
Thermosensitive PLGA–PEG–PLGA Hydrogel as Depot Matrix for Allergen-Specific Immunotherapy

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Hydrogel Synthesis
2.2. Determination of Gelling Temperature and Stability of Polymer Solution
2.3. Preparation of 20% (w/v) Hydrogel for Injection
2.4. In Vitro Protein Release Assays
2.5. Animals
2.6. MR Imaging
2.7. Murine Model of AIT
2.8. In Vivo Release Assay
2.9. Isolation of BALF Cells and Lung Lymphocytes
2.10. Flow Cytometry
2.11. Measurement of Immunoglobulins
2.12. Histological Analysis
2.13. Statistical Analysis
3. Results
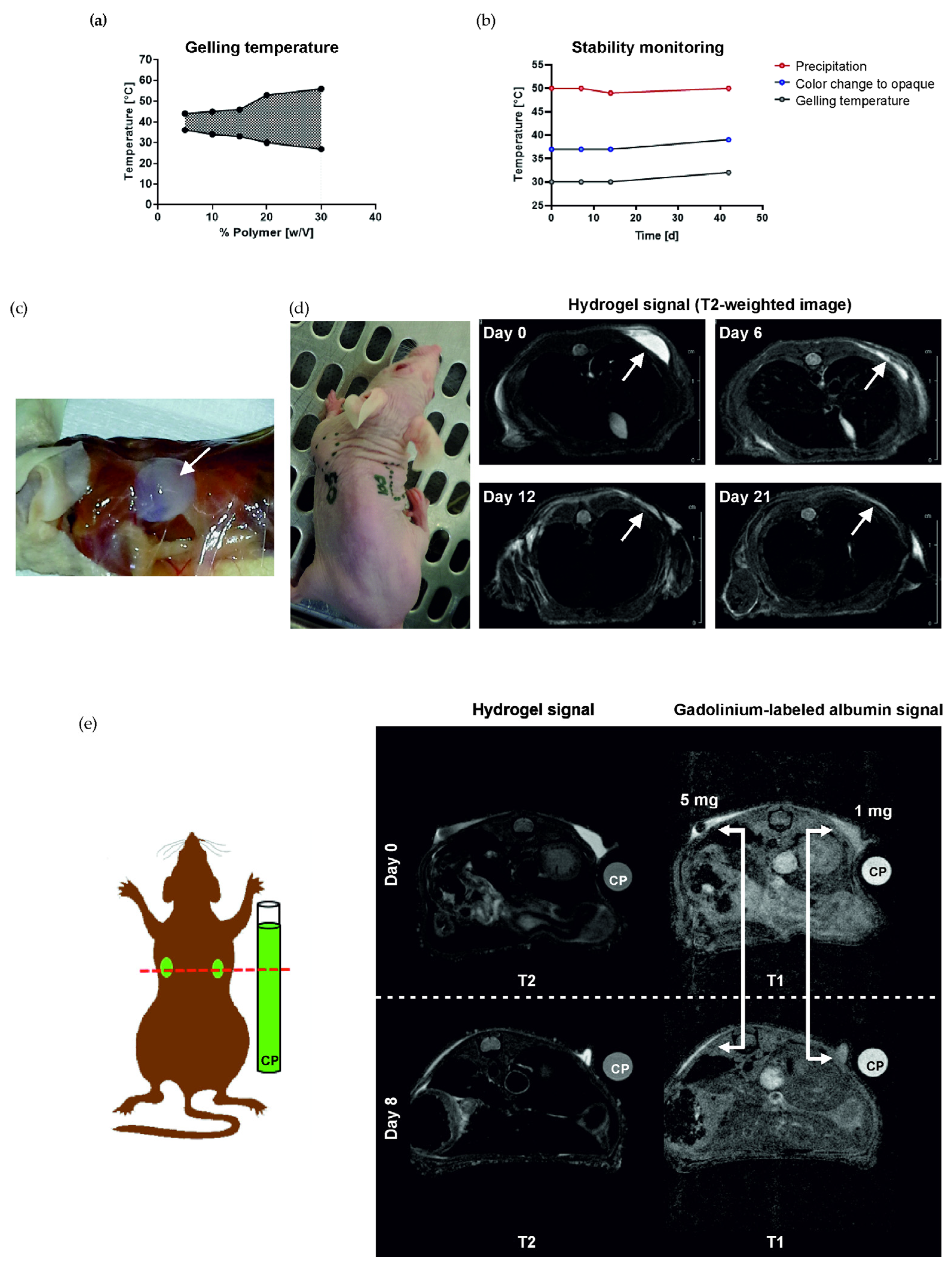
3.1. Hydrogel Forms a Solid Depot and Is Biodegradable In Vivo
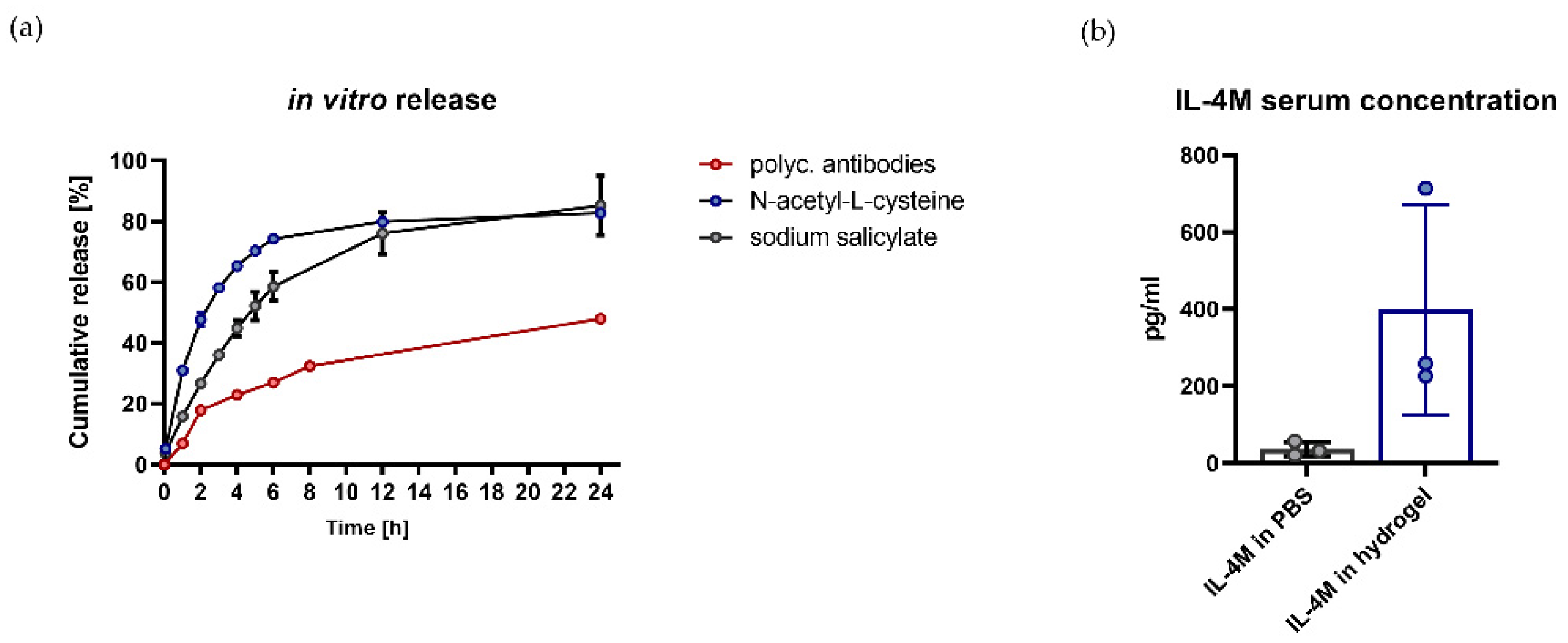
3.2. Hydrogel Is Suitable for In Vitro and In Vivo Release of Bioactive Molecules
3.3. Administration of AIT via a Hydrogel Depot Significantly Reduces the Infiltration of BALF Cells and the Production of Th2 Cytokines
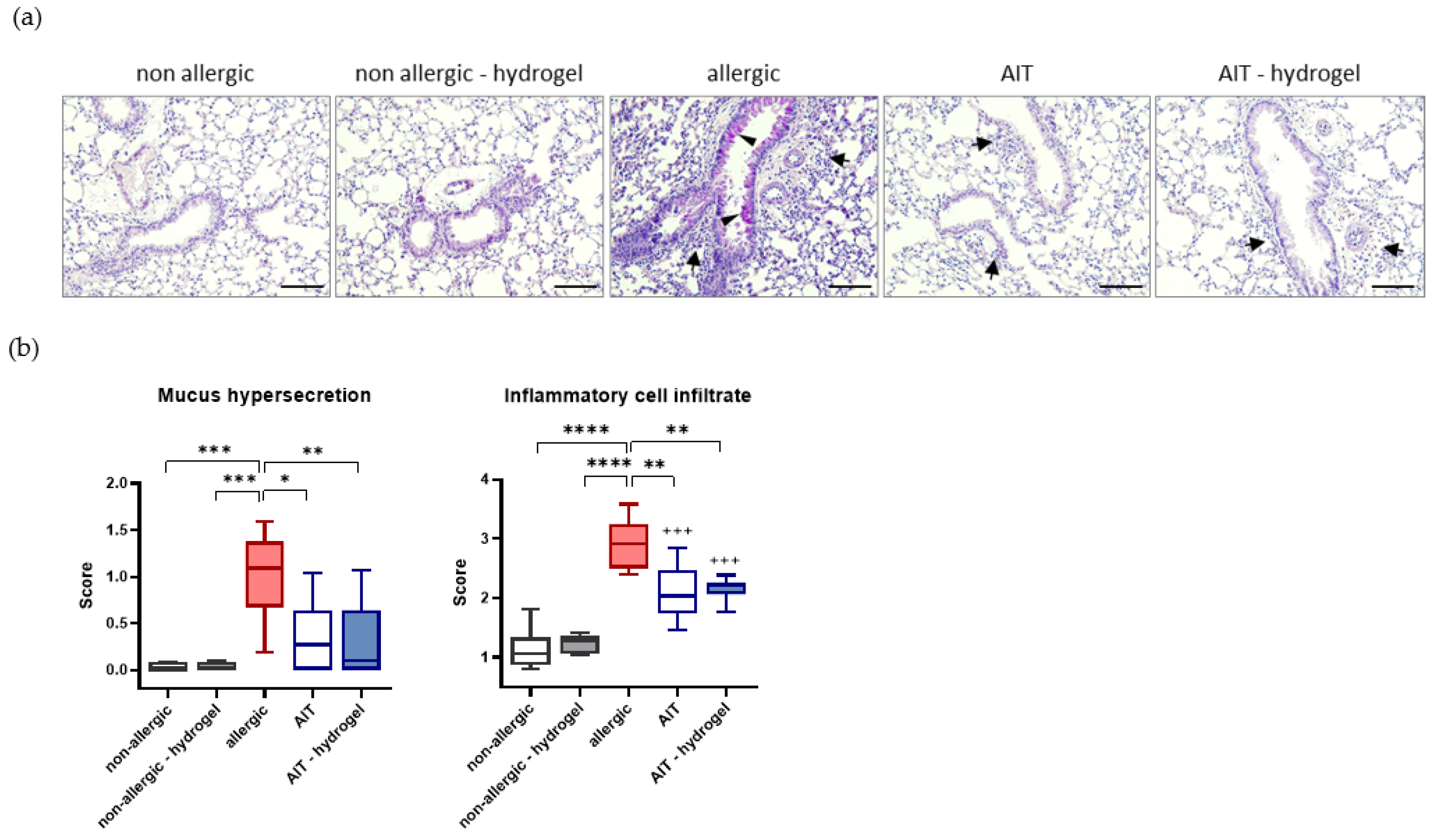
3.4. AIT Delivered with Hydrogel Significantly Reduces Mucus Hypersecretion and Cell Infiltration in Lung Tissue
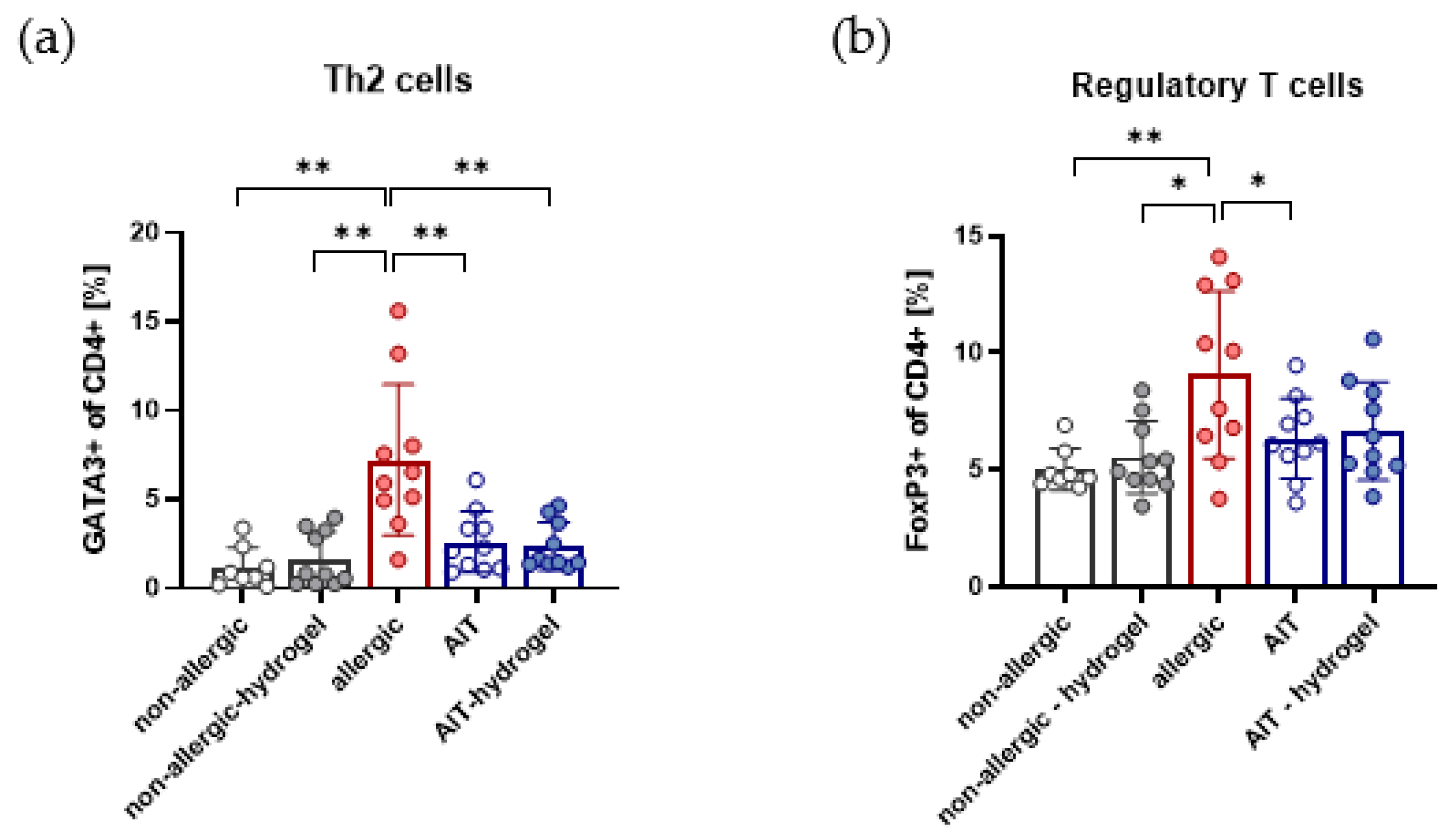
3.5. Effects on Th2 and Treg Cells in Lung Lymphocytes Are Independent of AIT Delivery Strategy
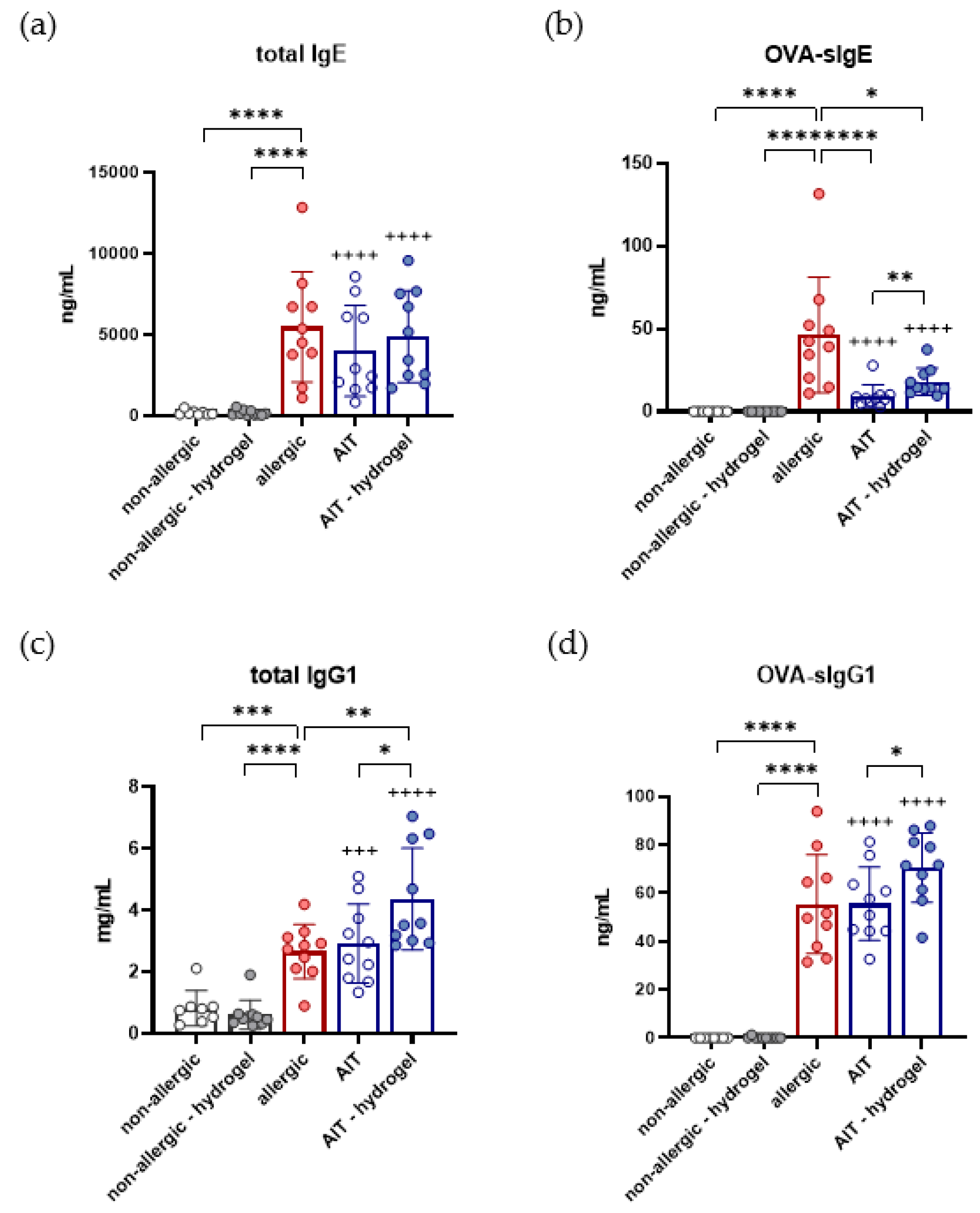
3.6. Total IgG1, OVA-Specific IgG1, and OVA-Specific IgE Are Increased in AIT–Hydrogel Treated Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pawankar, R. Allergic diseases and asthma: A global public health concern and a call to action. World Allergy Organ. J. 2014, 7, 12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pinart, M.; Albang, R.; Maier, D.; Duran-Tauleria, E.; Mena, G.; Gimeno-Santos, E.; Solà, I.; Garcia-Aymerich, J.; Guerra, S.; Stein, R.; et al. Systematic Review on the Definition of Allergic Diseases in Children: The MeDALL Study. Int. Arch. Allergy Immunol. 2015, 168, 110–121. [Google Scholar] [CrossRef] [PubMed]
- García-Almaraz, R.; Reyes-Noriega, N.; Del-Río-Navarro, B.E.; Berber, A.; Navarrete-Rodríguez, E.M.; Ellwood, P.; Álvarez, L.G.M.; Palacio, V.J.M.; García, B.D.C.R.; Domínguez, A.J.E.; et al. Prevalence and risk factors associated with allergic rhinitis in Mexican school children: Global Asthma Network Phase I. World Allergy Organ. J. 2021, 14, 100492. [Google Scholar] [CrossRef] [PubMed]
