
Daily Intraperitoneal Administration of Rosiglitazone Does Not Improve Lung Function or Alveolarization in Preterm Rabbits Exposed to Hyperoxia

 , , , , and add
Show full author list
, , , , and add
Show full author list
Abstract
:1. Introduction
2. Materials and Methods
2.1. In Vitro Selectivity Assay for Peroxisome Proliferator-Activated Receptor of Thiazolidinediones
2.2. Plasma Protein Binding, Lung Tissue Binding, and Caco-2 Permeability Assay
2.2.1. Plasma Protein Binding
2.2.2. Lung Tissue Binding
2.2.3. Caco-2 Permeability Assay
2.2.4. Liquid Chromatography–Tandem Mass Spectrometry (LC/MS/MS) Method
2.3. Animal Care and Cesarean Section
2.4. Pharmacokinetic Profiling of Rosiglitazone
2.4.1. In Vivo Phase
2.4.2. Bioanalysis and Pharmacokinetic Analysis
2.5. Pulmonary Efficacy Studies: Daily, Intraperitoneal Rosiglitazone Administration at 1 mg/kg
2.5.1. Lung Function Testing
2.5.2. Radial Alveolar Count
2.6. Daily, Intraperitoneal Rosiglitazone Administration at 10 mg/kg
2.7. Quantification of Blood Lipid Levels
2.8. Quantitative Proteomic Analysis by Mass Spectrometry
2.9. Statistical Analysis
3. Results and Discussion
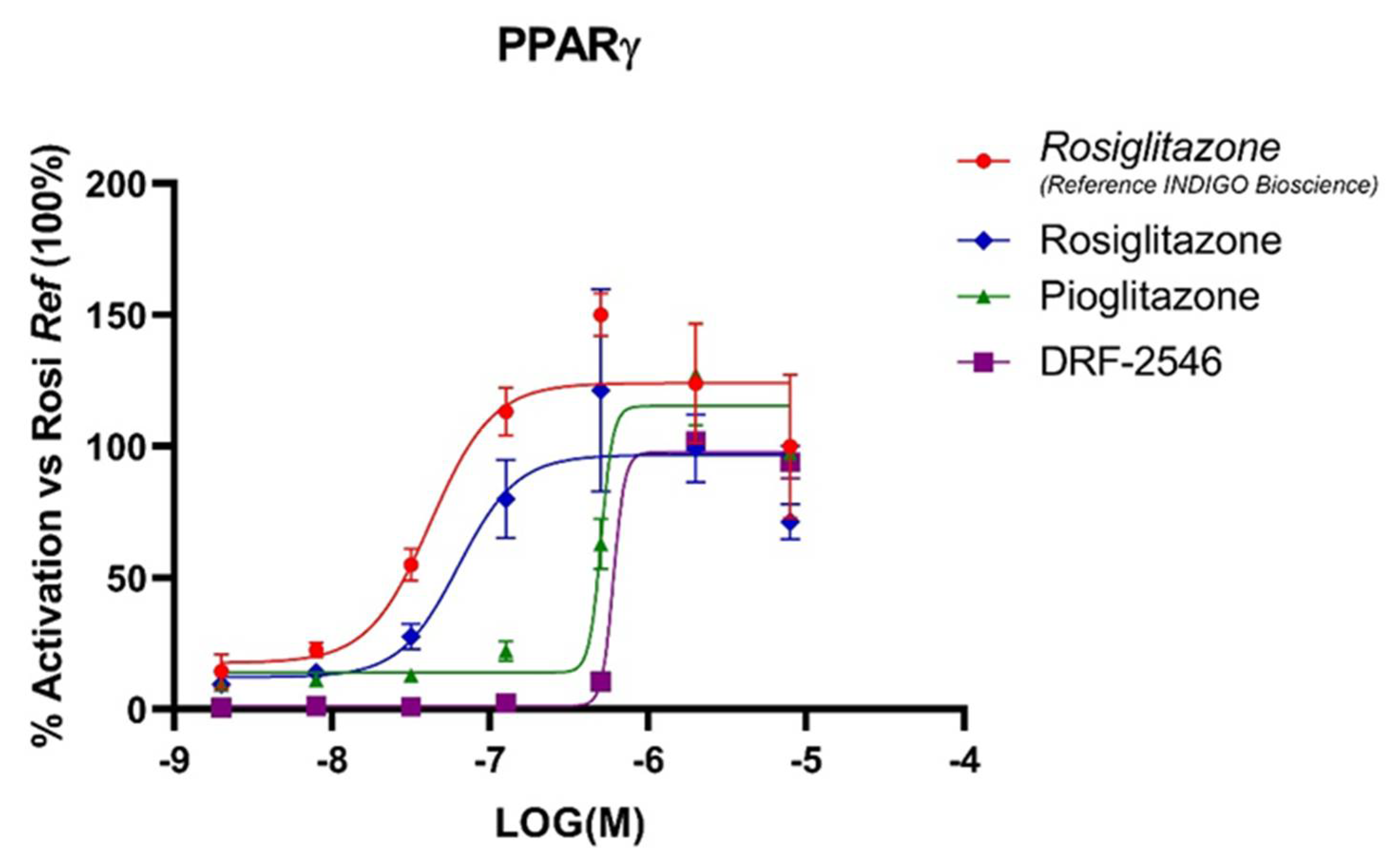
3.1. RGZ Has the Highest Affinity for Human PPARγ among TZDs
3.2. TZDs Show High Epithelial Permeability and Plasma Protein and Lung Tissue Binding
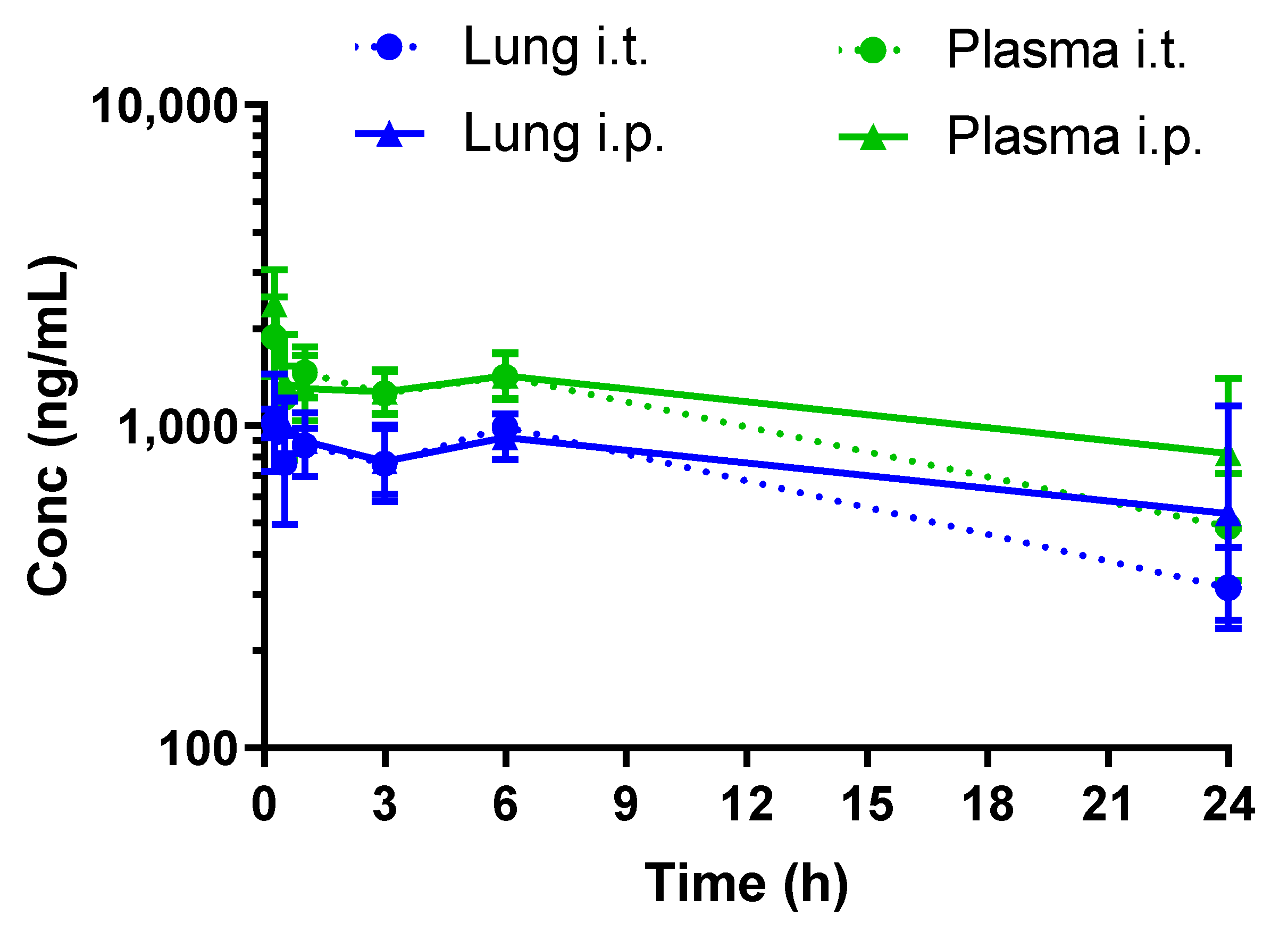
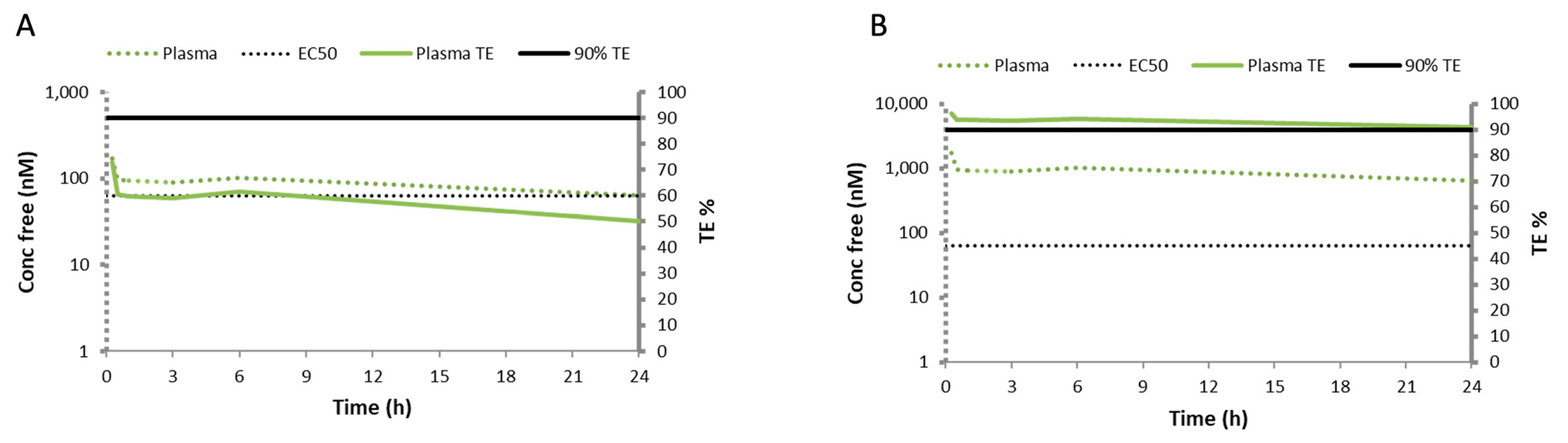
3.3. Rosiglitazone Displays a High Blood-to-Lung Delivery Irrespective of the Administration Route
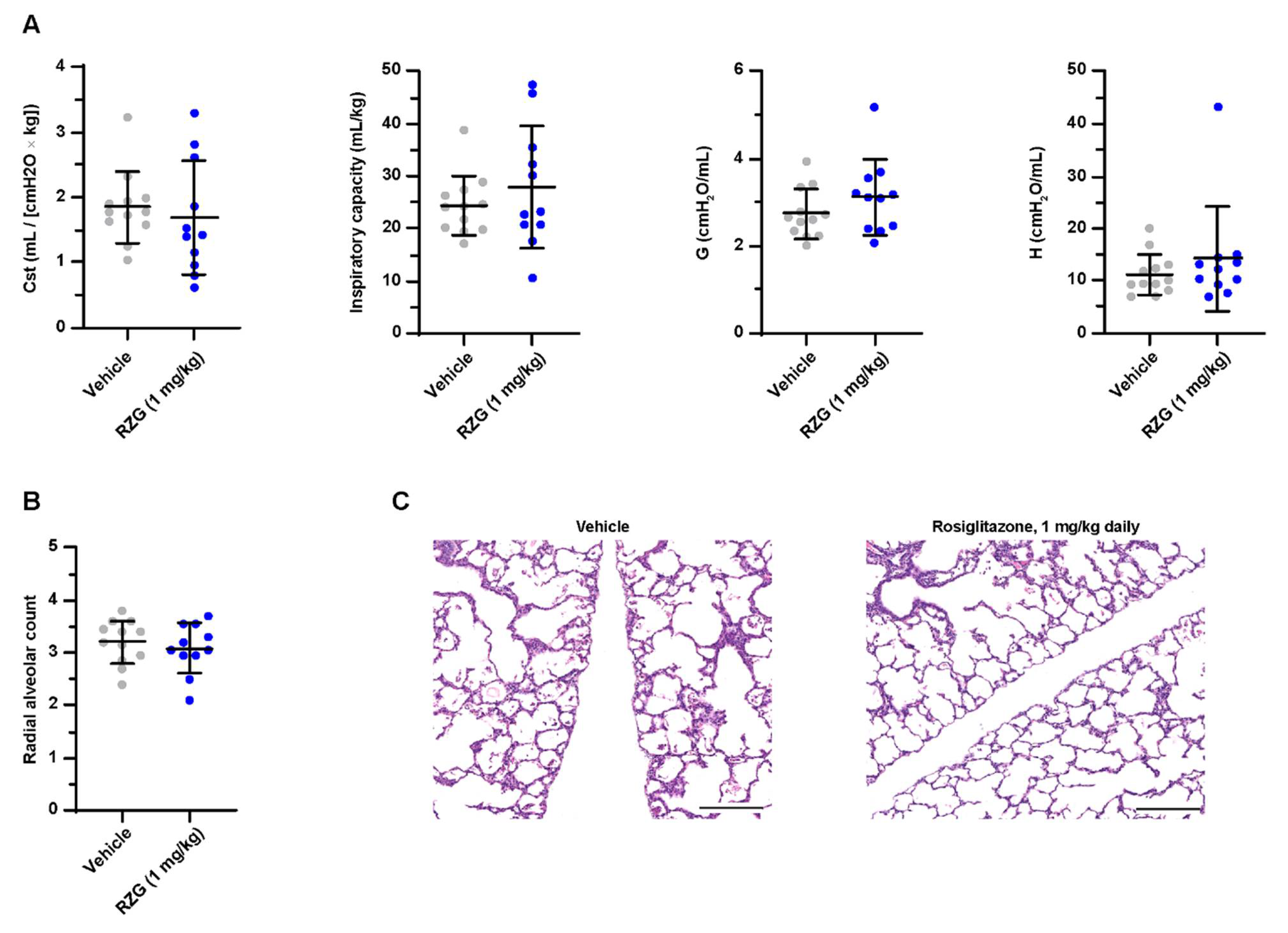
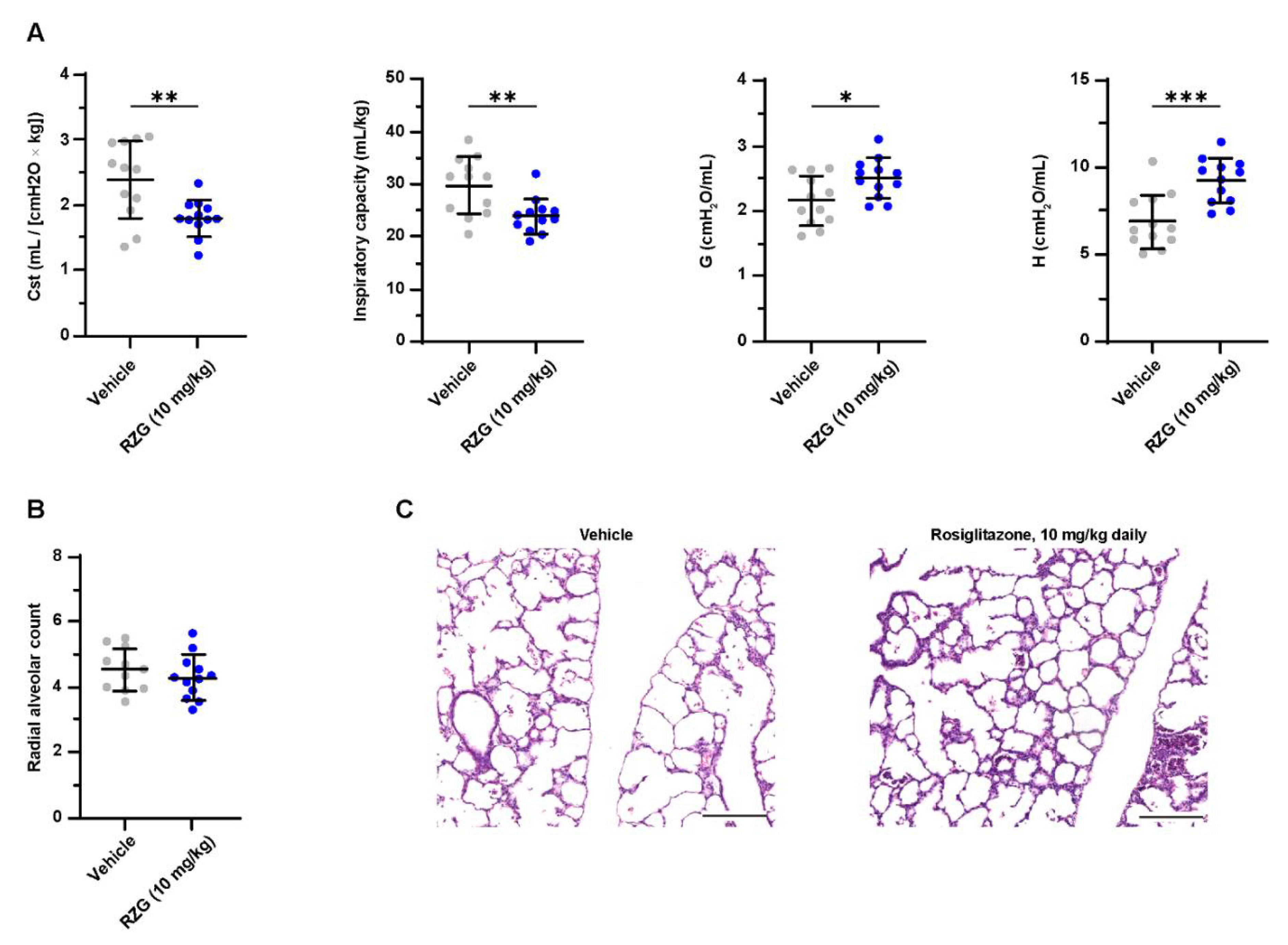
3.4. Daily Intraperitoneal Administration of Rosiglitazone Does Not Improve Lung Function or Alveolarization in Preterm Rabbits Exposed to Hyperoxia
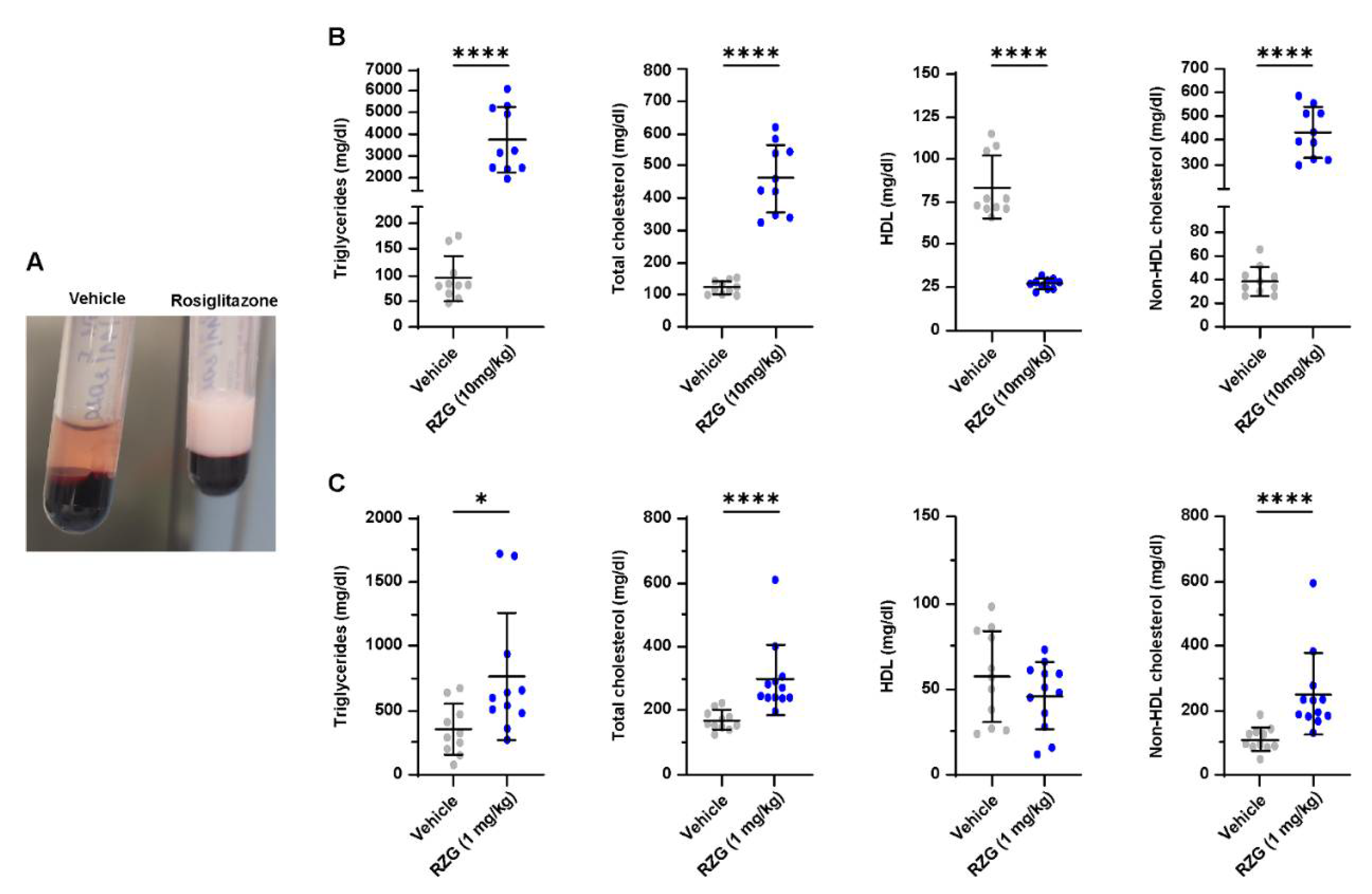
