
Activity of Apo-Lactoferrin on Pathogenic Protozoa


Abstract
:1. Introduction
1.1. Parasitism as a Way of Life
1.2. Opportunistic Free-Living Amoebae as Pathogenic Protozoa
2. Activities of Lactoferrin on Pathogenic Protozoa
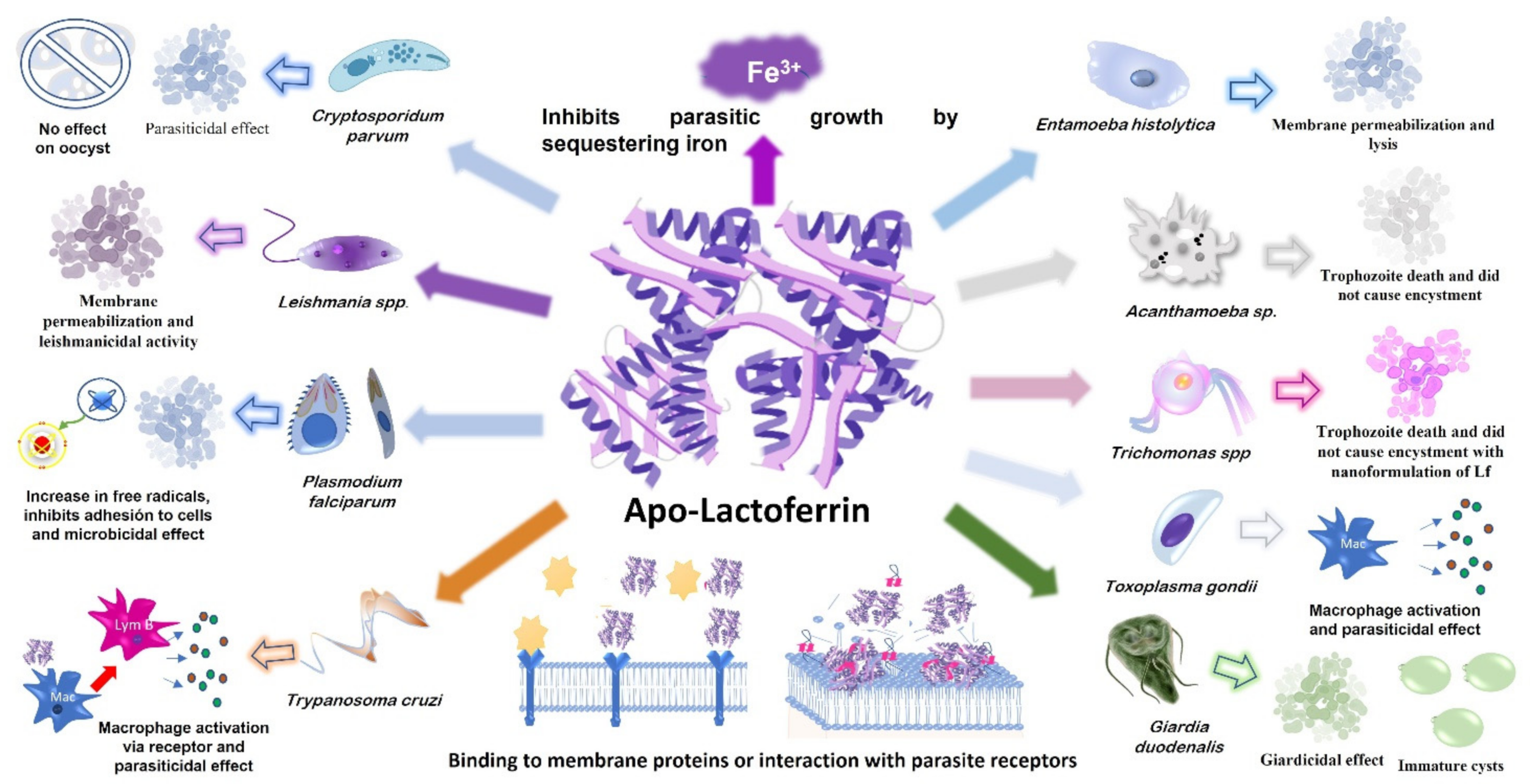
2.1. Apo-Lf Can Damage and Kill Pathogenic Protozoa
2.2. Holo-Lf Can Be an Iron Source for Pathogenic Protozoa
3. Effect of Lactoferrin on Pathogenic Parasites
3.1. Cryptosporidium spp.
3.2. Eimeria spp.
3.3. Entamoeba histolytica
3.4. Giardia duodenalis
3.5. Leishmania spp.
3.6. Trypanosoma spp.
3.7. Trypanosoma cruzi
3.8. Trypanosoma brucei
3.9. Plasmodium spp. and Babesia spp.
3.10. Toxoplasma gondii
3.11. Trichomonas vaginalis

{kind=link}
{kind=link}
| Disease/Pathogen | Treatment | Effect | Reference |
|---|---|---|---|
| Cryptosporidiosis/Cryptosporidium parvum | hLf, hLf hydrolyzed, hLfcin B 10 µg/mL | Lf hydrolyzed and Lfcin B affected the viability, showed parasiticidal effect, and decreased the infectivity. Lf had no effect. | Murdock, C.A. and K.R. Matthews, 2002 [27] |
| Lf 10 mg/mL | Lf affected the viability, showed parasiticidal effect, and decreased the infectivity. No significant effect was observed on the oocyst phase or parasite intracellular development. | Paredes, J.L., et al., 2017 [29] | |
| Eimeriosis/ Eimeria stiedai | bLfcin B 1000 ug/mL (In vivo rabbits and mice) | Less infectivity and less penetration into host cells, lower number of oocysts, the liver had fewer abscesses, and they did not present inflammation of the bile ducts. | Omata, Y., et al., 2001 [35] |
| Amoebiasis/ Entamoeba histolytica | hLf, hLfcin B, bLf, bLfcin B 100 µM Metronidazole 58.4 µM | The amoebicidal effect and the synergistic amoebicidal effect was found between metronidazole and Lf or metronidazole and Lfcin. | Leon-Sicairos, N., et al., 2006 [46] |
| hLf, sIgA, Lysozyme 100 µM | The amoebicidal effect, rearrangements, and disruption in the lipid pattern after being bound to the amoeba membrane. | Leon-Sicairos, N., et al., 2006 [51] | |
| Lfcin 17–30, Lfampin 265–284, Lfchimera 100 µM | Amoebicidal effect, Lfchimera showed the strongest amoebicidal activity. | Lopez-Soto, F., et al., 2010 [53] | |
| bLf 20 mg/kg (In vivo C3H/HeJ mice) | Elimination of amoebiasis by the production of anti-amoeba IgA antibodies and amoebicidal effect. | León-Sicairos, N., et al., 2012 [56] | |
| bLf 2.5 mg/100 g Metronidazole 0.5 mg/100 g (In vivo Syrian hamster) | No clinical signs of disease and amoebiasis was effectively decreased. Liver function and blood cells approached normal levels. | Ordaz-Pichardo, C., et al., 2012 [58] | |
| bLfcin-B, bLfcin 17–30, bLfampin 265–284, 250, 1000 µM bLfcin-B, bLfcin 17–30, bLfampin 265–284 10 mg/kg (In vivo C3H/HeJ mice) | Concentrations higher than 250 µM of Lfampin showed necrosis. No effect of Lfcin-B or Lfcin 17–30. Mice in the groups treated with Lfcin 17–30 or Lfcin B showed an absence of amoebic trophozoites in the intestinal lumen on 75% of the animals and treatment with Lfampin eradicated the infection in 100%. | Diaz-Godinez, C., et al., 2019 [59] | |
| Giardiasis/ Giardia duodenalis | hLf, 2.5 mg/mL bLf, 2.0 mg/mL hLfcin 18–40, 24 µg/mL bLfcin 17–41, 12 µg/mL. | bLfcin had the most potent giardicidal activity. Log-phase cells were more resistant to killing than stationary-phase cells | Turchany, J.M., 1995 [63] |
| bLf, 12.5 μM bLfcin 2.6 μM | Trophozoites exhibited structural changes in membranes, cell growth cessation, and production of immature cysts. | Frontera, L.S., et al., 2018 [60] | |
| bLfcin17-30, bLfampin265-284, Lfchimera 40 µM metronidazole 100 µM albendazole 5 µM | Parasiticidal effect. Apoptosis of trophozoites. Lfampin showed the best microbicidal activity. Lf and its derivative peptides showed synergy with the drugs. | Aguilar-Díaz, H., et al., 2017 [65,66] | |
| Lf, maltodextrin 0.5 g (in vivo, children aged 12–36 months) | No significant difference between the two groups in symptomatology. There was a lower prevalence of colonization with Giardia spp. and better growth in children of the group treated with Lf | Ochoa, T.J., et al., 2008 [67] | |
| Leishmaniasis/ Leishmania donovani | Lfcin 17–30, Lfampin 265–284, Lfchimera 10 µM | LFchimera was the most active peptide. All peptides induced plasma membrane permeabilization and bioenergetic collapse of the parasites | Silva, T., et al., 2012 [75] |
| Nanoparticles of Lf and amphotericin B. (In vivo Syrian hamsters) 0.8 µg/mL | Leishmanicidal effect. Increased protective of proinflammatory mediators expression and down-regulation of disease-promoting cytokines. | Asthana, S., et al., 2015 [76] | |
| Nanoparticles of Lf with betulinic acid (In vivo Balb/c mouse) 1.5 mg/mL | Leishmanicidal effect. Reduced anti-inflammatory cytokine IL-10 and increased nitric oxide. | Halder, A., et al., 2018 [77] | |
