
Research Progress and Potential Applications of Spermidine in Ocular Diseases


Abstract
:1. Introduction
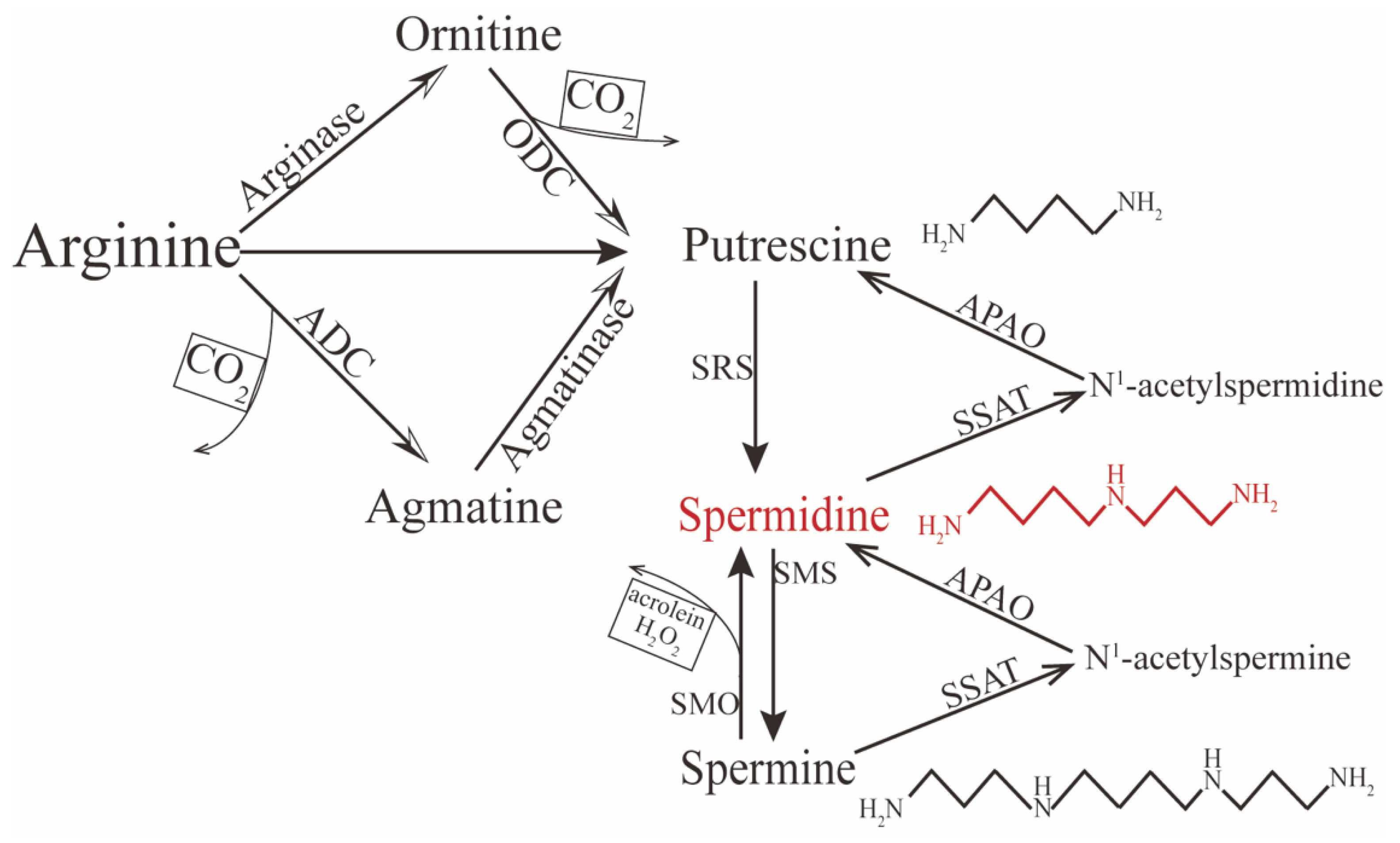
2. Metabolism of Spermidine
3. Biofunctions of Spermidine
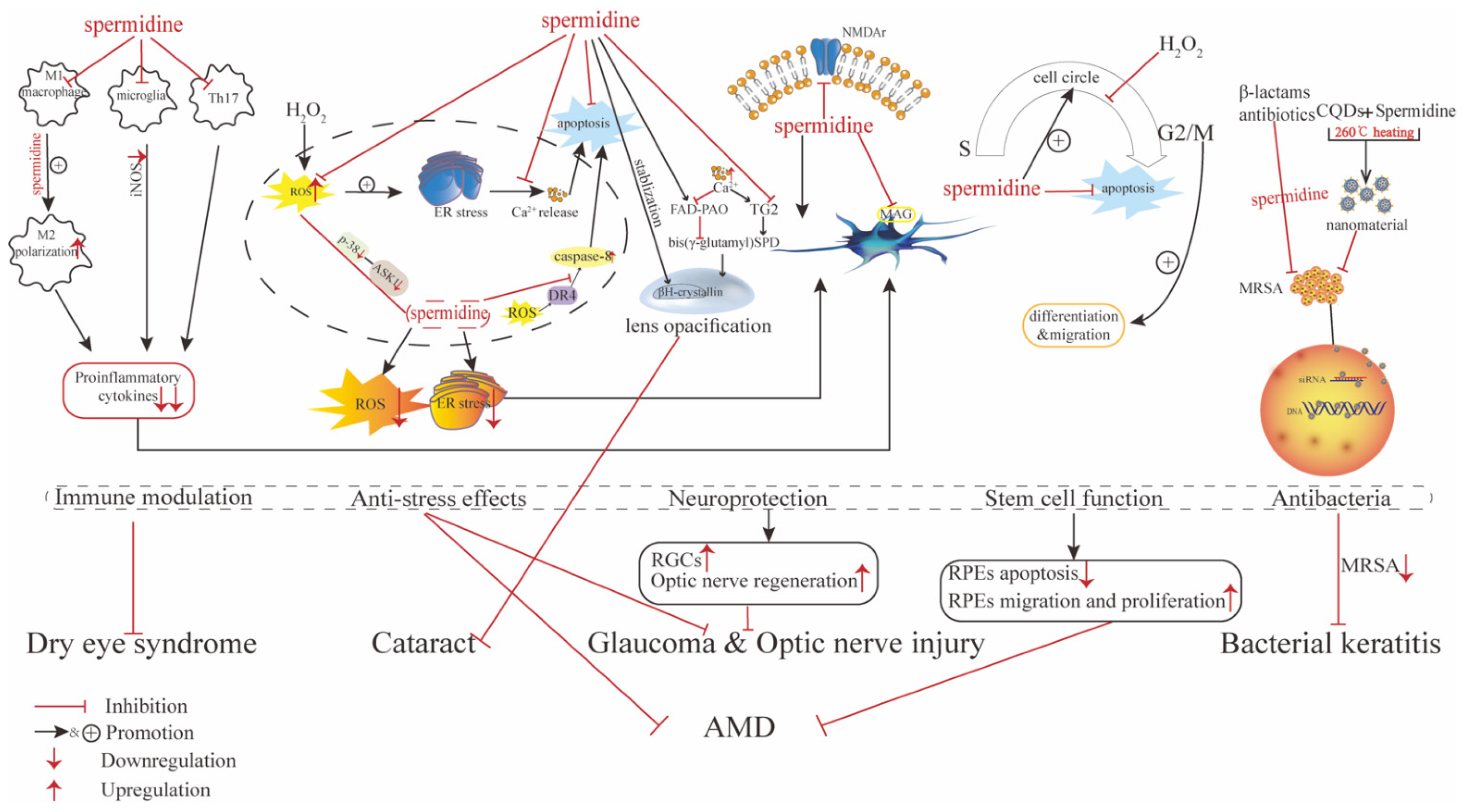
4. Role of Spermidine in Ocular Diseases
4.1. Role of Spermidine in Ocular Surface Disorders
4.1.1. Dry Eye Syndrome
4.1.2. Bacterial Keratitis
4.1.3. Keratoconus (KC)
4.2. Role of Spermidine in Cataracts
4.3. Role of Spermidine in Uveitis
4.4. Role of Spermidine in Ocular Neuroprotection and Retinal Disorders
4.4.1. Glaucoma, Optic Nerve Injury, and Retinal Nerve Degeneration
4.4.2. Retina Degeneration Induced by Different Causes
4.4.3. Altered Level of Spermidine in Proliferative Disorders of the Retina
5. Conclusions and Open Questions for Further Study
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Patocka, J.; Kuehn, G.D. Natural Polyamines and Their Biological Consequence in Mammals. Acta Med. (Hradec Kral.) 2000, 43, 119–124. [Google Scholar] [CrossRef] [Green Version]
- Partridge, L.; Fuentealba, M.; Kennedy, B.K. The Quest to Slow Ageing through Drug Discovery. Nat. Rev. Drug Discov. 2020, 19, 513–532. [Google Scholar] [CrossRef]
- Eisenberg, T.; Abdellatif, M.; Schroeder, S.; Primessnig, U.; Stekovic, S.; Pendl, T.; Harger, A.; Schipke, J.; Zimmermann, A.; Schmidt, A.; et al. Cardioprotection and Lifespan Extension by the Natural Polyamine Spermidine. Nat. Med. 2016, 22, 1428–1438. [Google Scholar] [CrossRef]
- Hofer, S.J.; Davinelli, S.; Bergmann, M.; Scapagnini, G.; Madeo, F. Caloric Restriction Mimetics in Nutrition and Clinical Trials. Front. Nutr. 2021, 8, 717343. [Google Scholar] [CrossRef] [PubMed]
- Schroeder, S.; Hofer, S.J.; Zimmermann, A.; Pechlaner, R.; Dammbrueck, C.; Pendl, T.; Marcello, G.M.; Pogatschnigg, V.; Bergmann, M.; Müller, M.; et al. Dietary Spermidine Improves Cognitive Function. Cell Rep. 2021, 35, 108985. [Google Scholar] [CrossRef]
- Kang, X.; Li, C.; Xie, Y.; He, L.-L.; Xiao, F.; Zhan, K.-B.; Tang, Y.-Y.; Li, X.; Tang, X.-Q. Hippocampal Ornithine Decarboxylase/Spermidine Pathway Mediates H2S-Alleviated Cognitive Impairment in Diabetic Rats: Involving Enhancment of Hippocampal Autophagic Flux. J. Adv. Res. 2021, 27, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Yin, J.; Zhang, L.; Qi, C.-C.; Ma, Z.-L.; Gao, L.-P.; Wang, D.-G.; Jing, Y.-H. Spermidine Preconditioning Ameliorates Laurate-Induced Brain Injury by Maintaining Mitochondrial Stability. Neurol. Res. 2017, 39, 248–258. [Google Scholar] [CrossRef] [PubMed]
- Eisenberg, T.; Abdellatif, M.; Zimmermann, A.; Schroeder, S.; Pendl, T.; Harger, A.; Stekovic, S.; Schipke, J.; Magnes, C.; Schmidt, A.; et al. Dietary Spermidine for Lowering High Blood Pressure. Autophagy 2017, 13, 767–769. [Google Scholar] [CrossRef] [PubMed]
- Messerer, J.; Wrede, C.; Schipke, J.; Brandenberger, C.; Abdellatif, M.; Eisenberg, T.; Madeo, F.; Sedej, S.; Mühlfeld, C. Spermidine Supplementation Influences Mitochondrial Number and Morphology in the Heart of Aged Mice. J. Anat. 2021, in press. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, A.; Kurihara, S.; Takahashi, D.; Ohashi, W.; Nakamura, Y.; Kimura, S.; Onuki, M.; Kume, A.; Sasazawa, Y.; Furusawa, Y.; et al. Symbiotic Polyamine Metabolism Regulates Epithelial Proliferation and Macrophage Differentiation in the Colon. Nat. Commun. 2021, 12, 2105. [Google Scholar] [CrossRef]