3.5. Daily Rosiglitazone Treatment Induces Dyslipidemia in Premature Rabbits
3.6. Lung Proteomics Reveal Dysregulation of Inflammatory Pathways and Protein–Lipid Complex by High-Dose Rosiglitazone
4. Limitations of the Study
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Thébaud, B.; Goss, K.N.; Laughon, M.; Whitsett, J.A.; Abman, S.H.; Steinhorn, R.H.; Aschner, J.L.; Davis, P.G.; McGrath-Morrow, S.A.; Soll, R.F.; et al. Bronchopulmonary dysplasia. Nat. Rev. Dis. Prim. 2019, 5, 78. [Google Scholar] [CrossRef] [PubMed]
- Sweet, D.G.; Carnielli, V.; Greisen, G.; Hallman, M.; Ozek, E.; Te Pas, A.; Plavka, R.; Roehr, C.C.; Saugstad, O.D.; Simeoni, U.; et al. European consensus guidelines on the management of respiratory distress syndrome – 2019 Update. Neonatology 2019, 115, 432–450. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Speer, C.P. Neonatal respiratory distress syndrome: An inflammatory disease? Neonatology 2011, 99, 316–319. [Google Scholar] [CrossRef] [PubMed]
- Hayes, D.; Feola, D.J.; Murphy, B.S.; Shook, L.A.; Ballard, H.O. Pathogenesis of bronchopulmonary dysplasia. Respiration 2010, 79, 425–436. [Google Scholar] [CrossRef]
- Rehan, V.K.; Torday, J.S. PPARγ signaling mediates the evolution, development, homeostasis, and repair of the lung. PPAR Res. 2012, 2012, 289867. [Google Scholar] [CrossRef] [Green Version]
- Grygiel-Górniak, B. Peroxisome proliferator-activated receptors and their ligands: Nutritional and clinical - a review. Nutr. J. 2014, 13, 17. [Google Scholar] [CrossRef] [Green Version]
- Rehan, V.K.; Torday, J.S. The lung alveolar lipofibroblast: An evolutionary strategy against neonatal hyperoxic lung injury. Antioxid. Redox Signal. 2014, 21, 1893–1904. [Google Scholar] [CrossRef] [Green Version]
