
The 46.1 Antibody Mediates Neurotensin Uptake into the CNS and the Effects Depend on the Route of Intravenous Administration

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
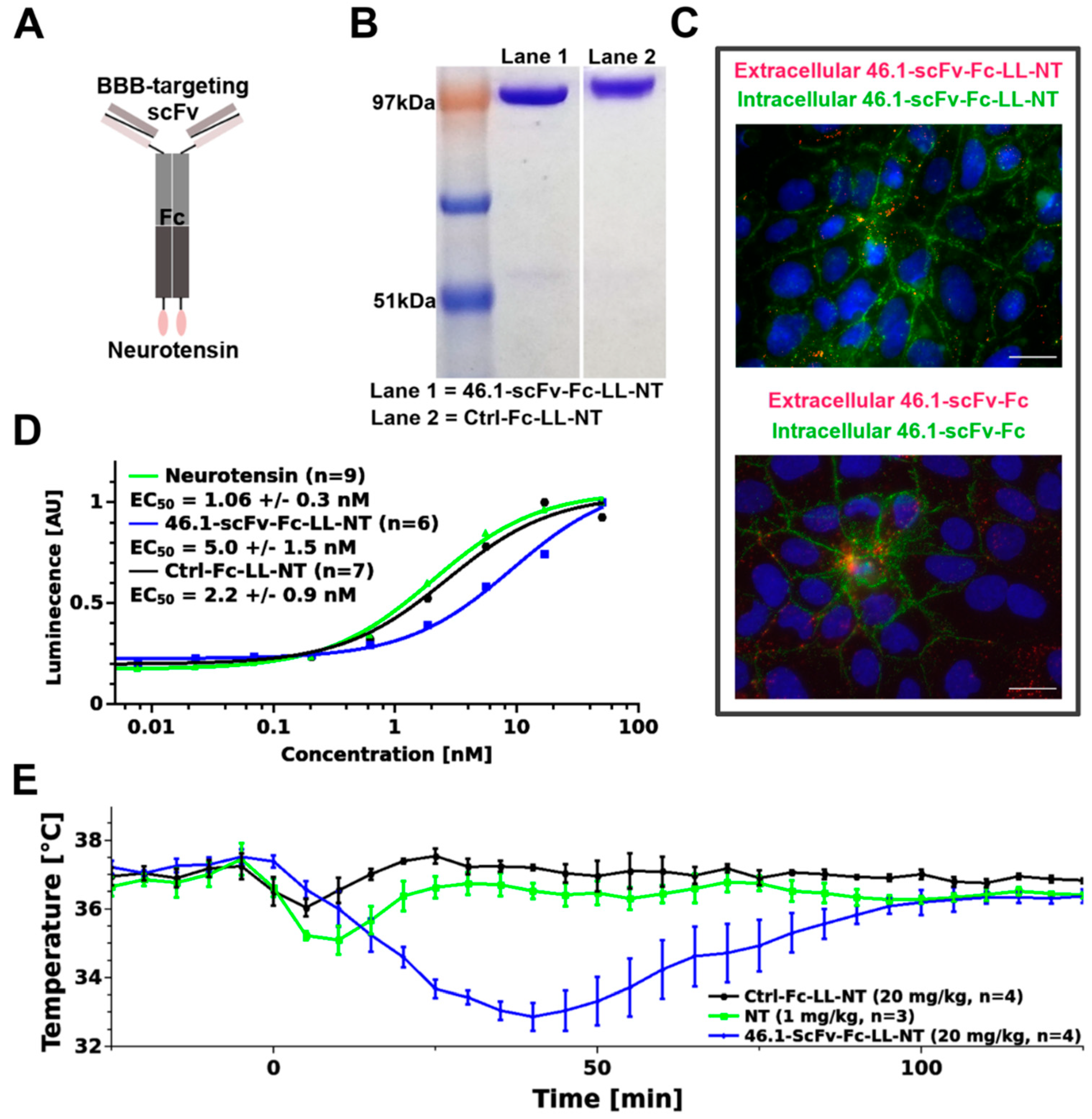
2.1. Construction of Neurotensin–Antibody Fusion Proteins
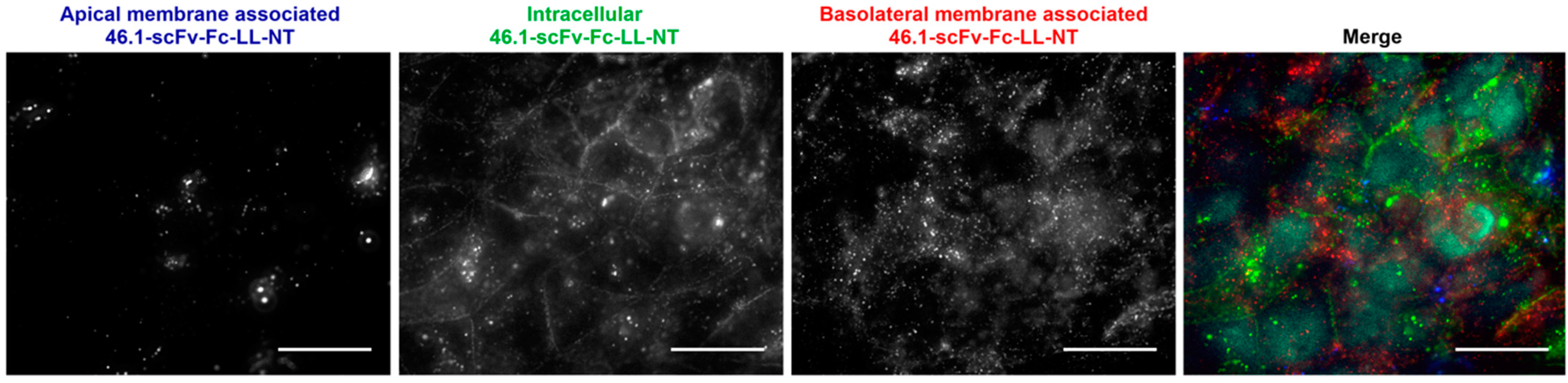
2.2. Microvascular Endothelial Cell Immunocytochemistry
2.3. Animal Experiments
2.4. Logger Implantation and Temperature Measurement
2.5. Locomotor Activity Measurement
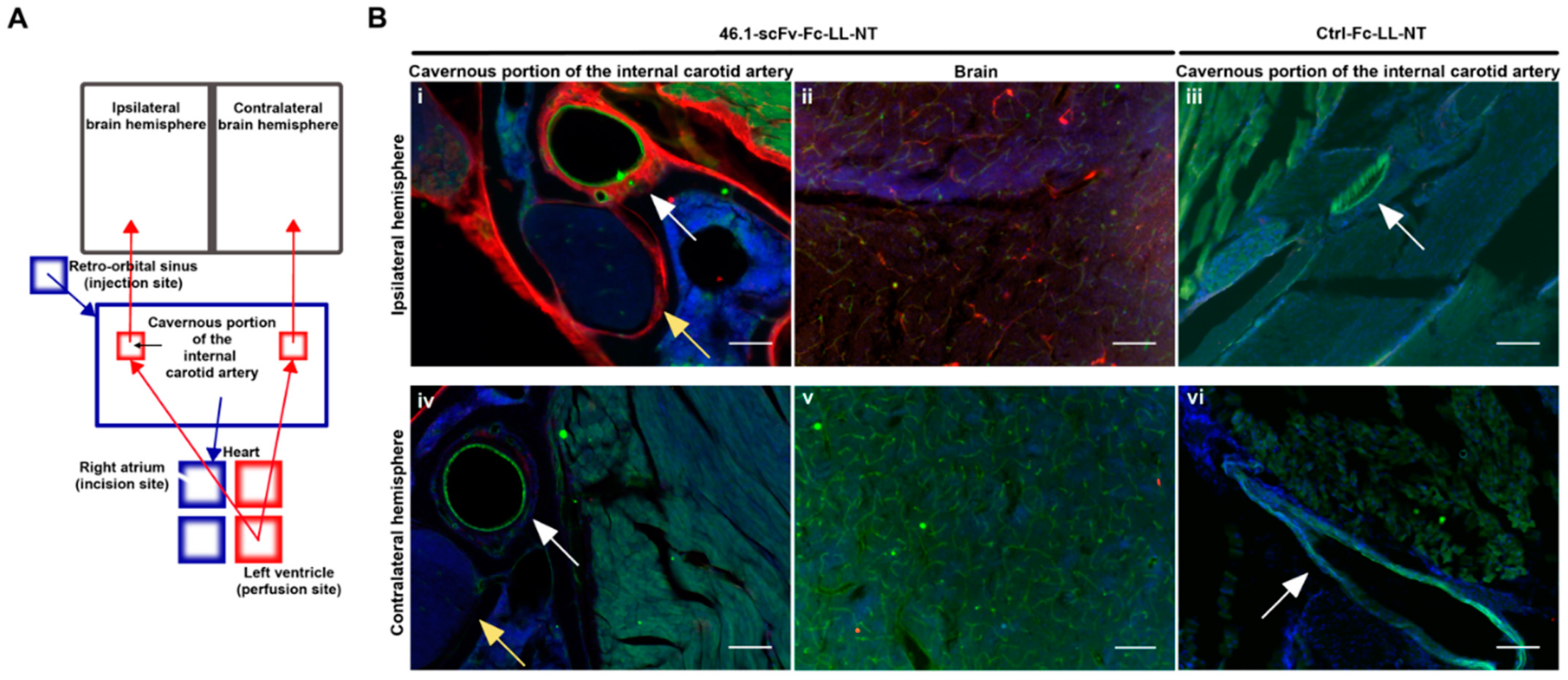
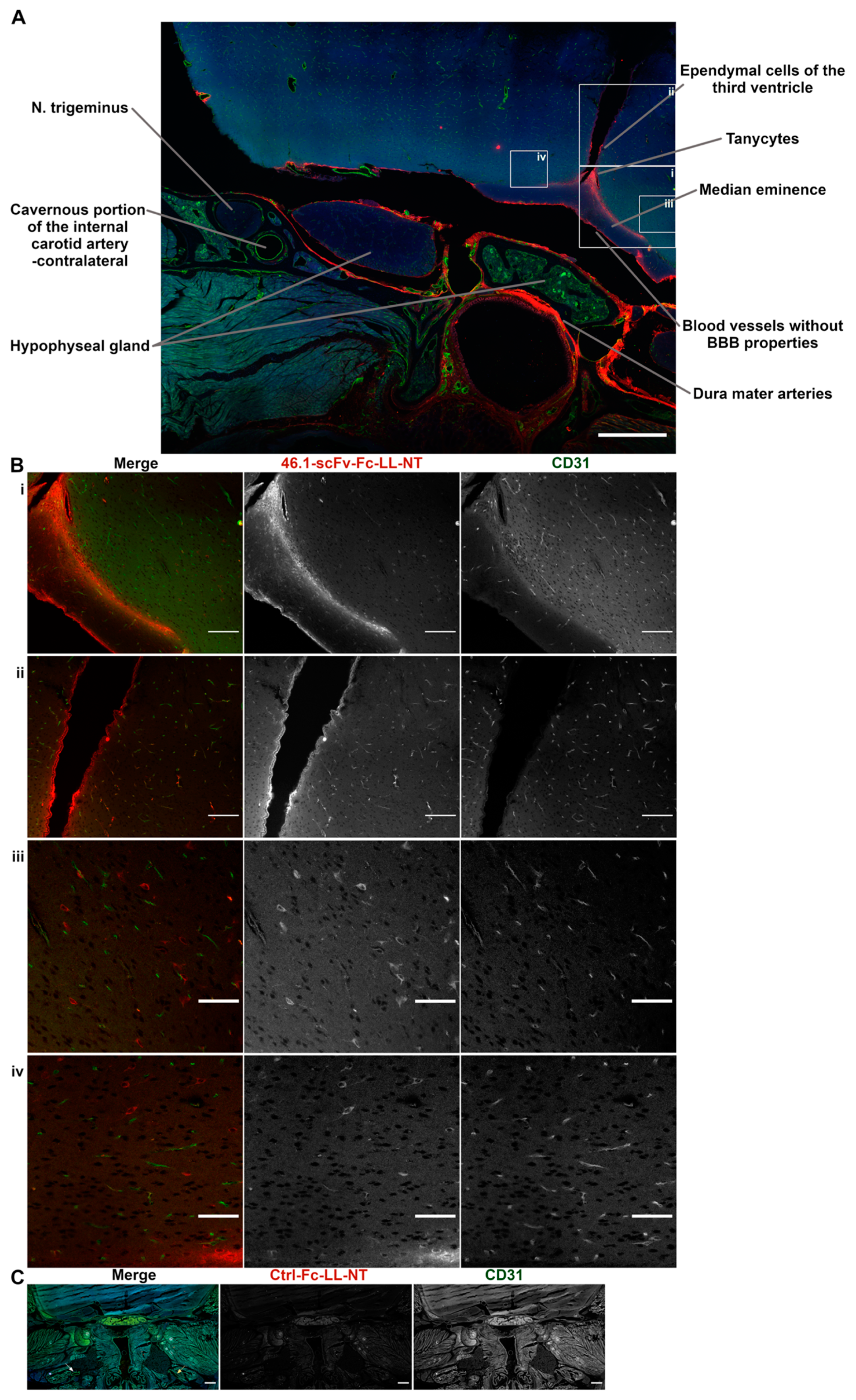
2.6. Surgery and Cavernous Sinus Immunohistochemistry
2.7. Statistical Analysis
3. Results
3.1. ScFv 46.1 Mediates the Transport of Neurotensin across the Blood Brain Barrier and Its Accumulation in the Median Preoptic Nucleus
3.2. ScFv 46.1 Mediates the Delivery of Neurotensin to the Striatum
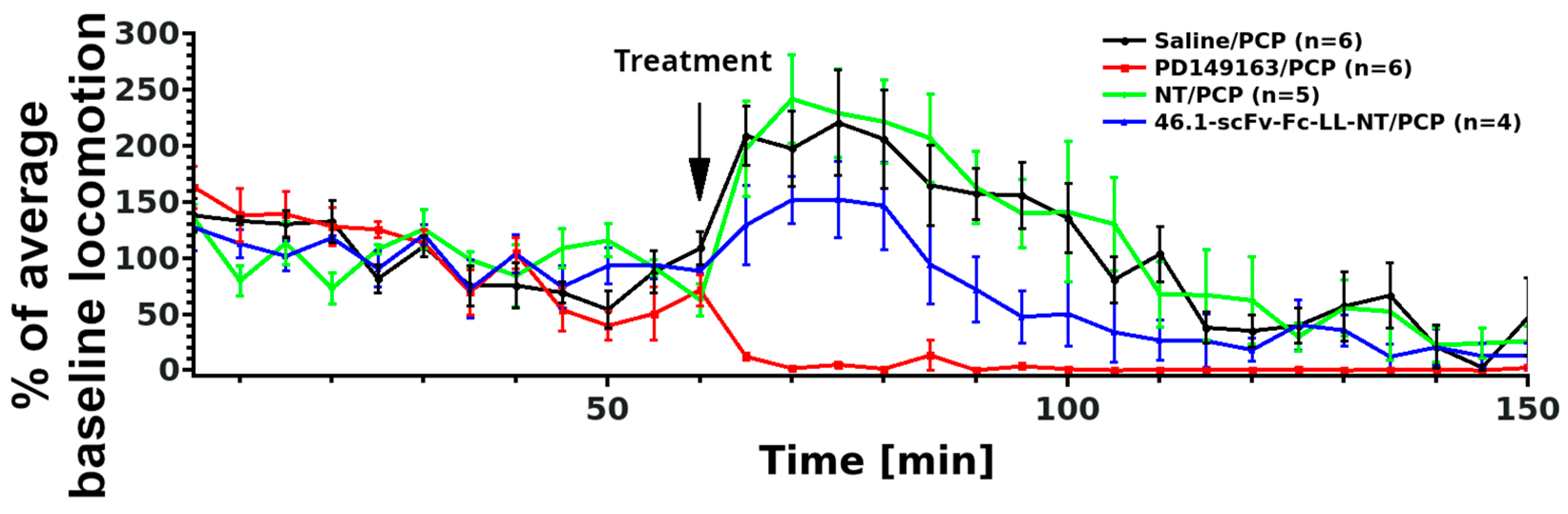
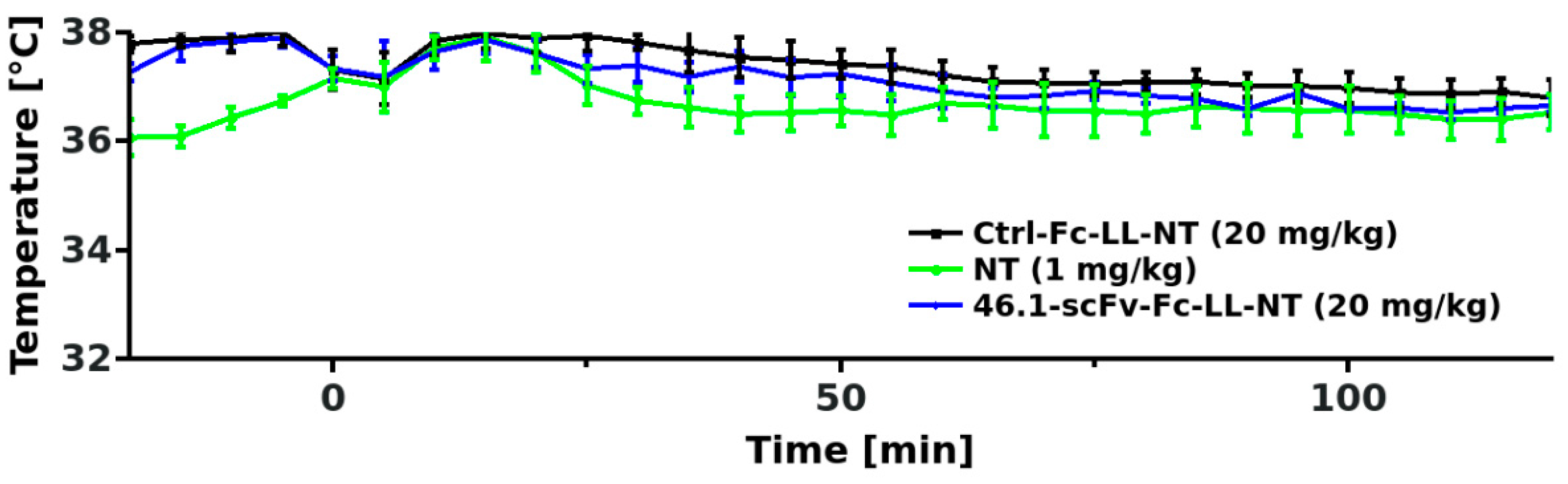
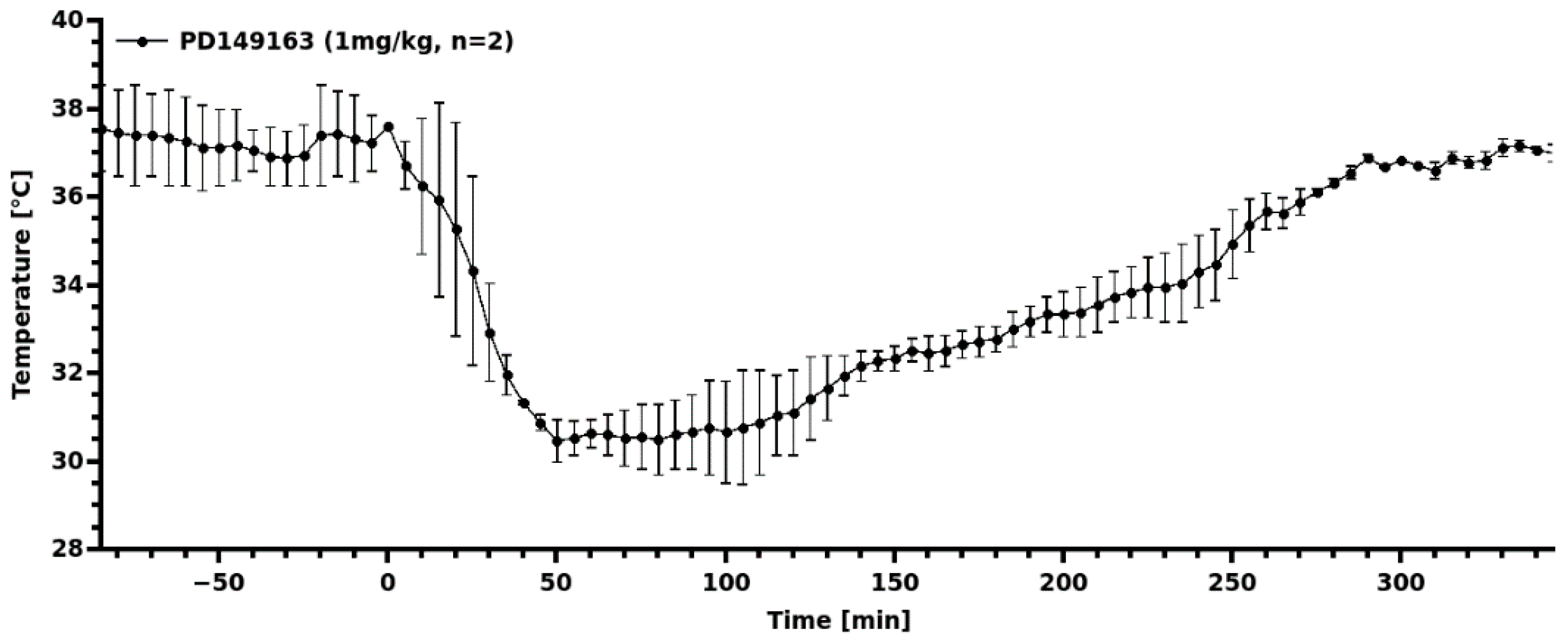
3.3. Transient Hypothermia Response Depends on the Route of Delivery
3.4. Trans-Carotid Transport of ScFv 46.1 at the Cavernous Sinus Allows for Enhanced Accumulation of the Antibody-NT Construct in the Brain after Retro-Orbital Sinus Administration
4. Discussion and Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Comparison | ||||||
|---|---|---|---|---|---|---|
| Treatment | Treatment | Mean Difference | SE | df | t | PHolm |
| 46.1-scFv-Fc-LL-NT (20 mg/kg) | Ctrl-Fc-LL-NT (20 mg/kg) | 2.8 | 1.22 | 19.0 | 2.3 | 0.066 |
| Neurotensin (1 mg/kg) | 3.95 | 1.16 | 19.0 | 3.41 | 0.009 | |
| Ctrl-Fc-LL-NT (20 mg/kg) | Neurotensin (1 mg/kg) | 1.14 | 1.11 | 19.0 | 1.03 | 0.314 |
| Comparison | ||||||
|---|---|---|---|---|---|---|
| Treatment | Treatment | Mean Difference | SE | df | t | PHolm |
| 46.1-scFv-Fc-LL-NT (20 mg/kg) | Ctrl-Fc-LL-NT (20 mg/kg) | −1.888 | 0.323 | 8.0 | −5.84 | 0.001 |
| Neurotensin (1 mg/kg) | −1.281 | 0.349 | 8.0 | −3.67 | 0.013 | |
| Ctrl-Fc-LL-NT (20 mg/kg) | Neurotensin (1 mg/kg) | −55.22 | 0.349 | 8.0 | 1.74 | 0.12 |



References