- Ma, L.; Ni, L.; Yang, T.; Mao, P.; Huang, X.; Luo, Y.; Jiang, Z.; Hu, L.; Zhao, Y.; Fu, Z.; et al. Preventive and Therapeutic Spermidine Treatment Attenuates Acute Colitis in Mice. J. Agric. Food Chem. 2021, 69, 1864–1876. [Google Scholar] [CrossRef] [PubMed]
- Morón, B.; Spalinger, M.; Kasper, S.; Atrott, K.; Frey-Wagner, I.; Fried, M.; McCole, D.F.; Rogler, G.; Scharl, M. Activation of Protein Tyrosine Phosphatase Non-Receptor Type 2 by Spermidine Exerts Anti-Inflammatory Effects in Human THP-1 Monocytes and in a Mouse Model of Acute Colitis. PLoS ONE 2013, 8, e73703. [Google Scholar] [CrossRef] [Green Version]
- Johnson, D.A.; Fields, C.; Fallon, A.; Fitzgerald, M.E.C.; Viar, M.J.; Johnson, L.R. Polyamine-Dependent Migration of Retinal Pigment Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2002, 43, 1228–1233. [Google Scholar] [PubMed]
- Kim, D.H.; Kim, J.-H.; Hwangbo, H.; Kim, S.Y.; Ji, S.Y.; Kim, M.Y.; Cha, H.-J.; Park, C.; Hong, S.H.; Kim, G.-Y.; et al. Spermidine Attenuates Oxidative Stress-Induced Apoptosis via Blocking Ca2+ Overload in Retinal Pigment Epithelial Cells Independently of ROS. Int. J. Mol. Sci. 2021, 22, 1361. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Rao, J.N.; Liu, L.; Zou, T.-T.; Turner, D.J.; Bass, B.L.; Wang, J.-Y. Regulation of Adherens Junctions and Epithelial Paracellular Permeability: A Novel Function for Polyamines. Am. J. Physiol. Cell Physiol. 2003, 285, C1174–C1187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noro, T.; Namekata, K.; Azuchi, Y.; Kimura, A.; Guo, X.; Harada, C.; Nakano, T.; Tsuneoka, H.; Harada, T. Spermidine Ameliorates Neurodegeneration in a Mouse Model of Normal Tension Glaucoma. Investig. Ophthalmol. Vis. Sci. 2015, 56, 5012–5019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, H.; Kim, D.H.; Hwangbo, H.; Kim, S.Y.; Ji, S.Y.; Kim, M.Y.; Shim, J.-H.; Leem, S.-H.; Hyun, J.W.; Kim, G.-Y.; et al. The Protective Effect of Topical Spermidine on Dry Eye Disease with Retinal Damage Induced by Diesel Particulate Matter2.5. Pharmaceutics 2021, 13, 1439. [Google Scholar] [CrossRef]
- Jian, H.-J.; Wu, R.-S.; Lin, T.-Y.; Li, Y.-J.; Lin, H.-J.; Harroun, S.G.; Lai, J.-Y.; Huang, C.-C. Super-Cationic Carbon Quantum Dots Synthesized from Spermidine as an Eye Drop Formulation for Topical Treatment of Bacterial Keratitis. ACS Nano 2017, 11, 6703–6716. [Google Scholar] [CrossRef]
- Lentini, A.; Tabolacci, C.; Mattioli, P.; Provenzano, B.; Beninati, S. Spermidine Delays Eye Lens Opacification in Vitro by Suppressing Transglutaminase-Catalyzed Crystallin Cross-Linking. Protein J. 2011, 30, 109–114. [Google Scholar] [CrossRef]
- Deng, K.; He, H.; Qiu, J.; Lorber, B.; Bryson, J.B.; Filbin, M.T. Increased Synthesis of Spermidine as a Result of Upregulation of Arginase I Promotes Axonal Regeneration in Culture and in Vivo. J. Neurosci. Off. J. Soc. Neurosci. 2009, 29, 9545–9552. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Wang, B.; Liu, H.; Wang, H.; Yin, P.; Dong, W.; Li, J.; Wang, Y.X.; Yusufu, M.; Briant, P.; et al. Prevalence and Causes of Vision Loss in China from 1990 to 2019: Findings from the Global Burden of Disease Study 2019. Lancet Public Health 2020, 5, e682–e691. [Google Scholar] [CrossRef]