- McGowan, S.E.; Torday, J.S. The pulmonary lipofibroblast (lipid interstitial cell) and its contributions to alveolar development. Annu. Rev. Physiol. 1997, 59, 43–62. [Google Scholar] [CrossRef]
- Mathew, R. Signaling pathways involved in the development of bronchopulmonary dysplasia and pulmonary hypertension. Children 2020, 7, 100. [Google Scholar] [CrossRef]
- Dasgupta, C.; Sakurai, R.; Wang, Y.; Guo, P.; Ambalavanan, N.; Torday, J.S.; Rehan, V.K. Hyperoxia-induced neonatal rat lung injury involves activation of TGF-β and Wnt signaling and is protected by rosiglitazone. Am. J. Physiol. Lung Cell Mol. Physiol. 2009, 296, 1031–1041. [Google Scholar] [CrossRef]
- Wu, S.; Savani, R.C. Chapter 1 - Molecular Bases for Lung Development, Injury, and Repair. In The Newborn Lung: Neonatology Questions and Controversies, 2nd ed.; Bancalari, E., Ed.; Elsevier: Philadelphia, PA, USA, 2012; Volume 1, pp. 3–28. [Google Scholar]
- Burgess, H.A.; Daugherty, L.E.; Thatcher, T.H.; Lakatos, H.F.; Ray, D.M.; Redonnet, M.; Phipps, R.P.; Sime, P.J. PPARγ agonists inhibit TGF-β induced pulmonary myofibroblast differentiation and collagen production: Implications for therapy of lung fibrosis. Am. J. Physiol. Lung Cell Mol. Physiol. 2005, 288, 1146–1153. [Google Scholar] [CrossRef] [Green Version]
- Lecarpentier, Y.; Gourrier, E.; Gobert, V.; Vallée, A. Bronchopulmonary dysplasia: Crosstalk between PPARγ, Wnt/β-catenin and TGF-β pathways; the potential therapeutic role of PPARγ agonists. Front. Pediatr. 2019, 7, 176. [Google Scholar] [CrossRef]
- Van Wijk, J.P.H.; De Koning, E.J.P.; Martens, E.P.; Rabelink, T.J. Thiazolidinediones and blood lipids in type 2 diabetes. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 1744–1749. [Google Scholar] [CrossRef]