- Kubo, T.; Morita, H.; Sugita, K.; Akdis, C.A. Introduction to Mechanisms of Allergic Diseases. In Middleton’s Allergy Essentials; Elsevier: Amsterdam, The Netherlands, 2017; pp. 1–27. [Google Scholar] [CrossRef]
- Nauta, A.J.; Engels, F.; Knippels, L.M.; Garssen, J.; Nijkamp, F.P.; Redegeld, F.A. Mechanisms of allergy and asthma. Eur. J. Pharmacol. 2008, 585, 354–360. [Google Scholar] [CrossRef]
- Holgate, S.T.; Wenzel, S.; Postma, D.S.; Weiss, S.T.; Renz, H.; Sly, P.D. Asthma. Nat. Rev. Dis. Primers 2015, 1, 15025. [Google Scholar] [CrossRef]
- Kucuksezer, U.C.; Ozdemir, C.; Cevhertas, L.; Ogulur, I.; Akdis, M.; Akdis, C.A. Mechanisms of allergen-specific immunotherapy and allergen tolerance. Allergol. Int. 2020, 69, 549–560. [Google Scholar] [CrossRef]
- Moote, W.; Kim, H.; Ellis, A.K. Allergen-specific immunotherapy. Allergy Asthma Clin. Immunol. 2018, 14 (Suppl. S2). [Google Scholar] [CrossRef] [Green Version]
- Głobińska, A.; Boonpiyathad, T.; Satitsuksanoa, P.; Kleuskens, M.; Van De Veen, W.; Sokolowska, M.; Akdis, M. Mechanisms of allergen-specific immunotherapy: Diverse mechanisms of immune tolerance to allergens. Ann. Allergy Asthma Immunol. 2018, 121, 306–312. [Google Scholar] [CrossRef]
- Tavares, L.P.; Galvão, I.; Ferrero, M.R. Novel Immunomodulatory Therapies for Respiratory Pathologies, in Reference Module in Biomedical Sciences. Compr. Pharmacol. 2021, 2022, 554–594. [Google Scholar]
- Moingeon, P. Adjuvants for allergy vaccines. Hum. Vaccines Immunother. 2012, 8, 1492–1498. [Google Scholar] [CrossRef] [Green Version]
- Chiang, P.-C.; Nagapudi, K.; Fan, P.W.; Liu, J. Investigation of Drug Delivery in Rats via Subcutaneous Injection: Case Study of Pharmacokinetic Modeling of Suspension Formulations. J. Pharm. Sci. 2019, 108, 109–119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dirnena-Fusini, I.; Åm, M.K.; Fougner, A.L.; Carlsen, S.M.; Christiansen, S.C. Intraperitoneal, subcutaneous and intravenous glucagon delivery and subsequent glucose response in rats: A randomized controlled crossover trial. BMJ Open Diabetes Res. Care 2018, 6, e000560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeong, S.-H.; Jang, J.-H.; Lee, Y.-B. Pharmacokinetic Comparison of Three Different Administration Routes for Topotecan Hydrochloride in Rats. Pharmaceuticals 2020, 13, 231. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Han, Y.; Huang, W.; Jin, M.; Gao, Z. The influence of the gut microbiota on the bioavailability of oral drugs. Acta Pharm. Sin. B 2020, 11, 1789–1812. [Google Scholar] [CrossRef]