- Pardridge, W.M. Alzheimer’s disease drug development and the problem of the blood-brain barrier. Alzheimer’s Dement. 2009, 5, 427–432. [Google Scholar] [CrossRef] [PubMed]
- Couch, J.A.; Yu, Y.J.; Zhang, Y.; Tarrant, J.M.; Fuji, R.N.; Meilandt, W.J.; Solanoy, H.; Tong, R.K.; Hoyte, K.; Luk, W.; et al. Addressing safety liabilities of TfR bispecific antibodies that cross the blood-brain barrier. Sci. Transl. Med. 2013, 5, 183ra57. [Google Scholar] [CrossRef] [PubMed]
- Jones, A.R.; Shusta, E.V. Blood-brain barrier transport of therapeutics via receptor-mediation. Pharm. Res. 2007, 24, 1759–1771. [Google Scholar] [CrossRef] [PubMed]
- Pardridge, W.M.; Kang, Y.S.; Buciak, J.L. Transport of human recombinant brain-derived neurotrophic factor ({BDNF}) through the rat blood-brain barrier in vivo using vector-mediated peptide drug delivery. Pharm. Res. 1994, 11, 738–746. [Google Scholar] [CrossRef]
- Pardridge, W.M.; Kang, Y.S.; Buciak, J.L.; Yang, J. Human insulin receptor monoclonal antibody undergoes high affinity binding to human brain capillaries in vitro and rapid transcytosis through the blood-brain barrier in vivo in the primate. Pharm. Res. 1995, 12, 807–816. [Google Scholar] [CrossRef]
- Ullman, J.C.; Arguello, A.; Getz, J.A.; Bhalla, A.; Mahon, C.S.; Wang, J.; Giese, T.; Bedard, C.; Kim, D.J.; Blumenfeld, J.R.; et al. Brain delivery and activity of a lysosomal enzyme using a blood-brain barrier transport vehicle in mice. Sci. Transl. Med. 2020, 12, eaay1163. [Google Scholar] [CrossRef]
- Kariolis, M.S.; Wells, R.C.; Getz, J.A.; Kwan, W.; Mahon, C.S.; Tong, R.; Kim, D.J.; Srivastava, A.; Bedard, C.; Henne, K.R.; et al. Brain delivery of therapeutic proteins using an Fc fragment blood-brain barrier transport vehicle in mice and monkeys. Sci. Transl. Med. 2020, 12, eaay1359. [Google Scholar] [CrossRef]
- Stocki, P.; Szary, J.; Rasmussen, C.L.M.; Demydchuk, M.; Northall, L.; Logan, D.B.; Gauhar, A.; Thei, L.; Moos, T.; Walsh, F.S.; et al. Blood-brain barrier transport using a high affinity, brain-selective VNAR antibody targeting transferrin receptor 1. FASEB J. 2021, 35, e21172. [Google Scholar] [CrossRef]