- Lee, C.; Sakurai, R.; Shin, E.; Wang, Y.; Liu, J.; Rehan, V.K. Antenatal PPAR-γ agonist pioglitazone stimulates fetal lung maturation equally in males and females. Am. J. Physiol. Cell. Mol. Physiol. 2020, 319, 435–443. [Google Scholar] [CrossRef]
- Rehan, V.K.; Sakurai, R.; Corral, J.; Krebs, M.; Ibe, B.; Ihida-Stansbury, K.; Torday, J.S. Antenatally administered PPAR-γ agonist rosiglitazone prevents hyperoxia-induced neonatal rat lung injury. Am. J. Physiol. Lung Cell Mol. Physiol. 2010, 299, 672–680. [Google Scholar] [CrossRef] [Green Version]
- Richter, J.; Toelen, J.; Nagatomo, T.; Jimenez, J.; Vanoirbeek, J.; Deprest, J. Transplacental Administration of Rosiglitazone Attenuates Hyperoxic Lung Injury in a Preterm Rabbit Model. Fetal Diagn. Ther. 2016, 39, 297–305. [Google Scholar] [CrossRef]
- Ren, J.; Lock, M.C.; Darby, J.R.T.; Orgeig, S.; Holman, S.L.; Quinn, M.; Seed, M.; Muhlhausler, B.S.; McMillen, I.C.; Morrison, J.L. PPARγ activation in late gestation does not promote surfactant maturation in the fetal sheep lung. J. Dev. Orig. Health Dis. 2021, 12, 963–974. [Google Scholar] [CrossRef]
- Sakurai, R.; Lee, C.; Shen, H.; Waring, A.J.; Walther, F.J.; Rehan, V.K. A Combination of the Aerosolized PPAR-γ Agonist Pioglitazone and a Synthetic Surfactant Protein B Peptide Mimic Prevents Hyperoxia-Induced Neonatal Lung Injury in Rats. Neonatology 2018, 113, 296–304. [Google Scholar] [CrossRef]
- Lee, H.J.; Lee, Y.J.; Choi, C.W.; Lee, J.A.; Kim, E.K.; Kim, H.S.; Kim, B.I.; Choi, J.H. Rosiglitazone, a peroxisome proliferator-activated receptor-γ agonist, restores alveolar and pulmonary vascular development in a rat model of bronchopulmonary dysplasia. Yonsei Med. J. 2014, 55, 99–106. [Google Scholar] [CrossRef] [Green Version]
