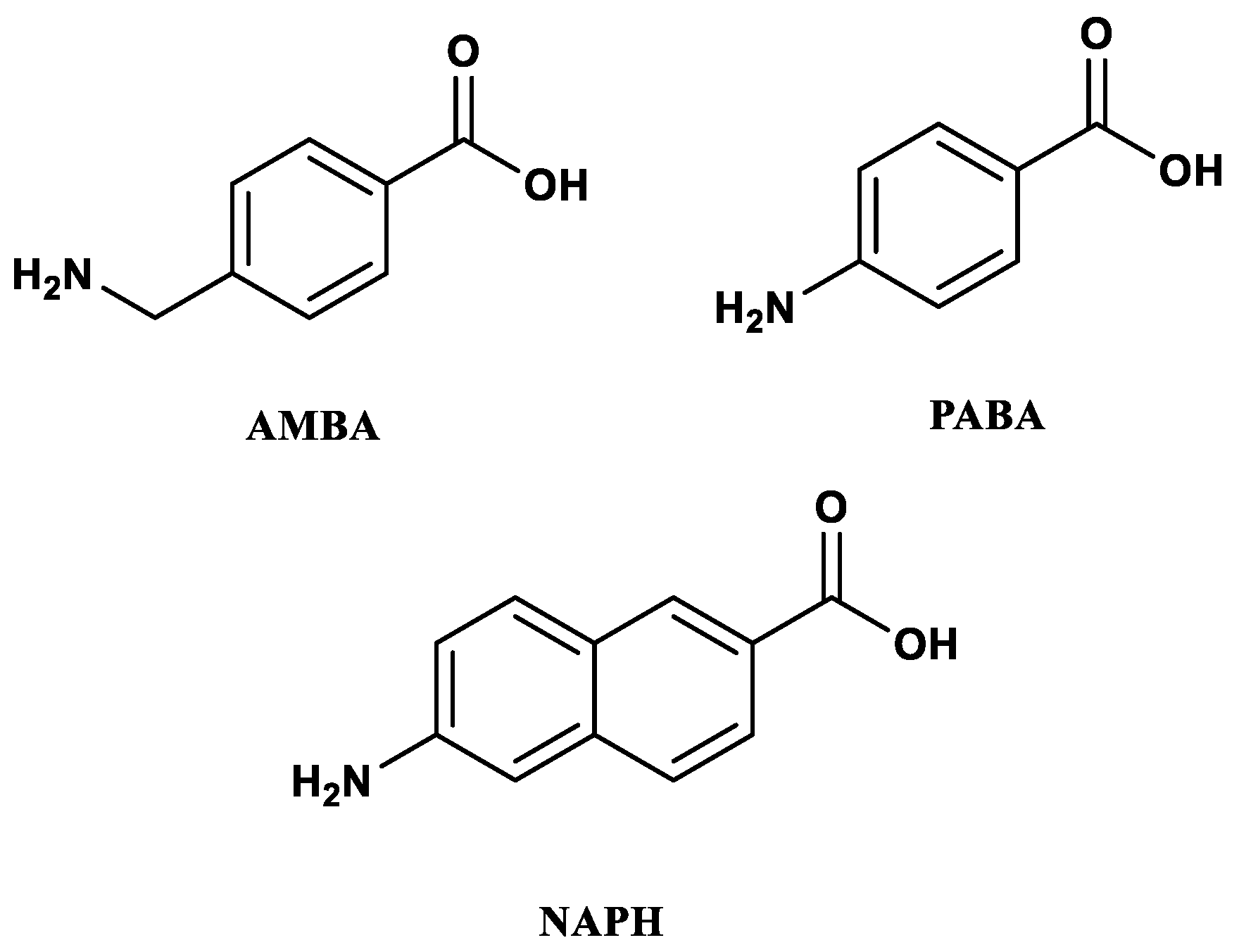
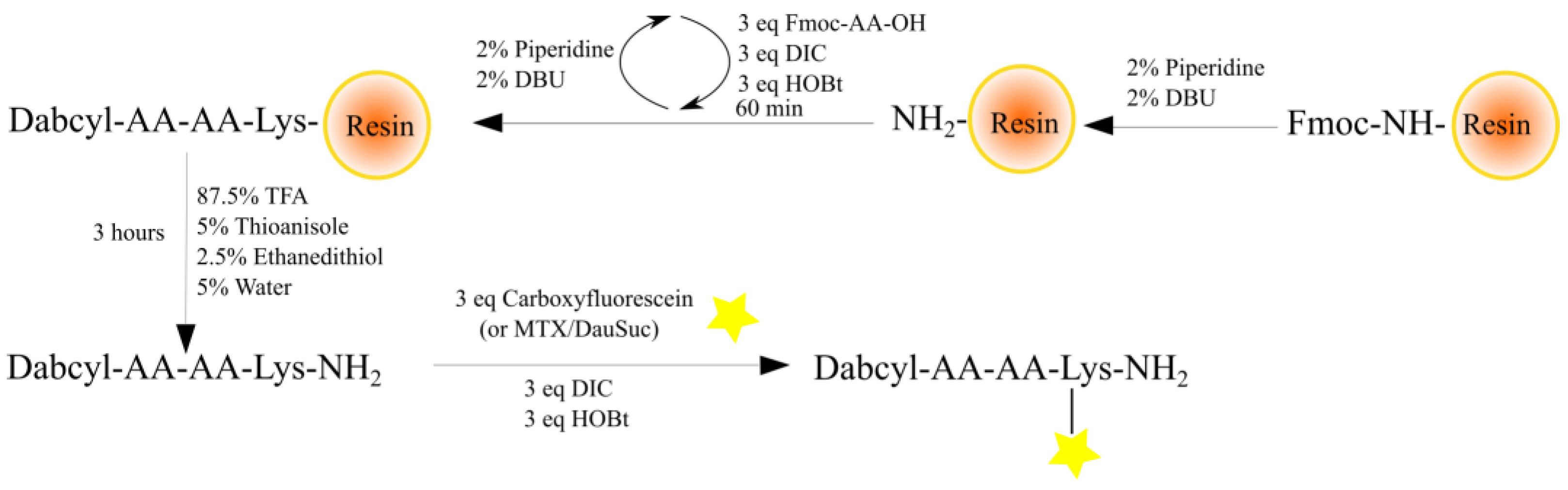
2.1. Synthesis of Peptides
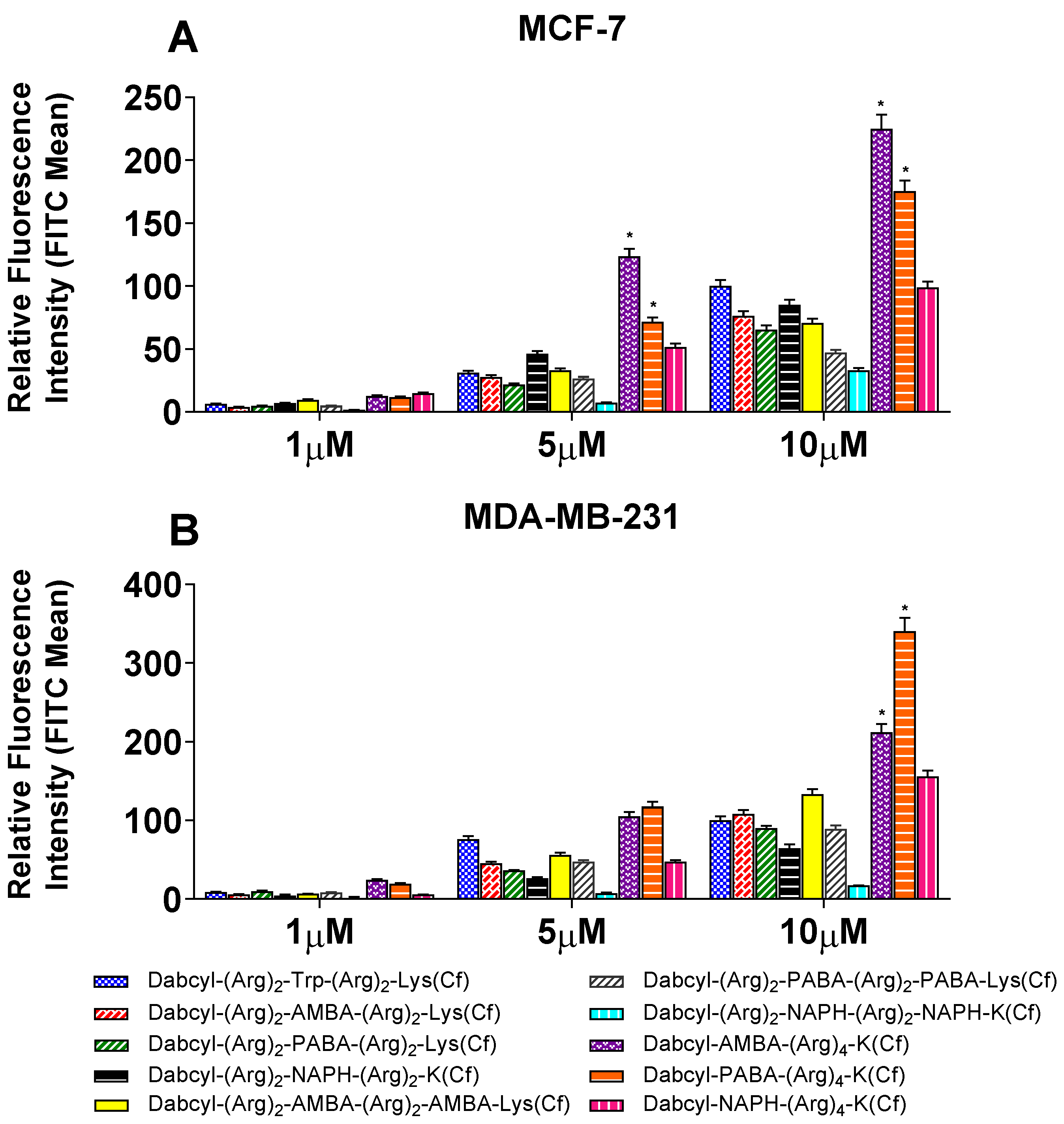
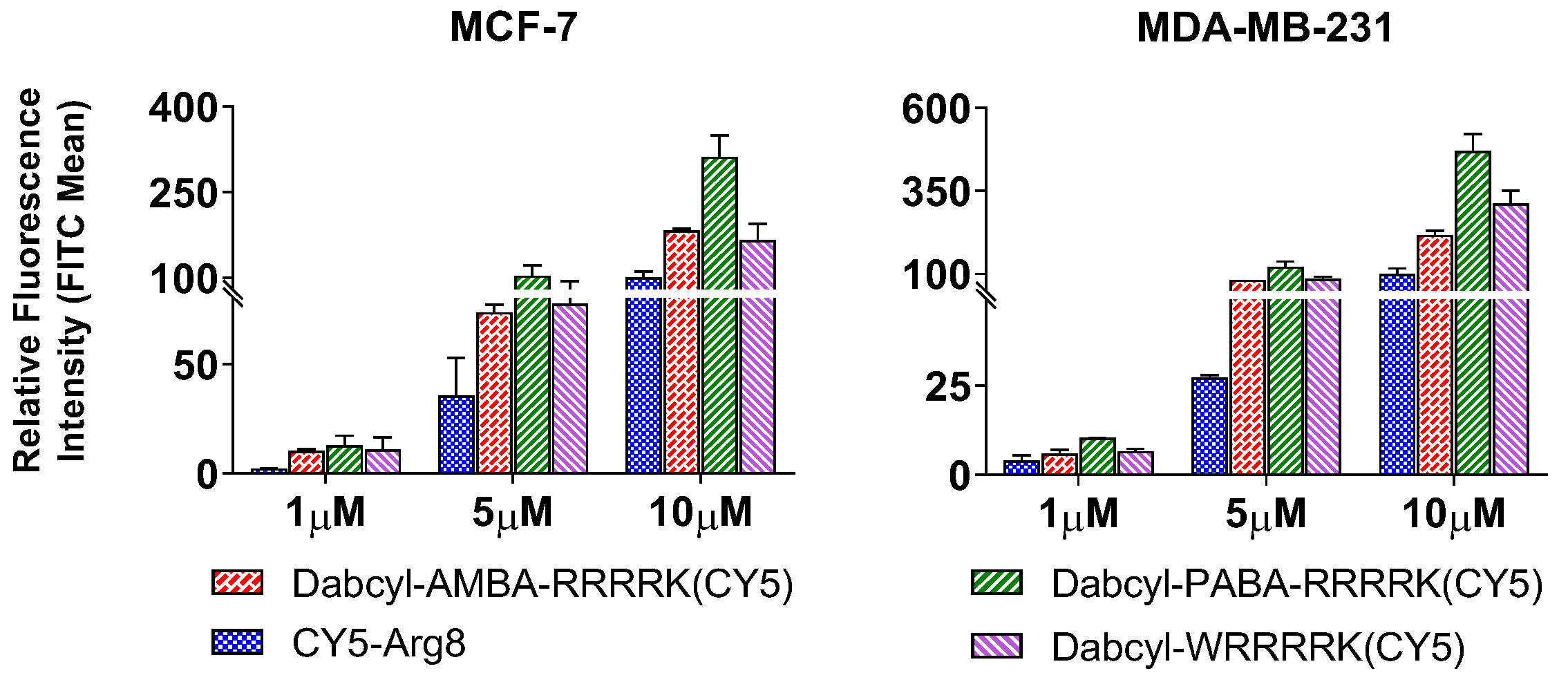
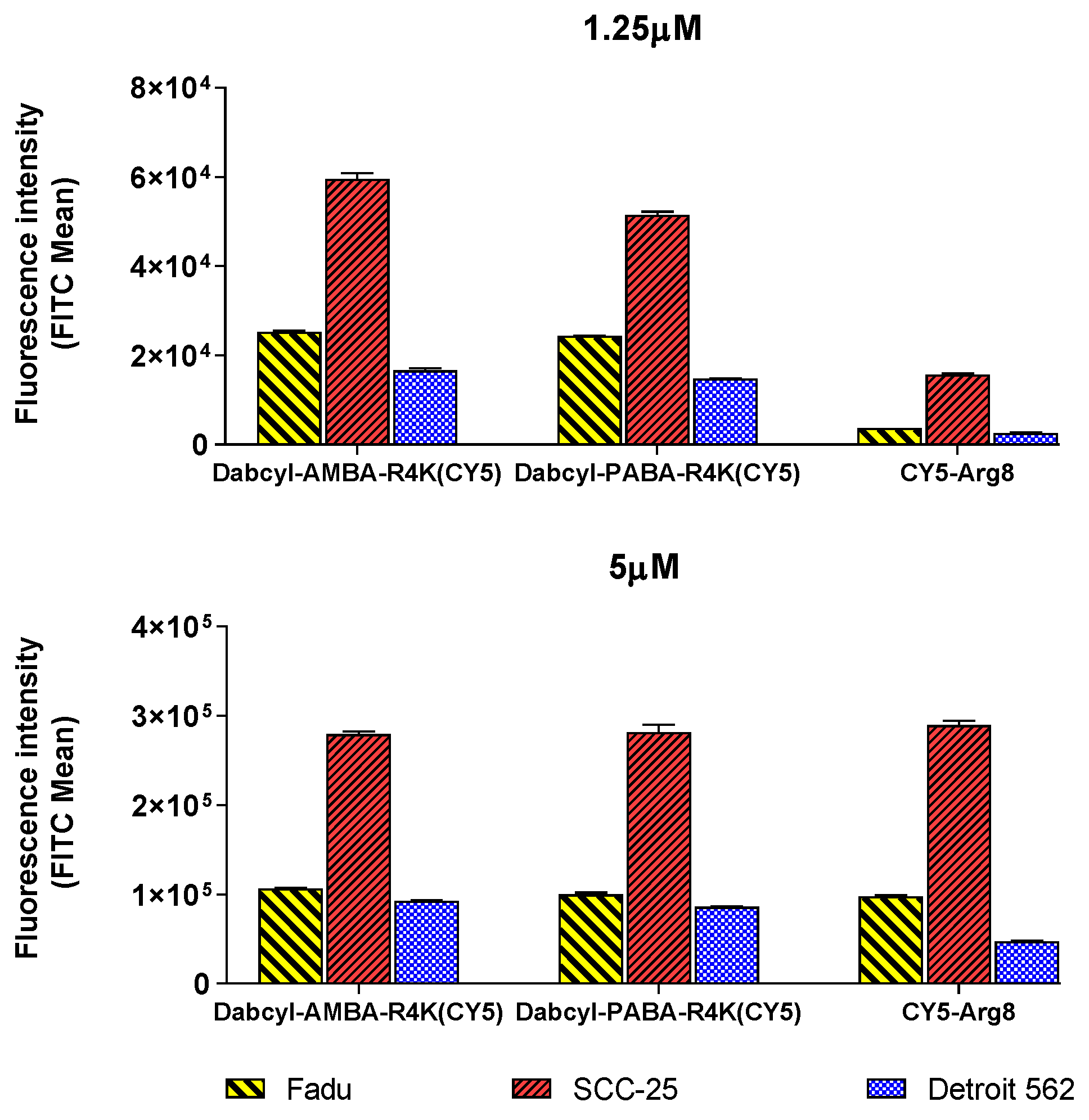
2.2. Cellular Uptake
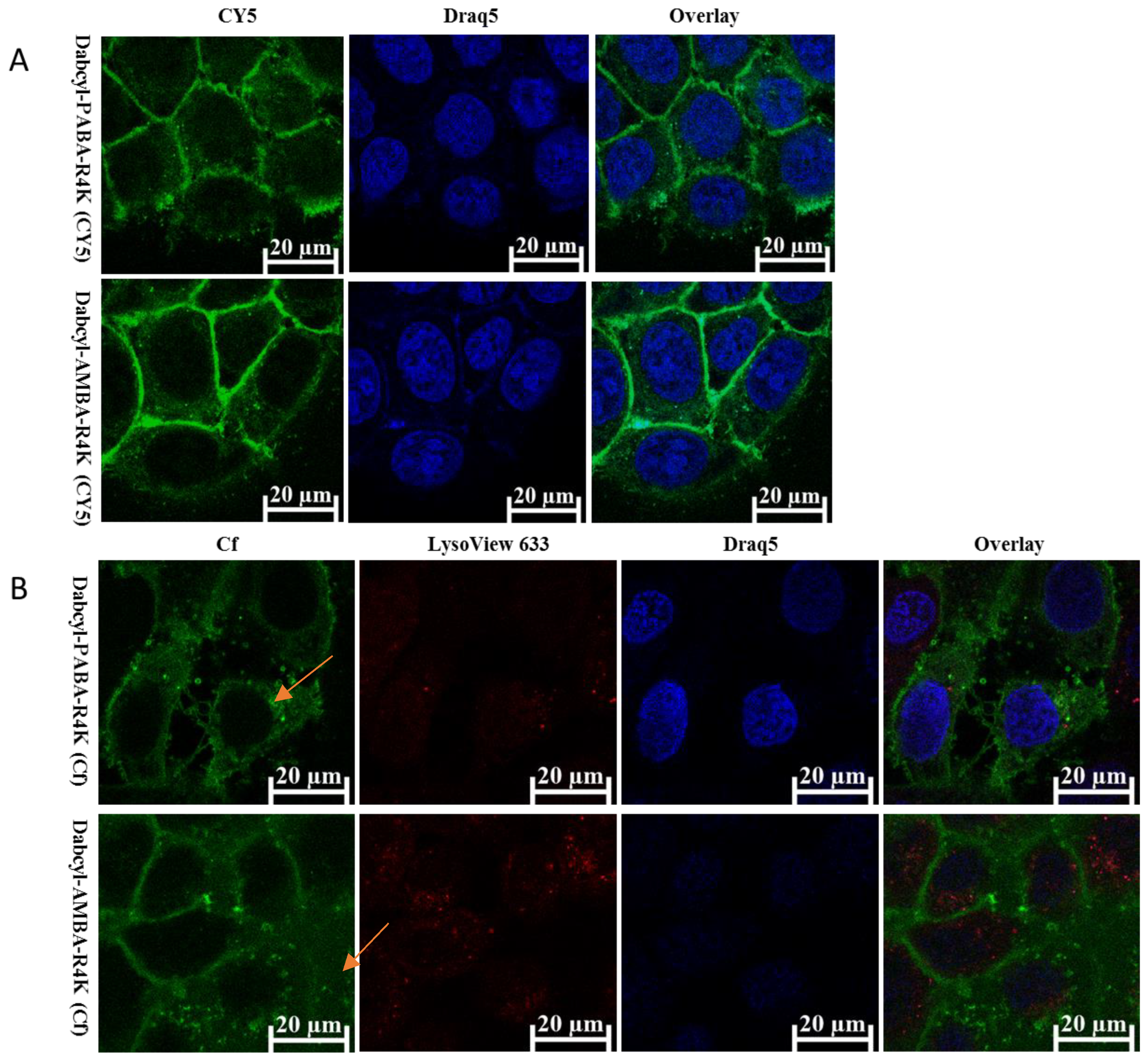
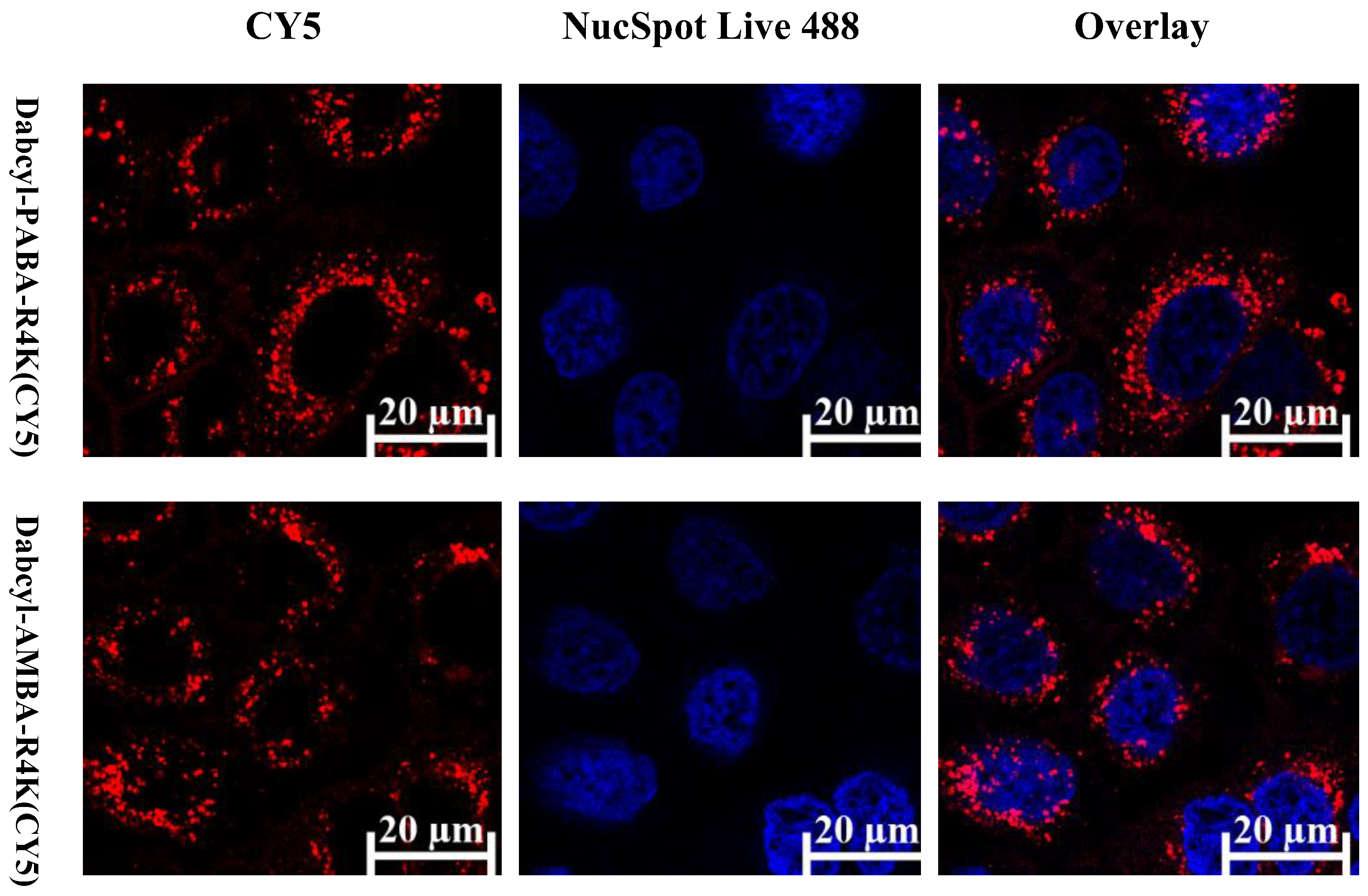
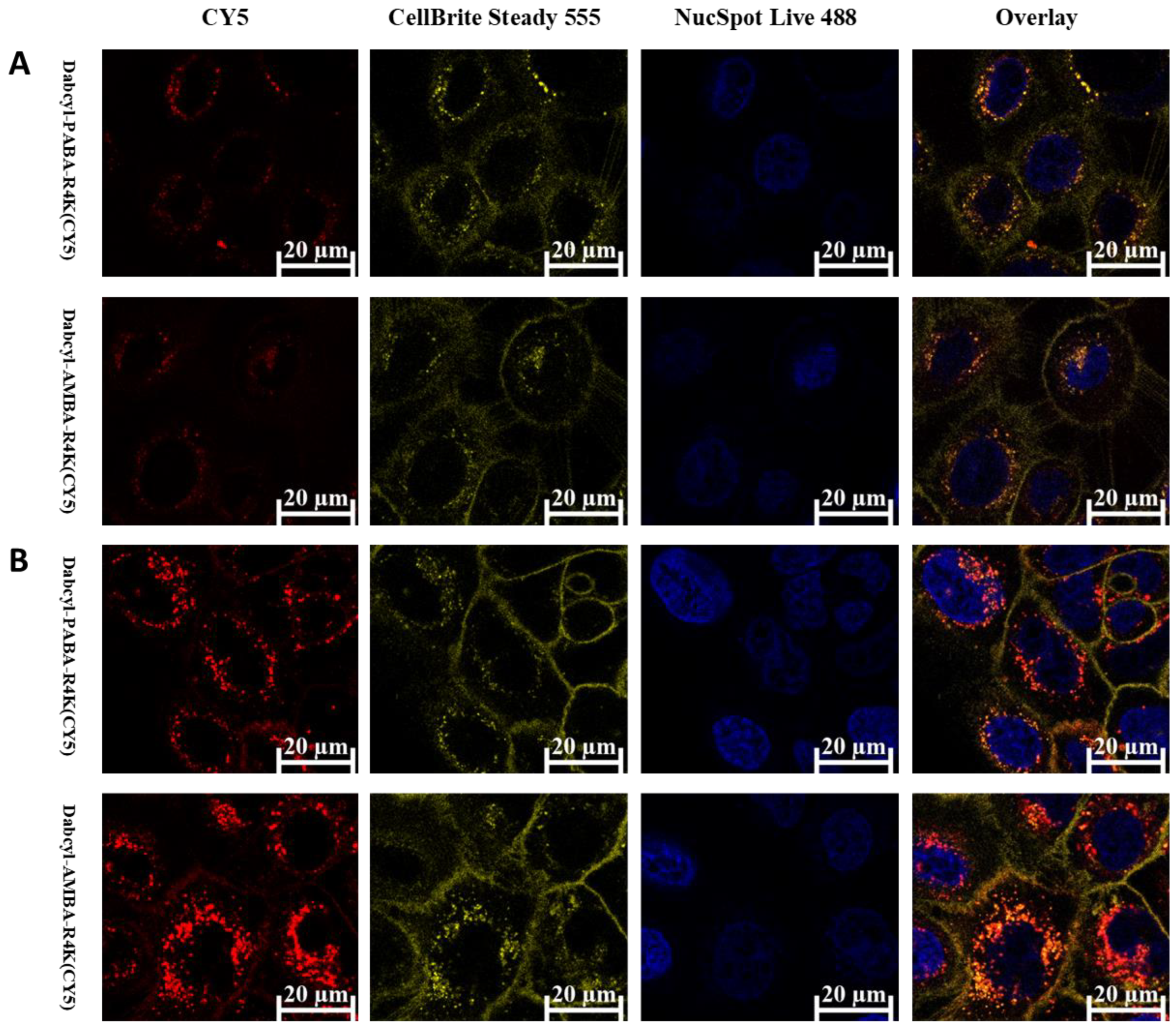
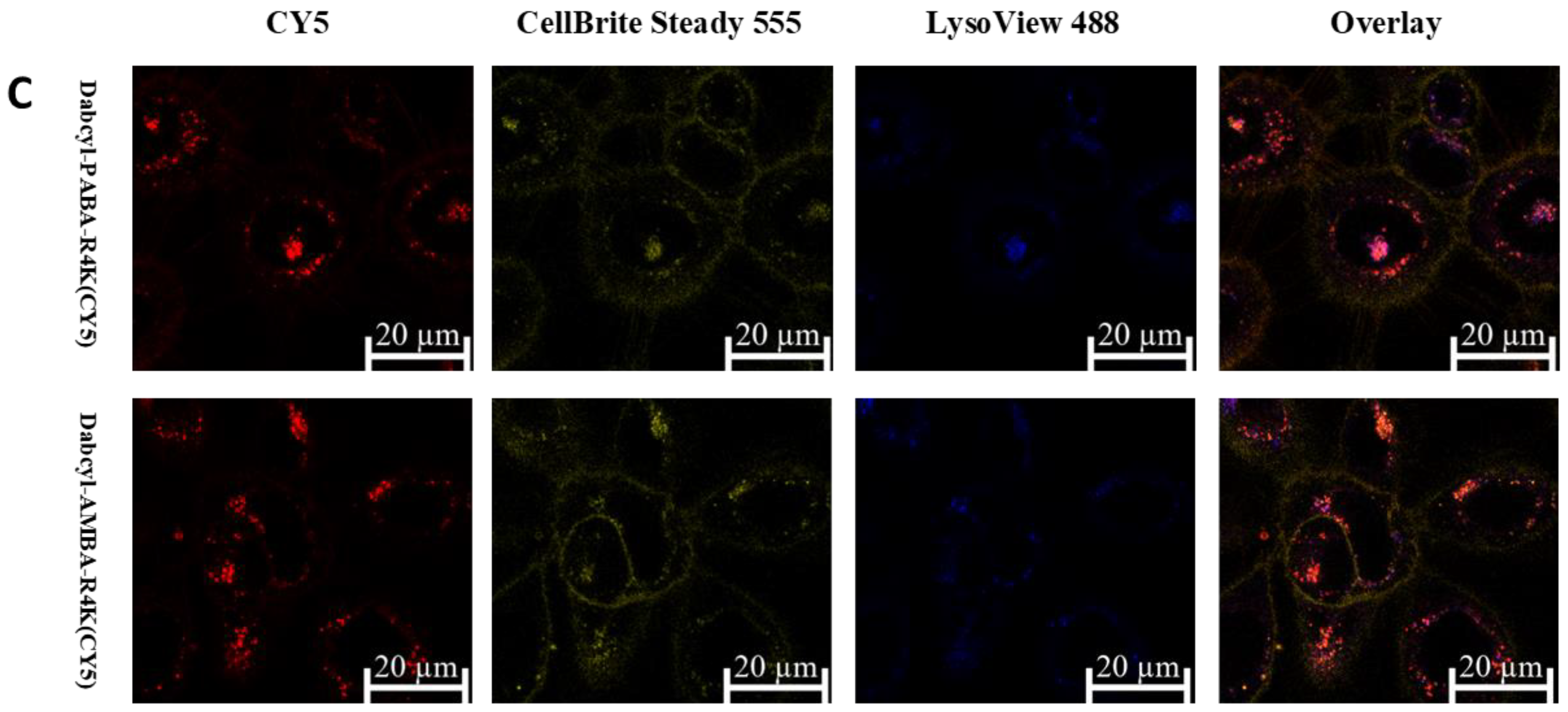
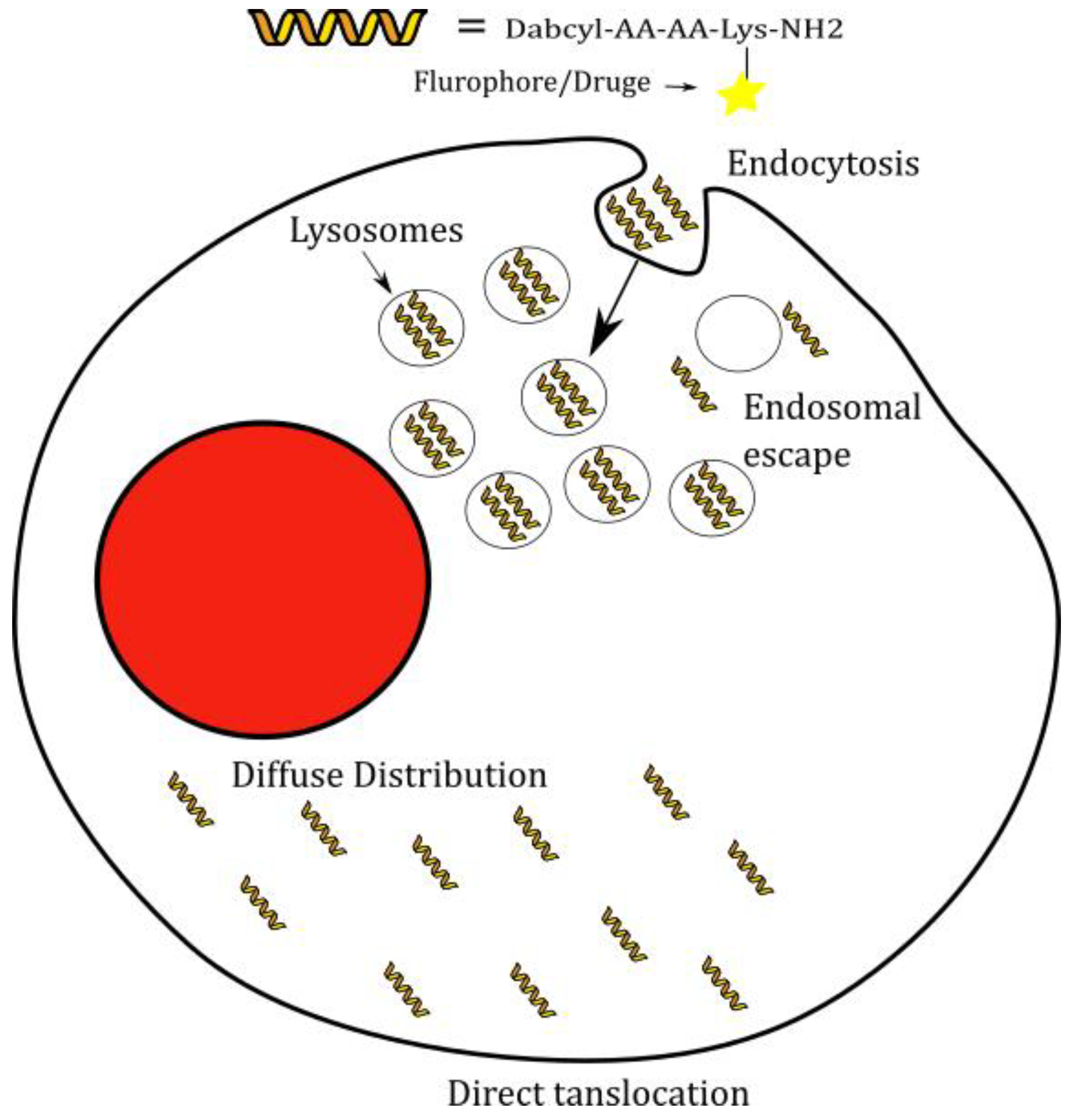
2.3. Cellular Distribution
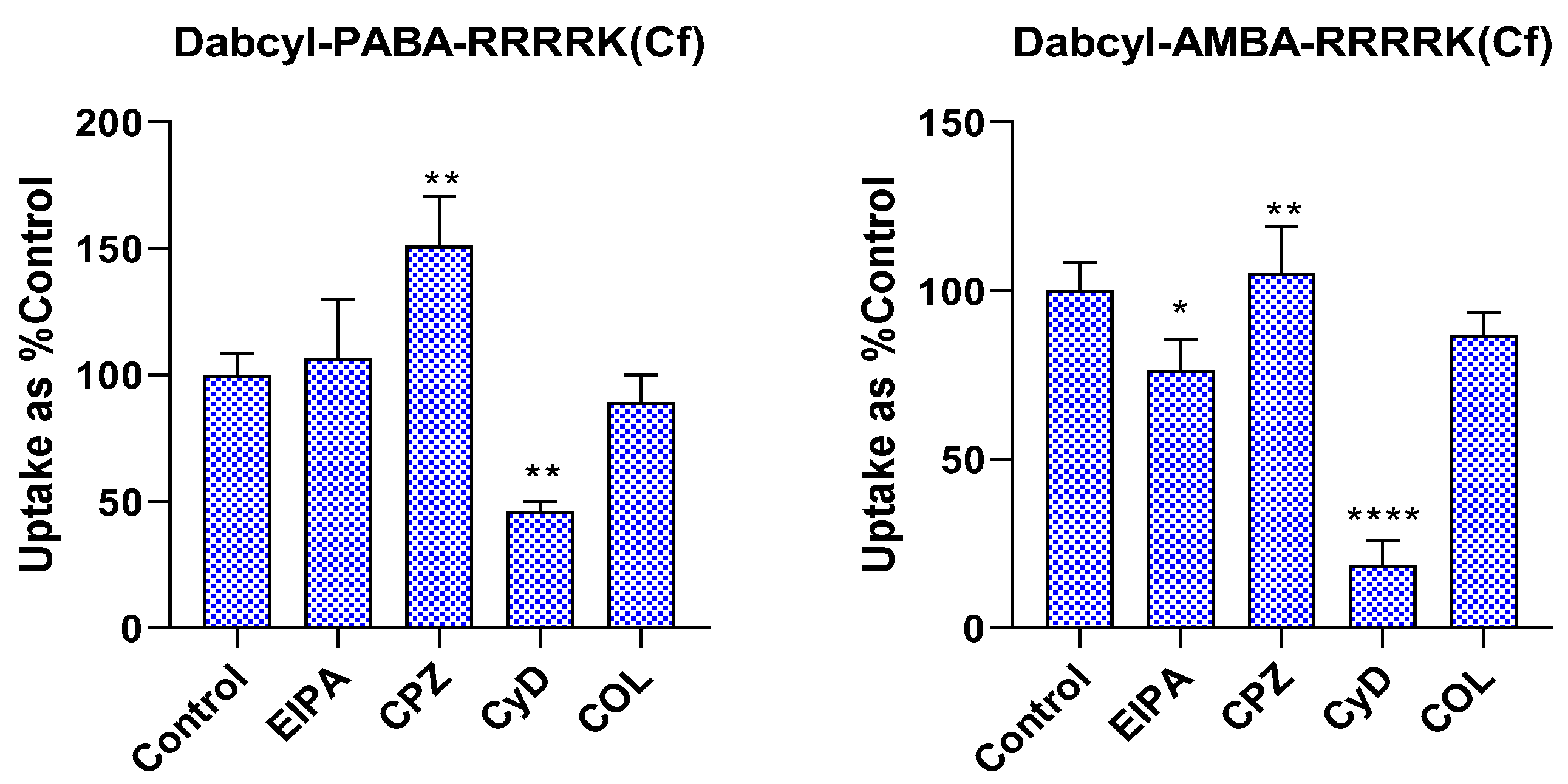
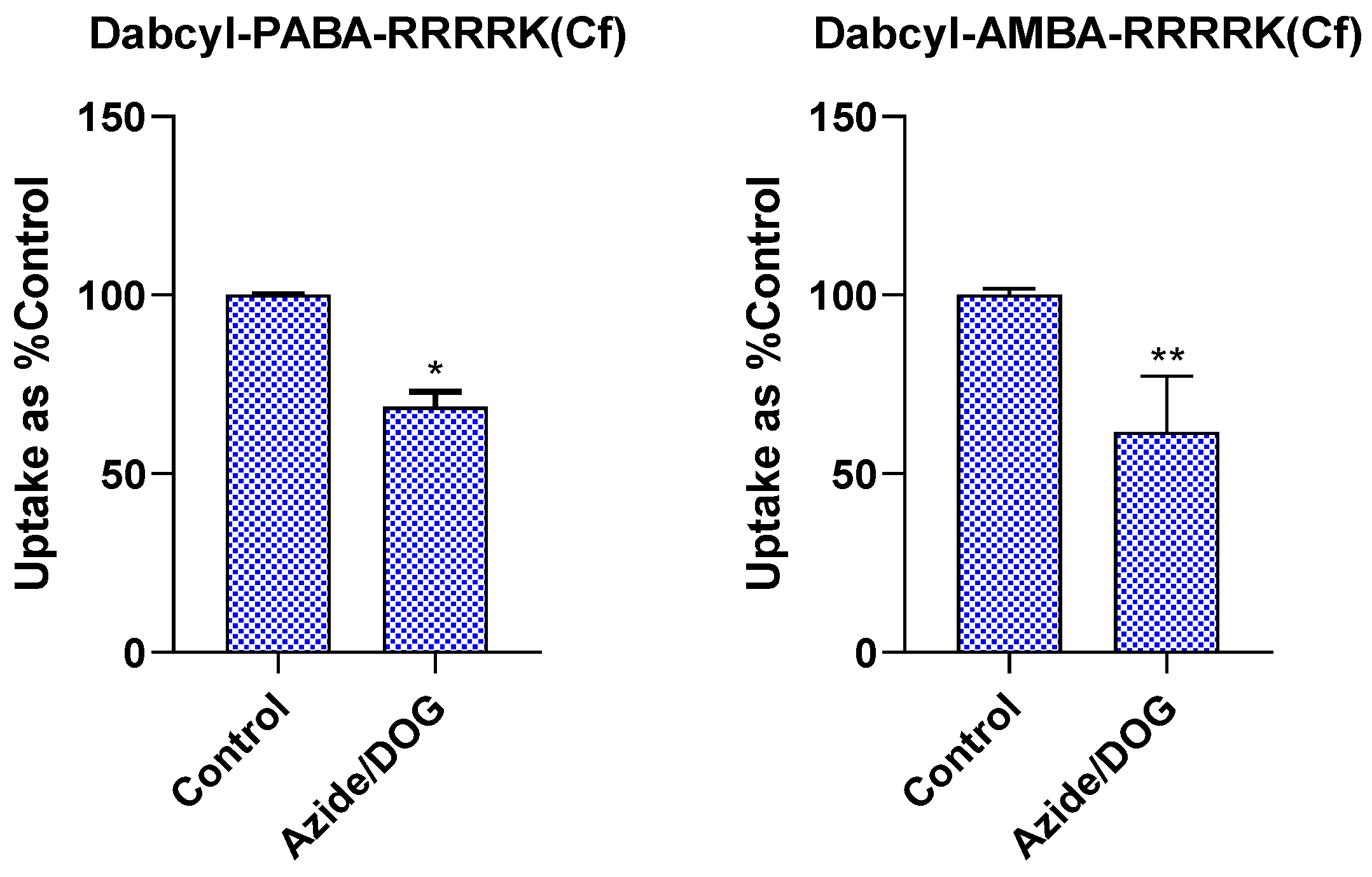
2.4. Investigation of Endocytic Pathways of Entry
2.5. Fluorimetry Based Measurement of Cellular Uptake at 4 °C and 37 °C
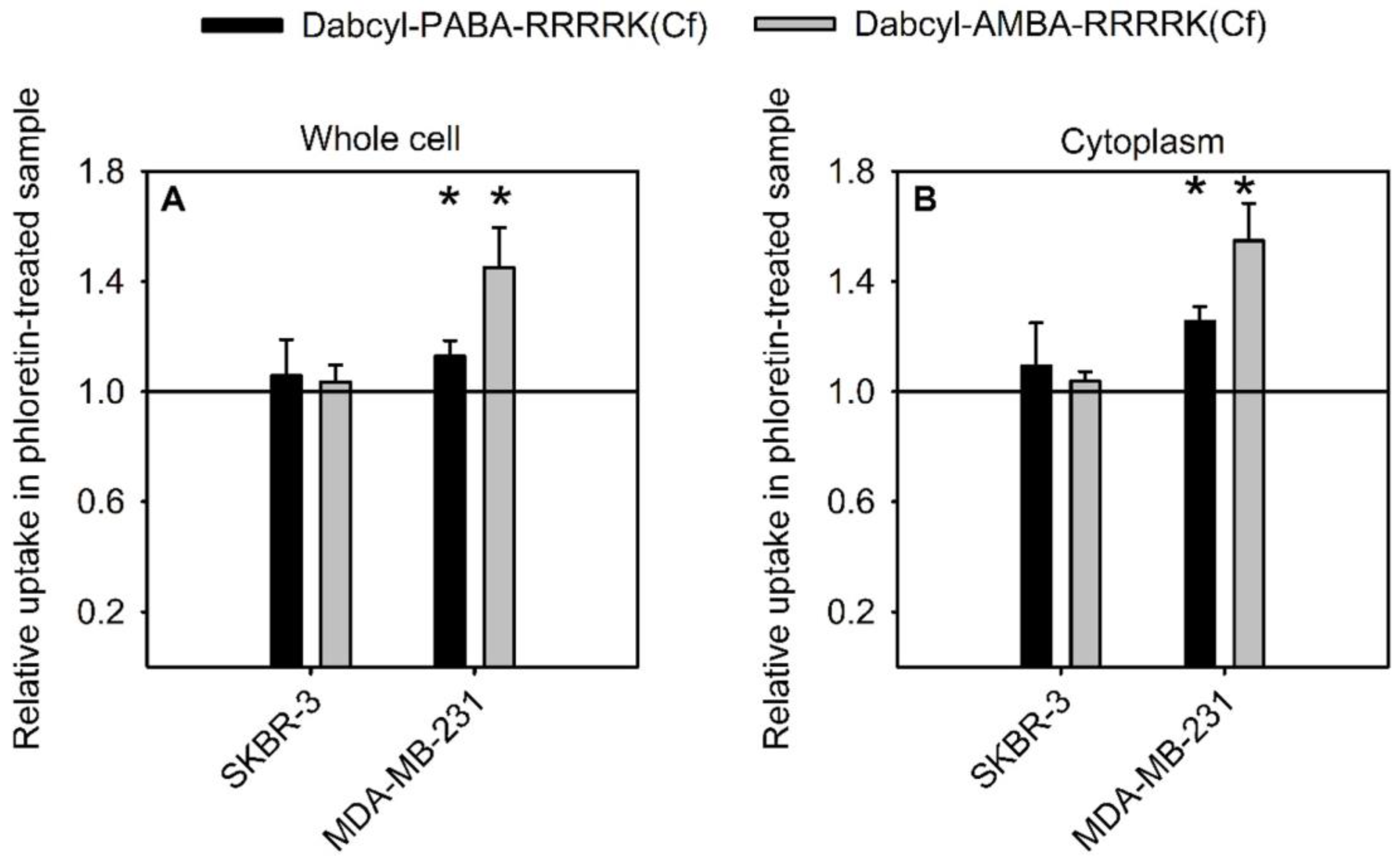
2.6. Effect of Membrane Dipole Potential on the Cellular Uptake
2.7. In Vitro Cytostatic Effect of Conjugates
3. Discussion
4. Materials and Methods
4.1. Synthesis of Peptides and Their Conjugates
4.2. Flow Cytometry
4.2.1. MCF-7 and MDA-MB-231
4.2.2. Detroit 562, FaDu and SCC-25
4.3. Internalization Quantification by Fluorometry of Lysed Cells at 37 °C or 4 °C
4.4. Confocal Microscopy
4.5. Study of the Effect of Membrane Dipole Potential on the Cellular Uptake
4.6. Analysis of In Vitro Cytostatic Activity of Conjugates
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bánóczi, Z.; Gorka-Kereskényi, Á.; Reményi, J.; Orbán, E.; Hazai, L.; Tokési, N.; Oláh, J.; Ovádi, J.; Béni, Z.; Háda, V.; et al. Synthesis and in vitro antitumor effect of vinblastine derivative- oligoarginine conjugates. Bioconjug. Chem. 2010, 21, 1948–1955. [Google Scholar] [CrossRef] [PubMed]
- Miklán, Z.; Orbán, E.; Csík, G.; Schlosser, G.; Magyar, A.; Hudecz, F. New daunomycin-oligoarginine conjugates: Synthesis, characterization, and effect on human leukemia and human hepatoma cells. Biopolymers 2009, 92, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Bánóczi, Z.; Keglevich, A.; Szabó, I.; Ranđelović, I.; Hegedüs, Z.; Regenbach, F.L.; Keglevich, P.; Lengyel, Z.; Gorka-Kereskényi, Á.; Dubrovay, Z.; et al. The effect of conjugation on antitumor activity of vindoline derivatives with octaarginine, a cell-penetrating peptide. J. Pept. Sci. 2018, 24, e3118. [Google Scholar] [CrossRef] [PubMed]
- Miklán, Z.; Szabó, R.; Zsoldos-Mády, V.; Reményi, J.; Bánóczi, Z.; Hudecz, F. New ferrocene containing peptide conjugates: Synthesis and effect on human leukemia (HL-60) cells. Biopolym.—Pept. Sci. Sect. 2007, 88, 108–114. [Google Scholar] [CrossRef]
