
In Vitro and In Silico Potential Inhibitory Effects of New Biflavonoids from Ochna rhizomatosa on HIV-1 Integrase and Plasmodium falciparum

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Materials Collection
2.2. General Experimental Procedures
2.3. Extraction of Plant Materials and Isolation of Compounds
2.4. Biological Activities
2.4.1. Evaluation of Anti-HIV Integrase Assay
2.4.2. Evaluation of Antiplasmodial Assay
2.4.3. Molecular Docking
2.4.4. Statistical Analysis
3. Results and Discussion
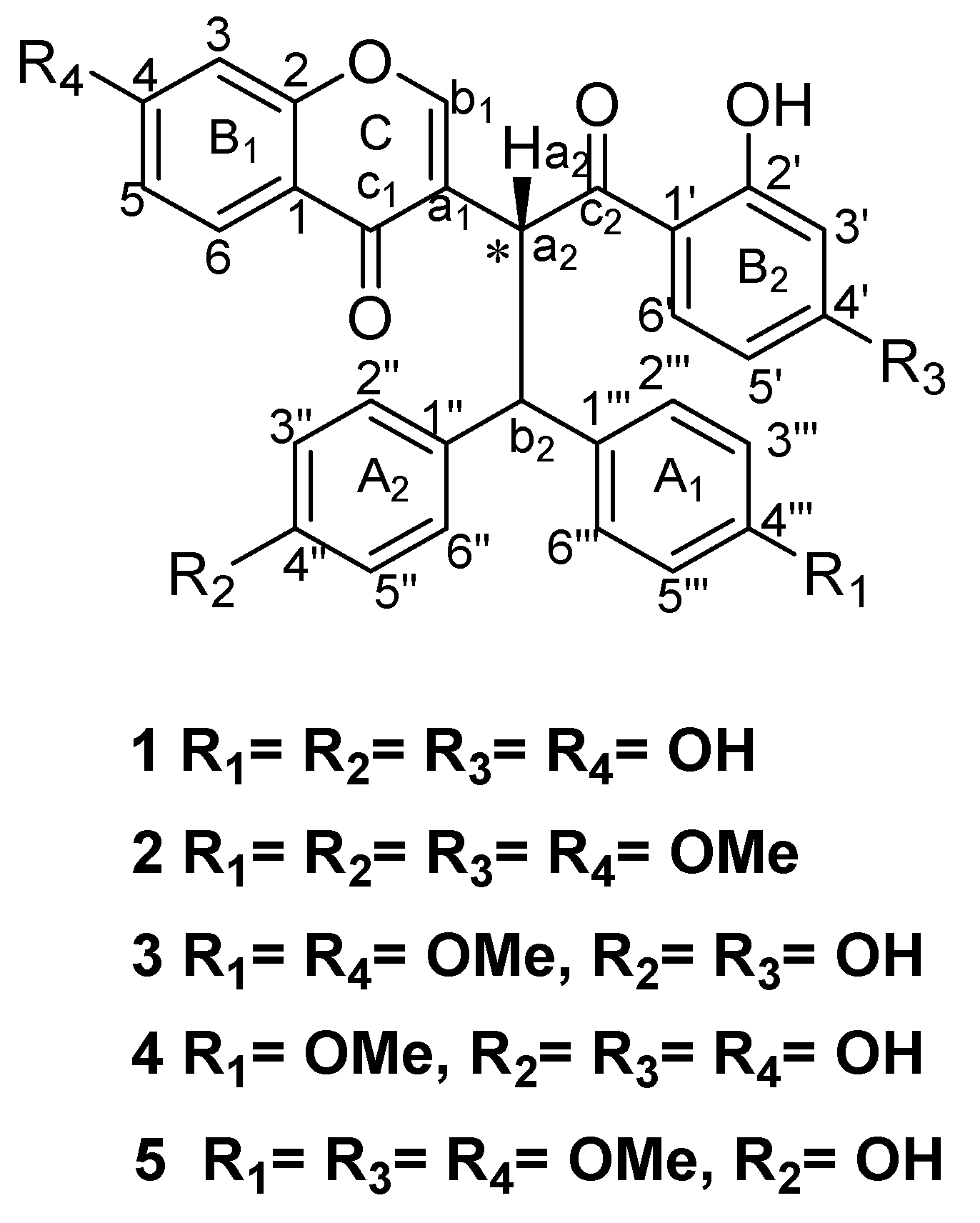
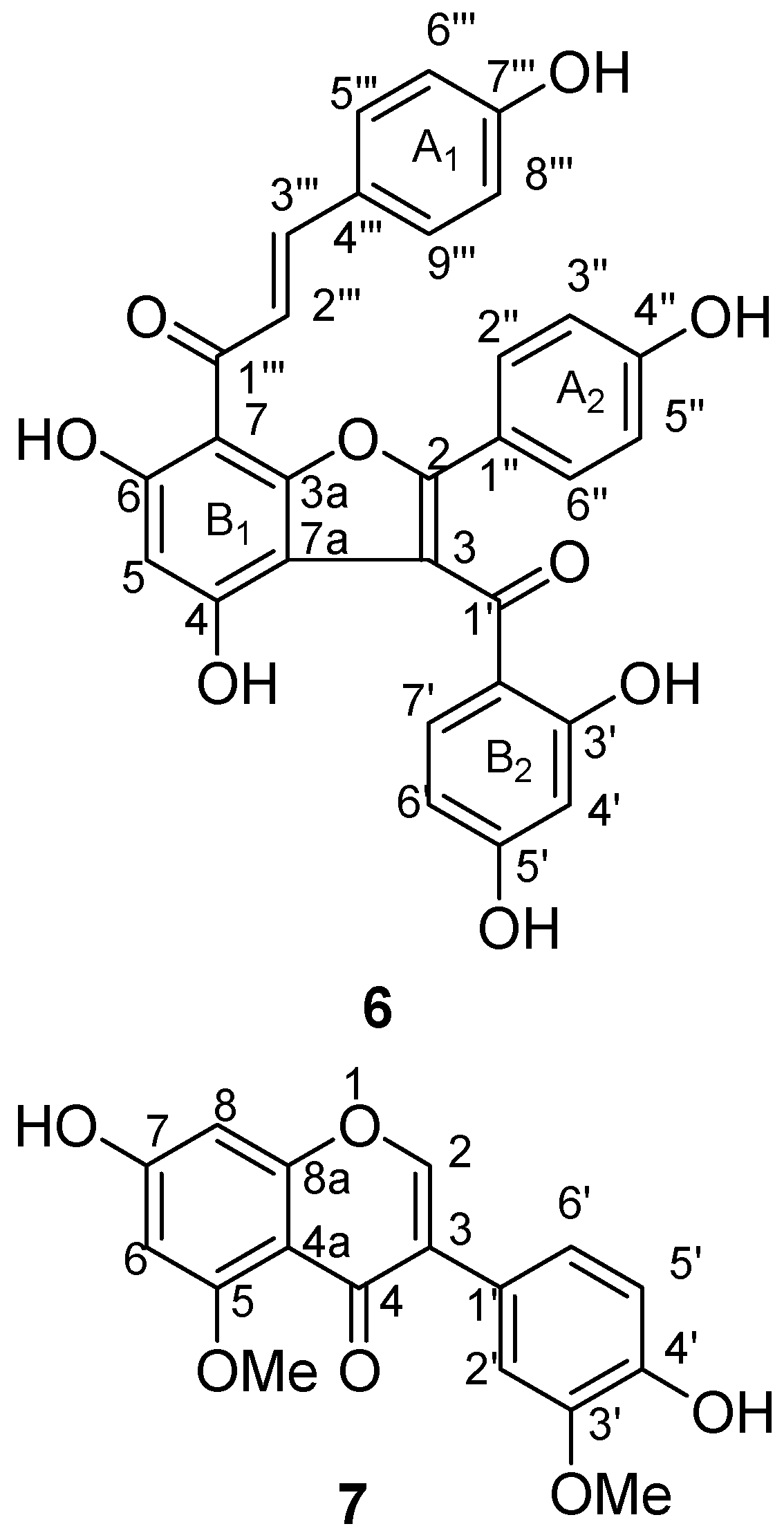
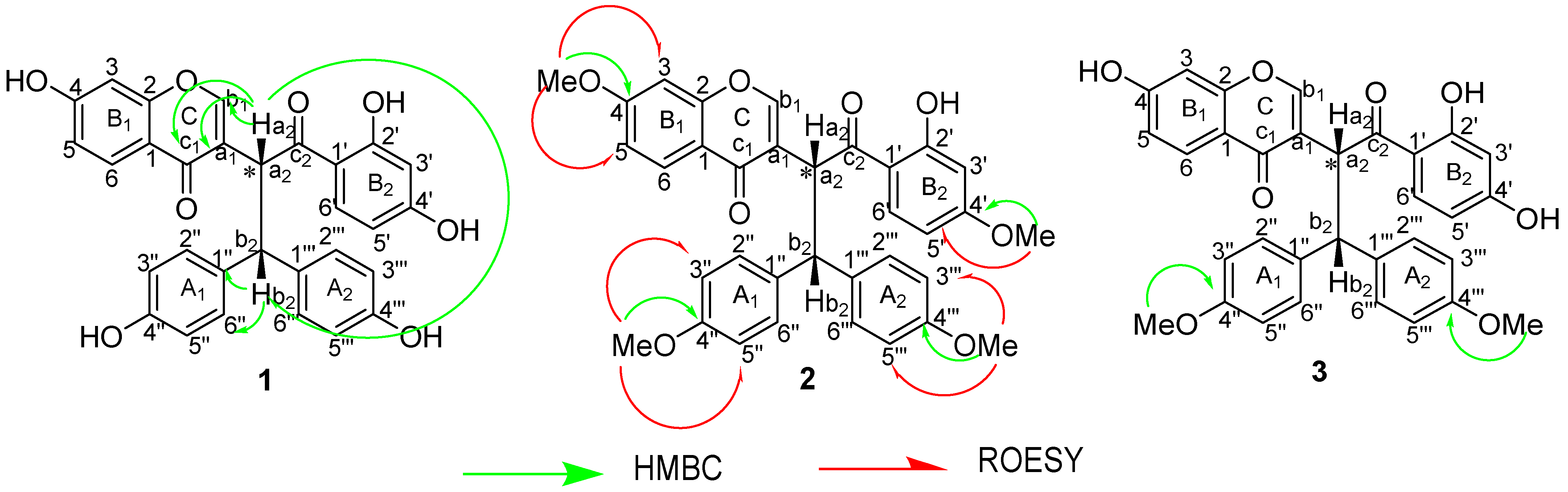
3.1. Identification of the Isolated Compounds
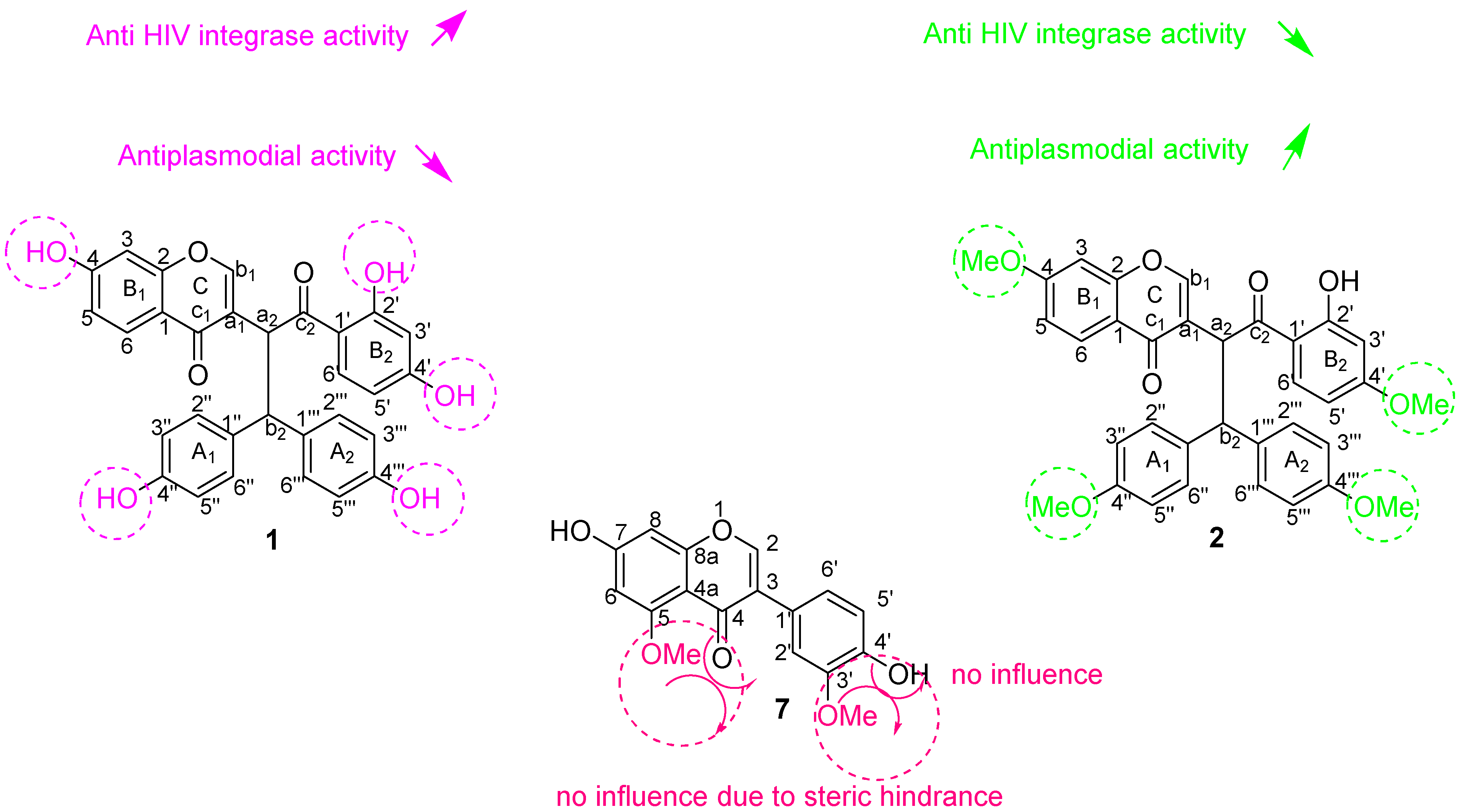
3.2. Biological Activities of Isolated Biflavonoids
3.2.1. Anti-HIV-1 Integrase Activity
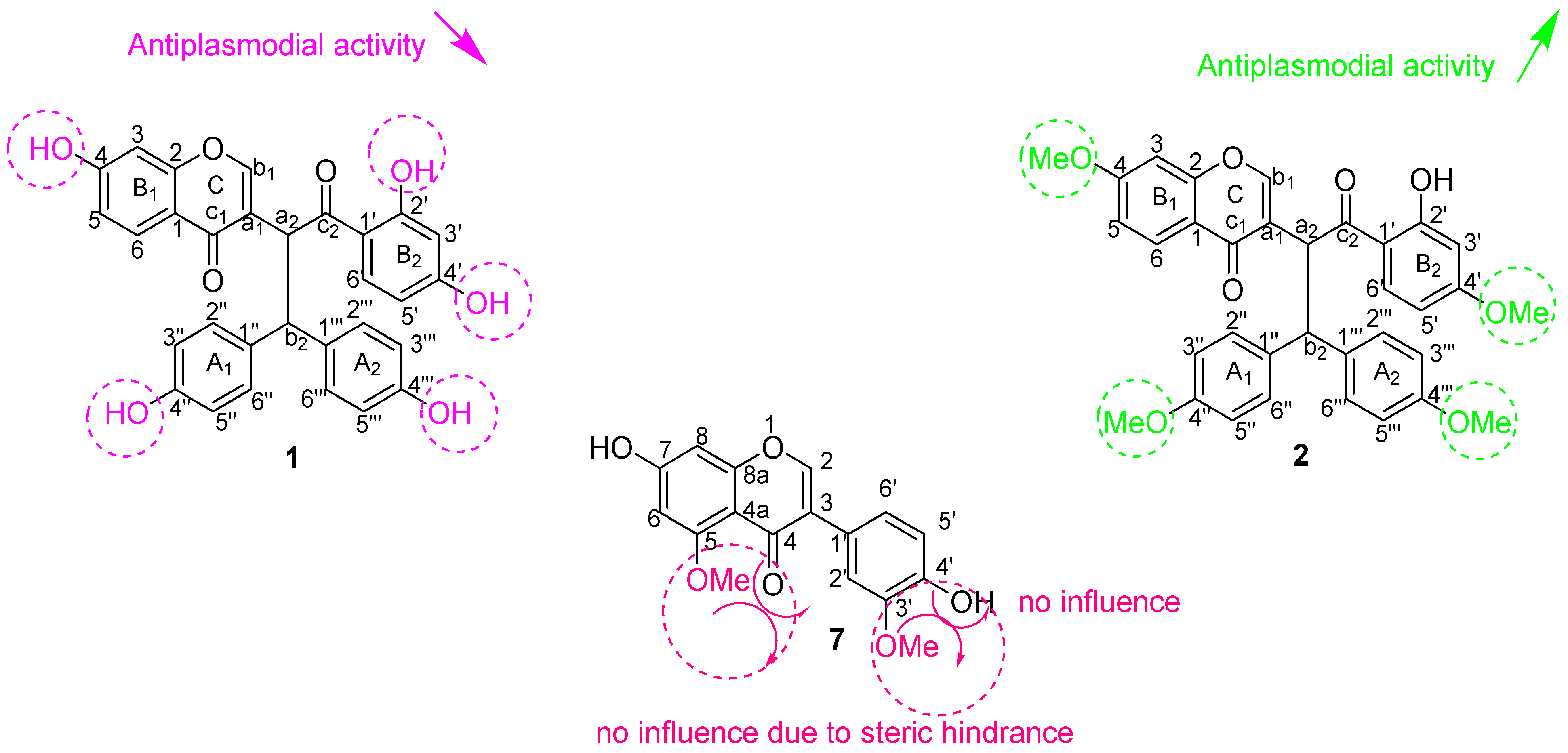
3.2.2. Antiplasmodial Activity
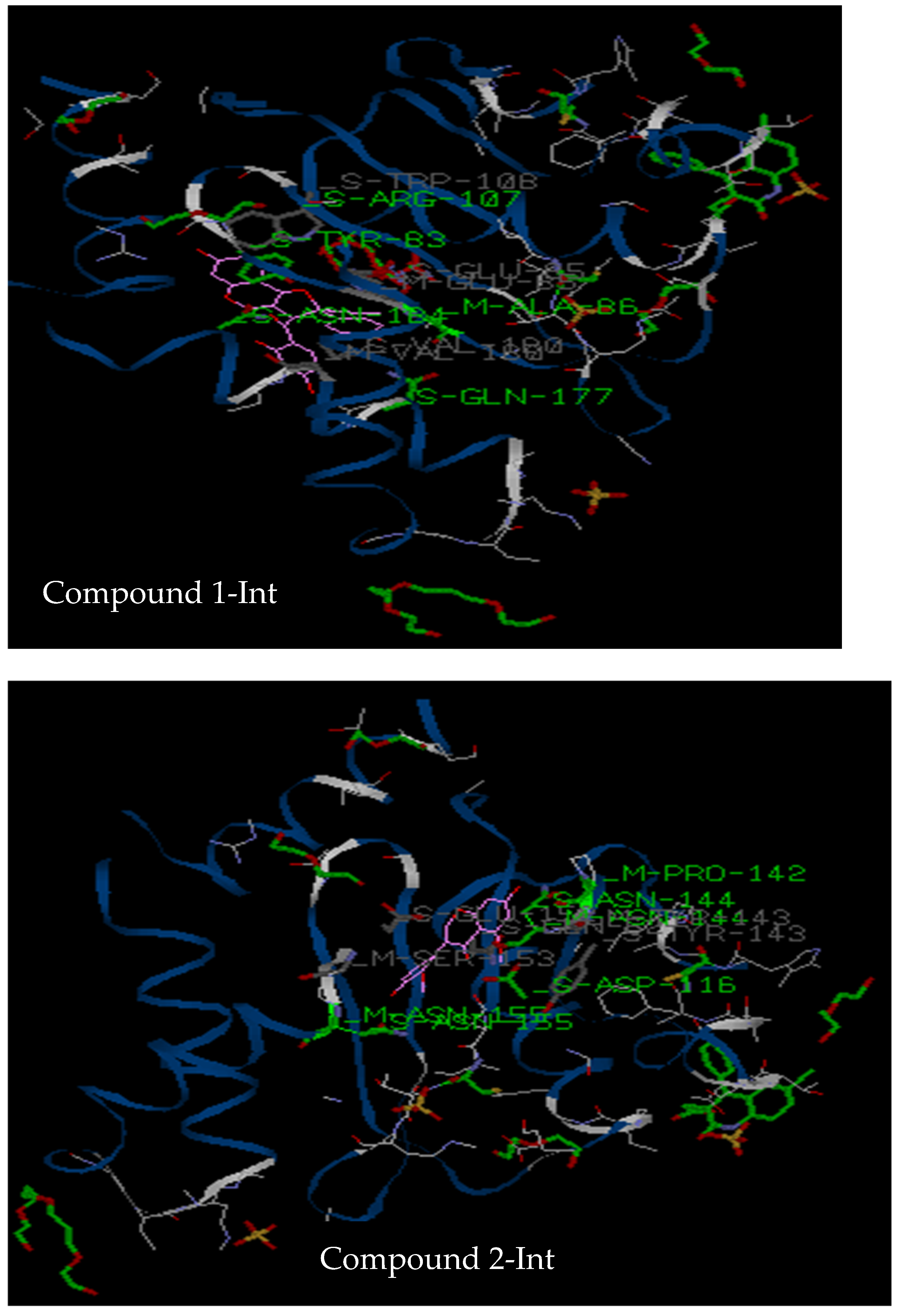
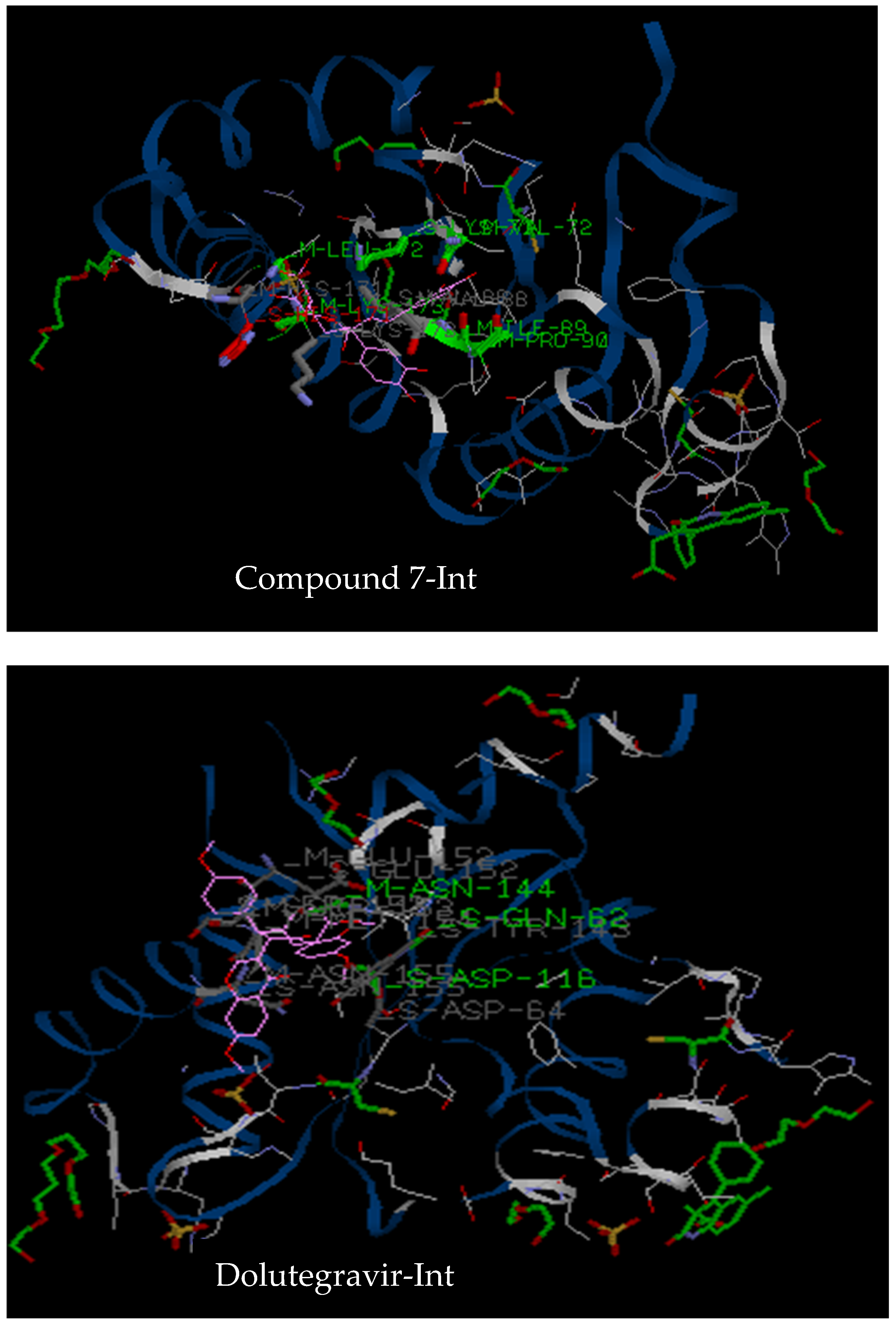
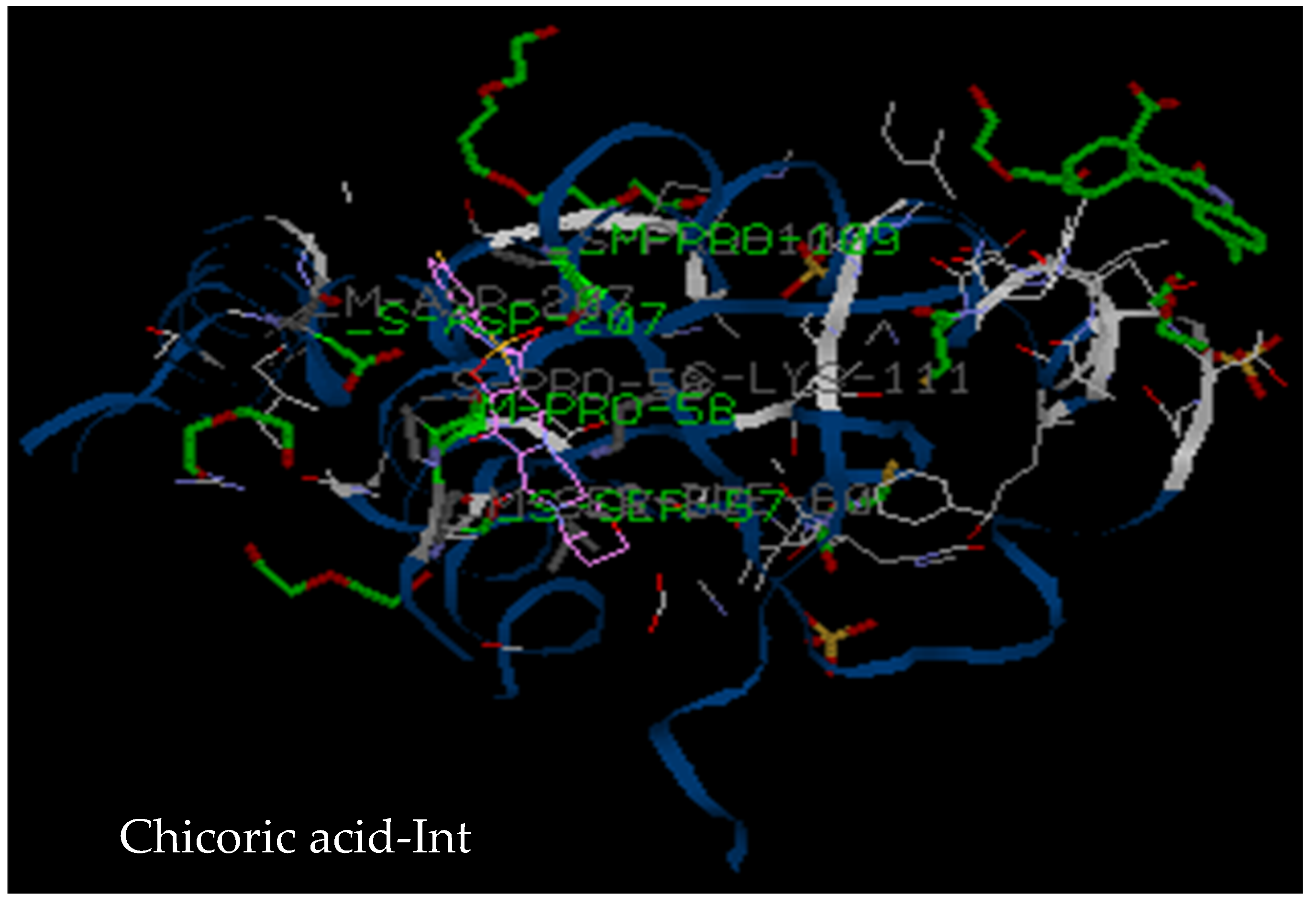
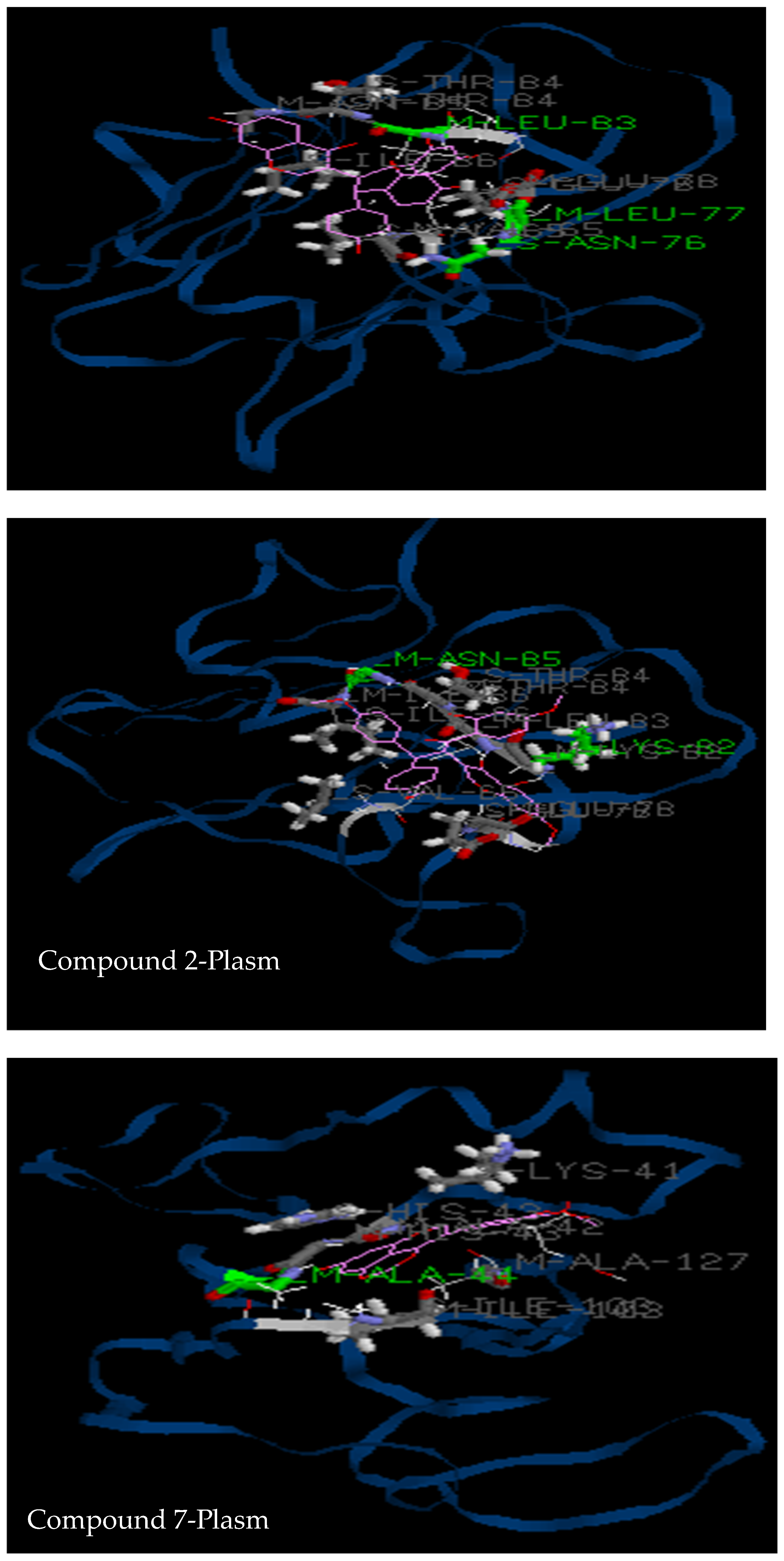
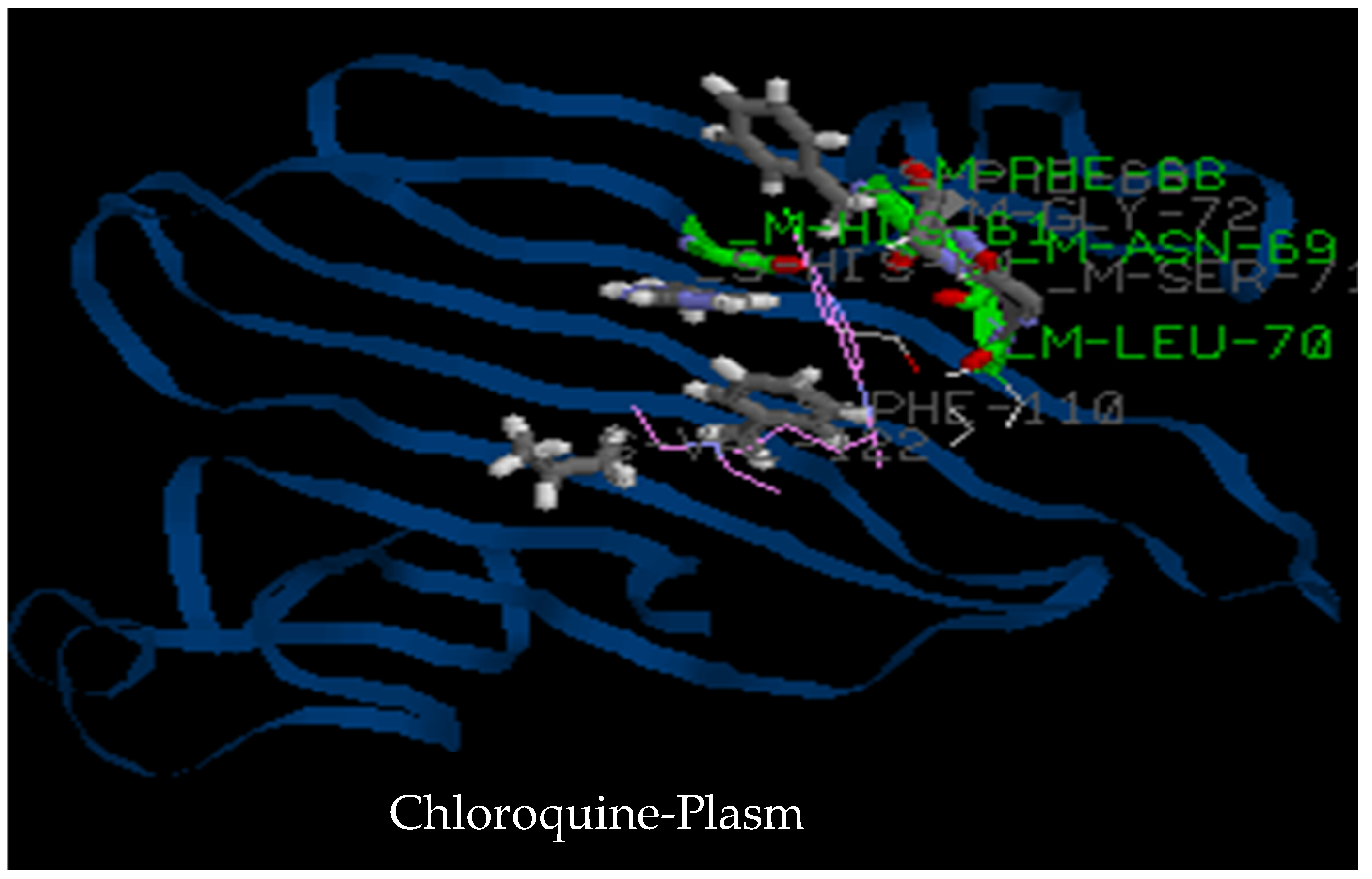
3.2.3. Molecular Docking
4. Conclusions and Recommendations
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kamya, M.R.; Gasasira, A.F.; Yeka, A.; Bakyaita, N.; Nsobya, S.L.; Francis, D.; Rosenthal, P.J.; Dorsey, G.; Havlir, D. Effect of HIV-1 Infection on Antimalarial Treatment Outcomes in Uganda: A Population-Based Study. J. Infect Dis. 2006, 193, 9–15. [Google Scholar] [CrossRef]
- United States Agency for International Development. U.S. President’s Malaria Initiative. USAID: USA, 2020. Available online: https://www.cdc.gov/malaria/malaria_worldwide/cdc_activities/pmi.html (accessed on 27 June 2021).