| Chagas disease/Trypanosoma cruzi | hLf 10 µg/mL | Microbicidal effect. Lf stimulated the killing of amastigotes by macrophage activation via oxygen reduction intermediates. | Lima, M.F. and F. Kierszenbaum, 1985 [78] |
| hApo-Lf hHolo-Lf 10 µg/mL | Microbicidal effect. Lf participated in the internalization of amastigotes in macrophages and stimulated respiratory burst. | Lima, M. F. and. Kierszenbaum, F. 1987 [79] | |
| Malaria/ Plasmodium falciparum | hApo-Lf hHolo-Lf 30 µM | Inhibition of Plasmodium depended on iron deprivation as well as on the generation of oxygen-free radicals that damaged the membrane. | Fritsch, G., et al., 1987 [78] |
| Heparinase 10 mU/mL, anti-LRP 10 µg/mL, GST-RAP 150 µg/mL, Lf 400 µg/mL | They all cause inhibition. Only Lf interfered with sporozoite attachment by binding to both receptor-related proteins and heparan sulfate proteoglycans. | Shakibaei, M. and U. Frevert, 1996 [87] | |
| Lf, Tf 1.6 µM ApoE, β3-VLDL apoE-enriched with 3-VLDL 250 µg/mL | Lf and lipoproteins inhibited the binding to liver cells by competition for the same binding sites as the CSA protein of sporozoites. No effect of 3-VLDL. | Sinnis, P., et al., 1996 [88] | |
| hLf, Lfcin 25–37 100 μg/mL KPSE peptide 394–409 of CD36 FASP peptide; 300–312 of CD36 | The inhibitory effects of Lf on P. falciparum–infected erythrocytes binding was a result of specific binding of Lf to CD36 and thrombospondin. No effect of KPSE peptide and FASP peptide. | Eda, S., et al., 1999 [89] | |
| Lf 50 µg/mL cathelicidin LL-37 peptide 50 µg/mL (in vivo naïve mice) | A 95% reduction in malaria infection was observed after treatment with Lf and 43% with cathelicidin. | Parra, M., et al., 2013 [90] | |
| Malaria/ Plasmodium berghei | BuLf, Nanoformulation with BuLf Chloroquine (in vivo, BALB/c mice) | BuLf and Nanoformulation with Lf showed lower parasitaemia, low inflammation in the spleen, free radical ion production, and higher survival tendency compared to the chloroquine group. | Anand N, et al., 2015 [91] |
| Babesiosis/ Babesia equi (Theileria equi) Babesia caballi. | Native-bLf bLf hydrolysate, bHolo-Lf bApo-Lf 2.5 and 5 mg/mL | Only Apo-Lf suppressed B. caballi. No effect on B. equi. | Ikadai, H., et al., 2005 [95] |
| Toxoplasmosis/Toxoplasma gondii | bLf, Lfcin B Lf C-terminal region 1000 µg/mL | Lfcin B produced an inhibitory effect and loss infectivity of parasites. | Tanaka, T., et al., 1995 [102] |
| Lfcin B, bLf Lf C-terminal region 1000 µg/mL | Parasiticidal effect and less penetration activity. Lfcin B was shown to be more effective than the other treatments. | Tanaka, T., et al., 1996 [103] | |
| Lfcin B 5 mg (In vivo, mice) | Lfcin orally administered caused survival and reduced the number of cysts in the brains of mice compared to the control group. | Isamida, T., et al., 1998 [106] | |
| Trichomoniasis/ Trichomonas gallinae | Nanoparticles with Lf and metronidazole, metronidazole 100 ug/mL (In vivo, pigeons 50 mg/kg) | 100% eradication of trophozoites with the nanoformulation. Pigeons treated with nanoparticles showed non-viable trophozoites in the crop lavage and all challenged animals recovered from the disease. | Tabari, M.A., et al., 2021 [112] |
| Primary amoebic meningoencephalitis/ Naegleria fowleri | Tritrypticin, Lf, killer decapeptide, scrambled peptide 100 µg/mL | Only tritrypticin showed a positive effect causing marked apoptosis. | Tiewcharoen, S., et al., 2014 [113] |
| Amoebic keratitis/ Acanthamoeba castellanii | Lf 1.48 mg/mL | No statistically significant difference was observed compared to the control treatment. | Alsam, S., et al., 2008 [114] |
| Apo-bLf, holo-bLf, native bLf 10 µM | Apo-bLf affected the viability of the parasite, caused trophozoite death and did not cause encystment. No effect with holo-bLf or native bLf. | Tomita et al. in 2017 [115] |
4. Effect of Lactoferrin on Free-Living Pathogenic Protozoa
4.1. Naegleria fowleri
4.2. Acanthamoeba castellanii
5. Conclusions
Funding
Conflicts of Interest
References
- Burchmore, R.J.; Barrett, M. Life in vacuoles—Nutrient acquisition by Leishmania amastigotes. Int. J. Parasitol. 2001, 31, 1311–1320. [Google Scholar] [CrossRef]
- Landfear, S.M. Nutrient Transport and Pathogenesis in Selected Parasitic Protozoa. Eukaryot. Cell 2011, 10, 483–493. [Google Scholar] [CrossRef] [PubMed]
- Rönnebäumer, K.; Gross, U.; Bohne, W.U. The Nascent Parasitophorous Vacuole Membrane of Encephalitozoon cuniculi Is Formed by Host Cell Lipids and Contains Pores which Allow Nutrient Uptake. Eukaryot. Cell 2008, 7, 1001–1008. [Google Scholar] [CrossRef] [PubMed]
- Blume, M.; Rodriguez-Contreras, D.; Landfear, S.; Fleige, T.; Soldati-Favre, D.; Lucius, R.; Gupta, N. Host-derived glucose and its transporter in the obligate intracellular pathogen Toxoplasma gondii are dispensable by glutaminolysis. Proc. Natl. Acad. Sci. USA 2009, 106, 12998–13003. [Google Scholar] [CrossRef]