- Miklán, Z.; Orbán, E.; Bánóczi, Z.; Hudecz, F. New pemetrexed-peptide conjugates: Synthesis, characterization and in vitro cytostatic effect on non-small cell lung carcinoma (NCI-H358) and human leukemia (HL-60) cells. J. Pept. Sci. 2011, 17, 805–811. [Google Scholar] [CrossRef]
- Szabó, I.; Orbán, E.; Schlosser, G.; Hudecz, F.; Bánóczi, Z. Cell-penetrating conjugates of pentaglutamylated methotrexate as potential anticancer drugs against resistant tumor cells. Eur. J. Med. Chem. 2016, 115, 361–368. [Google Scholar] [CrossRef] [Green Version]
- Bánóczi, Z.; Alexa, A.; Farkas, A.; Friedrich, P.; Hudecz, F. Novel cell-penetrating calpain substrate. Bioconjug. Chem. 2008, 19, 1375–1381. [Google Scholar] [CrossRef]
- Bánoczi, Z.; Tantos, Á.; Farkas, A.; Tompa, P.; Friedrich, P.; Hudecz, F. Synthesis of cell-penetrating conjugates of calpain activator peptides. Bioconjug. Chem. 2007, 18, 130–137. [Google Scholar] [CrossRef]
- Futaki, S.; Arafiles, J.V.V.; Hirose, H. Peptide-assisted intracellular delivery of biomacromolecules. Chem. Lett. 2020, 49, 1088–1094. [Google Scholar] [CrossRef]
- Jobin, M.-L.; Blanchet, M.; Henry, S.; Chaignepain, S.; Manigand, C.; Castano, S.; Lecomte, S.; Burlina, F.; Sagan, S.; Alves, I.D. The role of tryptophans on the cellular uptake and membrane interaction of arginine-rich cell penetrating peptides. Biochim. Biophys. Acta 2015, 1848, 593–602. [Google Scholar] [CrossRef]
- Jones, S.W.; Christison, R.; Bundell, K.; Voyce, C.J.; Brockbank, S.M.V.; Newham, P.; Lindsay, M.A. Characterisation of cell-penetrating peptide-mediated peptide delivery. Br. J. Pharmacol. 2005, 145, 1093–1102. [Google Scholar] [CrossRef] [PubMed]
- Hudecz, F.; Bánóczi, Z.; Csík, G. Medium-sized peptides as built in carriers for biologically active compounds. Med. Res. Rev. 2005, 25, 679–736. [Google Scholar] [CrossRef] [PubMed]
- Frankel, A.D.; Pabo, C.O. Cellular uptake of the tat protein from human immunodeficiency virus. Cell 1988, 55, 1189–1193. [Google Scholar] [CrossRef] [PubMed]
- Vivès, E.; Brodin, P.; Lebleu, B. A truncated HIV-1 Tat protein basic domain rapidly translocates through the plasma membrane and accumulates in the cell nucleus. J. Biol. Chem. 1997, 272, 16010–16017. [Google Scholar] [CrossRef] [Green Version]
- Derossi, D.; Joliot, A.H.; Chassaing, G.; Prochiantz, A. The third helix of the Antennapedia homeodomain translocates through biological membranes. J. Biol. Chem. 1994, 269, 10444–10450. [Google Scholar] [CrossRef]
