
New Strategies for Stroke Therapy: Nanoencapsulated Neuroglobin



Abstract
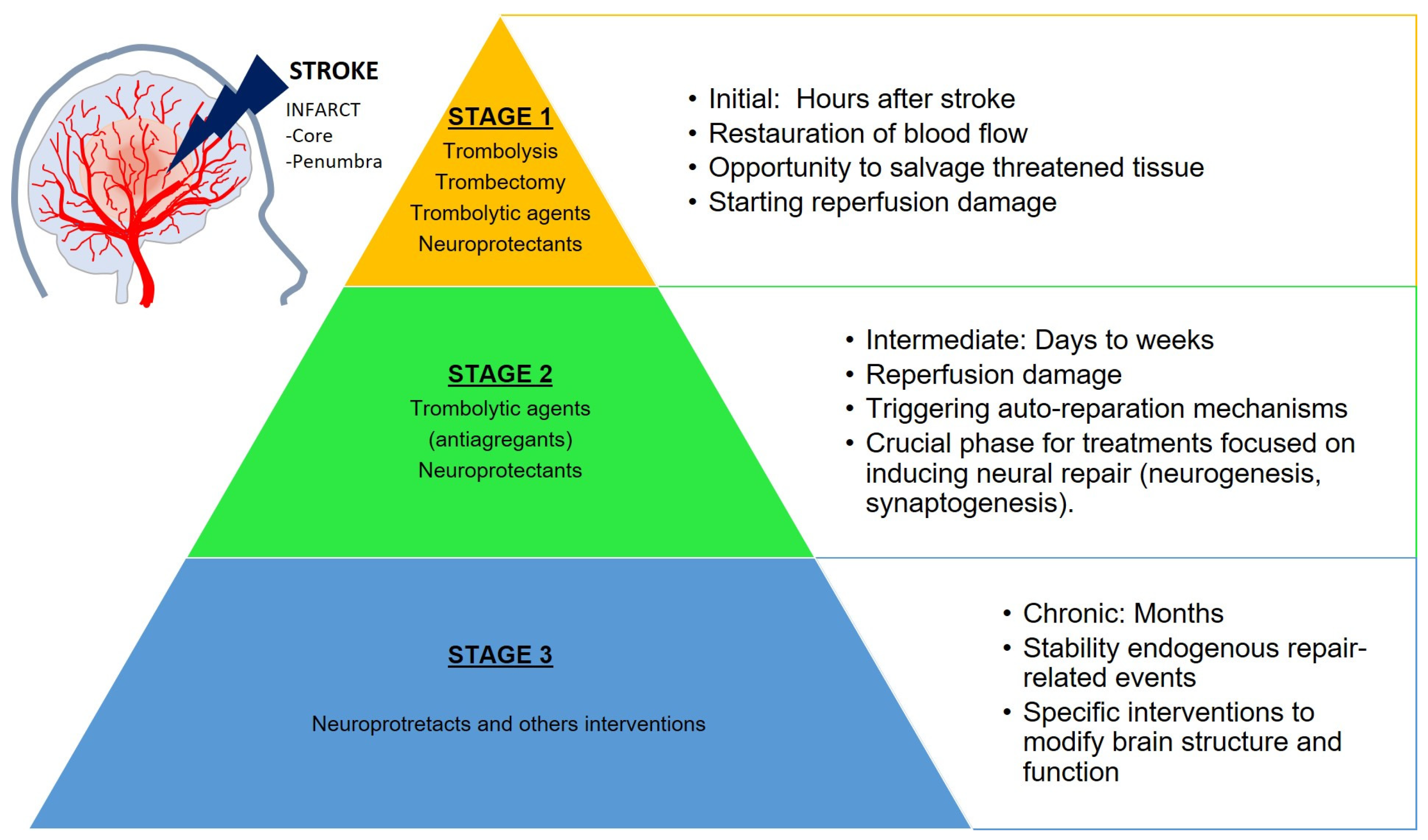
:1. Ischemic Stroke
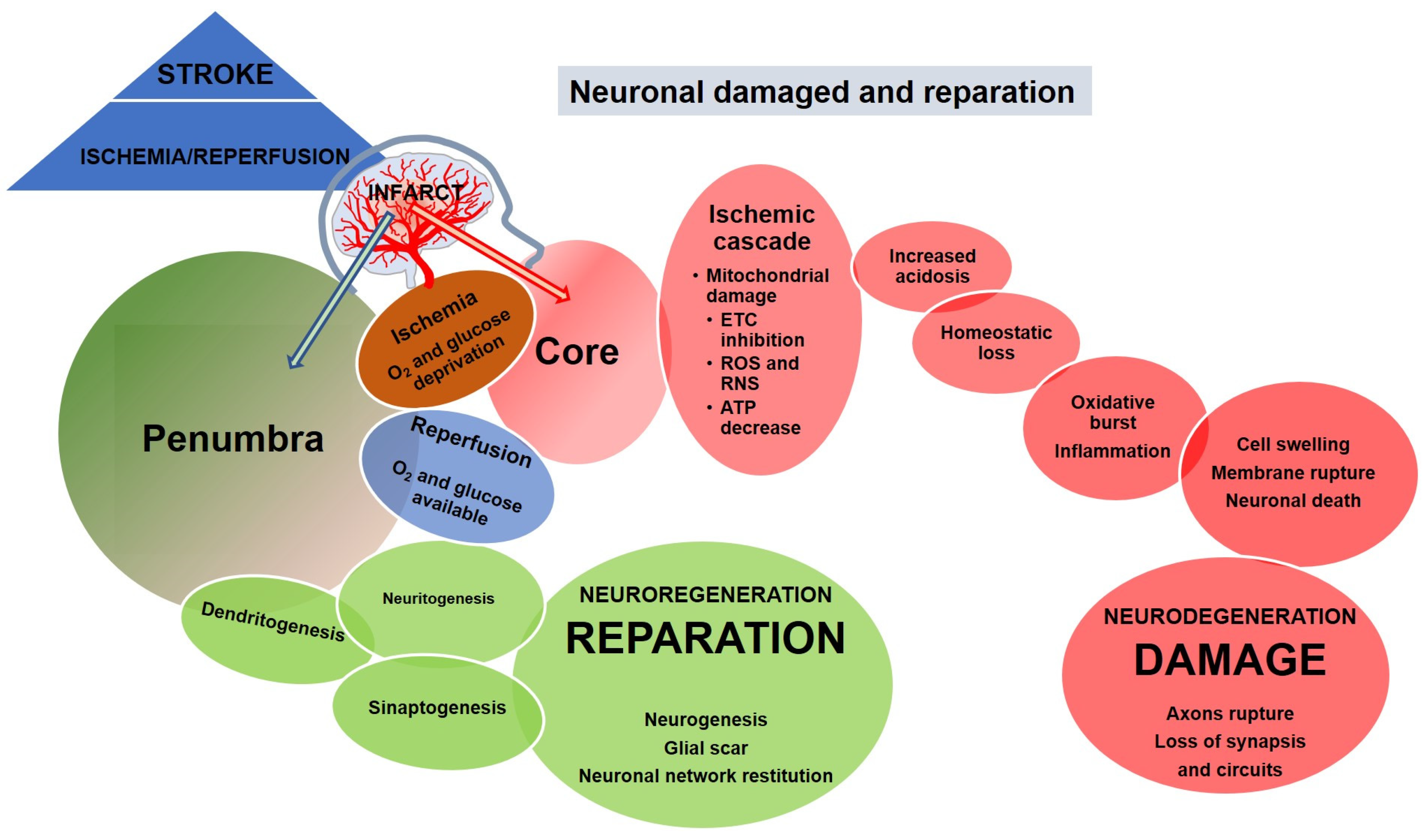
- The first occurs during the initial hours after the onset of stroke and represents an opportunity to salvage the tissue, eliminate clots (thrombolysis, thrombectomy and thrombolytic treatments), and restore the normal blood flow. The harmful consequences of stroke (ischemic cascade) begin as soon as the ischemia occurs and continue during the reperfusion onset. The prognosis of the stroke depends on the severity of the ischemia in this stage and determines the outcomes of the following stages.
- The second phase lasts from days to weeks following the stroke. The reperfusion damage may continue (including neuronal death and inflammation) but spontaneous neural auto-reparation mechanisms also occur. This stage is crucial for treatments focused on the induction of neural repair (neurogenesis and synaptogenesis).