- Azema, L.; Claustre, S.; Alric, I.; Blonski, C.; Willson, M.; Perié, J.; Baltz, T.; Tetaud, E.; Bringaud, F.; Cottem, D.; et al. Interaction of substituted hexose analogues with the Trypanosoma brucei hexose transporter. Biochem. Pharmacol. 2004, 67, 459–467. [Google Scholar] [CrossRef]
- Acosta-Smith, E.; Viveros-Jiménez, K.; Canizalez-Román, A.; Reyes-Lopez, M.; Bolscher, J.G.M.; Nazmi, K.; Flores-Villaseñor, H.; Alapizco-Castro, G.; de la Garza, M.; Martínez-Garcia, J.J.; et al. Bovine Lactoferrin and Lactoferrin-Derived Peptides Inhibit the Growth of Vibrio cholerae and Other Vibrio species. Front. Microbiol. 2017, 8, 2633. [Google Scholar] [CrossRef]
- Sibley, L.D. Invasion and intracellular survival by protozoan parasites. Immunol. Rev. 2011, 240, 72–91. [Google Scholar] [CrossRef]
- Lambert, H.; Barragan, A. Modelling parasite dissemination: Host cell subversion and immune evasion by Toxoplasma gondii. Cell. Microbiol. 2010, 12, 292–300. [Google Scholar] [CrossRef]
- Bellamy, W.; Takase, M.; Yamauchi, K.; Wakabayashi, H.; Kawase, K.; Tomita, M. Identification of the bactericidal domain of lactoferrin. Biochim. Biophys. Acta BBA-Protein Struct. Mol. Enzym. 1992, 1121, 130–136. [Google Scholar] [CrossRef]
- Gruden, Š.; Ulrih, N.P. Diverse Mechanisms of Antimicrobial Activities of Lactoferrins, Lactoferricins, and Other Lactoferrin-Derived Peptides. Int. J. Mol. Sci. 2021, 22, 11264. [Google Scholar] [CrossRef]
- Bruni, N.; Capucchio, M.T.; Biasibetti, E.; Pessione, E.; Cirrincione, S.; Giraudo, L.; Corona, A.; Dosio, F. Antimicrobial Activity of Lactoferrin-Related Peptides and Applications in Human and Veterinary Medicine. Molecules 2016, 21, 752. [Google Scholar] [CrossRef] [PubMed]
- Zarzosa-Moreno, D.; Avalos-Gómez, C.; Ramírez-Texcalco, L.S.; Torres-López, E.; Ramírez-Mondragón, R.; Hernández-Ramírez, J.O.; Serrano-Luna, J.; De La Garza, M. Lactoferrin and Its Derived Peptides: An Alternative for Combating Virulence Mechanisms Developed by Pathogens. Molecules 2020, 25, 5763. [Google Scholar] [CrossRef]
- Yamauchi, K.; Tomita, M.; Giehl, T.J.; Ellison, R.T. Antibacterial activity of lactoferrin and a pepsin-derived lactoferrin peptide fragment. Infect. Immun. 1993, 61, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Gifford, J.L.; Hunter, H.N.; Vogel, H.J. Lactoferricin: A lactoferrin-derived peptide with antimicrobial, antiviral, antitumor and immunological properties. Cell. Mol. Life Sci. 2005, 62, 2588–2598. [Google Scholar] [CrossRef] [PubMed]
- Ortíz-Estrada, G.; Luna-Castro, S.; Piña-Vázquez, C.; Samaniego-Barrón, L.; León-Sicairos, N.; Serrano-Luna, J.; de la Garza, M. Iron-saturated lactoferrin and pathogenic protozoa: Could this protein be an iron source for their parasitic style of life? Future Microbiol. 2012, 7, 149–164. [Google Scholar] [CrossRef] [PubMed]
- Saghaug, C.S.; Klotz, C.; Kallio, J.P.; Brattbakk, H.-R.; Stokowy, T.; Aebischer, T.; Kursula, I.; Langeland, N.; Hanevik, K. Genetic variation in metronidazole metabolism and oxidative stress pathways in clinical Giardia lamblia assemblage A and B isolates. Infect. Drug Resist. 2019, 12, 1221–1235. [Google Scholar] [CrossRef]
- Leitsch, D. A review on metronidazole: An old warhorse in antimicrobial chemotherapy. Parasitology 2019, 146, 1167–1178. [Google Scholar] [CrossRef]
- Krakovka, S.; Ribacke, U.; Miyamoto, Y.; Eckmann, L.; Svärd, S. Characterization of Metronidazole-Resistant Giardia intestinalis Lines by Comparative Transcriptomics and Proteomics. Front. Microbiol. 2022, 13, 834008. [Google Scholar] [CrossRef]
- Upcroft, P.; Upcroft, J.A. Drug Targets and Mechanisms of Resistance in the Anaerobic Protozoa. Clin. Microbiol. Rev. 2001, 14, 150–164. [Google Scholar] [CrossRef]
- Cudmore, S.L.; Delgaty, K.L.; Hayward-McClelland, S.F.; Petrin, D.P.; Garber, G.E. Treatment of Infections Caused by Metronidazole-Resistant Trichomonas vaginalis. Clin. Microbiol. Rev. 2004, 17, 783–793. [Google Scholar] [CrossRef] [PubMed]
- Monzote, L. Development of natural products as anti-parasitic agents. Curr. Clin. Pharmacol. 2014, 9, 181–186. [Google Scholar] [CrossRef]
- Christen, P.; Veuthey, J.L. New Trends in Extraction, Identification and Quantification of Artemisinin and its Derivatives. Curr. Med. Chem. 2001, 8, 1827–1839. [Google Scholar] [CrossRef]
- Duffey, M.; Blasco, B.; Burrows, J.N.; Wells, T.N.; Fidock, D.A.; Leroy, D. Assessing risks of Plasmodium falciparum resistance to select next-generation antimalarials. Trends Parasitol. 2021, 37, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; Ryan, U.M.; Xiao, L. Genetic Diversity and Population Structure of Cryptosporidium. Trends Parasitol. 2018, 34, 997–1011. [Google Scholar] [CrossRef]