- Shin, Y.S.; Takeda, K.; Gelfand, E.W. Understanding Asthma Using Animal Models. Allergy Asthma Immunol. Res. 2009, 1, 10–18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandrini, F.; Musiol, S.; Schneider, E.; Blanco-Pérez, F.; Albrecht, M. Mimicking Antigen-Driven Asthma in Rodent Models—How Close Can We Get? Front. Immunol. 2020, 11, 575936. [Google Scholar] [CrossRef]
- Pfaar, O.; Bachert, C.; Bufe, A.; Buhl, R.; Ebner, C.; Eng, P.; Friedrichs, F.; Fuchs, T.; Hamelmann, E.; Hartwig-Bade, D.; et al. Guideline on allergen-specific immunotherapy in IgE-mediated allergic diseases. Allergo J. Int. 2014, 23, 282–319. [Google Scholar] [CrossRef]
- Walker, S.; Varney, V.A.; Gaga, M.; Jacobson, M.; Durham, S.R. Grass pollen immunotherapy: Efficacy and safety during a 4-year follow-up study. Allergy 1995, 50, 405–413. [Google Scholar] [CrossRef]
- Albanesi, M.; Nico, A.; Sinisi, A.; Giliberti, L.; Rossi, M.P.; Rossini, M.; Kourtis, G.; Rucco, A.S.; LoConte, F.; Muolo, L.; et al. A 13-year real-life study on efficacy, safety and biological effects of Vespula venom immunotherapy. Clin. Mol. Allergy 2018, 16, 2. [Google Scholar] [CrossRef] [Green Version]
- Kool, M.; Soullié, T.; Van Nimwegen, M.; Willart, M.A.; Muskens, F.; Jung, S.; Hoogsteden, H.C.; Hammad, H.; Lambrecht, B.N. Alum adjuvant boosts adaptive immunity by inducing uric acid and activating inflammatory dendritic cells. J. Exp. Med. 2008, 205, 869–882. [Google Scholar] [CrossRef] [Green Version]
- Sokolovska, A.; Hem, S.L.; HogenEsch, H. Activation of dendritic cells and induction of CD4+ T cell differentiation by aluminum-containing adjuvants. Vaccine 2007, 25, 4575–4585. [Google Scholar] [CrossRef] [PubMed]
- Brewer, J.M.; Conacher, M.; Satoskar, A.; Bluethmann, H.; Alexander, J. In interleukin-4-deficient mice, alum not only generates T helper 1 responses equivalent to Freund’s complete adjuvant, but continues to induce T helper 2 cytokine production. Eur. J. Immunol. 1996, 26, 2062–2066. [Google Scholar] [CrossRef] [PubMed]
- Ghimire, T.R. The mechanisms of action of vaccines containing aluminum adjuvants: An in vitro vs in vivo paradigm. SpringerPlus 2015, 4, 118. [Google Scholar] [CrossRef] [Green Version]