- World Health Organization. Cameroon: Malaria Kills 3000 People in 2018. Published on 26 April 2019 at 16h21 by the Ministry of Public Health. Cameroon. 2019. Available online: https://www.journalducameroun.com/en/cameroon-malaria-kills-3000-people-in-2018 (accessed on 20 July 2022).
- World Health Organization. Guidelines for the Treatment of Malaria. WHO: Geneva, 2018. Available online: https://www.who.int/malaria/publications/atoz/9789241549127 (accessed on 20 July 2022).
- Ariey, F.; Witkowski, B.; Amaratunga, C.; Beghain, J.; Langlois, A.C.; Khim, S.; Khim, N.; Duru, V.; Bouchier, C.; Ma, L.; et al. A molecular marker of artemisinin resistant Plasmodium falciparum malaria. Nature 2014, 505, 50–55. [Google Scholar] [CrossRef]
- UNAIDS. Report on the Global Aids Epidemics Update. Geneva, UNAIDS. 2019. Available online: https://gapwatch.org/news/unaids-2019-annual-update/1474 (accessed on 20 July 2022).
- UNAIDS. Global HIV & AIDS Statistics Report-2019 Fact Sheet. UNAIDS: Cameroun, 2019. Available online: https://www.unaids.org/en/resources/fact-sheet (accessed on 20 July 2022).
- Subbaraman, R.; Chaguturu, S.K.; Mayer, K.H.; Flanigan, T.P.; Kumarasamy, N. Adverse effects of highly active antiretroviral therapy in developing countries. Clin. Infect. Dis. 2007, 45, 1093–1101. [Google Scholar] [CrossRef]
- Reutrakul, V.; Ningnuek, N.; Pohmakotr, M.; Yoosook, C.; Napaswad, C.; Kasisit, J.; Santisuk, T.; Tuchinda, P. Anti HIV-1 flavonoid glycosides from Ochna integerrima. Planta Med. 2007, 73, 683–688. [Google Scholar] [CrossRef]
- Rahman, N.; Riaz, M.; Desai, U.R. Synthesis of biologically relevant bioflavonoids a review. Chem. Biodivers. 2007, 4, 2495–2527. [Google Scholar] [CrossRef]