- Wang, R.-J.; Li, J.-Q.; Chen, Y.-C.; Zhang, L.-X.; Xiao, L.-H. Widespread occurrence of Cryptosporidium infections in patients with HIV/AIDS: Epidemiology, clinical feature, diagnosis, and therapy. Acta Trop. 2018, 187, 257–263. [Google Scholar] [CrossRef]
- Janssen, B.; Snowden, J. Cryptosporidiosis. In StatPearls; Stat Pearls Publishing: Treasure Island, FL, USA, 2022. [Google Scholar]
- Murdock, C.A.; Matthews, K.R. Antibacterial activity of pepsin-digested lactoferrin on foodborne pathogens in buffered broth systems and ultra-high temperature milk with EDTA. J. Appl. Microbiol. 2002, 93, 850–856. [Google Scholar] [CrossRef]
- Carryn, S.; Schaefer, D.A.; Imboden, M.; Homan, E.J.; Bremel, R.D.; Riggs, M.W. Phospholipases and Cationic Peptides Inhibit Cryptosporidium parvum Sporozoite Infectivity by Parasiticidal and Non-Parasiticidal Mechanisms. J. Parasitol. 2012, 98, 199–204. [Google Scholar] [CrossRef]
- Paredes, J.L.; Sparks, H.; White, A.C., Jr.; Martinez-Traverso, G.; Ochoa, T.; Castellanos-González, A. Killing of Cryptosporidium sporozoites by Lactoferrin. Am. J. Trop. Med. Hyg. 2017, 97, 774–776. [Google Scholar] [CrossRef]
- Blake, D.P. Eimeria genomics: Where are we now and where are we going? Vet. Parasitol. 2015, 212, 68–74. [Google Scholar] [CrossRef]
- Arisue, N.; Hashimoto, T. Phylogeny and evolution of apicoplasts and apicomplexan parasites. Parasitol. Int. 2015, 64, 254–259. [Google Scholar] [CrossRef] [PubMed]
- Walker, R.A.; Ferguson, D.J.P.; Miller, C.M.D.; Smith, N.C. Sex and Eimeria: A molecular perspective. Parasitology 2013, 140, 1701–1717. [Google Scholar] [CrossRef] [PubMed]
- Burrell, A.; Tomley, F.M.; Vaughan, S.; Marugan-Hernandez, V. Life cycle stages, specific organelles and invasion mechanisms of Eimeria species. Parasitology 2020, 147, 263–278. [Google Scholar] [CrossRef] [PubMed]
- Olajide, J.S.; Qu, Z.; Yang, S.; Oyelade, O.J.; Cai, J. Eimeria proteins: Order amidst disorder. Parasites Vectors 2022, 15, 38. [Google Scholar] [CrossRef] [PubMed]
- Omata, Y.; Satake, M.; Maeda, R.; Saito, A.; Shimazaki, K.; Uzuka, Y.; Tanabe, S.; Sarashina, T.; Mikami, T.; Yamauchi, K. Reduction of the Infectivity of Toxoplasma gondii and Eimeria stiedai Sporozoites by Treatment with Bovine Lactoferricin. J. Vet. Med. Sci. 2001, 63, 187–190. [Google Scholar] [CrossRef] [PubMed]
- World Health Organization. Amoebiasis = Amibiase. Wkly. Epidemiol. Rec. Relev. Épidémiol. Hebd. 1997, 72, 97–99. [Google Scholar]
- Stanley, S.L., Jr. Amoebiasis. Lancet 2003, 361, 1025–1034. [Google Scholar] [CrossRef]
- Carrero, J.C.; Reyes-López, M.; Serrano-Luna, J.; Shibayama, M.; Unzueta, J.; León-Sicairos, N.; de la Garza, M. Intestinal amoebiasis: 160 years of its first detection and still remains as a health problem in developing countries. Int. J. Med. Microbiol. 2020, 310, 151358. [Google Scholar] [CrossRef]
- Marie, C.; Petri, W.A., Jr. Regulation of Virulence of Entamoeba histolytica. Annu. Rev. Microbiol. 2014, 68, 493–520. [Google Scholar] [CrossRef] [PubMed]
- Murray, M.J.; Murray, A.; Murray, C.J. The salutary effect of milk on amoebiasis and its reversal by iron. Br. Med. J. 1980, 280, 1351–1352. [Google Scholar] [CrossRef] [PubMed]
- Bray, R.S.; Garnham, P.C. Effect of Milk Diet on P. cynomolgi Infections in Monkeys. Br. Med. J. 1953, 1, 1200–1201. [Google Scholar] [CrossRef] [PubMed]
- Leboffe, L.; Giansanti, F.; Antonini, G. Antifungal and Antiparasitic Activities of Lactoferrin. Anti-Infect. Agents Med. Chem. 2009, 8, 114–127. [Google Scholar] [CrossRef]
- Latour, N.G.; Reeves, R.E. An iron-requirement for growth of Entamoeba histolytica in culture, and the antiamebal activity of 7-iodo-8-hydroxy-quinoline-5-sulfonic acid. Exp. Parasitol. 1965, 17, 203–209. [Google Scholar] [CrossRef]
- Serrano, J.J.; Arzola, N.J.; Reyes, M.; de la Garza, M.; Serrano-Luna, J. Iron and Entamoeba histolytica HM1:IMSS. In Proceedings of the IX International Congress of Parasitoly ICOPA IX Makuhari Messe, Chiba, Japan, 24–26 August 1998. [Google Scholar]
- Gillin, F.D.; Reiner, D.S.; Wang, C.-S. Human Milk Kills Parasitic Intestinal Protozoa. Science 1983, 221, 1290–1292. [Google Scholar] [CrossRef] [PubMed]
- León-Sicairos, N.; Reyes-López, M.; Ordaz-Pichardo, C.; de la Garza, M. Microbicidal action of lactoferrin and lactoferricin and their synergistic effect with metronidazole in Entamoeba histolytica. Biochem. Cell Biol. 2006, 84, 327–336. [Google Scholar] [CrossRef]
- Leitch, E.C.; Willcox, M.D. Synergic antistaphylococcal properties of lactoferrin and lysozyme. J. Med. Microbiol. 1998, 47, 837–842. [Google Scholar] [CrossRef] [PubMed]