- Akdis, C.A.; Akdis, M. Mechanisms of allergen-specific immunotherapy. J. Allergy Clin. Immunol. 2011, 127, 18–27. [Google Scholar] [CrossRef] [Green Version]
- Lundberg, K.; Rydnert, F.; Broos, S.; Andersson, M.; Greiff, L.; Lindstedt, M. Allergen-Specific Immunotherapy Alters the Frequency, as well as the FcR and CLR Expression Profiles of Human Dendritic Cell Subsets. PLoS ONE 2016, 11, e0148838. [Google Scholar] [CrossRef]
- Radulovic, S.; Jacobson, M.R.; Durham, S.R.; Nouri-Aria, K.T. Grass pollen immunotherapy induces Foxp3-expressing CD4+CD25+ cells in the nasal mucosa. J. Allergy Clin. Immunol. 2008, 121, 1467–1472. [Google Scholar] [CrossRef] [Green Version]
- Woo, H.-Y.; Kim, Y.-S.; Kang, N.-I.; Chung, W.-C.; Song, C.-H.; Choi, I.-W.; Choi, I.-H.; Lee, H.-K. Mechanism for acute oral desensitization to antibiotics. Allergy 2006, 61, 954–958. [Google Scholar] [CrossRef] [PubMed]
- Jensen-Jarolim, E. Aluminium in Allergies and Allergen immunotherapy. World Allergy Organ. J. 2015, 8, 7. [Google Scholar] [CrossRef] [Green Version]
- Petrovsky, N. Comparative Safety of Vaccine Adjuvants: A Summary of Current Evidence and Future Needs. Drug Saf. 2015, 38, 1059–1074. [Google Scholar] [CrossRef]
- Principi, N.; Esposito, S. Aluminum in vaccines: Does it create a safety problem? Vaccine 2018, 36, 5825–5831. [Google Scholar] [CrossRef]
- Willhite, C.C.; Karyakina, N.A.; Yokel, R.; Yenugadhati, N.; Wisniewski, T.; Arnold, I.M.F.; Momoli, F.; Krewski, D. Systematic review of potential health risks posed by pharmaceutical, occupational and consumer exposures to metallic and nanoscale aluminum, aluminum oxides, aluminum hydroxide and its soluble salts. Crit. Rev. Toxicol. 2014, 44, 1–80. [Google Scholar] [CrossRef] [PubMed]
- Exley, C. The toxicity of aluminium in humans. Morphologie 2016, 100, 51–55. [Google Scholar] [CrossRef] [PubMed]
- Kopeček, J. Hydrogel biomaterials: A smart future? Biomaterials 2007, 28, 5185–5192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahinroosta, M.; Farsangi, Z.J.; Allahverdi, A.; Shakoori, Z. Hydrogels as intelligent materials: A brief review of synthesis, properties and applications. Mater. Today Chem. 2018, 8, 42–55. [Google Scholar] [CrossRef]
- Park, H.; Park, K.; Shalaby, W.S. Biodegradable Hydrogels for Drug Delivery; CRC Press: Boca Raton, FL, USA, 1993. [Google Scholar] [CrossRef]