- Georgieva, J.V.; Goulatis, L.I.; Stutz, C.C.; Canfield, S.G.; Song, H.W.; Gastfriend, B.D.; Shusta, E.V. Antibody screening using a human iPSC-based blood-brain barrier model identifies antibodies that accumulate in the CNS. FASEB J. 2020, 34, 12549–12564. [Google Scholar] [CrossRef]
- Wouters, Y.; Jaspers, T.; De Strooper, B.; Dewilde, M. Identification and in vivo characterization of a brain-penetrating nanobody. Fluids Barriers CNS 2020, 17, 62. [Google Scholar] [CrossRef]
- Demeule, M.; Beaudet, N.; Régina, A.; Besserer-Offroy, É.; Murza, A.; Tétreault, P.; Belleville, K.; Ché, C.; Larocque, A.; Thiot, C.; et al. Conjugation of a brain-penetrant peptide with neurotensin provides antinociceptive properties. J. Clin. Investig. 2014, 124, 1199–1213. [Google Scholar] [CrossRef] [PubMed]
- Ratner, C.; He, Z.; Grunddal, K.V.; Skov, L.J.; Hartmann, B.; Zhang, F.; Feuchtinger, A.; Bjerregaard, A.; Christoffersen, C.; Tschöp, M.H.; et al. Long-acting neurotensin synergizes with liraglutide to reverse obesity through a melanocortin-dependent pathway. Diabetes 2019, 68, 1329–1340. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.-P.; Wang, J.; Dong, Y.-L.; Wang, Y.-Y.; Li, Y.-Q. The Roles of Neurotensin and its Analogues in Pain. Curr. Pharm. Des. 2014, 21, 840–848. [Google Scholar] [CrossRef]
- Liang, Y.; Boules, M.; Shaw, A.M.; Williams, K.; Fredrickson, P.; Richelson, E. Effect of a novel neurotensin analog, NT69L, on nicotine-induced alterations in monoamine levels in rat brain. Brain Res. 2008, 1231, 6–15. [Google Scholar] [CrossRef] [PubMed]
- Boules, M.; Netz, R.; Fredrickson, P.A.; Richelson, E. A neurotensin analog blocks cocaine-conditioned place preference and reinstatement. Behav. Pharmacol. 2016, 27, 236–239. [Google Scholar] [CrossRef]
- Sharpe, A.L.; Varela, E.; Beckstead, M.J. Systemic PD149163, a neurotensin receptor 1 agonist, decreases methamphetamine self-administration in DBA/2J mice without causing excessive sedation. PLoS ONE 2017, 12, e0180710. [Google Scholar] [CrossRef]