- Benson, K.F.; Carter, S.G.; Patterson, K.M.; Patel, D.; Jensen, G.S. A novel extract from bovine colostrum whey supports anti-bacterial and anti-viral innate immune functions in vitro and in vivo: I. Enhanced immune activity in vitro translates to improved microbial clearance in animal infection models. Prev. Med. 2012, 54, S116–S123. [Google Scholar] [CrossRef]
- Kassim, O.O.; Ako-Anai, K.A.; Torimiro, S.E.; Hollowell, G.P.; Okoye, V.C.; Martin, S.K. Inhibitory factors in breastmilk, maternal and infant sera against in vitro growth of Plasmodium falciparum malaria parasite. J. Trop. Pediatr. 2000, 46, 92–96. [Google Scholar] [CrossRef]
- Mahdi, L.; Mahdi, N.; Al-Kakei, S.; Musafer, H.; Al-Joofy, I.; Essa, R.; Zwain, L.; Salman, I.; Mater, H.; Al-Alak, S.; et al. Treatment strategy by lactoperoxidase and lactoferrin combination: Immunomodulatory and antibacterial activity against multidrug-resistant Acinetobacter baumannii. Microb. Pathog. 2018, 114, 147–152. [Google Scholar] [CrossRef]
- León-Sicairos, N.; López-Soto, F.; Reyes-López, M.; Godínez-Vargas, D.; Ordaz-Pichardo, C.; de la Garza, M. Amoebicidal Activity of Milk, Apo-lactoferrin, sIgA and Lysozyme. Clin. Med. Res. 2006, 4, 106–113. [Google Scholar] [CrossRef]
- Bolscher, J.; Adao, R.; Nazmi, K.; VandenKeybus, P.; Vanthof, W.; Nieuwamerongen, A.; Bastos, M.; Veerman, E. Bactericidal activity of LFchimera is stronger and less sensitive to ionic strength than its constituent lactoferricin and lactoferrampin peptides. Biochimie 2009, 91, 123–132. [Google Scholar] [CrossRef]
- López-Soto, F.; Leon-Sicairos, N.; Nazmi, K.; Bolscher, J.G.; De La Garza, M. Microbicidal effect of the lactoferrin peptides Lactoferricin17–30, Lactoferrampin265–284, and Lactoferrin chimera on the parasite Entamoeba histolytica. BioMetals 2010, 23, 563–568. [Google Scholar] [CrossRef] [PubMed]
- Kuipers, M.E.; de Vries, H.G.; Eikelboom, M.C.; Meijer, D.K.F.; Swart, P.J. Synergistic Fungistatic Effects of Lactoferrin in Combination with Antifungal Drugs against Clinical Candida Isolates. Antimicrob. Agents Chemother. 1999, 43, 2635–2641. [Google Scholar] [CrossRef] [PubMed]
- Asgharpour, A.; Gilchrist, C.; Baba, D.; Hamano, S.; Houpt, E. Resistance to Intestinal Entamoeba histolytica Infection Is Conferred by Innate Immunity and Gr-1+ Cells. Infect. Immun. 2005, 73, 4522–4529. [Google Scholar] [CrossRef] [PubMed]
- León-Sicairos, N.; Martínez-Pardo, L.; Sánchez-Hernández, B.; de la Garza, M.; Carrero, J.C. Oral lactoferrin treatment resolves amoebic intracecal infection in C3H/HeJ mice. Biochem. Cell Biol. 2012, 90, 435–441. [Google Scholar] [CrossRef] [PubMed]
- León-Sicairos, N.; Ordaz-Pichardo, C.; Carrero, J.C.; de la Garza, M. Lactoferrin in the Battle against Intestinal Parasites: A Review. In Natural Remedies in the Fight against Parasites; IntechOpen: Rijeka, Croatia, 2017. [Google Scholar] [CrossRef]
- Ordaz-Pichardo, C.; León-Sicairos, N.; Hernández-Ramírez, V.I.; Talamás-Rohana, P.; de la Garza, M. Effect of bovine lactoferrin in a therapeutic hamster model of hepatic amoebiasis. Biochem. Cell Biol. 2012, 90, 425–434. [Google Scholar] [CrossRef]
- Díaz-Godínez, C.; González-Galindo, X.; Meza-Menchaca, T.; Bobes, R.J.; de la Garza, M.; León-Sicairos, N.; Laclette, J.P.; Carrero, J.C. Synthetic bovine lactoferrin peptide Lfampin kills Entamoeba histolytica trophozoites by necrosis and resolves amoebic intracecal infection in mice. Biosci. Rep. 2019, 39, BSR20180850. [Google Scholar] [CrossRef]
- Frontera, L.S.; Moyano, S.; Quassollo, G.; Lanfredi-Rangel, A.; Rópolo, A.S.; Touz, M.C. Lactoferrin and lactoferricin endocytosis halt Giardia cell growth and prevent infective cyst production. Sci. Rep. 2018, 8, 18020. [Google Scholar] [CrossRef]
- Gil, F.F.; Ventura, L.L.A.; Fonseca, J.F.; Saniago, H.C.; Busatti, H.; Santos, J.F.G.; Gomes, M.A. Hematological profile in natural progression of giardiasis: Kinetics of experimental infection in gerbils. J. Infect. Dev. Ctries. 2018, 12, 492–498. [Google Scholar] [CrossRef]
- Berkman, D.S.; Lescano, A.; Gilman, R.H.; Lopez, S.L.; Black, M.M. Effects of stunting, diarrhoeal disease, and parasitic infection during infancy on cognition in late childhood: A follow-up study. Lancet 2002, 359, 564–571. [Google Scholar] [CrossRef]
- Turchany, J.M.; Aley, S.B.; Gillin, F.D. Giardicidal activity of lactoferrin and N-terminal peptides. Infect. Immun. 1995, 63, 4550–4552. [Google Scholar] [CrossRef]
- Turchany, J.M.; Mccaffery, J.M.; Aley, S.B.; Gillin, F.D. Ultrastructural Effects of Lactoferrin Binding on Giardia lamblia Trophozoites. J. Eukaryot. Microbiol. 1997, 44, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Díaz, H.; Carrero, J.C.; Argüello-García, R.; Laclette, J.P.; Morales-Montor, J. Cyst and encystment in protozoan parasites: Optimal targets for new life-cycle interrupting strategies? Trends Parasitol. 2011, 27, 450–458. [Google Scholar] [CrossRef] [PubMed]