- Dhooghe, L.; Maregesi, S.; Mincheva, I.; Ferreira, D.; Marais, J.P.J.; Lemière, F.; Matheeussen, A.; Cos, P.; Maes, L.; Vlietinck, A.; et al. Antiplasmodial activity of (I-3, II-3)-biflavonoids and other constituents from Ormocarpum kirkii. Phytochemistry 2010, 71, 785–791. [Google Scholar] [CrossRef]
- Messi, A.N.; Mbing, J.N.; Ndongo, J.T.; Nyegue, M.A.; Tiabou Tchinda, A.; Ladoh Yemeda, F.; Frédérich, M.; Pegnyemb, D.E. Phenolic compounds from the roots of Ochna schweinfurthiana and their antioxidant and antiplasmodial activities. Phytochem. Lett. 2016, 17, 119–125. [Google Scholar] [CrossRef]
- Ren, D.; Meng, F.C.; Liua, H.; Xiao, T.; Lu, J.J.; Lin, L.G.; Chen, X.P.; Zhang, Q.W. Novel biflavonoids from Cephalotaxus oliveri Mast. Phytochem. Lett. 2018, 24, 150–153. [Google Scholar] [CrossRef]
- Abdullaha, I.; Phongpaichitb, S.; Voravuthikunchai, S.P.; Mahabusarakama, W. Prenylated biflavonoids from the green branches of Garcinia dulcis. Phytochem. Lett. 2018, 23, 176–179. [Google Scholar] [CrossRef]
- Shim, S.Y.; Lee, S.G.; Lee, M. Biflavonoids Isolated from Selaginella tamariscina and their Anti-Inflammatory Activities via ERK 1/2 Signaling. Molecules 2018, 23, 926. [Google Scholar] [CrossRef]
- Yan, H.W.; Zhu, H.; Yuan, X.; Yang, Y.N.; Feng, Z.M.; Jiang, J.S.; Zhang, P.C. Eight new biflavonoids with lavandulyl units from the roots of Sophora flavescens and their inhibitory effect on PTP1B. Bioorg. Chem. 2019, 86, 679–685. [Google Scholar] [CrossRef]
- Anuradha, V.; Srinivas, P.V.; Rao, R.R.; Manjulatha, K.; Purohit, M.G.; Rao, J.M. Isolation and synthesis of analgesic and anti-inflammatory compounds from Ochna squarrosa L. Bioorg. Med. Chem. 2006, 74, 6820–6826. [Google Scholar] [CrossRef]
- Pegnyemb, D.E.; Tih, R.G.; Sondengam, B.L.; Blond, A.B.; Bodo, B. Flavonoids of Ochna afzelii. Phytochemistry 2003, 64, 661–665. [Google Scholar] [CrossRef]