- Binder, E.B.; Kinkead, B.; Owens, M.J.; Nemeroff, C.B. Neurotensin and dopamine interactions. Pharmacol. Rev. 2001, 53, 453–486. [Google Scholar]
- Cáceda, R.; Kinkead, B.; Nemeroff, C.B. Neurotensin: Role in psychiatric and neurological diseases. Peptides 2006, 27, 2385–2404. [Google Scholar] [CrossRef]
- Tabarean, I.V. Neurotensin induces hypothermia by activating both neuronal neurotensin receptor 1 and astrocytic neurotensin receptor 2 in the median preoptic nucleus. Neuropharmacology 2020, 171, 108069. [Google Scholar] [CrossRef]
- Schoch, A.; Thorey, I.S.; Engert, J.; Winter, G.; Emrich, T. Comparison of the lateral tail vein and the retro-orbital venous sinus routes of antibody administration in pharmacokinetic studies. Lab. Anim. 2014, 43, 95–99. [Google Scholar] [CrossRef]
- Foust, K.D.; Nurre, E.; Montgomery, C.L.; Hernandez, A.; Chan, C.M.; Kaspar, B.K. Intravascular AAV9 preferentially targets neonatal neurons and adult astrocytes. Nat. Biotechnol. 2009, 27, 59–65. [Google Scholar] [CrossRef] [PubMed]
- Gruntman, A.M.; Su, L.; Flotte, T.R. Retro-Orbital Venous Sinus Delivery of rAAV9 Mediates High-Level Transduction of Brain and Retina Compared with Temporal Vein Delivery in Neonatal Mouse Pups. Hum. Gene Ther. 2017, 28, 228–230. [Google Scholar] [CrossRef] [PubMed]
- Liguore, W.A.; Domire, J.S.; Button, D.; Wang, Y.; Dufour, B.D.; Srinivasan, S.; McBride, J.L. AAV-PHP.B Administration Results in a Differential Pattern of CNS Biodistribution in Non-human Primates Compared with Mice. Mol. Ther. 2019, 27, 2018–2037. [Google Scholar] [CrossRef] [PubMed]
- Harris, F.S.; Rhoton, A.L. Anatomy of the cavernous sinus. A microsurgical study. J. Neurosurg. 1976, 45, 169–180. [Google Scholar] [CrossRef]
- Yang, W.J.; Wong, K.S.; Chen, X.Y. Intracranial atherosclerosis: From microscopy to high-resolution magnetic resonance imaging. J. Stroke 2017, 19, 249–262. [Google Scholar] [CrossRef]