- Taryam, M.O.; Rabiu, M.M.; Muhammad, N.; Oladigbolu, K.; Abdurrahman, H. Prevalence and Causes of Blindness and Visual Impairment; and Cataract Surgical Services in Katsina State of Nigeria. Br. J. Ophthalmol. 2020, 104, 752–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madeo, F.; Eisenberg, T.; Pietrocola, F.; Kroemer, G. Spermidine in Health and Disease. Science 2018, 359, eaan2788. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muñoz-Esparza, N.C.; Latorre-Moratalla, M.L.; Comas-Basté, O.; Toro-Funes, N.; Veciana-Nogués, M.T.; Vidal-Carou, M.C. Polyamines in Food. Front. Nutr. 2019, 6, 108. [Google Scholar] [CrossRef]
- Hofer, S.J.; Liang, Y.; Zimmermann, A.; Schroeder, S.; Dengjel, J.; Kroemer, G.; Eisenberg, T.; Sigrist, S.J.; Madeo, F. Spermidine-Induced Hypusination Preserves Mitochondrial and Cognitive Function during Aging. Autophagy 2021, 17, 2037–2039. [Google Scholar] [CrossRef]
- Kalač, P. Health Effects and Occurrence of Dietary Polyamines: A Review for the Period 2005-Mid 2013. Food Chem. 2014, 161, 27–39. [Google Scholar] [CrossRef]
- Soulet, D.; Gagnon, B.; Rivest, S.; Audette, M.; Poulin, R. A Fluorescent Probe of Polyamine Transport Accumulates into Intracellular Acidic Vesicles via a Two-Step Mechanism. J. Biol. Chem. 2004, 279, 49355–49366. [Google Scholar] [CrossRef] [Green Version]
- Moriyama, Y.; Hatano, R.; Moriyama, S.; Uehara, S. Vesicular Polyamine Transporter as a Novel Player in Amine-Mediated Chemical Transmission. Biochim. Biophys. Acta Biomembr. 2020, 1862, 183208. [Google Scholar] [CrossRef]
- Pegg, A.E. Functions of Polyamines in Mammals. J. Biol. Chem. 2016, 291, 14904–14912. [Google Scholar] [CrossRef] [Green Version]
- Malpica-Nieves, C.J.; Rivera-Aponte, D.E.; Tejeda-Bayron, F.A.; Mayor, A.M.; Phanstiel, O.; Veh, R.W.; Eaton, M.J.; Skatchkov, S.N. The Involvement of Polyamine Uptake and Synthesis Pathways in the Proliferation of Neonatal Astrocytes. Amino Acids 2020, 52, 1169–1180. [Google Scholar] [CrossRef]
- Fan, J.; Feng, Z.; Chen, N. Spermidine as a Target for Cancer Therapy. Pharmacol. Res. 2020, 159, 104943. [Google Scholar] [CrossRef] [PubMed]
- Çelik, V.K.; Kapancık, S.; Kaçan, T.; Kaçan, S.B.; Kapancık, S.; Kılıçgün, H. Serum Levels of Polyamine Synthesis Enzymes Increase in Diabetic Patients with Breast Cancer. Endocr. Connect. 2017, 6, 574–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pegg, A.E.; Casero, R.A. Current Status of the Polyamine Research Field. Methods Mol. Biol. Clifton N. J. 2011, 720, 3–35. [Google Scholar] [CrossRef] [Green Version]
- Alfarhan, M.; Liu, F.; Shan, S.; Pichavaram, P.; Somanath, P.R.; Narayanan, S.P. Pharmacological Inhibition of Spermine Oxidase Suppresses Excitotoxicity Induced Neuroinflammation in Mouse Retina. Int. J. Mol. Sci. 2022, 23, 2133. [Google Scholar] [CrossRef]
- Muth, A.; Madan, M.; Archer, J.J.; Ocampo, N.; Rodriguez, L.; Phanstiel, O. Polyamine Transport Inhibitors: Design, Synthesis, and Combination Therapies with Difluoromethylornithine. J. Med. Chem. 2014, 57, 348–363. [Google Scholar] [CrossRef]