- Rao, K.V.; Sreeramulu, K.; Rao, C.V.; Gunasekar, D.; Martin, M.T.; Bodo, B. Two new biflavonoids from Ochna obtusada. J. Nat. Prod. 1997, 60, 632–634. [Google Scholar] [CrossRef]
- Grobler, J.A.; Stillmock, K.; Hu, B.; Witmer, M.; Felock, P.; Espeseth, A.S.; Wolfe, A.; Egbertson, M.; Bourgeois, M.; Melamed, J.; et al. Diketo acid inhibitor mechanism and HIV-1 integrase: Implications for metal binding in the active site of phosphotransferase enzymes. Biochemistry 2002, 99, 6661–6666. [Google Scholar] [CrossRef]
- Siwe Noundou, X.; Musyoka, T.M.; Moses, V.; Ndinteh, D.T.; Mnkandhla, D.; Bishop, O.T.; Krause, R.W.M. Anti-HIV-1 integrase potency of methylgallate from Alchornea cordifolia using in vitro and in silico approaches. Sci. Rep. 2019, 9, 4718. [Google Scholar] [CrossRef]
- Thaithong, S.; Beale, G.H.; Chutmongkonkul, M. Susceptibility of Plasmodium falciparum to five drugs: An in vitro study of isolates mainly from Thailand. Trans. R. Soc. Trop. Med. Hyg. 1983, 77, 228–231. [Google Scholar] [CrossRef]
- Heble, N.K.; Mavillapalli, R.C.; Selvaraj, R.; Jeyabalan, S. Molecular docking studies of phytoconstituents identified in Crocus sativus, Curcuma longa, Cassia occidentalis and Moringa oleifera on thymidylate synthase—An enzyme target for anti-cancer activity. J. Appl. Pharm. Sci. 2016, 6, 131–135. [Google Scholar] [CrossRef]
- Madhuri, M.; Prasad, C.; Rao Avupati, V. In silico protein-ligand docking studies on thiazolidinediones as potential anticancer agents. Int. J. Comp. Appl. 2014, 95, 13–16. [Google Scholar] [CrossRef]
- Messanga, B.B.; Tih, R.G.; Kimbu, S.F.; Sondengam, B.L.; Martin, M.T.; Bodo, B. Calodenone, a new isobiflavonoid from Ochna calodendron. J. Nat. Prod. 1992, 55, 245–248. [Google Scholar] [CrossRef]