- The third stage is a chronic period lasting from months to years. The brain is relatively stable regarding the endogenous repair-related events, but modifications in brain structure and function are still possible with specific neuroprotectant interventions.
2. Neuronal Damage and Regeneration in Ischemic Stroke
3. Current Treatments against Stroke
4. Ngb: A Neuroprotective Protein against Stroke
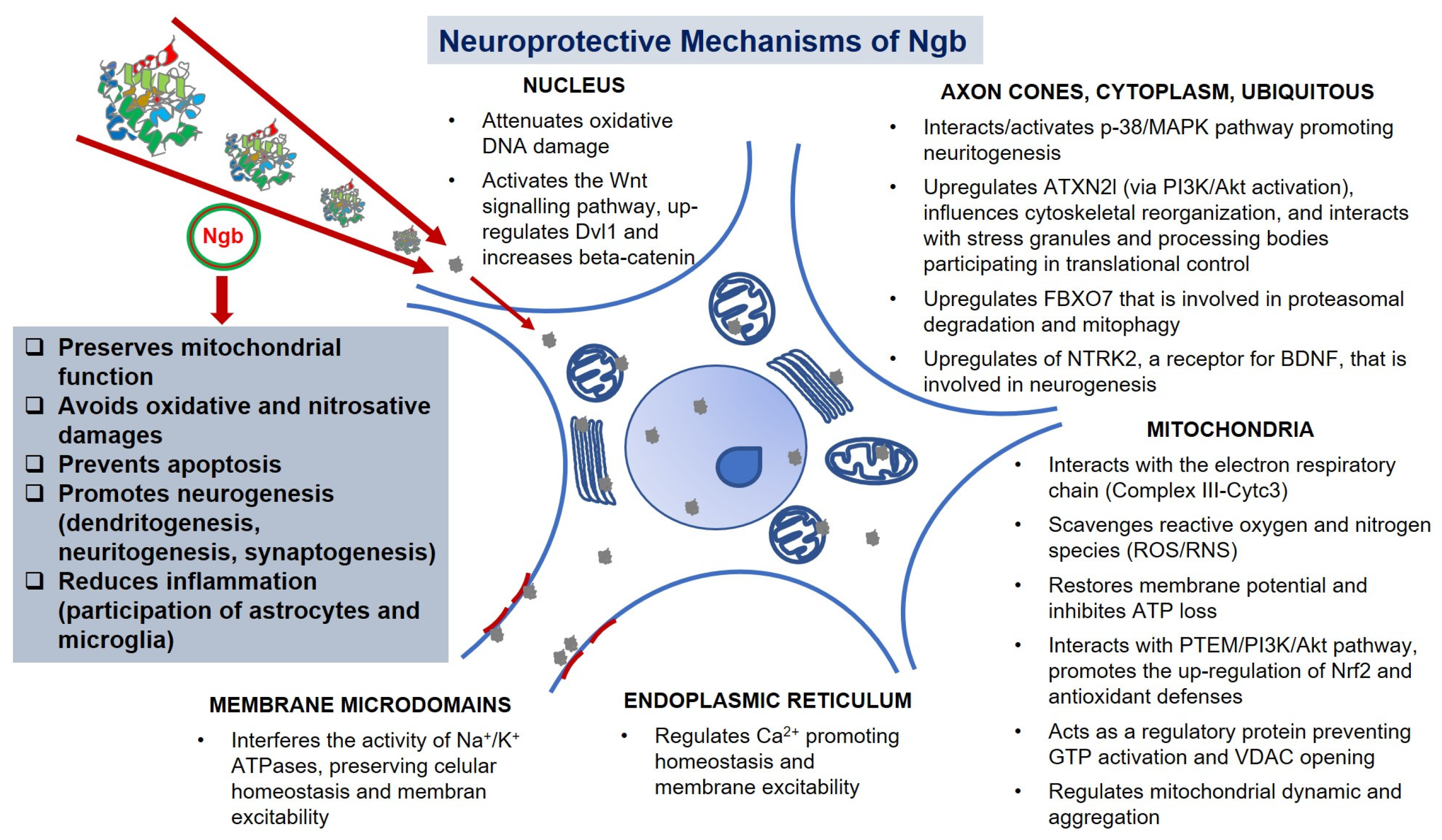
5. Unraveling the Neuroprotective Action Mechanisms of Ngb
6. Exogenous Ngb in Stroke Therapy: Nanotechnological Tools
6.1. BBB: A Problem to Overcome
6.2. CPPs
6.3. Polymeric NPs
6.4. New Challenges
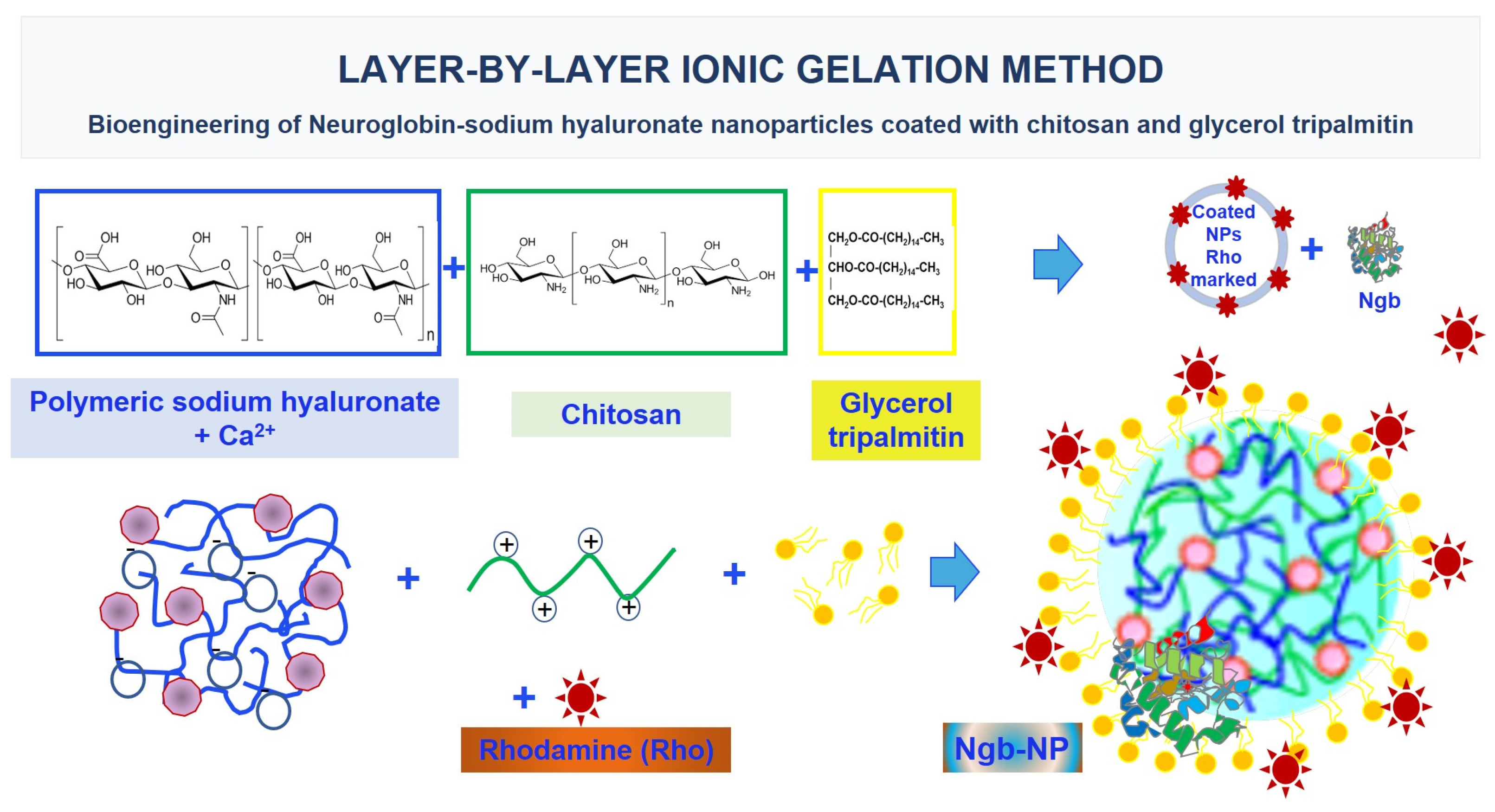
7. Delving into the Mechanisms of the Delivery of Ngb-NPs
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Alvarez-Sabín, J.; CONOCES Investigators Group; Quintana, M.; Masjuan, J.; Oliva-Moreno, J.; Mar, J.; Gonzalez-Rojas, N.; Becerra, V.; Torres, C.; Yebenes, M. Economic impact of patients admitted to stroke units in Spain. Eur. J. Health Econ. 2017, 18, 449–458. [Google Scholar] [CrossRef]
- Rajsic, S.; Gothe, H.; Borba, H.H.; Sroczynski, G.; Vujicic, J.; Toell, T.; Siebert, U. Economic burden of stroke: A systematic review on post-stroke care. Eur. J. Health Econ. 2019, 20, 107–134. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, J.; Wang, C.; Chen, X.; Zhao, X.; Jing, H.; Liu, H.; Li, Z.; Wang, L.; Shi, J. COVID-19 and ischemic stroke: Mechanisms of hypercoagulability (Review). Int. J. Mol. Med. 2021, 47, 21. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Asensio, F.J.; Hurtado, O.; Burguete, M.C.; Moro, M.A.; Salom, J.B.; Lizasoain, I.; Torregrosa, G.; Leza, J.C.; Alborch, E.; Castillo, J.; et al. Inhibition of iNOS activity by 1400W decreases glutamate release and ameliorates stroke outcome after experimental ischemia. Neurobiol. Dis. 2005, 18, 375–384. [Google Scholar] [CrossRef] [PubMed]
- Cramer, S.C. Treatments to Promote Neural Repair after Stroke. J. Stroke 2018, 20, 57–70. [Google Scholar] [CrossRef]
- Wu, Q.; Yan, R.; Sun, J. Probing the drug delivery strategies in ischemic stroke therapy. Drug Deliv. 2020, 27, 1644–1655. [Google Scholar] [CrossRef]
- Auriel, E.; Bornstein, N. Neuroprotection in acute ischemic stroke—current status. J. Cell. Mol. Med. 2010, 14, 2200–2202. [Google Scholar] [CrossRef]
- Di Domenico, F.; Casalena, G.; Jia, J.; Sultana, R.; Barone, E.; Cai, J.; Pierce, W.M.; Cini, C.; Mancuso, C.; Perluigi, M.; et al. Sex differences in brain proteomes of neuron-specific STAT3-null mice after cerebral ischemia/reperfusion. J. Neurochem. 2012, 121, 680–692. [Google Scholar] [CrossRef]
- Wu, M.-Y.; Yiang, G.-T.; Liao, W.-T.; Tsai, A.P.Y.; Cheng, Y.-L.; Cheng, P.-W.; Li, C.-Y.; Li, C.J. Current Mechanistic Concepts in Ischemia and Reperfusion Injury. Cell. Physiol. Biochem. 2018, 46, 1650–1667. [Google Scholar] [CrossRef] [PubMed]
- Carmichael, S.T.; Kathirvelu, B.; Schweppe, C.A.; Nie, E. Molecular, cellular and functional events in axonal sprouting after stroke. Exp. Neurol. 2017, 287, 384–394. [Google Scholar] [CrossRef]
- Koh, S.-H.; Park, H.-H. Neurogenesis in Stroke Recovery. Transl. Stroke Res. 2017, 8, 3–13. [Google Scholar] [CrossRef] [PubMed]
- Casares-Crespo, L.; Calatayud-Baselga, I.; García-Corzo, L.; Mira, H. On the Role of Basal Autophagy in Adult Neural Stem Cells and Neurogenesis. Front. Cell. Neurosci. 2018, 12, 339. [Google Scholar] [CrossRef] [PubMed]
- Xiong, X.X.; Pan, F.; Chen, R.Q.; Hu, D.X.; Qiu, X.Y.; Li, C.Y.; Xie, X.Q.; Tian, B.; Chen, X.Q. Neuroglobin boosts axon regeneration during ischemic reperfusion via p38 binding and activation depending on oxygen signal. Cell Death Dis. 2018, 9, 163. [Google Scholar] [CrossRef] [PubMed]
- Hermann, D.M.; Chopp, M. Promoting Neurological Recovery in the Post-Acute Stroke Phase: Benefits and Challenges. Eur. Neurol. 2014, 72, 317–325. [Google Scholar] [CrossRef] [PubMed]
- Beslow, L.A.; Smith, S.E.; Vossough, A.; Licht, D.J.; Kasner, S.E.; Favilla, C.; Halperin, A.R.; Gordon, D.M.; Jones, C.I.; Cucchiara, A.J.; et al. Hemorrhagic Transformation of Childhood Arterial Ischemic Stroke. Stroke 2011, 42, 941–946. [Google Scholar] [CrossRef] [PubMed]
- Terruso, V.; D’Amelio, M.; Di Benedetto, N.; Lupo, I.; Saia, V.; Famoso, G.; Mazzola, M.A.; Aridon, P.; Sarno, C.; Ragonese, P.; et al. Frequency and Determinants for Hemorrhagic Transformation of Cerebral Infarction. Neuroepidemiology 2009, 33, 261–265. [Google Scholar] [CrossRef] [PubMed]
- Van Den Berg, L.A.; Dijkgraaf, M.G.W.; Berkhemer, O.A.; Fransen, P.S.S.; Beumer, D.; Lingsma, H.F.; Majoie, C.B.L.M.; Dippel, D.W.J.; Van Der Lugt, A.; Van Oostenbrugge, R.J.; et al. Two-Year Outcome after Endovascular Treatment for Acute Ischemic Stroke. N. Engl. J. Med. 2017, 376, 1341–1349. [Google Scholar] [CrossRef] [PubMed]
- Bustamante, A.; García-Berrocoso, T.; Rodriguez, N.; Llombart, V.; Ribo, M.; Molina, C.; Montaner, J. Ischemic stroke outcome: A review of the influence of post-stroke complications within the different scenarios of stroke care. Eur. J. Intern. Med. 2016, 29, 9–21. [Google Scholar] [CrossRef]