- Morales, E.; Sakurai, R.; Husain, S.; Paek, D.; Gong, M.; Ibe, B.; Li, Y.; Husain, M.; Torday, J.S.; Rehan, V.K. Nebulized PPARγ agonists: A novel approach to augment neonatal lung maturation and injury repair in rats. Pediatr. Res. 2014, 75, 631–640. [Google Scholar] [CrossRef]
- Takeda, K.; Okamoto, M.; De Langhe, S.; Dill, E.; Armstrong, M.; Reisdorf, N.; Irwin, D.; Koster, M.; Wilder, J.; Stenmark, K.R.; et al. Peroxisome proliferator-activated receptor-g agonist treatment increases septation and angiogenesis and decreases airway hyperresponsiveness in a model of experimental neonatal chronic lung disease. Anat. Rec. 2009, 292, 1045–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, Y.; Santos, J.; Sakurai, R.; Shin, E.; Cerny, L.; Torday, J.S.; Rehan, V.K. Peroxisome proliferator-activated receptor γ agonists enhance lung maturation in a neonatal rat model. Pediatr. Res. 2009, 65, 150–155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehan, V.K.; Wang, Y.; Patel, S.; Santos, J.; Torday, J.S. Rosiglitazone, a peroxisome proliferator-activated receptor-γ agonist, prevents hyperoxia-induced neonatal rat lung injury in vivo. Pediatr. Pulmonol. 2006, 41, 558–569. [Google Scholar] [CrossRef]
- Salaets, T.; Gie, A.; Tack, B.; Deprest, J.; Toelen, J. Modelling bronchopulmonary dysplasia in animals: Arguments for the preterm rabbit model. Curr. Pharm. Des. 2017, 23, 5887–5901. [Google Scholar] [CrossRef] [PubMed]
- Gie, A.G.; Regin, Y.; Salaets, T.; Casiraghi, C.; Salomone, F.; Deprest, J.; Vanoirbeek, J.; Toelen, J. Intratracheal budesonide/surfactant attenuates hyperoxia-induced lung injury in preterm rabbits. Am. J. Physiol. Cell. Mol. Physiol. 2020, 319, 949–956. [Google Scholar] [CrossRef] [PubMed]
- Richter, J.; Jimenez, J.; Nagatomo, T.; Toelen, J.; Brady, P.; Salaets, T.; Lesage, F.; Vanoirbeek, J.; Deprest, J. Proton-pump inhibitor omeprazole attenuates hyperoxia induced lung injury. J. Transl. Med. 2016, 14, 247. [Google Scholar] [CrossRef] [Green Version]