- Pegnyemb, D.E.; Tih, R.G.; Sondengam, B.L.; Blond, A.; Bodo, B. Isolation and structure elucidation of a new isoflavonoid from Ochna afzelii. Pharm. Biol. 2003, 41, 218–220. [Google Scholar] [CrossRef]
- Abdullahi, M.I.; Musa, A.M.; Haruna, A.K.; Pateh, U.U.; Sule, M.I.; Abdullahi, M.S.; Abdulmalik, M.; Akinwande, Y.; Abimiku, A.G.; LLiya, L. Isolation and characterization of an anti-microbial biflavonoid from the chloroform-soluble fraction of methanolic root extract of Ochna schweinfurthiana (Ochnaceae). Afr. J. Pharm. Pharmacol. 2014, 8, 93–99. [Google Scholar] [CrossRef]
- Feng, S.; Hao, J.; Xu, Z.; Chen, T.; Qiu, S.X.S. Polyprenylated isoflavanone and isoflavonoids from Ormosia henryi and their cytotoxicity and anti-oxidation activity. Fitoterapia 2012, 83, 161–165. [Google Scholar] [CrossRef]
- Ghogomu, T.R.; Sondengam, B.L.; Martin, M.T.; Bodo, B. Lophirone A, a biflavonoid with unusual skeleton from Lophira lanceolata. Tetrahedron Lett. 1987, 28, 2967–2968. [Google Scholar] [CrossRef]
- Chaniad, P.; Mungthin, M.; Payaka, A.; Viriyavejakul, P.; Punsawad, C. Antimalarial properties and molecular docking analysis of compounds from Dioscorea bulbifera L. as new antimalarial agent candidates. BMC Complement. Med. Ther. 2021, 21, 144. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 1 | 2 | 3 | ||||||
|---|---|---|---|---|---|---|---|---|
| No. | δC | δH (J in Hz) | No. | δC | δH (J in Hz) | No. | δC | δH (J in Hz) |
| B1-1 | 108.6 | - | B1-1 | 108.6 | - | B1-1 | 108.6 | - |
| 2 | 159.5 | - | 2 | 159.5 | - | 2 | 159.5 | - |
| 3 | 103.3 | 6.71 (d, 2.5) | 3 | 103.3 | 6.71 (d, 2.5) | 3 | 103.3 | 6.71 (d, 2.5) |
| 4 | 164.7 | - | 4 | 164.9 | - | 4 | 164.9 | - |
| 5 | 116.6 | 6.85 (dd, 2.5, 9.0) | 5 | 116.6 | 6.85 (dd, 2.5, 9.0) | 5 | 116.6 | 6.85 (dd, 2.5, 9.0) |