- Ziegler, A.; Nervi, P.; Dürrenberger, M.; Seelig, J. The cationic cell-penetrating peptide CPPTAT derived from the HIV-1 protein TAT is rapidly transported into living fibroblasts: Optical, biophysical, and metabolic evidence. Biochemistry 2005, 44, 138–148. [Google Scholar] [CrossRef]
- Schmidt, N.; Mishra, A.; Lai, G.H.; Wong, G.C.L. Arginine-rich cell-penetrating peptides. FEBS Lett. 2010, 584, 1806–1813. [Google Scholar] [CrossRef] [Green Version]
- Khalil, I.A.; Kogure, K.; Futaki, S.; Harashima, H. High density of octaarginine stimulates macropinocytosis leading to efficient intracellular trafficking for gene expression. J. Biol. Chem. 2006, 281, 3544–3551. [Google Scholar] [CrossRef] [Green Version]
- Szabó, I.; Yousef, M.; Soltész, D.; Bató, C.; Mező, G.; Bánóczi, Z. Redesigning of Cell-Penetrating Peptides to Improve Their Efficacy as a Drug Delivery System. Pharmaceutics 2022, 14, 907. [Google Scholar] [CrossRef]
- Caesar, C.E.B.; Esbjörner, E.K.; Lincoln, P.; Nordén, B. Membrane interactions of cell-penetrating peptides probed by tryptophan fluorescence and dichroism techniques: Correlations of structure to cellular uptake. Biochemistry 2006, 45, 7682–7692. [Google Scholar] [CrossRef]
- Walrant, A.; Correia, I.; Jiao, C.Y.; Lequin, O.; Bent, E.H.; Goasdoué, N.; Lacombe, C.; Chassaing, G.; Sagan, S.; Alves, I.D. Different membrane behaviour and cellular uptake of three basic arginine-rich peptides. Biochim. Biophys. Acta-Biomembr. 2011, 1808, 382–393. [Google Scholar] [CrossRef] [PubMed]
- Bechara, C.; Pallerla, M.; Zaltsman, Y.; Burlina, F.; Alves, I.D.; Lequin, O.; Sagan, S. Tryptophan within basic peptide sequences triggers glycosaminoglycan- dependent endocytosis. FASEB J. 2013, 27, 738–749. [Google Scholar] [CrossRef] [PubMed]
- Bechara, C.; Pallerla, M.; Burlina, F.; Illien, F.; Cribier, S.; Sagan, S. Massive glycosaminoglycan-dependent entry of Trp-containing cell-penetrating peptides induced by exogenous sphingomyelinase or cholesterol depletion. Cell. Mol. Life Sci. 2015, 72, 809–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walrant, A.; Bauzá, A.; Girardet, C.; Alves, I.D.; Lecomte, S.; Illien, F.; Cardon, S.; Chaianantakul, N.; Pallerla, M.; Burlina, F.; et al. Ionpair-π interactions favor cell penetration of arginine/tryptophan-rich cell-penetrating peptides. Biochim. Biophys. Acta-Biomembr. 2020, 1862, 183098. [Google Scholar] [CrossRef]