- Jiménez, J.; Richter, J.; Nagatomo, T.; Salaets, T.; Quarck, R.; Wagennar, A.; Wang, H.; Vanoirbeek, J.; Deprest, J.; Toelen, J. Progressive vascular functional and structural damage in a bronchopulmonary dysplasia model in preterm rabbits exposed to hyperoxia. Int. J. Mol. Sci. 2016, 17, 1776. [Google Scholar] [CrossRef]
- Salaets, T.; Gie, A.; Jimenez, J.; Aertgeerts, M.; Gheysens, O.; Vande Velde, G.; Koole, M.; Murgia, X.; Casiraghi, C.; Ricci, F.; et al. Local pulmonary drug delivery in the preterm rabbit: Feasibility and efficacy of daily intratracheal injections. Am. J. Physiol. Cell. Mol. Physiol. 2019, 316, 589–597. [Google Scholar] [CrossRef]
- Cooney, T.P.; Thurlbeck, W.M. The radial alveolar count method of Emery and Mithal: A reappraisal 2--intrauterine and early postnatal lung growth. Thorax 1982, 37, 580–583. [Google Scholar] [CrossRef] [Green Version]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The Perseus computational platform for comprehensive analysis of (prote)omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, B.; Pache, L.; Chang, M.; Khodabakhshi, A.H.; Tanaseichuk, O.; Benner, C.; Chanda, S.K. Metascape provides a biologist-oriented resource for the analysis of systems-level datasets. Nat. Commun. 2019, 10, 1523. [Google Scholar] [CrossRef] [PubMed]
- Deeg, M.A.; Tan, M.H. Pioglitazone versus Rosiglitazone: Effects on lipids, lipoproteins, and apolipoproteins in head-to-head randomized clinical studies. PPAR Res. 2008, 2008, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Form 20-F Dr Reddys Laboratories Ltd. Available online: https://www.sec.gov/Archives/edgar/data/1135951/000119312513286265/d563639d20f.htm (accessed on 12 July 2022).