| 6 | 128.3 | 7.88 (d, 9.0) | 6 | 128.3 | 7.88 (d, 9.0) | 6 | 128.3 | 7.88 (d, 9.0) |
| C-1 | 177.2 | - | C-1 | 177.2 | - | C-1 | 177.2 | - |
| a1 | 122.7 | - | a1 | 122.7 | - | a1 | 122.7 | - |
| b1 | 157.5 | 8.23 (s) | b1 | 157.5 | 8.23 (s) | b1 | 157.5 | 8.23 (s) |
| B2-1′ | 114.4 | - | B2-1′ | 114.4 | - | B2-1′ | 114.4 | - |
| 2′ | 166.6 | - | 2′ | 166.6 | - | 2′ | 166.6 | - |
| 3′ | 103.6 | 6.14 (d, 2.0) | 3′ | 103.6 | 6.14 (d, 2.0) | 3′ | 103.6 | 6.14 (d, 2.0) |
| 4′ | 167.0 | - | 4′ | 167.0 | - | 4′ | 167.0 | - |
| 5′ | 109.3 | 6.34 (dd, 2.0, 9.0) | 5′ | 109.3 | 6.34 (dd, 2.0, 9.0) | 5′ | 109.3 | 6.34 (dd, 2.0, 9.0) |
| 6′ | 136.0 | 8.15 (d, 9.0) | 6′ | 136.0 | 8.15 (d, 9.0) | 6′ | 136.0 | 8.15 (d, 9.0) |
| C-2 | 204.9 | - | C-2 | 204.9 | - | C-2 | 204.9 | - |
| a2 | 44.8 | 6.01 (d, 11.0) | a2 | 44.8 | 6.01 (d, 11.0) | a2 | 44.8 | 6.01 (d, 11.0) |
| b2 | 54.4 | 4.67 (d, 12.0) | b2 | 54.4 | 4.67 (d, 12.0) | b2 | 54.4 | 4.67 (d, 12.0) |
| A1-1″ | 134.4 | - | A1-1″ | 134.4 | - | A1-1″ | 134.4 | - |
| 2″ | 129.9 | 7.13 (d, 8.5) | 2″ | 129.9 | 7.13 (d, 8.5) | 2″ | 129.9 | 7.13 (d, 8.5) |
| 3″ | 116.1 | 6.65 (d, 8.5) | 3″ | 116.1 | 6.65 (d, 8.5) | 3″ | 116.1 | 6.65 (d, 8.5) |
| 4″ | 156.7 | - | 4″ | 159.5 | - | 4″ | 159.7 | - |
| 5″ | 116.1 | 6.65 (d, 8.5) | 5″ | 116.1 | 6.65 (d, 8.5) | 5″ | 116.1 | 6.65 (d, 8.5) |
| 6″ | 129.9 | 7.13 (d, 8.5) | 6″ | 129.9 | 7.13 (d, 8.5) | 6″ | 129.9 | 7.13 (d, 8.5) |
| A2-1‴ | 134.9 | - | A2-1‴ | 134.9 | - | A2-1‴ | 134.9 | - |
| 2‴ | 130.5 | 7.17 (d, 8.5) | 2‴ | 130.5 | 7.17 (d, 8.5) | 2‴ | 130.5 | 7.17 (d, 8.5) |
| 3‴ | 116.2 | 6.60 (d, 8.5) | 3‴ | 116.2 | 6.60 (d, 8.5) | 3‴ | 116.2 | 6.60 (d, 8.5) |
| 4‴ | 156.8 | - | 4‴ | 159.6 | - | 4‴ | 159.6 | - |