- Wang, Y.; Devereux, W.; Woster, P.M.; Stewart, T.M.; Hacker, A.; Casero, R.A. Cloning and Characterization of a Human Polyamine Oxidase That Is Inducible by Polyamine Analogue Exposure. Cancer Res. 2001, 61, 5370–5373. [Google Scholar]
- Kaushik, S.; Tasset, I.; Arias, E.; Pampliega, O.; Wong, E.; Martinez-Vicente, M.; Cuervo, A.M. Autophagy and the Hallmarks of Aging. Ageing Res. Rev. 2021, 72, 101468. [Google Scholar] [CrossRef]
- Ramot, Y.; Tiede, S.; Bíró, T.; Abu Bakar, M.H.; Sugawara, K.; Philpott, M.P.; Harrison, W.; Pietilä, M.; Paus, R. Spermidine Promotes Human Hair Growth and Is a Novel Modulator of Human Epithelial Stem Cell Functions. PLoS ONE 2011, 6, e22564. [Google Scholar] [CrossRef]
- D’Adamo, S.; Cetrullo, S.; Guidotti, S.; Silvestri, Y.; Minguzzi, M.; Santi, S.; Cattini, L.; Filardo, G.; Flamigni, F.; Borzì, R.M. Spermidine Rescues the Deregulated Autophagic Response to Oxidative Stress of Osteoarthritic Chondrocytes. Free Radic. Biol. Med. 2020, 153, 159–172. [Google Scholar] [CrossRef]
- Liu, R.; Li, X.; Ma, H.; Yang, Q.; Shang, Q.; Song, L.; Zheng, Z.; Zhang, S.; Pan, Y.; Huang, P.; et al. Spermidine Endows Macrophages Anti-Inflammatory Properties by Inducing Mitochondrial Superoxide-Dependent AMPK Activation, Hif-1α Upregulation and Autophagy. Free Radic. Biol. Med. 2020, 161, 339–350. [Google Scholar] [CrossRef]
- Carriche, G.M.; Almeida, L.; Stüve, P.; Velasquez, L.; Dhillon-LaBrooy, A.; Roy, U.; Lindenberg, M.; Strowig, T.; Plaza-Sirvent, C.; Schmitz, I.; et al. Regulating T-Cell Differentiation through the Polyamine Spermidine. J. Allergy Clin. Immunol. 2021, 147, 335–348. [Google Scholar] [CrossRef] [PubMed]
- Baek, A.R.; Hong, J.; Song, K.S.; Jang, A.S.; Kim, D.J.; Chin, S.S.; Park, S.W. Spermidine Attenuates Bleomycin-Induced Lung Fibrosis by Inducing Autophagy and Inhibiting Endoplasmic Reticulum Stress (ERS)-Induced Cell Death in Mice. Exp. Mol. Med. 2020, 52, 2034–2045. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Li, S.; Wang, J.; Wu, F.; Chen, Y.; Zhang, H.; Guo, Y.; Lin, Y.; Li, L.; Yu, X.; et al. Spermidine Alleviates Cardiac Aging by Improving Mitochondrial Biogenesis and Function. Aging 2020, 12, 650–671. [Google Scholar] [CrossRef] [PubMed]
- Jamwal, S.; Singh, S.; Kaur, N.; Kumar, P. Protective Effect of Spermidine against Excitotoxic Neuronal Death Induced by Quinolinic Acid in Rats: Possible Neurotransmitters and Neuroinflammatory Mechanism. Neurotox. Res. 2015, 28, 171–184. [Google Scholar] [CrossRef]
- Ghosh, I.; Sankhe, R.; Mudgal, J.; Arora, D.; Nampoothiri, M. Spermidine, an Autophagy Inducer, as a Therapeutic Strategy in Neurological Disorders. Neuropeptides 2020, 83, 102083. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.; He, D.; Yao, Z.; Klionsky, D.J. The Machinery of Macroautophagy. Cell Res. 2014, 24, 24–41. [Google Scholar] [CrossRef] [Green Version]
- Mizushima, N.; Komatsu, M. Autophagy: Renovation of Cells and Tissues. Cell 2011, 147, 728–741. [Google Scholar] [CrossRef] [Green Version]
- Leidal, A.M.; Levine, B.; Debnath, J. Autophagy and the Cell Biology of Age-Related Disease. Nat. Cell Biol. 2018, 20, 1338–1348. [Google Scholar] [CrossRef]