- Madhavan, G.R.; Chakrabarti, R.; Kumar, S.K.B.; Misra, P.; Mamidi, R.N.V.S.; Balraju, V.; Kasiram, K.; Babu, R.K.; Suresh, J.; Lohray, B.B.; et al. Novel phthalazinone and benzoxazinone containing thiazolidinediones as antidiabetic and hypolipidemic agents. Eur. J. Med. Chem. 2001, 36, 627–637. [Google Scholar] [CrossRef]
- Thangavel, N.; Al Bratty, M.; Akhtar Javed, S.; Ahsan, W.; Alhazmi, H.A. Targeting Peroxisome Proliferator-Activated Receptors Using Thiazolidinediones: Strategy for Design of Novel Antidiabetic Drugs. Int. J. Med. Chem. 2017, 2017, 1069718. [Google Scholar] [CrossRef] [Green Version]
- Salaets, T.; Aertgeerts, M.; Gie, A.; Vignero, J.; de Winter, D.; Regin, Y.; Jimenez, J.; Vande Velde, G.; Allegaert, K.; Deprest, J.; et al. Preterm birth impairs postnatal lung development in the neonatal rabbit model. Respir. Res. 2020, 21, 59. [Google Scholar] [CrossRef] [Green Version]
- GlaxoSmithKline Avandia: FDA—Avandia Prescribing Information. 2007, 1–34. Available online: https://www.accessdata.fda.gov/drugsatfda_docs/label/2008/021071s034lbl.pdf (accessed on 12 July 2022).
- Hong, F.; Pan, S.; Guo, Y.; Xu, P.; Zhai, Y. PPARs as nuclear receptors for nutrient and energy metabolism. Molecules 2019, 24, 2545. [Google Scholar] [CrossRef] [Green Version]
- Krishna, S.; Cheng, B.; Sharma, D.R.; Yadav, S.; Stempinski, E.S.; Mamtani, S.; Shah, E.; Deo, A.; Acherjee, T.; Thomas, T.; et al. PPAR-γ activation enhances myelination and neurological recovery in premature rabbits with intraventricular hemorrhage. Proc. Natl. Acad. Sci. USA 2021, 118, 1–12. [Google Scholar] [CrossRef]
- Nissen, S.E.; Wolski, K. Effect of rosiglitazone on the risk of myocardial infarction and death from cardiovascular causes. N. Engl. J. Med. 2007, 356, 2457–2471. [Google Scholar] [CrossRef] [Green Version]
- Giretti, I.; D’Ascenzo, R.; Correani, A.; Antognoli, L.; Monachesi, C.; Biagetti, C.; Pompilio, A.; Marinelli, L.; Burattini, I.; Cogo, P.; et al. Hypertriglyceridemia and lipid tolerance in preterm infants with a birth weight of less than 1250 g on routine parenteral nutrition. Clin. Nutr. 2021, 40, 4444–4448. [Google Scholar] [CrossRef]
- Salaets, T.; Richter, J.; Brady, P.; Jimenez, J.; Nagatomo, T.; Deprest, J.; Toelen, J. Transcriptome analysis of the preterm rabbit lung after seven days of hyperoxic exposure. PLoS ONE 2015, 10, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, A.K.; Gopal, V.Y.N.; Menon, R.T.; Hagan, J.L.; Huang, S.; Shivanna, B. Lung omics signatures in a bronchopulmonary dysplasia and pulmonary hypertension-like murine model. Am. J. Physiol. Cell. Mol. Physiol. 2018, 315, 734–741. [Google Scholar] [CrossRef] [PubMed]
- Salaets, T.; Tack, B.; Jimenez, J.; Gie, A.; Lesage, F.; de Winter, D.; Berghen, N.; Allegaert, K.; Deprest, J.; Toelen, J. Simvastatin attenuates lung functional and vascular effects of hyperoxia in preterm rabbits. Pediatr. Res. 2019, 87, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Goldberg, R.B.; Kendall, D.M.; Deeg, M.A.; Buse, J.B.; Zagar, A.J.; Pinaire, J.A.; Tan, M.H.; Khan, M.A.; Perez, A.T.; Jacober, S.J.; et al. A Comparison of Lipid and Glycemic Effects of Pioglitazone and Rosiglitazone in Patients with Type 2 Diabetes and Dyslipidemia. Diabetes Care 2005, 28, 1547–1554. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TZD Type | Plasma Protein Binding (%) | Lung Tissue Binding (%) | Papp (A–B/B–A, cm/sec) | Papp with Pgp Inhibitor (A–B/B–A, cm/sec) |