- Chen, S.; Singh, J. Controlled release of growth hormone from thermosensitive triblock copolymer systems: In vitro and in vivo evaluation. Int. J. Pharm. 2008, 352, 58–65. [Google Scholar] [CrossRef]
- Chen, D.C.D.; Ma, X.M.X.; Qiao, M.Q.M. Injectable thermosensitive PLGA-PEG-PLGA triblock copolymers-based hydrogels as carriers for interleukin-2. Pharmazie 2007, 63, 27–30. [Google Scholar] [CrossRef]
- Makadia, H.K.; Siegel, S.J. Poly lactic-co-glycolic acid (PLGA) as biodegradable controlled drug delivery carrier. Polymers 2011, 3, 1377–1397. [Google Scholar] [CrossRef]
- Jeong, B.; Bea, Y.H.; Kim, S.W. In situ gelation of PEG-PLGA-PEG triblock copolymer aqueous solutions and degradation thereof. J. Biomed. Mater. Res. 1999, 50, 171–177. [Google Scholar] [CrossRef]
- Qiao, M.; Chen, D.; Ma, X.; Liu, Y. Injectable biodegradable temperature-responsive PLGA–PEG–PLGA copolymers: Synthesis and effect of copolymer composition on the drug release from the copolymer-based hydrogels. Int. J. Pharm. 2005, 294, 103–112. [Google Scholar] [CrossRef]
- Russkamp, D.; Aguilar-Pimentel, A.; Alessandrini, F.; Gailus-Durner, V.; Fuchs, H.; Ohnmacht, C.; Chaker, A.; de Angelis, M.H.; Ollert, M.; Schmidt-Weber, C.B.; et al. IL-4 receptor α blockade prevents sensitization and alters acute and long-lasting effects of allergen-specific immunotherapy of murine allergic asthma. Allergy 2019, 74, 1549–1560. [Google Scholar] [CrossRef] [Green Version]
- Grassetti, D.; Murray, J. Determination of sulfhydryl groups with 2,2′- or 4,4′-dithiodipyridine. Arch. Biochem. Biophys. 1967, 119, 41–49. [Google Scholar] [CrossRef]
- Kukoc-Modun, L.; Radić, N. Spectrophotometric Determination of N-Acetyl-L-Cysteine and N-(2-Mercaptopropionyl)-Glycine in Pharmaceutical Preparations. Int. J. Anal. Chem. 2011, 2011, 140756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alessandrini, F.; Schulz, H.; Takenaka, S.; Lentner, B.; Karg, E.; Behrendt, H.; Jakob, T. Effects of ultrafine carbon particle inhalation on allergic inflammation of the lung. J. Allergy Clin. Immunol. 2006, 117, 824–830. [Google Scholar] [CrossRef]
- Abbas, M.; Moussa, M.; Akel, H. Type I Hypersensitivity Reaction; StatPearls: Treasure Island, FL, USA, 2021. [Google Scholar]
- George, L.; Brightling, C.E. Eosinophilic airway inflammation: Role in asthma and chronic obstructive pulmonary disease. Ther. Adv. Chronic Dis. 2016, 7, 34–51. [Google Scholar] [CrossRef] [Green Version]