- Aguilar-Diaz, H.; Canizalez-Roman, A.; Nepomuceno-Mejia, T.; Gallardo-Vera, F.; Hornelas-Orozco, Y.; Nazmi, K.; Bolscher, J.G.; Carrero, J.C.; Leon-Sicairos, C.; Leon-Sicairos, N. Parasiticidal effect of synthetic bovine lactoferrin peptides on the enteric parasite Giardia intestinalis. Biochem. Cell Biol. 2017, 95, 82–90. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, T.J.; Chea-Woo, E.; Campos, M.; Pecho, I.; Prada, A.; McMahon, R.J.; Cleary, T.G. Impact of Lactoferrin Supplementation on Growth and Prevalence of Giardia Colonization in Children. Clin. Infect. Dis. 2008, 46, 1881–1883. [Google Scholar] [CrossRef] [PubMed]
- Steverding, D. The history of leishmaniasis. Parasites Vectors 2017, 10, 82. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Tovar, A.E.; Osorio-Trujillo, C.; Talamás-Rohana, P. Leishmaniosis. Rev. Cienc. Acad. Mex. Cienc. 2017, 68, 38–41. [Google Scholar]
- Dai, J.H.; Lee, Y.S.; Wong, H.C. Effects of iron limitation on production of a siderophore, outer membrane proteins, and hemolysin and on hydrophobicity, cell adherence, and lethality for mice of Vibrio parahaemolyticus. Infect. Immun. 1992, 60, 2952–2956. [Google Scholar] [CrossRef]
- Chang, K.-P.; Hendricks, L. Laboratory cultivation and maintenance of Leishmania. In Leishmaniasis (Human Parasitic Diseases); Elsevier: Amsterdam, The Netherlands, 1985; Volume 1, pp. 213–244. [Google Scholar]
- Wilson, M.E.; Vorhies, R.W.; Andersen, K.A.; Britigan, B.E. Acquisition of iron from transferrin and lactoferrin by the protozoan Leishmania chagasi. Infect. Immun. 1994, 62, 3262–3269. [Google Scholar] [CrossRef]
- Zarley, J.H.; Britigan, B.E.; Wilson, M. Hydrogen peroxide-mediated toxicity for Leishmania donovani chagasi promastigotes. Role of hydroxyl radical and protection by heat shock. J. Clin. Investig. 1991, 88, 1511–1521. [Google Scholar] [CrossRef]
- Wilson, M.E.; Lewis, T.S.; Miller, M.A.; McCormick, M.L.; Britigan, B.E. Leishmania chagasi: Uptake of iron bound to lactoferrin or transferrin requires an iron reductase. Exp. Parasitol. 2002, 100, 196–207. [Google Scholar] [CrossRef]
- Silva, T.; Abengózar, M.; Fernández-Reyes, M.; Andreu, D.; Nazmi, K.; Bolscher, J.G.M.; Bastos, M.; Rivas, L. Enhanced leishmanicidal activity of cryptopeptide chimeras from the active N1 domain of bovine lactoferrin. Amino Acids 2012, 43, 2265–2277. [Google Scholar] [CrossRef] [PubMed]
- Asthana, S.; Gupta, P.K.; Jaiswal, A.K.; Dube, A.; Chourasia, M.K. Targeted chemotherapy of visceral leishmaniasis by lactoferrin-appended amphotericin B-loaded nanoreservoir: In vitro and in vivo studies. Nanomedicine 2015, 10, 1093–1109. [Google Scholar] [CrossRef] [PubMed]
- Halder, A.; Shukla, D.; Das, S.; Roy, P.; Mukherjee, A.; Saha, B. Lactoferrin-modified Betulinic Acid-loaded PLGA nanoparticles are strong anti-leishmanials. Cytokine 2018, 110, 412–415. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.F.; Beltz, L.A.; Kierszenbaum, F. Trypanosoma cruzi: A Specific Surface Marker for the Amastigote Form. J. Protozool. 1988, 35, 108–110. [Google Scholar] [CrossRef] [PubMed]
- Lima, M.F.; Kierszenbaum, F. Lactoferrin effects on phagocytic cell function. I. Increased uptake and killing of an intracellular parasite by murine macrophages and human monocytes. J. Immunol. 1985, 134, 4176–4183. [Google Scholar]
- Lima, M.F.; Kierszenbaum, F. Lactoferrin effects of phagocytic cell function. II. The presence of iron is required for the lactoferrin molecule to stimulate intracellular killing by macrophages but not to enhance the uptake of particles and microorganisms. J. Immunol. 1987, 139, 1647–1651. [Google Scholar]
- Hansen, N.E.; Karle, H.; Andersen, V.; Malmquist, J.; Hoff, G.E. Neutrophilic granulocytes in acute bacterial infection. Sequential studies on lysozyme, myeloperoxidase and lactoferrin. Clin. Exp. Immunol. 1976, 26, 463–468. [Google Scholar]
- Tanaka, T.; Abe, Y.; Inoue, N.; Kim, W.-S.; Kumura, H.; Nagasawa, H.; Igarashi, I.; Shimazaki, K.-I. The Detection of Bovine Lactoferrin Binding Protein on Trypanosoma brucei. J. Vet. Med. Sci. 2004, 66, 619–625. [Google Scholar] [CrossRef]
- Mendis, K.; Rietveld, A.; Warsame, M.; Bosman, A.; Greenwood, B.; Wernsdorfer, W.H. From malaria control to eradication: The WHO perspective. Trop. Med. Int. Health 2009, 14, 802–809. [Google Scholar] [CrossRef]
- Foth, B.J.; Zhang, N.; Chaal, B.K.; Sze, S.K.; Preiser, P.R.; Bozdech, Z. Quantitative Time-course Profiling of Parasite and Host Cell Proteins in the Human Malaria Parasite Plasmodium falciparum. Mol. Cell. Proteom. 2011, 10, M110006411. [Google Scholar] [CrossRef]
- Soulard, V.; Bosson-Vanga, H.; Lorthiois, A.; Roucher, C.; Franetich, J.F.; Zanghi, G.; Bordessoulles, M.; Tefit, M.; Thellier, M.; Morosan, S.; et al. Plasmodium falciparum full life cycle and Plasmodium ovale liver stages in humanized mice. Nat. Commun. 2015, 6, 7690. [Google Scholar] [CrossRef] [PubMed]