- Masuoka, T.; Hayashi, N.; Hori, E.; Kuwayama, N.; Ohtani, O.; Endo, S. Distribution of internal elastic lamina and external elastic lamina in the internal carotid artery: Possible relationship with atherosclerosis. Neurol. Med. Chir. 2010, 50, 179–182. [Google Scholar] [CrossRef]
- Lippmann, E.S.; Azarin, S.M.; Kay, J.E.; Nessler, R.A.; Wilson, H.K.; Al-Ahmad, A.; Palecek, S.P.; Shusta, E. V Derivation of blood-brain barrier endothelial cells from human pluripotent stem cells. Nat. Biotechnol. 2012, 30, 783–791. [Google Scholar] [CrossRef]
- Lippmann, E.S.; Al-Ahmad, A.; Azarin, S.M.; Palecek, S.P.; Shusta, E.V. A retinoic acid-enhanced, multicellular human blood-brain barrier model derived from stem cell sources. Sci. Rep. 2014, 4, 4160. [Google Scholar] [CrossRef]
- Westfall, P.H.; Johnson, W.O.; Utts, J.M. A bayesian perspective on the bonferroni adjustment. Biometrika 1997, 84, 419–427. [Google Scholar] [CrossRef]
- Bissette, G.; Nemeroff, C.B.; Loosen, P.T.; Prange, A.J.; Lipton, M.A. Hypothermia and intolerance to cold induced by intracisternal administration of the hypothalamic peptide neurotensin. Nature 1976, 262, 607–609. [Google Scholar] [CrossRef]
- Han, B.W.; Herrin, B.R.; Cooper, M.D.; Wilson, I.A. Antigen recognition by variable lymphocyte receptors. Science 2008, 321, 1834–1837. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Boules, M.; Williams, K.; Peris, J.; Richelson, E. The novel neurotensin analog NT69L blocks phencyclidine (PCP)-induced increases in locomotor activity and PCP-induced increases in monoamine and amino acids levels in the medial prefrontal cortex. Brain Res. 2010, 1311, 28–36. [Google Scholar] [CrossRef] [PubMed]
- Chou, S.; Davis, C.; Jones, S.; Li, M. Repeated effects of the neurotensin receptor agonist PD149163 in three animal tests of antipsychotic activity: Assessing for tolerance and cross-tolerance to clozapine. Pharmacol. Biochem. Behav. 2014, 128, 78–88. [Google Scholar] [CrossRef]
- Ervin, G.N.; Birkemo, L.S.; Nemeroff, C.B.; Prange, A.J. Neurotensin blocks certain amphetamine-induced behaviours. Nature 1981, 291, 73–76. [Google Scholar] [CrossRef] [PubMed]