- Lantz, C.S.; Yamaguchi, M.; Oettgen, H.C.; Katona, I.M.; Miyajima, I.; Kinet, J.P.; Galli, S.J. IgE regulates mouse basophil Fc epsilon RI expression in vivo. J. Immunol. 1997, 158, 2517–2521. [Google Scholar] [PubMed]
- Yamaguchi, M.; Lantz, C.S.; Oettgen, H.C.; Katona, I.M.; Fleming, T.; Miyajima, I.; Kinet, J.-P.; Galli, S.J. IgE Enhances Mouse Mast Cell FcεRI Expression In Vitro and In Vivo: Evidence for a Novel Amplification Mechanism in IgE-dependent Reactions. J. Exp. Med. 1997, 185, 663–672. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamilton, R.G.; Saito, H. IgE antibody concentration, specific activity, clonality, and affinity measures from future diagnostic confirmatory tests. J. Allergy Clin. Immunol. 2008, 122, 305–306. [Google Scholar] [CrossRef]
- Shamji, M.H.; Valenta, R.; Jardetzky, T.; Verhasselt, V.; Durham, S.R.; Würtzen, P.A.; van Neerven, R.J. The role of allergen-specific IgE, IgG and IgA in allergic disease. Allergy 2021, 76, 3627–3641. [Google Scholar] [CrossRef]
- Strait, R.T.; Morris, S.C.; Finkelman, F.D. IgG-blocking antibodies inhibit IgE-mediated anaphylaxis in vivo through both antigen interception and Fc RIIb cross-linking. J. Clin. Investig. 2006, 116, 833–841. [Google Scholar] [CrossRef] [Green Version]
- Platts-Mills, T.A.; Von Maur, R.K.; Ishizaka, K.; Norman, P.S.; Lichtenstein, L.M. IgA and IgG anti-ragweed antibodies in nasal secretions. Quantitative measurements of antibodies and correlation with inhibition of histamine release. J. Clin. Investig. 1976, 57, 1041–1050. [Google Scholar] [CrossRef]
- Shamji, M.H.; Larson, D.; Eifan, A.; Scadding, G.W.; Qin, T.; Lawson, K.; Sever, M.L.; Macfarlane, E.; Layhadi, J.A.; Würtzen, P.A.; et al. Differential induction of allergen-specific IgA responses following timothy grass subcutaneous and sublingual immunotherapy. J. Allergy Clin. Immunol. 2021, 148, 1061–1071. [Google Scholar] [CrossRef]
- Shamji, M.H.; Kappen, J.H.; Akdis, M.; Jensen-Jarolim, E.; Knol, E.F.; Kleine-Tebbe, J.; Bohle, B.; Chaker, A.M.; Till, S.J.; Valenta, R.; et al. Biomarkers for monitoring clinical efficacy of allergen immunotherapy for allergic rhinoconjunctivitis and allergic asthma: An EAACI Position Paper. Allergy 2017, 72, 1156–1173. [Google Scholar] [CrossRef] [PubMed]