- Shirani, A.; Mojarrad, J.S.; Farkhani, S.M.; Khosroshahi, A.Y.; Zakeri-Milani, P.; Samadi, N.; Sharifi, S.; Mohammadi, S.; Valizadeh, H. The relation between thermodynamic and structural properties and cellular uptake of peptides containing tryptophan and arginine. Adv. Pharm. Bull. 2015, 5, 161–168. [Google Scholar] [CrossRef] [Green Version]
- Rydberg, H.A.; Matson, M.; Amand, H.L.; Esbjörner, E.K.; Nordén, B. Effects of tryptophan content and backbone spacing on the uptake efficiency of cell-penetrating peptides. Biochemistry 2012, 51, 5531–5539. [Google Scholar] [CrossRef]
- Zakeri-Milani, P.; Farkhani, S.M.; Shirani, A.; Mohammadi, S.; Mojarrad, J.S.; Akbari, J.; Valizadeh, H. Cellular uptake and anti-tumor activity of gemcitabine conjugated with new amphiphilic cell penetrating peptides. EXCLI J. 2017, 16, 650–662. [Google Scholar] [CrossRef]
- Khemaissa, S.; Walrant, A.; Sagan, S. Tryptophan, more than just an interfacial amino acid in the membrane activity of cationic cell-penetrating and antimicrobial peptides. Q. Rev. Biophys. 2022, 55, e10. [Google Scholar] [CrossRef]
- Tyagi, S.; Bratu, D.P.; Kramer, F.R. Multicolor molecular beacons for allele discrimination. Nat. Biotechnol. 1998, 16, 49–53. [Google Scholar] [CrossRef]
- Moss, M.L.; Miller, M.A.; Vujanovic, N.; Yoneyama, T.; Rasmussen, F.H. Fluorescent substrates for ADAM15 useful for assaying and high throughput screening. Anal. Biochem. 2016, 514, 42–47. [Google Scholar] [CrossRef]
- Tsuji, M.; Ueda, S.; Hirayama, T.; Okuda, K.; Sakaguchi, Y.; Isono, A.; Nagasawa, H. FRET-based imaging of transbilayer movement of pepducin in living cells by novel intracellular bioreductively activatable fluorescent probes. Org. Biomol. Chem. 2013, 11, 3030–3037. [Google Scholar] [CrossRef] [PubMed]
- Szabó, I.; Illien, F.; Dókus, L.E.; Yousef, M.; Baranyai, Z.; Bősze, S.; Ise, S.; Kawano, K.; Sagan, S.; Futaki, S.; et al. Influence of the Dabcyl group on the cellular uptake of cationic peptides: Short oligoarginines as efficient cell-penetrating peptides. Amino Acids 2021, 53, 1033–1049. [Google Scholar] [CrossRef] [PubMed]
- Yousef, M.; Szabó, I.; Biri-Kovács, B.; Szeder, B.; Illien, F.; Sagan, S.; Bánóczi, Z. Modification of Short Non-Permeable Peptides to Increase Cellular Uptake and Cytostatic Activity of Their Conjugates. ChemistrySelect 2021, 6, 10111–10120. [Google Scholar] [CrossRef]