- Rajkovic, O.; Potjewyd, G.; Pinteaux, E. Regenerative Medicine Therapies for Targeting Neuroinflammation after Stroke. Front. Neurol. 2018, 9, 734. [Google Scholar] [CrossRef]
- Van Der Worp, H.B.; Macleod, M.R.; Bath, P.; Demotes, J.; Durand-Zaleski, I.; Gebhardt, B.; Gluud, C.; Kollmar, R.; Krieger, D.W.; Lees, K.R.; et al. EuroHYP-1: European Multicenter, Randomized, Phase III Clinical Trial of Therapeutic Hypothermia plus Best Medical Treatment vs. Best Medical Treatment Alone for Acute Ischemic Stroke. Int. J. Stroke 2014, 9, 642–645. [Google Scholar] [CrossRef]
- A Donnan, G.; Davis, S.M. Breaking the 3 h barrier for treatment of acute ischaemic stroke. Lancet Neurol. 2008, 7, 981–982. [Google Scholar] [CrossRef]
- Broussalis, E.; Killer, M.; McCoy, M.; Harrer, A.; Trinka, E.; Kraus, J. Current therapies in ischemic stroke. Part A. Recent developments in acute stroke treatment and in stroke prevention. Drug Discov. Today 2012, 17, 296–309. [Google Scholar] [CrossRef] [PubMed]
- Ronaldson, P.T.; Davis, T.P. Transport Mechanisms at the Blood–Brain Barrier and in Cellular Compartments of the Neurovascular Unit: Focus on CNS Delivery of Small Molecule Drugs. Pharmaceutics 2022, 14, 1501. [Google Scholar] [CrossRef] [PubMed]
- Williams, E.I.; Betterton, R.D.; Davis, T.P.; Ronaldson, P.T. Transporter-Mediated Delivery of Small Molecule Drugs to the Brain: A Critical Mechanism That Can Advance Therapeutic Development for Ischemic Stroke. Pharmaceutics 2020, 12, 154. [Google Scholar] [CrossRef]
- Castillo, J.; Loza, M.I.; Mirelman, D.; Brea, J.; Blanco, M.; Sobrino, T.; Campos, F. A novel mechanism of neuroprotection: Blood glutamate grabber. J. Cereb. Blood Flow Metab. 2016, 36, 292–301. [Google Scholar] [CrossRef]
- da Silva-Candal, A.; Perez-Diaz, A.; Santamaría, M.; Correa-Paz, C.; Rodríguez-Yáñez, M.; Ardá, A.; Pérez-Mato, M.; Iglesias-Rey, R.; Brea, J.; Azuaje, J.; et al. Clinical validation of blood/brain glutamate grabbing in acute ischemic stroke. Ann. Neurol. 2018, 84, 260–273. [Google Scholar] [CrossRef] [PubMed]
- García-Bonilla, L.; Campos, M.; Giralt, D.; Salat, D.; Chacón, P.; Guillamon, M.M.H.; Rosell, A.; Montaner, J. Evidence for the efficacy of statins in animal stroke models: A meta-analysis. J. Neurochem. 2012, 122, 233–243. [Google Scholar] [CrossRef]
- Zhao, S.-P.; Zhao, W.; Xiao, Z.-J. The Benefits and Risks of Statin Therapy in Ischemic Stroke: A Review of the Literature. Neurol. India 2019, 67, 983–992. [Google Scholar] [CrossRef]
- Qian, Y.; Chopp, M.; Chen, J. Emerging role of microRNAs in ischemic stroke with comorbidities. Exp. Neurol. 2020, 331, 113382. [Google Scholar] [CrossRef]
- Jiang, X.; Andjelkovic, A.V.; Zhu, L.; Yang, T.; Bennett, M.V.L.; Chen, J.; Keep, R.F.; Shi, Y. Blood-brain barrier dysfunction and recovery after ischemic stroke. Prog. Neurobiol. 2018, 163–164, 144–171. [Google Scholar] [CrossRef]
- Dong, X.; Gao, J.; Su, Y.; Wang, Z. Nanomedicine for Ischemic Stroke. Int. J. Mol. Sci. 2020, 21, 7600. [Google Scholar] [CrossRef] [PubMed]
- Fukuta, T.; Oku, N.; Kogure, K. Application and Utility of Liposomal Neuroprotective Agents and Biomimetic Nanoparticles for the Treatment of Ischemic Stroke. Pharmaceutics 2022, 14, 361. [Google Scholar] [CrossRef] [PubMed]
- Teleanu, D.M.; Chircov, C.; Grumezescu, A.M.; Teleanu, R.I. Neuronanomedicine: An Up-to-Date Overview. Pharmaceutics 2019, 11, 101. [Google Scholar] [CrossRef] [PubMed]
- Durzyńska, J.; Przysiecka, Ł.; Nawrot, R.; Barylski, J.; Nowicki, G.; Warowicka, A.; Musidlak, O.; Goździcka-Józefiak, A. Viral and Other Cell-Penetrating Peptides as Vectors of Therapeutic Agents in Medicine. J. Pharmacol. Exp. Ther. 2015, 354, 32–42. [Google Scholar] [CrossRef]
- Kurrikoff, K.; Vunk, B.; Langel, Ü. Status update in the use of cell-penetrating peptides for the delivery of macromolecular therapeutics. Expert Opin. Biol. Ther. 2021, 21, 361–370. [Google Scholar] [CrossRef]
- Johnson, R.M.; Harrison, S.D.; Maclean, D. Therapeutic Applications of Cell-Penetrating Peptides. Methods Mol. Biol. 2011, 683, 535–551. [Google Scholar] [CrossRef]
- Wang, F.; Cerione, R.A.; Antonyak, M.A. Isolation and characterization of extracellular vesicles produced by cell lines. STAR Protoc. 2021, 2, 100295. [Google Scholar] [CrossRef]
- Alkaff, S.A.; Radhakrishnan, K.; Nedumaran, A.M.; Liao, P.; Czarny, B. Nanocarriers for Stroke Therapy: Advances and Obstacles in Translating Animal Studies. Int. J. Nanomed. 2020, 15, 445–464. [Google Scholar] [CrossRef]
- Fatima, S.; Quadri, S.N.; Parveen, S.; Beg, S.; Barkat, A.; Samim, M.; Abdin, M.; Ahmad, F.J. Nanomedicinal Strategies as Emerging Therapeutic Avenues to Treat and Manage Cerebral Ischemia. CNS Neurol. Disord. Drug Targets 2021, 20, 125–144. [Google Scholar] [CrossRef]
- Bernardo-Castro, S.; Albino, I.; Barrera-Sandoval, Á.M.; Tomatis, F.; Sousa, J.; Martins, E.; Simões, S.; Lino, M.; Ferreira, L.; Sargento-Freitas, J. Therapeutic Nanoparticles for the Different Phases of Ischemic Stroke. Life 2021, 11, 482. [Google Scholar] [CrossRef]