- Zha, L.; Leoratti, F.M.; He, L.; Mohsen, M.O.; Cragg, M.; Storni, F.; Vogel, M.; Bachmann, M.F. An unexpected protective role of low-affinity allergen-specific IgG through the inhibitory receptor FcγRIIb. J. Allergy Clin. Immunol. 2018, 142, 1529–1536. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, M.; Su, Q.; Lai, X.; Xian, M.; Shi, X.; Wurtzen, P.A.; Qin, R.; Zeng, X.; Li, J. Functional and Immunoreactive Levels of IgG4 Correlate with Clinical Responses during the Maintenance Phase of House Dust Mite Immunotherapy. J. Immunol. 2018, 200, 3897–3904. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gehlhar, K.; Schlaak, M.; Becker, W.-M.; Bufe, A. Monitoring allergen immunotherapy of pollen-allergic patients: The ratio of allergen-specific IgG4 to IgG1 correlates with clinical outcome. Clin. Exp. Allergy 1999, 29, 497–506. [Google Scholar] [CrossRef]
- Schmid, J.M.; Würtzen, P.A.; Dahl, R.; Hoffmann, H.J. Pretreatment IgE sensitization patterns determine the molecular profile of the IgG4 response during updosing of subcutaneous immunotherapy with timothy grass pollen extract. J. Allergy Clin. Immunol. 2016, 137, 562–570. [Google Scholar] [CrossRef] [Green Version]
- Lilienthal, G.-M.; Rahmöller, J.; Petry, J.; Bartsch, Y.C.; Leliavski, A.; Ehlers, M. Potential of Murine IgG1 and Human IgG4 to Inhibit the Classical Complement and Fcγ Receptor Activation Pathways. Front. Immunol. 2018, 9, 958. [Google Scholar] [CrossRef]
- Casale, T.B.; Stokes, J.R. Immunotherapy: What lies beyond. J. Allergy Clin. Immunol. 2014, 133, 612–619. [Google Scholar] [CrossRef]
- Ekramoddoullah, A.; Kisil, F.; Sehon, A. Suppression of the IgE Antibody Response in Mice to Kentucky Blue Grass Pollen Allergens. Int. Arch. Allergy Immunol. 1977, 55, 247–254. [Google Scholar] [CrossRef]
- Marichal, T.; Ohata, K.; Bedoret, D.; Mesnil, C.; Sabatel, C.; Kobiyama, K.; Lekeux, P.; Coban, C.; Akira, S.; Ishii, K.; et al. DNA released from dying host cells mediates aluminum adjuvant activity. Nat. Med. 2011, 17, 996–1002. [Google Scholar] [CrossRef]
- HogenEsch, H. Mechanism of Immunopotentiation and Safety of Aluminum Adjuvants. Front. Immunol. 2012, 3, 406. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klimek, L.; Brehler, R.; Hamelmann, E.; Kopp, M.; Ring, J.; Treudler, R.; Jakob, T.; Worm, M.; Pfaar, O. Development of subcutaneous allergen immunotherapy (part 2): Preventive aspects and innovations. Allergo J. Int. 2019, 28, 107–119. [Google Scholar] [CrossRef] [Green Version]