- Futaki, S.; Suzuki, T.; Ohashi, W.; Yagami, T.; Tanaka, S.; Ueda, K.; Sugiura, Y. Arginine-rich peptides. An abundant source of membrane-permeable peptides having potential as carriers for intracellular protein delivery. J. Biol. Chem. 2001, 276, 5836–5840. [Google Scholar] [CrossRef] [Green Version]
- Letoha, T.; Gaá, S.; Somlai, C.; Czajlik, A.; Perczel, A.; Penke, B. Membrane translocation of penetratin and its derivatives in different cell lines. J. Mol. Recognit. 2003, 16, 272–279. [Google Scholar] [CrossRef]
- Delaroche, D.; Aussedat, B.; Aubry, S.; Chassaing, G.; Burlina, F.; Clodic, G.; Bolbach, G.; Lavielle, S.; Sagan, S. Tracking a new cell-penetrating (W/R) nonapeptide, through an enzyme-stable mass spectrometry reporter tag. Anal. Chem. 2007, 79, 1932–1938. [Google Scholar] [CrossRef]
- Alvarado-González, M.; Gallo, M.; Lopez-Albarran, P.; Flores-Holguín, N.; Glossman-Mitnik, D. DFT study of the interaction between the conjugated fluorescein and dabcyl system, using fluorescene quenching method. J. Mol. Model. 2012, 18, 4113–4120. [Google Scholar] [CrossRef]
- Lansdorp, P.M.; Smith, C.; Safford, M.; Terstappen, L.W.M.M.; Thomas, T.E. Single laser three color immunofluorescence staining procedures based on energy transfer between phycoerythrin and cyanine 5. Cytometry 1991, 12, 723–730. [Google Scholar] [CrossRef]
- Koivusalo, M.; Welch, C.; Hayashi, H.; Scott, C.C.; Kim, M.; Alexander, T.; Touret, N.; Hahn, K.M.; Grinstein, S. Amiloride inhibits macropinocytosis by lowering submembranous pH and preventing Rac1 and Cdc42 signaling. J. Cell Biol. 2010, 188, 547–563. [Google Scholar] [CrossRef] [Green Version]
- Gomes dos Reis, L.; Lee, W.H.; Svolos, M.; Moir, L.M.; Jaber, R.; Engel, A.; Windhab, N.; Young, P.M.; Traini, D. Delivery of pDNA to lung epithelial cells using PLGA nanoparticles formulated with a cell-penetrating peptide: Understanding the intracellular fate. Drug Dev. Ind. Pharm. 2020, 46, 427–442. [Google Scholar] [CrossRef]
- Fittipaldi, A.; Ferrari, A.; Zoppé, M.; Arcangeli, C.; Pellegrini, V.; Beltram, F.; Giacca, M. Cell Membrane Lipid Rafts Mediate Caveolar Endocytosis of HIV-1 Tat Fusion Proteins. J. Biol. Chem. 2003, 278, 34141–34149. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Johnson, J.R.; Kocher, B.; Barnett, E.M.; Marasa, J.; Piwnica-Worms, D. Caspase-activated cell-penetrating peptides reveal temporal coupling between endosomal release and apoptosis in an RGC-5 cell model. Bioconjug. Chem. 2012, 23, 1783–1793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nolasco, S.; Bellido, J.; Serna, M.; Carmona, B.; Soares, H.; Zabala, J.C. Colchicine Blocks Tubulin Heterodimer Recycling by Tubulin Cofactors TBCA, TBCB, and TBCE. Front. Cell Dev. Biol. 2021, 9, 950. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Liu, K.; Manaloto, E.; Casey, A.; Cribaro, G.P.; Byrne, H.J.; Tian, F.; Barcia, C.; Conway, G.E.; Cullen, P.J.; et al. Cold Atmospheric Plasma Induces ATP-Dependent Endocytosis of Nanoparticles and Synergistic U373MG Cancer Cell Death. Sci. Rep. 2018, 8, 5298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gross, E.; Bedlack, R.S.; Loew, L.M. Dual-wavelength ratiometric fluorescence measurement of the membrane dipole potential. Biophys. J. 1994, 67, 208–216. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batta, G.; Kárpáti, L.; Henrique, G.F.; Tóth, G.; Tarapcsák, S.; Kovacs, T.; Zakany, F.; Mándity, I.M.; Nagy, P. Statin-boosted cellular uptake and endosomal escape of penetratin due to reduced membrane dipole potential. Br. J. Pharmacol. 2021, 178, 3667–3681. [Google Scholar] [CrossRef] [PubMed]
- Lättig-Tünnemann, G.; Prinz, M.; Hoffmann, D.; Behlke, J.; Palm-Apergi, C.; Morano, I.; Herce, H.D.; Cardoso, M.C. Backbone rigidity and static presentation of guanidinium groups increases cellular uptake of arginine-rich cell-penetrating peptides. Nat. Commun. 2011, 2, 453. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Zhang, Y.; Chen, S.W.; Li, F.J.; Zhuang, S.M.; Wang, L.P.; Zhang, J.; Song, M. Prognostic significance of Flotillin1 expression in clinically N0 tongue squamous cell cancer. Int. J. Clin. Exp. Pathol. 2014, 7, 996. [Google Scholar]
- Pang, L.; Yang, S.; Dai, W.; Wu, S.; Kong, J. Role of caveolin-1 in human organ function and disease: Friend or foe? Carcinogenesis 2022, 43, 2–11. [Google Scholar] [CrossRef]
- Richard, J.P.; Melikov, K.; Brooks, H.; Prevot, P.; Lebleu, B.; Chernomordik, L.V. Cellular uptake of unconjugated TAT peptide involves clathrin-dependent endocytosis and heparan sulfate receptors. J. Biol. Chem. 2005, 280, 15300–15306. [Google Scholar] [CrossRef] [Green Version]