- Minois, N.; Carmona-Gutierrez, D.; Madeo, F. Polyamines in Aging and Disease. Aging 2011, 3, 716–732. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Alsaleh, G.; Feltham, J.; Sun, Y.; Napolitano, G.; Riffelmacher, T.; Charles, P.; Frau, L.; Hublitz, P.; Yu, Z.; et al. Polyamines Control EIF5A Hypusination, TFEB Translation, and Autophagy to Reverse B Cell Senescence. Mol. Cell 2019, 76, 110–125. [Google Scholar] [CrossRef] [Green Version]
- Ni, Y.-Q.; Liu, Y.-S. New Insights into the Roles and Mechanisms of Spermidine in Aging and Age-Related Diseases. Aging Dis. 2021, 12, 1948–1963. [Google Scholar] [CrossRef] [PubMed]
- Madeo, F.; Pietrocola, F.; Eisenberg, T.; Kroemer, G. Caloric Restriction Mimetics: Towards a Molecular Definition. Nat. Rev. Drug Discov. 2014, 13, 727–740. [Google Scholar] [CrossRef] [PubMed]
- Mackeh, R.; Lorin, S.; Ratier, A.; Mejdoubi-Charef, N.; Baillet, A.; Bruneel, A.; Hamaï, A.; Codogno, P.; Poüs, C.; Perdiz, D. Reactive Oxygen Species, AMP-Activated Protein Kinase, and the Transcription Cofactor P300 Regulate α-Tubulin Acetyltransferase-1 (ATAT-1/MEC-17)-Dependent Microtubule Hyperacetylation during Cell Stress. J. Biol. Chem. 2014, 289, 11816–11828. [Google Scholar] [CrossRef] [Green Version]
- Pällmann, N.; Braig, M.; Sievert, H.; Preukschas, M.; Hermans-Borgmeyer, I.; Schweizer, M.; Nagel, C.H.; Neumann, M.; Wild, P.; Haralambieva, E.; et al. Biological Relevance and Therapeutic Potential of the Hypusine Modification System. J. Biol. Chem. 2015, 290, 18343–18360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghisalberti, C.A.; Borzì, R.M.; Cetrullo, S.; Flamigni, F.; Cairo, G. Soft TCPTP Agonism-Novel Target to Rescue Airway Epithelial Integrity by Exogenous Spermidine. Front. Pharmacol. 2016, 7, 147. [Google Scholar] [CrossRef] [Green Version]
- Yuan, H.; Wu, S.-X.; Zhou, Y.-F.; Peng, F. Spermidine Inhibits Joints Inflammation and Macrophage Activation in Mice with Collagen-Induced Arthritis. J. Inflamm. Res. 2021, 14, 2713–2721. [Google Scholar] [CrossRef]
- Yuan, X.; Tian, G.G.; Pei, X.; Hu, X.; Wu, J. Spermidine Induces Cytoprotective Autophagy of Female Germline Stem Cells in Vitro and Ameliorates Aging Caused by Oxidative Stress through Upregulated Sequestosome-1/P62 Expression. Cell Biosci. 2021, 11, 107. [Google Scholar] [CrossRef]
- Stout, R.D.; Jiang, C.; Matta, B.; Tietzel, I.; Watkins, S.K.; Suttles, J. Macrophages Sequentially Change Their Functional Phenotype in Response to Changes in Microenvironmental Influences. J. Immunol. Baltim. Md. 2005, 175, 342–349. [Google Scholar] [CrossRef]
- Jamerson, E.C.; Elhusseiny, A.M.; ElSheikh, R.H.; Eleiwa, T.K.; El Sayed, Y.M. Role of Matrix Metalloproteinase 9 in Ocular Surface Disorders. Eye Contact Lens 2020, 46 (Suppl. S2), S57–S63. [Google Scholar] [CrossRef]
- Shapouri-Moghaddam, A.; Mohammadian, S.; Vazini, H.; Taghadosi, M.; Esmaeili, S.-A.; Mardani, F.; Seifi, B.; Mohammadi, A.; Afshari, J.T.; Sahebkar, A. Macrophage Plasticity, Polarization, and Function in Health and Disease. J. Cell. Physiol. 2018, 233, 6425–6440. [Google Scholar] [CrossRef]
- Igarashi, K.; Kashiwagi, K. Characteristics of Cellular Polyamine Transport in Prokaryotes and Eukaryotes. Plant Physiol. Biochem. PPB 2010, 48, 506–512. [Google Scholar] [CrossRef] [PubMed]