| 5‴ | 116.2 | 6.60 (d, 8.5) | 5‴ | 116.2 | 6.60 (d, 8.5) | 5‴ | 116.2 | 6.60 (d, 8.5) |
| 6‴ | 130.5 | 7.17 (d, 8.5) | 6‴ | 130.5 | 7.17 (d, 8.5) | 6‴ | 130.5 | 7.17 (d, 8.5) |
| OH | - | - | 4‴-OMe | 54.9 | 3.63 | 4‴-OMe | 55.4 | 3.65 |
| OH | - | - | 4″-OMe | 55.7 | 3.63 | 4-OMe | 55.5 | 3.65 |
| OH | - | - | 4′-OMe | 56.2 | 3.78 | OH | - | - |
| OH | - | - | 4-OMe | 55.6 | 3.75 | OH | - | - |
| OH | - | OH | - | - | OH | - | - |
| Compounds | Assembly IC50 (μM) |
|---|---|
| 1 | 0.047 ± 0.021 b |
| 2 | - |
| 3 | - |
| 4 | - |
| 5 | - |
| 6 | - |
| 7 | - |
| Chicoric Acid | 0.006 ± 0.002 a |
| Compounds | NF54: IC50 (μM) |
|---|---|
| 1 | - |
| 2 | 4.60 ± 6.09 b |
| 3 | 7.86 ± 5.12 b |
| 4 | 8.20 ± 0.93 b |
| 5 | 5.11 ± 13.7 b |
| 6 | - |
| 7 | - |
| Chloroquine (CQ) | 0.006 ± 0.002 a |
| Artesumate | 0.002 ± ND |
| Compound/Drug | Energy (Kcal/mol) | VDW (Kcal/mol) | Hbond (Kcal/mol) | Elec (Kcal/mol) |
|---|---|---|---|---|
| Compound 1 | −121.8 | −96.54 | −24.46 | 0 |
| Compound 2 | −131.88 | −116.91 | −14.97 | 0 |
| Compound 7 | −92.85 | −68.24 | −24.6 | 0 |
| Chicoric acid | −116.06 | −83.27 | −28.28 | −4.52 |
| Dolutegravir | −100.27 | −77.05 | −23.22 | 0 |
| Compound/Drug | Energy (Kcal/mol) | VDW (Kcal/mol) | Hbond (Kcal/mol) | Elec (Kcal/mol) |
|---|---|---|---|---|
| Compound 1 | −125.33 | −107.75 | −17.59 | 0 |
| Compound 2 | −124.95 | −116.11 | −8.84 | 0 |
| Compound 7 | −90.99 | −85.06 | −5.93 | 0 |
| Chloroquine | −84.48 | −69.36 | −15.11 | 0 |
| Target | Compound 1 | Compound 2 | Compound 7 | Chicoric Acid | Dolutegravir |
|---|---|---|---|---|---|