- Kaplan, I.M.; Wadia, J.S.; Dowdy, S.F. Cationic TAT peptide transduction domain enters cells by macropinocytosis. J. Control. Release 2005, 102, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, G.; Nakase, I.; Fukuda, Y.; Masuda, R.; Oishi, S.; Shimura, K.; Kawaguchi, Y.; Takatani-Nakase, T.; Langel, Ü.; Gräslund, A.; et al. CXCR4 stimulates macropinocytosis: Implications for cellular uptake of arginine-rich cell-penetrating peptides and HIV. Chem. Biol. 2012, 19, 1437–1446. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kawaguchi, Y.; Takeuchi, T.; Kuwata, K.; Chiba, J.; Hatanaka, Y.; Nakase, I.; Futaki, S. Syndecan-4 Is a Receptor for Clathrin-Mediated Endocytosis of Arginine-Rich Cell-Penetrating Peptides. Bioconjug. Chem. 2016, 27, 1119–1130. [Google Scholar] [CrossRef] [PubMed]
- Cleal, K.; He, L.; Watson, P.D.; Jones, A.T. Endocytosis, Intracellular Traffic and Fate of Cell Penetrating Peptide Based Conjugates and Nanoparticles. Curr. Pharm. Des. 2013, 19, 2878–2894. [Google Scholar] [CrossRef]
- Shin, J.S.; Abraham, S.N. Caveolae as portals of entry for microbes. Microbes Infect. 2001, 3, 755–761. [Google Scholar] [CrossRef]
- Gawande, M.; Branco, P. An efficient and expeditious Fmoc protection of amines and amino acids in aqueous media. Green Chem. 2011, 13, 3355–3359. [Google Scholar] [CrossRef]
- Bánóczi, Z.; Peregi, B.; Orbán, E.; Szabó, R.; Hudecz, F. Synthesis of daunomycin-oligoarginine conjugates and their effect on human leukemia cells (HL-60). Arkivoc 2008, 2008, 140–153. [Google Scholar] [CrossRef]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sequence | Rt | Mcalc | Mmeas |
|---|---|---|---|
| Cf-Arg8 a | 11.4 | 1623.9 | 1624.4 |
| Dabcyl-(Arg)2-Trp-(Arg)2-Lys(Cf) | 14.7 | 1564.8 | 1565.6 |
| Dabcyl-AMBA-(Arg)4-Lys(Cf) | 14.9 | 1511.7 | 1511.6 |
| Dabcyl-(Arg)2-AMBA-(Arg)2-Lys(Cf) | 13.7 | 1511.7 | 1512.0 |
| Dabcyl-(Arg)2-AMBA-(Arg)2-AMBA-Lys(Cf) | 13.9 | 1644.8 | 1644.8 |
| Dabcyl-NAPH-(Arg)4-Lys(Cf) | 16.2 | 1547.7 | 1547.7 |
| Dabcyl-(Arg)2-NAPH-(Arg)2-Lys(Cf) | 13.8 | 1547.7 | 1547.7 |
| Dabcyl-(Arg)2-NAPH-(Arg)2-NAPH-Lys(Cf) | 14.2 | 1716.8 | 1716.4 |
| Dabcyl-(Arg)2-PABA-(Arg)2R-Lys(Cf) | 13.6 | 1497.7 | 1497.6 |
| Dabcyl-PABA-(Arg)4-Lys(Cf) | 15.3 | 1497.7 | 1497.3 |
| Dabcyl-(Arg)2-PABA-(Arg)2-PABA-Lys(Cf) | 13.6 | 1616.8 | 1616.8 |
| CY5-Arg8 b | 15.2 | 1731.1 | 1731.4 |
| Dabcyl-PABA-(Arg)4-Lys(CY5)b | 17.7 | 1605.0 | 1605.2 |
| Dabcyl-AMBA-(Arg)4-Lys(CY5)b | 17.6 | 1619.0 | 1619.1 |
| Dabcyl-AMBA-(Arg)4-Lys(DauSuc) | 16.1 | 1761.9 | 1761.6 |
| Dabcyl-PABA-(Arg)4- Lys(DauSuc) | 16.5 | 1747.8 | 1747.5 |
| Dabcyl-NAPH-(Arg)4-Lys(DauSuc) | 16.9 | 1797.9 | 1798.1 |
| Dabcyl-AMBA-(Arg)4-Lys(MTX) | 13.6 | 1589.8 | 1589.6 |