- Gordon, S.; Taylor, P.R. Monocyte and Macrophage Heterogeneity. Nat. Rev. Immunol. 2005, 5, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zheng, C.; Cao, J.; Cao, G.; Shou, P.; Lin, L.; Velletri, T.; Jiang, M.; Chen, Q.; Han, Y.; et al. Spermidine Alleviates Experimental Autoimmune Encephalomyelitis through Inducing Inhibitory Macrophages. Cell Death Differ. 2016, 23, 1850–1861. [Google Scholar] [CrossRef] [PubMed]
- Lee, G.R. The Balance of Th17 versus Treg Cells in Autoimmunity. Int. J. Mol. Sci. 2018, 19, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stepp, M.A.; Menko, A.S. Immune Responses to Injury and Their Links to Eye Disease. Transl. Res. J. Lab. Clin. Med. 2021, 236, 52–71. [Google Scholar] [CrossRef]
- Wang, C.; Zhou, W.; Su, G.; Hu, J.; Yang, P. Progranulin Suppressed Autoimmune Uveitis and Autoimmune Neuroinflammation by Inhibiting Th1/Th17 Cells and Promoting Treg Cells and M2 Macrophages. Neurol. Neuroimmunol. Neuroinflamm. 2022, 9, e1133. [Google Scholar] [CrossRef]
- Cong, H.; Jiang, H.; Peng, J.; Cui, S.; Liu, L.; Wang, J.; Zhang, X. Change of Th17 Lymphocytes and Treg/Th17 in Typical and Atypical Optic Neuritis. PLoS ONE 2016, 11, e0146270. [Google Scholar] [CrossRef]
- Verstappen, G.M.; Corneth, O.B.J.; Bootsma, H.; Kroese, F.G.M. Th17 Cells in Primary Sjögren’s Syndrome: Pathogenicity and Plasticity. J. Autoimmun. 2018, 87, 16–25. [Google Scholar] [CrossRef]
- Zhao, J.; Chen, W.; Huang, X.; Peng, S.; Zhu, T.; Deng, Z.; Liang, P.; Chang, H.; Fan, B.J. Serum Th1 and Th17 Related Cytokines and Autoantibodies in Patients with Posner-Schlossman Syndrome. PLoS ONE 2017, 12, e0175519. [Google Scholar] [CrossRef]
- Zhang, Z.; Cao, K.; Liu, J.; Wei, Z.; Xu, X.; Liang, Q. Pathogens and Antibiotic Susceptibilities of Global Bacterial Keratitis: A Meta-Analysis. Antibiotics 2022, 11, 238. [Google Scholar] [CrossRef]
- Bispo, P.J.M.; Sahm, D.F.; Asbell, P.A. A Systematic Review of Multi-Decade Antibiotic Resistance Data for Ocular Bacterial Pathogens in the United States. Ophthalmol. Ther. 2022, 11, 503–520. [Google Scholar] [CrossRef] [PubMed]
- Malone, L.; Kwon, D.H. Carbapenem-Associated Multidrug-Resistant Acinetobacter Baumannii Are Sensitised by Aztreonam in Combination with Polyamines. Int. J. Antimicrob. Agents 2013, 41, 70–74. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, D.-H.; Lu, C.-D. Polyamine Effects on Antibiotic Susceptibility in Bacteria. Antimicrob. Agents Chemother. 2007, 51, 2070–2077. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, L.; Ni, Y.; Wang, Z.; Tu, W.; Ni, L.; Zhuge, F.; Zheng, A.; Hu, L.; Zhao, Y.; Zheng, L.; et al. Spermidine Improves Gut Barrier Integrity and Gut Microbiota Function in Diet-Induced Obese Mice. Gut Microbes 2020, 12, 1–19. [Google Scholar] [CrossRef]
- Petrillo, F.; Pignataro, D.; Lavano, M.A.; Santella, B.; Folliero, V.; Zannella, C.; Astarita, C.; Gagliano, C.; Franci, G.; Avitabile, T.; et al. Current Evidence on the Ocular Surface Microbiota and Related Diseases. Microorganisms 2020, 8, 1033. [Google Scholar] [CrossRef]