|---|---|---|---|---|
| RGZ | 97.5 ± 0.2 | 94.2 ± 0.7 | 1.45 × 10−5 ± 0.06 × 10−5 / 1.00 × 10−5 ± 0.06 × 10−5 | 1.63 × 10−5 ± 0.05 × 10−5 / 1.14 ×10−5 ± 0.04 × 10−5 |
| PGZ | 96.7 ± 0.6 | 95.6 ± 2.1 | 1.18 × 10−5 ± 0.05 × 10−5 / 1.53 × 10−5 ± 0.36 × 10−5 | 1.23 × 10−5 ± 0.17 × 10−5 / 2.54 × 10−5 ± 0.55 × 10−5 |
| DRF-2546 | 98.4 ± 0.3 | 91.8 ± 1.8 | 2.29 × 10−5 ± 0.03 × 10−5 / 1.87 × 10−5 ± 0.80 × 10−5 | 2.86 × 10−5 ± 0.15× 10−5 / 1.58 × 10−5 ± 0.34 × 10−5 |
| Tmax (h) | Cmax (ng/mL) | t1/2 (h) | AUClast (ng/mL∙h) | AUCLung/AUCPlasma | |
|---|---|---|---|---|---|
| Intratracheal Administration | |||||
| Lung | 0.25 | 1067 | 15 | 17,020 | 0.7 |
| Plasma | 0.25 | 1950 | 15 | 25,785 | |
| Intraperitoneal Administration | |||||
| Lung | 0.25 | 1024 | n.c. | 19,752 | 0.7 |
| Plasma | 0.25 | 2443 | n.c. | 29,490 |
| Pathway Description | Downregulated Log (q-Value) | Upregulated Log (q-Value) | Dysregulated Proteins in the Pathway *** |
|---|---|---|---|
| 1.Neutrophil degranulation | - | −12.0 | AHSG, CAMP, CHIT1, FTL, HK3, HP, ITGAM, LCN2, LTF, MPO, CFP, S100A8, S100A9, S100A12, PGLYRP1, SNAP29, GCA, PYCARD, RETN |
| 2. MCM * complex | −7.6 | - | MCM2, MCM3, MCM4, MCM5, MCM7 |
| 3. Regulation of inflammatory response | - | −7.1 | AGT, AHSG, APOE, IL16, LBP, MVK, S100A8, S100A9, S100A12, SNCA, PGLYRP1, PYCARD, PGLYRP2 |
| 4. Humoral immune response | - | −7.0 | C6, C8A, C8B, C8G, CAMP, CRP, HPX, LTF, CFP, S100A9, S100A12, PGLYRP1 |
| 5. Transition metal ion homeostasis | −5.4 | FTL, HPX, LCN2, LTF, S100A8, S100A9, ABCB6, STEAP4 | |
| 6. Positive regulation of reactive oxygen species metabolic process | - | −4.7 | AGT, CRP, ITGAM, LCN2, SNCA, XDH |
| 7. Protein–lipid complex remodeling | - | −5.0 | AGT, APOB, APOC3, APOE, MPO |
| 8. Regulation of endopeptidase activity | −5.0 | AGT, AHSG, BAX, LTF, S100A8, S100A9, SNCA, XDH, LAMTOR5, FETUB, PYCARD | |
| 9. Extracellular matrix organization | −5.1 | COL4A6, HAPLN1, DCN, FBLN2, FBN2, ICAM2, ITGA6, ITGB3, SPARC, PXDN, CRTAP | |
| 10. NABA CORE MATRISOME ** | −4.6 | COL4A6, HAPLN1, DCN, FBLN2, FBN2, SPARC, PXDN, SPARCL1, POSTN, NPNT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aquila, G.; Regin, Y.; Murgia, X.; Salomone, F.; Casiraghi, C.; Catozzi, C.; Scalera, E.; Storti, M.; Stretti, F.; Aquino, G.; et al. Daily Intraperitoneal Administration of Rosiglitazone Does Not Improve Lung Function or Alveolarization in Preterm Rabbits Exposed to Hyperoxia. Pharmaceutics 2022, 14, 1507. https://doi.org/10.3390/pharmaceutics14071507
Aquila G, Regin Y, Murgia X, Salomone F, Casiraghi C, Catozzi C, Scalera E, Storti M, Stretti F, Aquino G, et al. Daily Intraperitoneal Administration of Rosiglitazone Does Not Improve Lung Function or Alveolarization in Preterm Rabbits Exposed to Hyperoxia. Pharmaceutics. 2022; 14(7):1507. https://doi.org/10.3390/pharmaceutics14071507
Chicago/Turabian StyleAquila, Giorgio, Yannick Regin, Xabier Murgia, Fabrizio Salomone, Costanza Casiraghi, Chiara Catozzi, Enrica Scalera, Matteo Storti, Francesca Stretti, Giancarlo Aquino, and et al. 2022. "Daily Intraperitoneal Administration of Rosiglitazone Does Not Improve Lung Function or Alveolarization in Preterm Rabbits Exposed to Hyperoxia" Pharmaceutics 14, no. 7: 1507. https://doi.org/10.3390/pharmaceutics14071507