- Kalivas, P.W.; Nemeroff, C.B.; Prange, A.J. Neurotensin microinjection into the nucleus accumbens antagonizes dopamine-induced increase in locomotion and rearing. Neuroscience 1984, 11, 919–930. [Google Scholar] [CrossRef]
- Sarhan, S.; Hitchcock, J.M.; Grauffel, C.A.; Wettstein, J.G. Comparative antipsychotic profiles of neurotensin and a related systemically active peptide agonist. Peptides 1997, 18, 1223–1227. [Google Scholar] [CrossRef]
- Hackler, E.A.; Byun, N.E.; Jones, C.K.; Williams, J.M.; Baheza, R.; Sengupta, S.; Grier, M.D.; Avison, M.; Conn, P.J.; Gore, J.C. Selective potentiation of the metabotropic glutamate receptor subtype 2 blocks phencyclidine-induced hyperlocomotion and brain activation. Neuroscience 2010, 168, 209–218. [Google Scholar] [CrossRef]
- Lee, M.D.; Wagenmakers, E.J. Bayesian Cognitive Modeling: A Practical Course; Cambridge University Press: Cambridge, UK, 2014; ISBN 9781139087759. [Google Scholar]
- Krzymowski, T.; Stefanczyk-Krzymowska, S. New facts and the concept of physiological regulation of the dopaminergic system function and its disorders. J. Physiol. Pharmacol. 2015, 66, 331–341. [Google Scholar]
- Socher, M.; Kuntz, J.; Sawall, S.; Bartling, S.; Kachelrieß, M. The retrobulbar sinus is superior to the lateral tail vein for the injection of contrast media in small animal cardiac imaging. Lab. Anim. 2014, 48, 105–113. [Google Scholar] [CrossRef]
- Petrie, K.A.; Bubser, M.; Casey, C.D.; Davis, M.D.; Roth, B.L.; Deutch, A.Y. The neurotensin agonist PD149163 increases fos expression in the prefrontal cortex of the rat. Neuropsychopharmacology 2004, 29, 1878–1888. [Google Scholar] [CrossRef]
- Gene Erwin, V.; Jones, B.C. Comparison of neurotensin levels, receptors and actions in LS/Ibg and SS/Ibg mice. Peptides 1989, 10, 435–440. [Google Scholar] [CrossRef]
- Einer-Jensen, N.; Larsen, L. Transfer of tritiated water, tyrosine, and propanol from the nasal cavity to cranial arterial blood in rats. Exp. Brain Res. 2000, 130, 216–220. [Google Scholar] [CrossRef] [PubMed]