| Dabcyl-PABA-(Arg)4-Lys(MTX) | 13.9 | 1575.8 | 1575.6 |
| Dabcyl-NAPH-(Arg)4-Lys(MTX) | 14.6 | 1625.8 | 1625.6 |
| Dabcyl-AMBA-(Arg)4-Lys(GFLG-E5-MTX) | 14.1 | 2609.3 | 2608.4 |
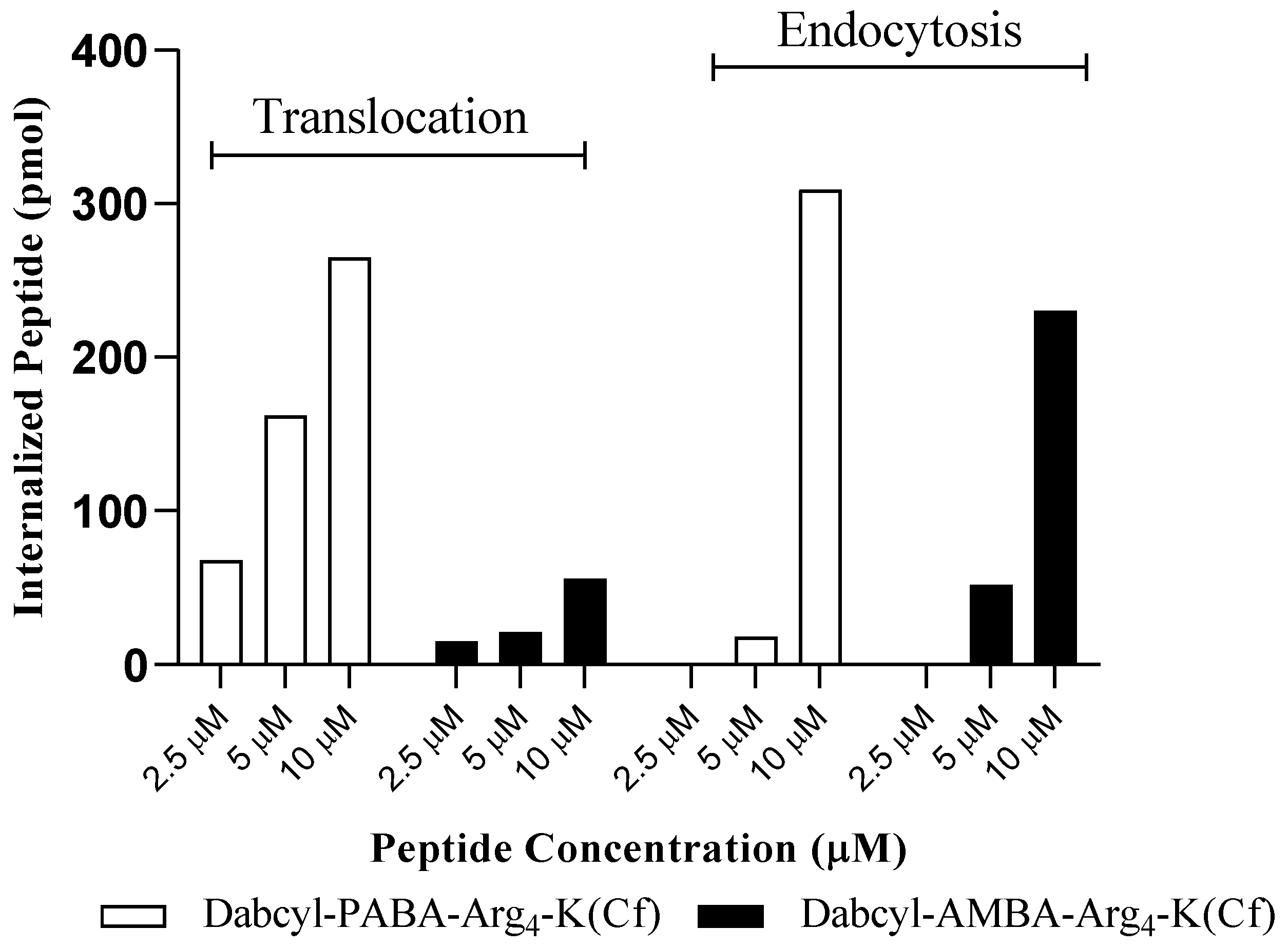
| Peptide | Conc. | Total Uptake (37 °C) | Direct Translocation (4 °C) | Endocytosis (Difference between Uptake at 37 and 4 °C) |
|---|---|---|---|---|
| Dabcyl-PABA-Arg4-K(Cf) | 2.5 µM | 50 ± 9 | 68 ± 14 | 0 |
| 5 µM | 180 ± 24 | 162 ± 33 | 18 | |
| 10 µM | 574 ± 100 | 265 ± 70 | 309 | |
| Dabcyl-AMBA-Arg4-K(Cf) | 2.5 µM | 15 ± 5 | 15 ± 5 | 0 |
| 5 µM | 73 ± 9 | 21 ± 5 | 52 | |
| 10 µM | 286 ± 19 | 56 ± 14 | 230 |
| Conjugate | IC50 ±SD (µM) a | |
|---|---|---|
| MCF-7 | MDA-MB-231 | |
| Dabcyl-AMBA-Arg-Arg-Arg-Arg-Lys(DauSuc) | 10.3 ± 5.0 | 10.1 ± 6.0 |
| Dabcyl-PABA- Arg-Arg-Arg-Arg-Lys(DauSuc) | 17.7 ± 1.2 | 14.3 ± 1.1 |
| Dabcyl-NAPH- Arg-Arg-Arg-Arg-Lys(DauSuc) | 15.0 ± 4.2 | 16.6 ± 0.5 |
| Dabcyl-Trp-Arg-Arg-Arg-Arg-Lys(DauSuc) b | 26.4 ± 4.2 | 18.0 ± 6.7 |
| DauSuc-Arg8 b | 14.4 ± 0.2 | 19.5 ± 0.4 |
| Dabcyl-AMBA-Arg-Arg-Arg-Arg-Lys(MTX) | 11.8 ± 6.9 | 19.2 ± 7.1 |
| Dabcyl-PABA- Arg-Arg-Arg-Arg-Lys(MTX) | 18.6 ± 3.3 | 16.8 ± 4.7 |
| Dabcyl-NAPH- Arg-Arg-Arg-Arg-Lys(MTX) | 14.8 ± 2.5 | 16.9 ± 2.8 |
| Dabcyl-AMBA-Arg-Arg-Arg-Arg-Lys(GFLG-E5-MTX) | 8.0 ± 2.4 | 16.3 ±0.7 |
| Dabcyl-Trp-Arg-Arg-Arg-Arg-Lys(MTX) b | 36.9 ± 3.0 | 7.4 ± 2.2 |
| MTX-Arg8 c | >100 | >100 |
| DauSuc d | 24.5 ± 7.1 | >100 |
| MTX c | 0.8 ± 0.3 | >100 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yousef, M.; Szabó, I.; Murányi, J.; Illien, F.; Soltész, D.; Bató, C.; Tóth, G.; Batta, G.; Nagy, P.; Sagan, S.; et al. Cell-Penetrating Dabcyl-Containing Tetraarginines with Backbone Aromatics as Uptake Enhancers. Pharmaceutics 2023, 15, 141. https://doi.org/10.3390/pharmaceutics15010141
Yousef M, Szabó I, Murányi J, Illien F, Soltész D, Bató C, Tóth G, Batta G, Nagy P, Sagan S, et al. Cell-Penetrating Dabcyl-Containing Tetraarginines with Backbone Aromatics as Uptake Enhancers. Pharmaceutics. 2023; 15(1):141. https://doi.org/10.3390/pharmaceutics15010141
Chicago/Turabian StyleYousef, Mo’ath, Ildikó Szabó, József Murányi, Françoise Illien, Dóra Soltész, Csaba Bató, Gabriella Tóth, Gyula Batta, Péter Nagy, Sandrine Sagan, and et al. 2023. "Cell-Penetrating Dabcyl-Containing Tetraarginines with Backbone Aromatics as Uptake Enhancers" Pharmaceutics 15, no. 1: 141. https://doi.org/10.3390/pharmaceutics15010141