- Kramer, M.; Heath, M.D. Aluminium in allergen-specific subcutaneous immunotherapy–A German perspective. Vaccine 2014, 32, 4140–4148. [Google Scholar] [CrossRef] [Green Version]
- Exley, C.; Siesjö, P.; Eriksson, H. The immunobiology of aluminium adjuvants: How do they really work? Trends Immunol. 2010, 31, 103–109. [Google Scholar] [CrossRef] [PubMed]
- Banasik, A.; Lankoff, A.; Piskulak, A.; Adamowska, K.; Lisowska, H.; Wojcik, A. Aluminum-induced micronuclei and apoptosis in human peripheral-blood lymphocytes treated during different phases of the cell cycle. Environ. Toxicol. 2005, 20, 402–406. [Google Scholar] [CrossRef] [PubMed]
- Leuthard, D.S.; Duda, A.; Freiberger, S.N.; Weiss, S.; Dommann, I.; Fenini, G.; Contassot, E.; Kramer, M.F.; Skinner, M.A.; Kündig, T.M.; et al. Microcrystalline Tyrosine and Aluminum as Adjuvants in Allergen-Specific Immunotherapy Protect from IgE-Mediated Reactivity in Mouse Models and Act Independently of Inflammasome and TLR Signaling. J. Immunol. 2018, 200, 3151–3159. [Google Scholar] [CrossRef] [Green Version]
- Pavón-Romero, G.F.; Parra-Vargas, M.I.; Ramírez-Jiménez, F.; Melgoza-Ruiz, E.; Serrano-Pérez, N.H.; Teran, L.M. Allergen Immunotherapy: Current and Future Trends. Cells 2022, 11, 212. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heine, S.; Aguilar-Pimentel, A.; Russkamp, D.; Alessandrini, F.; Gailus-Durner, V.; Fuchs, H.; Ollert, M.; Bredehorst, R.; Ohnmacht, C.; Zissler, U.M.; et al. Thermosensitive PLGA–PEG–PLGA Hydrogel as Depot Matrix for Allergen-Specific Immunotherapy. Pharmaceutics 2022, 14, 1527. https://doi.org/10.3390/pharmaceutics14081527
Heine S, Aguilar-Pimentel A, Russkamp D, Alessandrini F, Gailus-Durner V, Fuchs H, Ollert M, Bredehorst R, Ohnmacht C, Zissler UM, et al. Thermosensitive PLGA–PEG–PLGA Hydrogel as Depot Matrix for Allergen-Specific Immunotherapy. Pharmaceutics. 2022; 14(8):1527. https://doi.org/10.3390/pharmaceutics14081527
Chicago/Turabian StyleHeine, Sonja, Antonio Aguilar-Pimentel, Dennis Russkamp, Francesca Alessandrini, Valerie Gailus-Durner, Helmut Fuchs, Markus Ollert, Reinhard Bredehorst, Caspar Ohnmacht, Ulrich M. Zissler, and et al. 2022. "Thermosensitive PLGA–PEG–PLGA Hydrogel as Depot Matrix for Allergen-Specific Immunotherapy" Pharmaceutics 14, no. 8: 1527. https://doi.org/10.3390/pharmaceutics14081527