- Einer-Jensen, N.; Larsen, L. Local transfer of diazepam, but not of cocaine, from the nasal cavities to the brain arterial blood in rats. Pharmacol. Toxicol. 2000, 87, 276–278. [Google Scholar] [CrossRef]
- Muszak, J.; Krzymowski, T.; Gilun, P.; Stefanczyk-Krzymowska, S. Countercurrent transfer of dopamine from venous blood in the cavernous sinus to the arterial blood supplying the brain—Tlhe perfused rabbit head as an experimental model. J. Physiol. Pharmacol. 2014, 65, 641–648. [Google Scholar]
- Wolak, D.J.; Pizzo, M.E.; Thorne, R.G. Probing the extracellular diffusion of antibodies in brain using in vivo integrative optical imaging and ex vivo fluorescence imaging. J. Control. Release 2015, 197, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Dye, J.; Duckwiler, G.; Gonzalez, N.; Kaneko, N.; Goldberg, R.; Rootman, D.; Jahan, R.; Tateshima, S.; Szeder, V. Endovascular approaches to the cavernous sinus in the setting of dural arteriovenous fistula. Brain Sci. 2020, 10, 554. [Google Scholar] [CrossRef]
- Teramoto, A.; Nemoto, S.; Takakura, K.; Sasaki, Y.; Machida, T. Selective venous sampling directly from cavernous sinus in cushing’s syndrome. J. Clin. Endocrinol. Metab. 1993, 76, 637–641. [Google Scholar] [CrossRef]
- Wahl, A.S.; Correa, D.; Imobersteg, S.; Maurer, M.A.; Kaiser, J.; Augath, M.A.; Schwab, M.E. Targeting Therapeutic Antibodies to the CNS: A Comparative Study of Intrathecal, Intravenous, and Subcutaneous Anti-Nogo A Antibody Treatment after Stroke in Rats. Neurotherapeutics 2020, 17, 1153–1159. [Google Scholar] [CrossRef]
- Doyle, T.D.; Anand, S.; Edens, M.A. Scalp Catheterization; StatPearls Publishing: Treasure Island, FL, USA, 2021. [Google Scholar]
- Takaku, A.; Suzuki, J. A new method of orbital and cavernous sinus venography. Technical note. J. Neurosurg. 1969, 30, 200–205. [Google Scholar] [CrossRef]
- Hoy, S.M. Onasemnogene Abeparvovec: First Global Approval. Drugs 2019, 79, 1255–1262. [Google Scholar] [CrossRef]




| Comparison | |||||||
|---|---|---|---|---|---|---|---|
| Treatment | Treatment | Mean Difference | SE | df | t | p | PHolm |