- He, W.; Zhang, Z.; Sha, X. Nanoparticles-mediated emerging approaches for effective treatment of ischemic stroke. Biomaterials 2021, 277, 121111. [Google Scholar] [CrossRef] [PubMed]
- Ruíz, M.A.; Clares, B.; Morales, M.E.; Gallardo, V. Vesicular Lipidic Systems, Liposomes, PLO, and Liposomes–PLO: Characterization by Electronic Transmission Microscopy. Drug Dev. Ind. Pharm. 2008, 34, 1269–1276. [Google Scholar] [CrossRef] [PubMed]
- Spuch, C.; Navarro, C. Liposomes for Targeted Delivery of Active Agents against Neurodegenerative Diseases (Alzheimer’s Disease and Parkinson’s Disease). J. Drug Deliv. 2011, 2011, 469679. [Google Scholar] [CrossRef] [PubMed]
- Teleanu, R.I.; Preda, M.D.; Niculescu, A.-G.; Vladâcenco, O.; Radu, C.I.; Grumezescu, A.M.; Teleanu, D.M. Current Strategies to Enhance Delivery of Drugs across the Blood–Brain Barrier. Pharmaceutics 2022, 14, 987. [Google Scholar] [CrossRef] [PubMed]
- Satapathy, M.; Yen, T.-L.; Jan, J.-S.; Tang, R.-D.; Wang, J.-Y.; Taliyan, R.; Yang, C.-H. Solid Lipid Nanoparticles (SLNs): An Advanced Drug Delivery System Targeting Brain through BBB. Pharmaceutics 2021, 13, 1183. [Google Scholar] [CrossRef]
- Gastaldi, L.; Battaglia, L.; Peira, E.; Chirio, D.; Muntoni, E.; Solazzi, I.; Gallarate, M.; Dosio, F. Solid lipid nanoparticles as vehicles of drugs to the brain: Current state of the art. Eur. J. Pharm. Biopharm. 2014, 87, 433–444. [Google Scholar] [CrossRef]
- Landowski, L.M.; Niego, B.; Sutherland, B.A.; Hagemeyer, C.E.; Howells, D.W. Applications of Nanotechnology in the Diagnosis and Therapy of Stroke. Semin. Thromb. Hemost. 2020, 46, 592–605. [Google Scholar] [CrossRef]
- Nair, R.; Kumar, A.C.; Priya, V.K.; Yadav, C.M.; Raju, P.Y. Formulation and evaluation of chitosan solid lipid nanoparticles of carbamazepine. Lipids Health Dis. 2012, 11, 72. [Google Scholar] [CrossRef]
- Panagiotou, S.; Esaha, S. Therapeutic benefits of nanoparticles in stroke. Front. Neurosci. 2015, 9, 182. [Google Scholar] [CrossRef]
- Karatas, H.; Aktas, Y.; Gursoy-Ozdemir, Y.; Bodur, E.; Yemisci, M.; Caban, S.; Vural, A.; Pinarbasli, O.; Capan, Y.; Fernandez-Megia, E.; et al. A Nanomedicine Transports a Peptide Caspase-3 Inhibitor across the Blood–Brain Barrier and Provides Neuroprotection. J. Neurosci. 2009, 29, 13761–13769. [Google Scholar] [CrossRef]
- Reddy, M.K.; Labhasetwar, V. Nanoparticle-mediated delivery of superoxide dismutase to the brain: An effective strategy to reduce ischemia-reperfusion injury. FASEB J. 2009, 23, 1384–1395. [Google Scholar] [CrossRef] [PubMed]
- Ortega, E.; Blanco, S.; Ruiz, A.; Peinado, M.A.; Peralta, S.; Morales, M.E. Lipid nanoparticles for the transport of drugs like dopamine through the blood-brain barrier. J. Nanopart. Res. 2021, 23, 106. [Google Scholar] [CrossRef]
- Liu, X.; Ye, M.; An, C.; Pan, L.; Ji, L. The effect of cationic albumin-conjugated PEGylated tanshinone IIA nanoparticles on neuronal signal pathways and neuroprotection in cerebral ischemia. Biomaterials 2013, 34, 6893–6905. [Google Scholar] [CrossRef] [PubMed]
- Blanco, S.; Peralta, S.; Morales, M.E.; Martínez-Lara, E.; Pedrajas, J.R.; Castán, H.; Peinado, M.; Ruiz, M.A. Hyaluronate Nanoparticles as a Delivery System to Carry Neuroglobin to the Brain after Stroke. Pharmaceutics 2020, 12, 40. [Google Scholar] [CrossRef]
- Peinado, M.; Ovelleiro, D.; del Moral, M.L.; Hernández, R.; Martínez-Lara, E.; Siles, E.; Pedrajas, J.R.; García-Martín, M.L.; Caro, C.; Peralta, S.; et al. Biological Implications of a Stroke Therapy Based in Neuroglobin Hyaluronate Nanoparticles. Neuroprotective Role and Molecular Bases. Int. J. Mol. Sci. 2021, 23, 247. [Google Scholar] [CrossRef]
- Peralta, S.; Blanco, S.; Hernández, R.; Castán, H.; Siles, E.; Martínez-Lara, E.; Morales, M.E.; Peinado, M.; Ruiz, M.A. Synthesis and characterization of different sodium hyaluronate nanoparticles to transport large neurotherapheutic molecules through blood brain barrier after stroke. Eur. Polym. J. 2019, 112, 433–441. [Google Scholar] [CrossRef]
- Burmester, T.; Weich, B.; Reinhardt, S.; Hankeln, T. A vertebrate globin expressed in the brain. Nature 2000, 407, 520–523. [Google Scholar] [CrossRef]
- Exertier, C.; Montemiglio, L.C.; Freda, I.; Gugole, E.; Parisi, G.; Savino, C.; Vallone, B. Neuroglobin, clues to function and mechanism. Mol. Asp. Med. 2022, 84, 101055. [Google Scholar] [CrossRef]
- Keppner, A.; Maric, D.; Correia, M.; Koay, T.W.; Orlando, I.M.; Vinogradov, S.N.; Hoogewijs, D. Lessons from the post-genomic era: Globin diversity beyond oxygen binding and transport. Redox Biol. 2020, 37, 101687. [Google Scholar] [CrossRef]
- Gorabi, A.M.; Aslani, S.; Barreto, G.E.; Báez-Jurado, E.; Kiaie, N.; Jamialahmadi, T.; Sahebkar, A. The potential of mitochondrial modulation by neuroglobin in treatment of neurological disorders. Free Radic. Biol. Med. 2020, 162, 471–477. [Google Scholar] [CrossRef]
- Fordel, E.; Geuens, E.; Dewilde, S.; De Coen, W.; Moens, L. Hypoxia/Ischemia and the Regulation of Neuroglobin and Cytoglobin Expression. IUBMB Life 2004, 56, 681–687. [Google Scholar] [CrossRef] [PubMed]