| 3LPT | Tyr83, Ala86, Arg107, Asn184, Tyr83, Glu85, Trp108, Glu177, Val180, Asn184 (5 hydrogen bonds) | Gln62, Asp116, Asn 144, Asp64, Tyr143, Glu152, Ser153, Met154, Asn155 (3 hydrogen bonds) | Asp116, Pro142, Asn144, Asn155, Glu62, Tyr143, Glu152, Ser153 (4 hydrogen bonds) | His 171, Lys71, Val72, Ile89, Pro90, His171, Lys 173, Leu172 (6 hydrogen bonds) | Asp116, Asn144, Gln62, Glu152, Ser153, Met154, Asn155 (3 hydrogen bonds) |
| Target | Compound 1 | Compound 2 | Compound 7 | Chloroquine |
|---|---|---|---|---|
| 2LOE | Asn76, Leu77, Leu83, Val65, Glu78, Thr84, Asn85, Ile86 (Hydro 3) | Lys82, Asn85, Val65, Glu78, Leu83, Thr84, Ile86 (Hydro2) | Ala44, Lys41, Ala42, His43, Ile103, Ala127 (Hydro 1) | His61, Phe68, Asn69, Leu70, Ser71, Gly72, Phe110, Val122 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Messi, A.N.; Bonnet, S.L.; Owona, B.A.; Wilhelm, A.; Kamto, E.L.D.; Ndongo, J.T.; Siwe-Noundou, X.; Poka, M.; Demana, P.H.; Krause, R.W.M.; et al. In Vitro and In Silico Potential Inhibitory Effects of New Biflavonoids from Ochna rhizomatosa on HIV-1 Integrase and Plasmodium falciparum. Pharmaceutics 2022, 14, 1701. https://doi.org/10.3390/pharmaceutics14081701
Messi AN, Bonnet SL, Owona BA, Wilhelm A, Kamto ELD, Ndongo JT, Siwe-Noundou X, Poka M, Demana PH, Krause RWM, et al. In Vitro and In Silico Potential Inhibitory Effects of New Biflavonoids from Ochna rhizomatosa on HIV-1 Integrase and Plasmodium falciparum. Pharmaceutics. 2022; 14(8):1701. https://doi.org/10.3390/pharmaceutics14081701
Chicago/Turabian StyleMessi, Angélique Nicolas, Susan Lucia Bonnet, Brice Ayissi Owona, Anke Wilhelm, Eutrophe Le Doux Kamto, Joseph Thierry Ndongo, Xavier Siwe-Noundou, Madan Poka, Patrick H. Demana, Rui W. M. Krause, and et al. 2022. "In Vitro and In Silico Potential Inhibitory Effects of New Biflavonoids from Ochna rhizomatosa on HIV-1 Integrase and Plasmodium falciparum" Pharmaceutics 14, no. 8: 1701. https://doi.org/10.3390/pharmaceutics14081701