| Saline | 46.1-scFv-Fc-LL-NT (20 mg/kg) | 47.46 | 22.3 | 17.0 | 2.128 | 0.048 | 0.096 |
| Neurotensin (1 mg/kg) | −7.76 | 20.9 | 17.0 | −0.371 | 0.715 | 0.715 | |
| PD149163 (1 mg/kg) | 105.2 | 19.9 | 17.0 | 5.275 | <0.001 | <0.001 | |
| 46.1-scFv-Fc-LL-NT (20 mg/kg) | Neurotensin (1 mg/kg) | −55.22 | 23.2 | 17.0 | −2.383 | 0.029 | 0.087 |
| PD149163 (1 mg/kg) | 57.74 | 22.3 | 17.0 | 2.589 | 0.019 | 0.076 | |
| Neurotensin (1 mg/kg) | PD149163 (1 mg/kg) | 112.96 | 20.9 | 17.0 | 5.400 | <0.001 | <0.001 |
| Comparison | |||||
|---|---|---|---|---|---|
| Treatment | Treatment | Prior Odds | Posterior Odds | BF 10, U | Error (%) |
| Saline | 46.1-scFv-Fc-LL-NT (20 mg/kg) | 0.414 | 29.136 | 70.34 | 2.603 × 10−8 |
| Neurotensin (1 mg/kg) | 0.414 | 0.074 | 0.178 | 4.210 × 10−6 | |
| PD149163 (1 mg/kg) | 0.414 | 1.093 × 1020 | 2.638 × 1020 | 3.010 × 10−27 | |
| 46.1-scFv-Fc-LL-NT (20 mg/kg) | Neurotensin (1 mg/kg) | 0.414 | 61.063 | 147.42 | 1.607 × 10−9 |
| PD149163 (1 mg/kg) | 0.414 | 5.637 × 1012 | 1.361 × 1013 | 2.716 × 10−17 | |
| Neurotensin (1 mg/kg) | PD149163 (1 mg/kg) | 0.414 | 1.033 × 1019 | 2.493 × 1019 | 2.053 × 10−26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Georgieva, J.V.; Katt, M.; Ye, Z.; Umlauf, B.J.; Wenthur, C.J.; Shusta, E.V. The 46.1 Antibody Mediates Neurotensin Uptake into the CNS and the Effects Depend on the Route of Intravenous Administration. Pharmaceutics 2022, 14, 1706. https://doi.org/10.3390/pharmaceutics14081706
Georgieva JV, Katt M, Ye Z, Umlauf BJ, Wenthur CJ, Shusta EV. The 46.1 Antibody Mediates Neurotensin Uptake into the CNS and the Effects Depend on the Route of Intravenous Administration. Pharmaceutics. 2022; 14(8):1706. https://doi.org/10.3390/pharmaceutics14081706
Chicago/Turabian StyleGeorgieva, Julia V., Moriah Katt, Zhou Ye, Benjamin J. Umlauf, Cody J. Wenthur, and Eric V. Shusta. 2022. "The 46.1 Antibody Mediates Neurotensin Uptake into the CNS and the Effects Depend on the Route of Intravenous Administration" Pharmaceutics 14, no. 8: 1706. https://doi.org/10.3390/pharmaceutics14081706