- Liu, N.; Yu, Z.; Gao, X.; Song, Y.; Yuan, J.; Xun, Y.; Wang, T.; Yan, F.; Yuan, S.; Zhang, J.; et al. Establishment of Cell-Based Neuroglobin Promoter Reporter Assay for Neuroprotective Compounds Screening. CNS Neurol. Disord. Drug Targets 2016, 15, 629–639. [Google Scholar] [CrossRef] [PubMed]
- Ren, C.; Wang, P.; Wang, B.; Li, N.; Li, W.; Zhang, C.; Jin, K.; Ji, X. Limb remote ischemic per-conditioning in combination with post-conditioning reduces brain damage and promotes neuroglobin expression in the rat brain after ischemic stroke. Restor. Neurol. Neurosci. 2015, 33, 369–379. [Google Scholar] [CrossRef] [PubMed]
- Schmidt-Kastner, R.; Haberkamp, M.; Schmitz, C.; Hankeln, T.; Burmester, T. Neuroglobin mRNA expression after transient global brain ischemia and prolonged hypoxia in cell culture. Brain Res. 2006, 1103, 173–180. [Google Scholar] [CrossRef] [PubMed]
- Shang, A.; Zhou, D.; Wang, L.; Gao, Y.; Fan, M.; Wang, X.; Zhou, R.; Zhang, C. Increased neuroglobin levels in the cerebral cortex and serum after ischemia–reperfusion insults. Brain Res. 2006, 1078, 219–226. [Google Scholar] [CrossRef]
- Song, X.; Xu, R.; Xie, F.; Zhu, H.; Zhu, J.; Wang, X. Hemin offers neuroprotection through inducing exogenous neuroglobin in focal cerebral hypoxic-ischemia in rats. Int. J. Clin. Exp. Pathol. 2014, 7, 2163. [Google Scholar]
- Pandya, R.S.; Mao, L.; Zhou, H.; Zhou, S.; Zeng, J.; Popp, A.J.; Wang, X. Central nervous system agents for ischemic stroke: Neuroprotection mechanisms. Central Nerv. Syst. Agents Med. Chem. 2011, 11, 81–97. [Google Scholar] [CrossRef]
- Sun, Y.; Jin, K.; Mao, X.O.; Zhu, Y.; Greenberg, D.A. Neuroglobin is up-regulated by and protects neurons from hypoxic-ischemic injury. Proc. Natl. Acad. Sci. USA 2001, 98, 15306–15311. [Google Scholar] [CrossRef]
- Jin, K.; Mao, X.; Xie, L.; Greenberg, D.A. Neuroglobin Expression in Human Arteriovenous Malformation and Intracerebral Hemorrhage. Acta Neurochir. Suppl. 2011, 111, 315–319. [Google Scholar] [CrossRef]
- Jin, K.; Mao, Y.; Mao, X.; Xie, L.; Greenberg, D.A. Neuroglobin Expression in Ischemic Stroke. Stroke 2010, 41, 557–559. [Google Scholar] [CrossRef]
- Fiocchetti, M.; Cipolletti, M.; Brandi, V.; Polticelli, F.; Ascenzi, P. Neuroglobin and friends. J. Mol. Recognit. 2017, 30, e2654. [Google Scholar] [CrossRef] [PubMed]
- Raida, Z.; Hundahl, C.A.; Nyengaard, J.R.; Hay-Schmidt, A. Neuroglobin Over Expressing Mice: Expression Pattern and Effect on Brain Ischemic Infarct Size. PLoS ONE 2013, 8, e76565. [Google Scholar] [CrossRef] [PubMed]
- Hundahl, C.; Kelsen, J.; Kjær, K.; Rønn, L.C.B.; Weber, R.E.; Geuens, E.; Hay-Schmidt, A.; Nyengaard, J.R. Does neuroglobin protect neurons from ischemic insult? A quantitative investigation of neuroglobin expression following transient MCAo in spontaneously hypertensive rats. Brain Res. 2006, 1085, 19–27. [Google Scholar] [CrossRef] [PubMed]
- Raida, Z.; Hundahl, C.A.; Kelsen, J.; Nyengaard, J.R.; Hay-Schmidt, A. Reduced infarct size in neuroglobin-null mice after experimental stroke in vivo. Exp. Transl. Stroke Med. 2012, 4, 15. [Google Scholar] [CrossRef]
- Shang, A.; Liu, K.; Wang, H.; Wang, J.; Hang, X.; Yang, Y.; Wang, Z.; Zhang, C.; Zhou, D. Neuroprotective effects of neuroglobin after mechanical injury. Neurol. Sci. 2011, 33, 551–558. [Google Scholar] [CrossRef]
- Khan, A.A.; Wang, Y.; Sun, Y.; Mao, X.O.; Xie, L.; Miles, E.; Graboski, J.; Chen, S.; Ellerby, L.M.; Jin, K.; et al. Neuroglobin-overexpressing transgenic mice are resistant to cerebral and myocardial ischemia. Proc. Natl. Acad. Sci. USA 2006, 103, 17944–17948. [Google Scholar] [CrossRef]
- Wang, J.-Y.; Shen, J.; Gao, Q.; Ye, Z.-G.; Yang, S.-Y.; Liang, H.-W.; Bruce, I.C.; Luo, B.-Y.; Xia, Q. Ischemic Postconditioning Protects Against Global Cerebral Ischemia/Reperfusion-Induced Injury in Rats. Stroke 2008, 39, 983–990. [Google Scholar] [CrossRef]
- Raychaudhuri, S.; Skommer, J.; Henty, K.; Birch, N.; Brittain, T. Neuroglobin protects nerve cells from apoptosis by inhibiting the intrinsic pathway of cell death. Apoptosis 2010, 15, 401–411. [Google Scholar] [CrossRef]
- Sun, Y.; Jin, K.; Peel, A.; Mao, X.O.; Xie, L.; Greenberg, D.A. Neuroglobin protects the brain from experimental stroke in vivo. Proc. Natl. Acad. Sci. USA 2003, 100, 3497–3500. [Google Scholar] [CrossRef]
- Greenberg, D.A.; Jin, K.; Khan, A.A. Neuroglobin: An endogenous neuroprotectant. Curr. Opin. Pharmacol. 2008, 8, 20–24. [Google Scholar] [CrossRef]
- Yu, Z.; Fan, X.; Lo, E.H.; Wang, X. Neuroprotective roles and mechanisms of neuroglobin. Neurol. Res. 2009, 31, 122–127. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Sun, Y.; Jin, K.; Greenberg, D.A. Hemin induces neuroglobin expression in neural cells. Blood 2002, 100, 2494–2498. [Google Scholar] [CrossRef] [PubMed]
- Avila-Rodríguez, M.; Garcia-Segura, L.M.; Lanussa, O.A.H.; Baez, E.; Gonzalez, J.; Barreto, G.E. Tibolone protects astrocytic cells from glucose deprivation through a mechanism involving estrogen receptor beta and the upregulation of neuroglobin expression. Mol. Cell. Endocrinol. 2016, 433, 35–46. [Google Scholar] [CrossRef] [PubMed]