- Hume, E.B.; Dajcs, J.J.; Moreau, J.M.; Sloop, G.D.; Willcox, M.D.; O’Callaghan, R.J. Staphylococcus Corneal Virulence in a New Topical Model of Infection. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2904–2908. [Google Scholar]
- Moreau, J.M.; Girgis, D.O.; Hume, E.B.; Dajcs, J.J.; Austin, M.S.; O’Callaghan, R.J. Phospholipase A(2) in Rabbit Tears: A Host Defense against Staphylococcus aureus. Investig. Ophthalmol. Vis. Sci. 2001, 42, 2347–2354. [Google Scholar]
- Rabinowitz, Y.S. Keratoconus. Surv. Ophthalmol. 1998, 42, 297–319. [Google Scholar] [CrossRef]
- Kenney, M.C.; Nesburn, A.B.; Burgeson, R.E.; Butkowski, R.J.; Ljubimov, A.V. Abnormalities of the Extracellular Matrix in Keratoconus Corneas. Cornea 1997, 16, 345–351. [Google Scholar] [CrossRef]
- McKay, T.B.; Priyadarsini, S.; Rowsey, T.; Karamichos, D. Arginine Supplementation Promotes Extracellular Matrix and Metabolic Changes in Keratoconus. Cells 2021, 10, 2076. [Google Scholar] [CrossRef]
- Maekawa, S.; Hibasami, H.; Uji, Y.; Nakashima, K. Active Transport and Metabolic Characteristics of Polyamines in the Rat Lens. Biochim. Biophys. Acta 1989, 993, 199–203. [Google Scholar] [CrossRef]
- Kremzner, L.T.; Roy, D.; Spector, A. Polyamines in Normal and Cataractous Human Lenses: Evidence for Post-Translational Modification. Exp. Eye Res. 1983, 37, 649–659. [Google Scholar] [CrossRef]
- Maekawa, S.; Hibasami, H.; Tsukada, T.; Furusako, S.; Nakashima, K.; Yokoyama, M. Induction of Spermidine/Spermine N1-Acetyltransferase in Needle-Punctured Rat Lens as a Model of Traumatic Cataract. Biochim. Biophys. Acta 1986, 883, 501–505. [Google Scholar] [CrossRef]
- Mischiati, C.; Feriotto, G.; Tabolacci, C.; Domenici, F.; Melino, S.; Borromeo, I.; Forni, C.; De Martino, A.; Beninati, S. Polyamine Oxidase Is Involved in Spermidine Reduction of Transglutaminase Type 2-Catalyzed ΒH-Crystallins Polymerization in Calcium-Induced Experimental Cataract. Int. J. Mol. Sci. 2020, 21, 5427. [Google Scholar] [CrossRef]
- Duncan, G.; Jacob, T.J. Calcium and the Physiology of Cataract. In Ciba Foundation Symposium 106; Pitman Publishing: London, UK, 1984; pp. 132–152. [Google Scholar] [CrossRef]
- Duncan, G.; Wormstone, I.M. Calcium Cell Signalling and Cataract: Role of the Endoplasmic Reticulum. Eye Lond. Engl. 1999, 13 Pt 3b, 480–483. [Google Scholar] [CrossRef]
- Wildner, G.; Diedrichs-Möhring, M. Resolution of Uveitis. Semin. Immunopathol. 2019, 41, 727–736. [Google Scholar] [CrossRef]
- Wickström, K. Polyamine and Histopathological Changes after Unilateral Endotoxin-Induced Uveitis and Its Contralateral Effects. Acta Ophthalmol. (Copenh.) 1992, 70, 506–514. [Google Scholar] [CrossRef]
- Wickström, K.; Lundgren, B.; Törngren, L.; Ostberg, C. Aqueous Humor Polyamines and Alkaline Phosphatase Activity in Endotoxin-Induced Uveitis: Correlations to Diverse Leukocyte Subsets. Ophthalmic Res. 1992, 24, 175–180. [Google Scholar] [CrossRef]
- Wickström, K. Polyamines in Rabbit Aqueous Humor after Surgical Trauma to the Eye. Curr. Eye Res. 1991, 10, 463–469. [Google Scholar] [CrossRef]
- Jonas, J.B.; Aung, T.; Bourne, R.R.; Bron, A.M.; Ritch, R.; Panda-Jonas, S. Glaucoma. Lancet Lond. Engl. 2017, 390, 2183–2193. [Google Scholar] [CrossRef]