- Fritsch, G.; Sawatzki, G.; Treumer, J.; Jung, A.; Spira, D.T. Plasmodium falciparum: Inhibition in vitro with lactoferrin, desferriferrithiocin, and desferricrocin. Exp. Parasitol. 1987, 63, 1–9. [Google Scholar] [CrossRef]
- Shakibaei, M.; Frevert, U. Dual interaction of the malaria circumsporozoite protein with the low density lipoprotein receptor-related protein (LRP) and heparan sulfate proteoglycans. J. Exp. Med. 1996, 184, 1699–1711. [Google Scholar] [CrossRef]
- Sinnis, P.; Willnow, T.E.; Briones, M.R.; Herz, J.; Nussenzweig, V. Remnant lipoproteins inhibit malaria sporozoite invasion of hepatocytes. J. Exp. Med. 1996, 184, 945–954. [Google Scholar] [CrossRef] [PubMed]
- Eda, S.; Eda, K.; Prudhomme, J.G.; Sherman, I.W. Inhibitory activity of human lactoferrin and its peptide on chondroitin sulfate A-, CD36-, and thrombospondin-mediated cytoadherence of Plasmodium falciparum-infected erythrocytes. Blood 1999, 94, 326–332. [Google Scholar] [CrossRef]
- Parra, M.; Liu, X.; Derrick, S.C.; Yang, A.; Tian, J.; Kolibab, K.; Kumar, S.; Morris, S.L. Molecular Analysis of Non-Specific Protection against Murine Malaria Induced by BCG Vaccination. PLoS ONE 2013, 8, e66115. [Google Scholar] [CrossRef]
- Anand, N.; Kanwar, R.K.; Sehgal, R.; Kanwar, J.R. Antiparasitic and immunomodulatory potential of oral nanocapsules encapsulated lactoferrin protein against Plasmodium berghei. Nanomedicine 2016, 11, 47–62. [Google Scholar] [CrossRef]
- Skrabalo, Z.; Deanovic, Z. Piroplasmosis in man; report of a case. Doc. Med. Geogr. Trop. 1957, 9, 11–16. [Google Scholar]
- Montero, E.; Gray, J.; Lobo, C.A.; González, L.M. Babesia and Human Babesiosis. Pathogens 2022, 11, 399. [Google Scholar] [CrossRef]
- Elsworth, B.; Duraisingh, M.T. A framework for signaling throughout the life cycle of Babesia species. Mol. Microbiol. 2021, 115, 882–890. [Google Scholar] [CrossRef]
- Ikadai, H.; Tanaka, T.; Igarashi, I.; Oyamada, T.; Matsuu, A.; Kudo, N.; Shimazaki, K.-I.; Shibahara, N.; Tanaka, H. Inhibitory effect of lactoferrin on in vitro growth of Babesia caballi. Am. J. Trop. Med. Hyg. 2005, 73, 710–712. [Google Scholar] [CrossRef] [PubMed]
- Köksal, Z.S.; Yanik, K.; Bilgin, K.; Yılmaz, E.M.; Hokelek, M. In Vivo Efficacy of Drugs against Toxoplasma gondii Combined with Immunomodulators. Jpn. J. Infect. Dis. 2016, 69, 113–117. [Google Scholar] [CrossRef] [PubMed]
- Jones, T.C.; Yeh, S.; Hirsch, J.G. The interaction between Toxoplasma gondii and mammalian cells. I. Mechanism of entry and intracellular fate of the parasite. J. Exp. Med. 1972, 136, 1157–1172. [Google Scholar] [CrossRef]
- Vidigal, P.V.T.; Santos, D.V.V.; Castro, F.C.; Couto, J.C.D.F.; Vitor, R.W.D.A.; Filho, G.B. Prenatal toxoplasmosis diagnosis from amniotic fluid by PCR. Rev. Soc. Bras. Med. Trop. 2002, 35, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Fachado, A.; Fonte, L.; Alberti, E.; Hadad, P.; Fonseca, L.; Machin, R.; Finlay, C. Usefulness of the detection of Toxoplasma gondii antigens in AIDS patients. Rev. Inst. Med. Trop. Sao Paulo 1994, 36, 525–529. [Google Scholar] [CrossRef] [PubMed]
- Gómez, J.E. Guía de práctica clínica para toxoplasmosis durante el embarazo y toxoplasmosis congénita en Colombia. Infectio 2007, 11, 129–141. [Google Scholar]
- Ellison, R.T., 3rd; Giehl, T.J.; LaForce, F.M. Damage of the outer membrane of enteric gram-negative bacteria by lactoferrin and transferrin. Infect. Immun. 1988, 56, 2774–2781. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Omata, Y.; Saito, A.; Shimazaki, K.; Yamauchi, K.; Takase, M.; Kawase, K.; Igarashi, I.; Suzuki, N. Toxoplasma gondii: Parasiticidal Effects of Bovine Lactoferricin against Parasites. Exp. Parasitol. 1995, 81, 614–617. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, T.; Omata, Y.; Narisawa, M.; Saito, A.; Shimazaki, K.; Igarashi, I.; Hirumi, H.; Suzuki, N. Growth inhibitory effect of bovine lactoferrin on Toxoplasma gondii tachyzoites in murine macrophages: Role of radical oxygen and inorganic nitrogen oxide in Toxoplasma growth-inhibitory activity. Vet. Parasitol. 1997, 68, 27–33. [Google Scholar] [CrossRef]
- Dzitko, K.; Dziadek, B.; Dziadek, J.; Długońska, H. Toxoplasma gondii: Inhibition of the intracellular growth by human lactoferrin. Pol. J. Microbiol. 2007, 56, 25–32. [Google Scholar] [PubMed]
- Tanaka, T.; Omata, Y.; Saito, A.; Shimazaki, K.; Igarashi, I.; Suzuki, N. Growth Inhibitory Effects of Bovine Lactoferrin to Toxoplasma gondii Parasites in Murine Somatic Cells. J. Vet. Med. Sci. 1996, 58, 61–65. [Google Scholar] [CrossRef] [PubMed]
- Isamida, T.; Tanaka, T.; Omata, Y.; Yamauchi, K.; Shimazaki, K.-I.; Saito, A. Protective Effect of Lactoferricin against Toxoplasma gondii Infection in Mice. J. Vet. Med Sci. 1998, 60, 241–244. [Google Scholar] [CrossRef] [PubMed]