- Lanussa, O.A.H.; Avila-Rodríguez, M.; Baez-Jurado, E.; Zamudio-Rodriguez, J.A.; Echeverria, V.; Garcia-Segura, L.M.; Barreto, G.E. Tibolone Reduces Oxidative Damage and Inflammation in Microglia Stimulated with Palmitic Acid through Mechanisms Involving Estrogen Receptor Beta. Mol. Neurobiol. 2018, 55, 5462–5477. [Google Scholar] [CrossRef] [PubMed]
- Barreto, G.; McGovern, A.; Garcia-Segura, L. Role of Neuroglobin in the Neuroprotective Actions of Estradiol and Estrogenic Compounds. Cells 2021, 10, 1907. [Google Scholar] [CrossRef]
- Acaz-Fonseca, E.; Castelló-Ruiz, M.; Burguete, M.C.; Aliena-Valero, A.; Salom, J.B.; Torregrosa, G.; García-Segura, L.M. Insight into the molecular sex dimorphism of ischaemic stroke in rat cerebral cortex: Focus on neuroglobin, sex steroids and autophagy. Eur. J. Neurosci. 2020, 52, 2756–2770. [Google Scholar] [CrossRef]
- Baez, E.; Echeverria, V.; Cabezas, R.; Ávila-Rodriguez, M.; Garcia-Segura, L.M.; Barreto, G.E. Protection by Neuroglobin Expression in Brain Pathologies. Front. Neurol. 2016, 7, 146. [Google Scholar] [CrossRef]
- Kakar, S.; Hoffman, F.G.; Storz, J.F.; Fabian, M.; Hargrove, M.S. Structure and reactivity of hexacoordinate hemoglobins. Biophys. Chem. 2010, 152, 1–14. [Google Scholar] [CrossRef]
- Liu, J.; Yu, Z.; Guo, S.; Lee, S.-R.; Xing, C.; Zhang, C.; Gao, Y.; Nicholls, D.G.; Lo, E.H.; Wang, X. Effects of neuroglobin overexpression on mitochondrial function and oxidative stress following hypoxia/reoxygenation in cultured neurons. J. Neurosci. Res. 2009, 87, 164–170. [Google Scholar] [CrossRef]
- Tejero, J.; Sparacino-Watkins, C.E.; Ragireddy, V.; Frizzell, S.; Gladwin, M.T. Exploring the Mechanisms of the Reductase Activity of Neuroglobin by Site-Directed Mutagenesis of the Heme Distal Pocket. Biochemistry 2015, 54, 722–733. [Google Scholar] [CrossRef]
- Brunori, M.; Giuffrè, A.; Nienhaus, K.; Nienhaus, G.U.; Scandurra, F.M.; Vallone, B. Neuroglobin, nitric oxide, and oxygen: Functional pathways and conformational changes. Proc. Natl. Acad. Sci. USA 2005, 102, 8483–8488. [Google Scholar] [CrossRef] [PubMed]
- Van Doorslaer, S.; Dewilde, S.; Kiger, L.; Nistor, S.V.; Goovaerts, E.; Marden, M.C.; Moens, L. Nitric oxide binding properties of neuroglobin: A characterization by EPR and flash photolysis. J. Biol. Chem. 2003, 278, 4919–4925. [Google Scholar] [CrossRef] [PubMed]
- Hundahl, C.A.; Allen, G.C.; Nyengaard, J.R.; Dewilde, S.; Carter, B.D.; Kelsen, J.; Hay-Schmidt, A. Neuroglobin in the Rat Brain: Localization. Neuroendocrinology 2008, 88, 173–182. [Google Scholar] [CrossRef] [PubMed]
- Mammen, P.P.; Shelton, J.M.; Goetsch, S.C.; Williams, S.C.; Richardson, J.A.; Garry, M.G.; Garry, D.J. Neuroglobin, A Novel Member of the Globin Family, Is Expressed in Focal Regions of the Brain. J. Histochem. Cytochem. 2002, 50, 1591–1598. [Google Scholar] [CrossRef]
- Eliasson, M.J.L.; Huang, Z.; Ferrante, R.J.; Sasamata, M.; Molliver, M.E.; Snyder, S.H.; Moskowitz, M.A. Neuronal Nitric Oxide Synthase Activation and Peroxynitrite Formation in Ischemic Stroke Linked to Neural Damage. J. Neurosci. 1999, 19, 5910–5918. [Google Scholar] [CrossRef]
- Herold, S.; Fago, A. Reactions of peroxynitrite with globin proteins and their possible physiological role. Comp. Biochem. Physiol. Part A Mol. Integr. Physiol. 2005, 142, 124–129. [Google Scholar] [CrossRef]
- Nicolis, S.; Monzani, E.; Ciaccio, C.; Ascenzi, P.; Moens, L.; Casella, L. Reactivity and endogenous modification by nitrite and hydrogen peroxide: Does human neuroglobin act only as a scavenger? Biochem. J. 2007, 407, 89–99. [Google Scholar] [CrossRef]
- Yu, Z.; Zhang, Y.; Liu, N.; Yuan, J.; Lin, L.; Zhuge, Q.; Xiao, J.; Wang, X. Roles of Neuroglobin Binding to Mitochondrial Complex III Subunit Cytochrome c1 in Oxygen-Glucose Deprivation-Induced Neurotoxicity in Primary Neurons. Mol. Neurobiol. 2016, 53, 3249–3257. [Google Scholar] [CrossRef]
- Chen, L.-M.; Xiong, Y.-S.; Kong, F.-L.; Qu, M.; Wang, Q.; Chen, X.-Q.; Wang, J.-Z.; Zhu, L.-Q. Neuroglobin attenuates Alzheimer-like tau hyperphosphorylation by activating Akt signaling. J. Neurochem. 2012, 120, 157–164. [Google Scholar] [CrossRef]
- Lee, H.M.; Greeley, G.H., Jr.; Englander, E.W. Transgenic overexpression of neuroglobin attenuates formation of smoke-inhalation-induced oxidative DNA damage, in vivo, in the mouse brain. Free Radic. Biol. Med. 2011, 51, 2281–2287. [Google Scholar] [CrossRef]
- Antao, S.T.; Duong, T.H.; Aran, R.; Witting, P.K. Neuroglobin Overexpression in Cultured Human Neuronal Cells Protects Against Hydrogen Peroxide Insult via Activating Phosphoinositide-3 Kinase and Opening the Mitochondrial KATP Channel. Antioxid. Redox Signal. 2010, 13, 769–781. [Google Scholar] [CrossRef] [PubMed]
- Cai, B.; Li, W.; Mao, X.; Winters, A.; Ryou, M.-G.; Liu, R.; Greenberg, D.A.; Wang, N.; Jin, K.; Yang, S.-H. Neuroglobin Overexpression Inhibits AMPK Signaling and Promotes Cell Anabolism. Mol. Neurobiol. 2016, 53, 1254–1265. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Hao, X.; Chang, J.; Geng, Z.; Wang, Z. Mn-TAT PTD-Ngb attenuates oxidative injury by an enhanced ROS scavenging ability and the regulation of redox signaling pathway. Sci. Rep. 2019, 9, 20103. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Ji, X.; Zhang, S.; Ren, H.; Wang, M.; Guo, C.; Li, Y. Hemin-mediated neuroglobin induction exerts neuroprotection following ischemic brain injury through PI3K/Akt signaling. Mol. Med. Rep. 2013, 8, 681–685. [Google Scholar] [CrossRef] [PubMed]
- Zhang, B.; Liu, Y.; Li, Y.; Zhe, X.; Zhang, S.; Zhang, L. Neuroglobin promotes the proliferation and suppresses the apoptosis of glioma cells by activating the PI3K/AKT pathway. Mol. Med. Rep. 2018, 17, 2757–2763. [Google Scholar] [CrossRef] [PubMed]
- Yu, Z.; Liu, N.; Li, Y.; Xu, J.; Wang, X. Neuroglobin overexpression inhibits oxygen–glucose deprivation-induced mitochondrial permeability transition pore opening in primary cultured mouse cortical neurons. Neurobiol. Dis. 2013, 56, 95–103. [Google Scholar] [CrossRef] [PubMed]
- Fago, A.; Mathews, A.J.; Moens, L.; Dewilde, S.; Brittain, T. The reaction of neuroglobin with potential redox protein partners cytochrome b5 and cytochrome c. FEBS Lett. 2006, 580, 4884–4888. [Google Scholar] [CrossRef]
- Duong, T.T.H.; Witting, P.K.; Antao, S.T.; Parry, S.N.; Kennerson, M.; Lai, B.; Vogt, S.; Lay, P.A.; Harris, H.H. Multiple protective activities of neuroglobin in cultured neuronal cells exposed to hypoxia re-oxygenation injury. J. Neurochem. 2009, 108, 1143–1154. [Google Scholar] [CrossRef]
- Goldsmith, K.C.; Gross, M.; Peirce, S.; Luyindula, D.; Liu, X.; Vu, A.; Sliozberg, M.; Guo, R.; Zhao, H.; Reynolds, C.P.; et al. Mitochondrial Bcl-2 Family Dynamics Define Therapy Response and Resistance in Neuroblastoma. Cancer Res. 2012, 72, 2565–2577. [Google Scholar] [CrossRef]
- Boldogh, I.R.; Pon, L.A. Interactions of mitochondria with the actin cytoskeleton. Biochim. Biophys. Acta 2006, 1763, 450–462. [Google Scholar] [CrossRef]
- Yu, Z.; Cheng, C.; Liu, Y.; Liu, N.; Lo, E.H.; Wang, X. Neuroglobin promotes neurogenesis through Wnt signaling pathway. Cell Death Dis. 2018, 9, 945. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Liu, L.; Zhan, L.; Liang, D.; Li, L.; Liu, D.; Sun, W.; Xu, E. Neuroglobin mediates neuroprotection of hypoxic postconditioning against transient global cerebral ischemia in rats through preserving the activity of Na+/K+ ATPases. Cell Death Dis. 2018, 9, 635. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Liu, Q.R.; Xiong, X.X.; Liu, J.M.; Lai, X.J.; Cheng, C.; Pan, F.; Chen, Y.; Bin Yu, S.; Yu, A.C.H.; et al. Neuroglobin Promotes Neurite Outgrowth via Differential Binding to PTEN and Akt. Mol. Neurobiol. 2014, 49, 149–162. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Liu, Y.; Zhang, L.; Zhu, P.; Zhu, H.; Yang, Y.; Guan, P. Long-term neuroglobin expression of human astrocytes following brain trauma. Neurosci. Lett. 2015, 606, 194–199. [Google Scholar] [CrossRef]
- Hernández, R.; Blanco, S.; Peragón, J.; Pedrosa, J.; Peinado, M. Hypobaric Hypoxia and Reoxygenation Induce Proteomic Profile Changes in the Rat Brain Cortex. Neuromol. Med. 2013, 15, 82–94. [Google Scholar] [CrossRef]
- Ovelleiro, D.; Blanco, S.; Hernández, R.; Peinado, M. Comparative proteomic study of early hypoxic response in the cerebral cortex of rats submitted to two different hypoxic models. Proteom. Clin. Appl. 2017, 11, 1700058. [Google Scholar] [CrossRef]
- Peinado, M.; Hernández, R.; Peragón, J.; Ovelleiro, D.; Pedrosa, J.; Blanco, S. Proteomic characterization of nitrated cell targets after hypobaric hypoxia and reoxygenation in rat brain. J. Proteom. 2014, 109, 309–321. [Google Scholar] [CrossRef]
- Haines, B.A.; Davis, D.A.; Zykovich, A.; Peng, B.; Rao, R.; Mooney, S.D.; Jin, K.; Greenberg, D.A. Comparative protein interactomics of neuroglobin and myoglobin. J. Neurochem. 2012, 123, 192–198. [Google Scholar] [CrossRef]
- Lin, L.; Li, X.; Pan, C.; Lin, W.; Shao, R.; Liu, Y.; Zhang, J.; Luo, Y.; Qian, K.; Shi, M.; et al. ATXN2L upregulated by epidermal growth factor promotes gastric cancer cell invasiveness and oxaliplatin resistance. Cell Death Dis. 2019, 10, 173. [Google Scholar] [CrossRef]
- Peroni, D.; Negro, A.; Bähr, M.; Dietz, G.P. Intracellular delivery of Neuroglobin using HIV-1 TAT protein transduction domain fails to protect against oxygen and glucose deprivation. Neurosci. Lett. 2007, 421, 110–114. [Google Scholar] [CrossRef]
- Zhou, G.-Y.; Zhou, S.-N.; Lou, Z.-Y.; Zhu, C.-S.; Zheng, X.-P.; Hu, X.-Q. Translocation and neuroprotective properties of transactivator-of-transcription protein-transduction domain–neuroglobin fusion protein in primary cultured cortical neurons. Biotechnol. Appl. Biochem. 2008, 49, 25–33. [Google Scholar] [CrossRef] [PubMed]
- Belayev, L.; Busto, R.; Zhao, W.; Ginsberg, M.D. Quantitative evaluation of blood-brain barrier permeability following middle cerebral artery occlusion in rats. Brain Res. 1996, 739, 88–96. [Google Scholar] [CrossRef]
- Rosenberg, G.A.; Estrada, E.Y.; Dencoff, J.E. Matrix Metalloproteinases and TIMPs Are Associated with Blood-Brain Barrier Opening After Reperfusion in Rat Brain. Stroke 1998, 29, 2189–2195. [Google Scholar] [CrossRef] [PubMed]