- McClelland, R.S. Trichomonas vaginalis Infection: Can We Afford to Do Nothing? J. Infect. Dis. 2008, 197, 487–489. [Google Scholar] [CrossRef] [PubMed]
- Petrin, D.; Delgaty, K.; Bhatt, R.; Garber, G. Clinical and Microbiological Aspects of Trichomonas vaginalis. Clin. Microbiol. Rev. 1998, 11, 300–317. [Google Scholar] [CrossRef] [PubMed]
- Schwebke, J.R.; Burgess, D. Trichomoniasis. Clin. Microbiol. Rev. 2004, 17, 794–803. [Google Scholar] [CrossRef]
- Peterson, K.M.; Alderete, J.F. Iron uptake and increased intracellular enzyme activity follow host lactoferrin binding by Trichomonas vaginalis receptors. J. Exp. Med. 1984, 160, 398–410. [Google Scholar] [CrossRef] [PubMed]
- Lehker, M.W.; Alderete, J.F. Iron regulates growth of Trichomonas vaginalis and the expression of immunogenic trichomonad proteins. Mol. Microbiol. 1992, 6, 123–132. [Google Scholar] [CrossRef]
- Tabari, M.A.; Poźniak, B.; Abrishami, A.; Moradpour, A.A.; Shahavi, M.H.; Kazemi, S.; Youssefi, M.R. Antitrichomonal activity of metronidazole-loaded lactoferrin nanoparticles in pigeon trichomoniasis. Parasitol. Res. 2021, 120, 3263–3272. [Google Scholar] [CrossRef] [PubMed]
- Tiewcharoen, S.; Phurttikul, W.; Rabablert, J.; Auewarakul, P.; Roytrakul, S.; Chetanachan, P.; Atithep, T.; Junnu, V. Effect of synthetic antimicrobial peptides on Naegleria fowleri trophozoites. Southeast Asian J. Trop. Med. Public Health 2014, 45, 537–546. [Google Scholar]
- Alsam, S.; Jeong, S.R.; Dudley, R.; Khan, N.A. Role of human tear fluid in Acanthamoeba interactions with the human corneal epithelial cells. Int. J. Med. Microbiol. 2008, 298, 329–336. [Google Scholar] [CrossRef] [PubMed]
- Tomita, S.; Suzuki, C.; Wada, H.; Nomachi, M.; Imayasu, M.; Araki-Sasaki, K. Effects of lactoferrin on the viability and the encystment of Acanthamoeba trophozoites. Biochem. Cell Biol. 2017, 95, 48–52. [Google Scholar] [CrossRef]
- Król-Turmińska, K.; Olender, A. Human infections caused by free-living amoebae. Ann. Agric. Environ. Med. 2017, 24, 254–260. [Google Scholar] [CrossRef] [PubMed]
- Ong, T.Y.Y.; Khan, N.A.; Siddiqui, R. Brain-Eating Amoebae: Predilection Sites in the Brain and Disease Outcome. J. Clin. Microbiol. 2017, 55, 1989–1997. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Castillo, M.; Cárdenas-Guerra, R.E.; Arroyo, R.; Debnath, A.; Rodríguez, M.A.; Sabanero, M.; Flores-Sánchez, F.; Navarro-Garcia, F.; Serrano-Luna, J.; Shibayama, M. Nf-GH, a glycosidase secreted by Naegleria fowleri, causes mucin degradation: An in vitro and in vivo study. Future Microbiol. 2017, 12, 781–799. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Castillo, M.; Ramírez-Rico, G.; Serrano-Luna, J.; Shibayama, M. Iron-Binding Protein Degradation by Cysteine Proteases of Naegleria fowleri. BioMed Res. Int. 2015, 2015, 416712. [Google Scholar] [CrossRef] [PubMed]
- Güémez, A.; García, E. Primary Amoebic Meningoencephalitis by Naegleria fowleri: Pathogenesis and Treatments. Biomolecules 2021, 11, 1320. [Google Scholar] [CrossRef] [PubMed]
- Haney, E.F.; Nazmi, K.; Bolscher, J.G.; Vogel, H.J. Structural and biophysical characterization of an antimicrobial peptide chimera comprised of lactoferricin and lactoferrampin. Biochim. Biophys. Acta BBA-Biomembr. 2012, 1818, 762–775. [Google Scholar] [CrossRef] [PubMed]
- Khan, N.A. Acanthamoeba: Biology and increasing importance in human health. FEMS Microbiol. Rev. 2006, 30, 564–595. [Google Scholar] [CrossRef] [PubMed]
- Fanselow, N.; Sirajuddin, N.; Yin, X.-T.; Huang, A.; Stuart, P. Acanthamoeba keratitis, Pathology, Diagnosis and Treatment. Pathogens 2021, 10, 323. [Google Scholar] [CrossRef]
- Siddiqui, R.; Khan, N.A. Biology and pathogenesis of Acanthamoeba. Parasites Vectors 2012, 5, 6. [Google Scholar] [CrossRef]
- Ramírez-Rico, G.; Martínez-Castillo, M.; de la Garza, M.; Shibayama, M.; Serrano-Luna, J. Acanthamoeba castellanii Proteases are Capable of Degrading Iron-Binding Proteins as a Possible Mechanism of Pathogenicity. J. Eukaryot. Microbiol. 2015, 62, 614–622. [Google Scholar] [CrossRef] [PubMed]


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Reyes-López, M.; Ramírez-Rico, G.; Serrano-Luna, J.; de la Garza, M. Activity of Apo-Lactoferrin on Pathogenic Protozoa. Pharmaceutics 2022, 14, 1702. https://doi.org/10.3390/pharmaceutics14081702
Reyes-López M, Ramírez-Rico G, Serrano-Luna J, de la Garza M. Activity of Apo-Lactoferrin on Pathogenic Protozoa. Pharmaceutics. 2022; 14(8):1702. https://doi.org/10.3390/pharmaceutics14081702
Chicago/Turabian StyleReyes-López, Magda, Gerardo Ramírez-Rico, Jesús Serrano-Luna, and Mireya de la Garza. 2022. "Activity of Apo-Lactoferrin on Pathogenic Protozoa" Pharmaceutics 14, no. 8: 1702. https://doi.org/10.3390/pharmaceutics14081702