- McColl, B.W.; Rothwell, N.J.; Allan, S. Systemic Inflammation Alters the Kinetics of Cerebrovascular Tight Junction Disruption after Experimental Stroke in Mice. J. Neurosci. 2008, 28, 9451–9462. [Google Scholar] [CrossRef]
- Cai, B.; Lin, Y.; Xue, X.-H.; Fang, L.; Wang, N.; Wu, Z.-Y. TAT-mediated delivery of neuroglobin protects against focal cerebral ischemia in mice. Exp. Neurol. 2011, 227, 224–231. [Google Scholar] [CrossRef]
- Fiocchetti, M.; De Marinis, E.; Ascenzi, P.; Marino, M. Neuroglobin and neuronal cell survival. Biochim. Biophys. Acta Proteins Proteom. 2013, 1834, 1744–1749. [Google Scholar] [CrossRef]
- Yu, Z.; Liu, N.; Liu, J.; Yang, K.; Wang, X. Neuroglobin, a Novel Target for Endogenous Neuroprotection against Stroke and Neurodegenerative Disorders. Int. J. Mol. Sci. 2012, 13, 6995–7014. [Google Scholar] [CrossRef]
- Dietz, G.P.H.; Bähr, M. Delivery of bioactive molecules into the cell: The Trojan horse approach. Mol. Cell. Neurosci. 2004, 27, 85–131. [Google Scholar] [CrossRef]
- Luyckx, E.; Van Acker, Z.P.; Ponsaerts, P.; Dewilde, S. Neuroglobin Expression Models as a Tool to Study Its Function. Oxidative Med. Cell. Longev. 2019, 2019, 5728129. [Google Scholar] [CrossRef]
- Hanson, L.R.; Frey, W.H. Intranasal delivery bypasses the blood-brain barrier to target therapeutic agents to the central nervous system and treat neurodegenerative disease. BMC Neurosci. 2008, 9, S5. [Google Scholar] [CrossRef]
- Monge-Fuentes, V.; Mayer, A.B.; Lima, M.R.; Geraldes, L.R.; Zanotto, L.N.; Moreira, K.G.; Martins, O.P.; Piva, H.L.; Felipe, M.S.S.; Amaral, A.C.; et al. Dopamine-loaded nanoparticle systems circumvent the blood–brain barrier restoring motor function in mouse model for Parkinson’s Disease. Sci. Rep. 2021, 11, 15185. [Google Scholar] [CrossRef] [PubMed]
- Fortuni, B.; Inose, T.; Ricci, M.; Fujita, Y.; Van Zundert, I.; Masuhara, A.; Fron, E.; Mizuno, H.; Latterini, L.; Rocha, S.; et al. Polymeric Engineering of Nanoparticles for Highly Efficient Multifunctional Drug Delivery Systems. Sci. Rep. 2019, 9, 2666. [Google Scholar] [CrossRef] [PubMed]
- Ahlawat, J.; Henriquez, G.; Narayan, M. Enhancing the Delivery of Chemotherapeutics: Role of Biodegradable Polymeric Nanoparticles. Molecules 2018, 23, 2157. [Google Scholar] [CrossRef] [PubMed]
- Cortés, H.; Alcalá-Alcalá, S.; Caballero-Florán, I.H.; Bernal-Chávez, S.A.; Ávalos-Fuentes, A.; González-Torres, M.; Carmen, M.G.-D.; Figueroa-González, G.; Reyes-Hernández, O.D.; Floran, B.; et al. A Reevaluation of Chitosan-Decorated Nanoparticles to Cross the Blood-Brain Barrier. Membranes 2020, 10, 212. [Google Scholar] [CrossRef] [PubMed]
- Jensen, G.; Holloway, J.L.; Stabenfeldt, S.E. Hyaluronic Acid Biomaterials for Central Nervous System Regenerative Medicine. Cells 2020, 9, 2113. [Google Scholar] [CrossRef] [PubMed]
- Sawada, R.; Nakano-Doi, A.; Matsuyama, T.; Nakagomi, N.; Nakagomi, T.T. CD44 expression in stem cells and niche microglia/macrophages following ischemic stroke. Stem Cell Investig. 2020, 7, 4. [Google Scholar] [CrossRef]
- Donahue, N.D.; Acar, H.; Wilhelm, S. Concepts of nanoparticle cellular uptake, intracellular trafficking, and kinetics in nanomedicine. Adv. Drug Deliv. Rev. 2019, 143, 68–96. [Google Scholar] [CrossRef] [PubMed]
- Varkouhi, A.K.; Scholte, M.; Storm, G.; Haisma, H.J. Endosomal escape pathways for delivery of biologicals. J. Control. Release 2011, 151, 220–228. [Google Scholar] [CrossRef]
- Pei, D.; Buyanova, M. Overcoming Endosomal Entrapment in Drug Delivery. Bioconj. Chem. 2019, 30, 273–283. [Google Scholar] [CrossRef]
- Correa-Paz, C.; da Silva-Candal, A.; Polo, E.; Parcq, J.; Vivien, D.; Maysinger, D.; Pelaz, B.; Campos, F. New Approaches in Nanomedicine for Ischemic Stroke. Pharmaceutics 2021, 13, 757. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
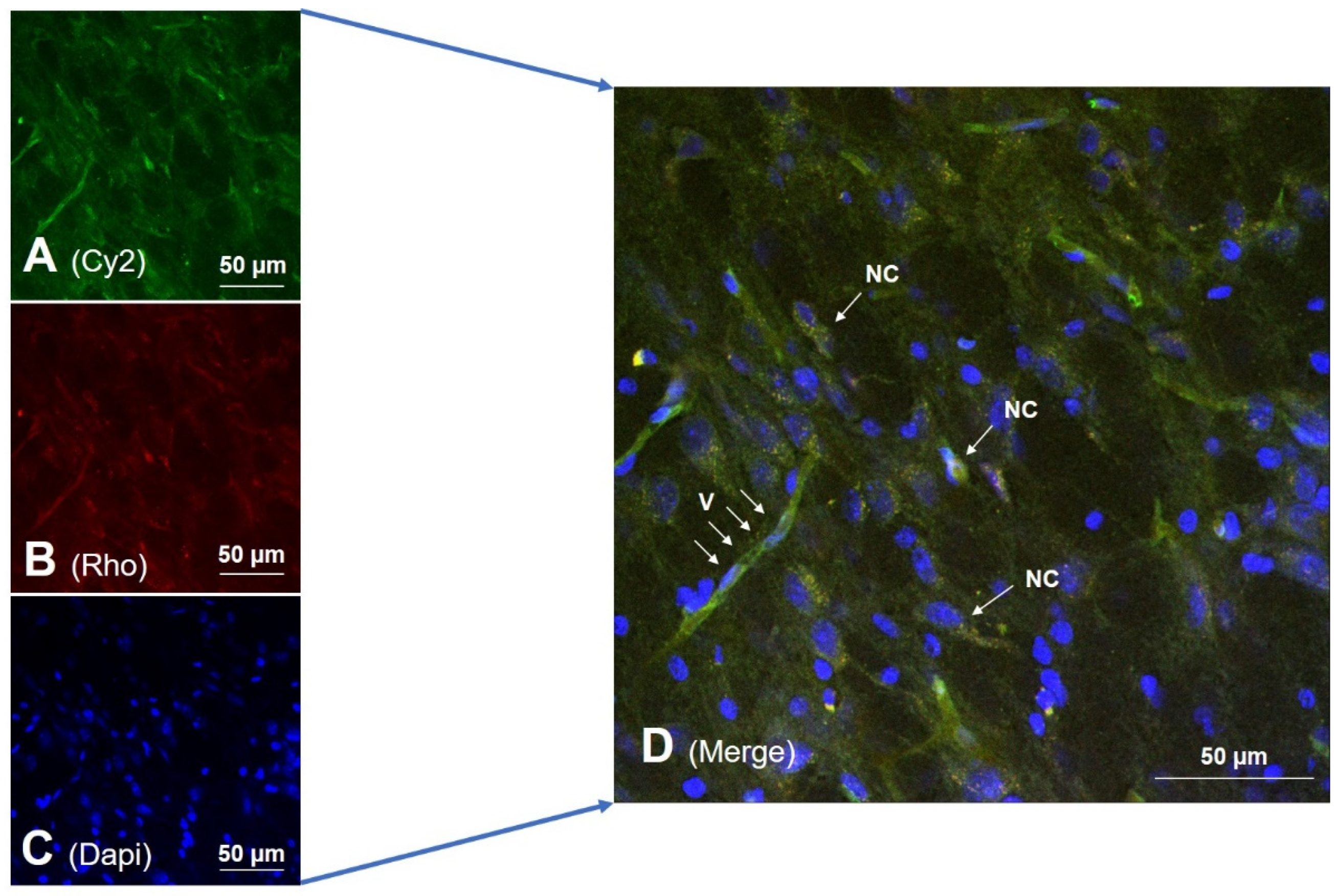{kind=link}
| Therapy | Advantages | Disadvantages |
|---|---|---|
| Intravenous thrombolysis |
|
|
| Thrombectomy combined with arterial thrombolysis |
|
|
| Decompressive surgery |
|
|
| Hypothermia |
|
|
Several pharmacological therapies:
|
|
|
| Blood glutamate grabbers |
|
|
| Statins |
|
|
| miRNAs |
|
|
| Drug delivery systems (DDS) | ||
| Cell penetrating peptides (CPPs) |
|
|
Cell membrane-derived vesicles (CMVs):
|
|
|
| NPs: |
| |
|
|
|
|
|
|
|
|
|
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Blanco, S.; Martínez-Lara, E.; Siles, E.; Peinado, M.Á. New Strategies for Stroke Therapy: Nanoencapsulated Neuroglobin. Pharmaceutics 2022, 14, 1737. https://doi.org/10.3390/pharmaceutics14081737
Blanco S, Martínez-Lara E, Siles E, Peinado MÁ. New Strategies for Stroke Therapy: Nanoencapsulated Neuroglobin. Pharmaceutics. 2022; 14(8):1737. https://doi.org/10.3390/pharmaceutics14081737
Chicago/Turabian StyleBlanco, Santos, Esther Martínez-Lara, Eva Siles, and María Ángeles Peinado. 2022. "New Strategies for Stroke Therapy: Nanoencapsulated Neuroglobin" Pharmaceutics 14, no. 8: 1737. https://doi.org/10.3390/pharmaceutics14081737