- Zukerman, R.; Harris, A.; Vercellin, A.V.; Siesky, B.; Pasquale, L.R.; Ciulla, T.A. Molecular Genetics of Glaucoma: Subtype and Ethnicity Considerations. Genes 2020, 12, 55. [Google Scholar] [CrossRef] [PubMed]
- Leruez, S.; Marill, A.; Bresson, T.; de Saint Martin, G.; Buisset, A.; Muller, J.; Tessier, L.; Gadras, C.; Verny, C.; Gohier, P.; et al. A Metabolomics Profiling of Glaucoma Points to Mitochondrial Dysfunction, Senescence, and Polyamines Deficiency. Investig. Ophthalmol. Vis. Sci. 2018, 59, 4355–4361. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leruez, S.; Bresson, T.; Chao de la Barca, J.M.; Marill, A.; de Saint Martin, G.; Buisset, A.; Muller, J.; Tessier, L.; Gadras, C.; Verny, C.; et al. A Plasma Metabolomic Signature of the Exfoliation Syndrome Involves Amino Acids, Acylcarnitines, and Polyamines. Investig. Ophthalmol. Vis. Sci. 2018, 59, 1025–1032. [Google Scholar] [CrossRef]
- Pan, C.-W.; Ke, C.; Chen, Q.; Tao, Y.-J.; Zha, X.; Zhang, Y.-P.; Zhong, H. Differential Metabolic Markers Associated with Primary Open-Angle Glaucoma and Cataract in Human Aqueous Humor. BMC Ophthalmol. 2020, 20, 183. [Google Scholar] [CrossRef] [PubMed]
- Chao de la Barca, J.M.; Simard, G.; Sarzi, E.; Chaumette, T.; Rousseau, G.; Chupin, S.; Gadras, C.; Tessier, L.; Ferré, M.; Chevrollier, A.; et al. Targeted Metabolomics Reveals Early Dominant Optic Atrophy Signature in Optic Nerves of Opa1delTTAG/+ Mice. Investig. Ophthalmol. Vis. Sci. 2017, 58, 812–820. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pang, Y.; Qin, M.; Hu, P.; Ji, K.; Xiao, R.; Sun, N.; Pan, X.; Zhang, X. Resveratrol Protects Retinal Ganglion Cells against Ischemia Induced Damage by Increasing Opa1 Expression. Int. J. Mol. Med. 2020, 46, 1707–1720. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Harada, C.; Namekata, K.; Kimura, A.; Mitamura, Y.; Yoshida, H.; Matsumoto, Y.; Harada, T. Spermidine Alleviates Severity of Murine Experimental Autoimmune Encephalomyelitis. Investig. Ophthalmol. Vis. Sci. 2011, 52, 2696–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noro, T.; Namekata, K.; Kimura, A.; Guo, X.; Azuchi, Y.; Harada, C.; Nakano, T.; Tsuneoka, H.; Harada, T. Spermidine Promotes Retinal Ganglion Cell Survival and Optic Nerve Regeneration in Adult Mice Following Optic Nerve Injury. Cell Death Dis. 2015, 6, e1720. [Google Scholar] [CrossRef] [Green Version]
- Gilad, G.M.; Gilad, V.H. Treatment with Polyamines Can Prevent Monosodium Glutamate Neurotoxicity in the Rat Retina. Life Sci. 1989, 44, 1963–1969. [Google Scholar] [CrossRef]
- Wareham, L.K.; Calkins, D.J. The Neurovascular Unit in Glaucomatous Neurodegeneration. Front. Cell Dev. Biol. 2020, 8, 452. [Google Scholar] [CrossRef]
- Rinaldi, C.; Donato, L.; Alibrandi, S.; Scimone, C.; D’Angelo, R.; Sidoti, A. Oxidative Stress and the Neurovascular Unit. Life 2021, 11, 767. [Google Scholar] [CrossRef] [PubMed]
- Harada, T.; Harada, C.; Nakamura, K.; Quah, H.-M.A.; Okumura, A.; Namekata, K.; Saeki, T.; Aihara, M.; Yoshida, H.; Mitani, A.; et al. The Potential Role of Glutamate Transporters in the Pathogenesis of Normal Tension Glaucoma. J. Clin. Investig. 2007, 117, 1763–1770. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharif, N.A.; Xu, S.X. Human Retina Contains Polyamine Sensitive [3H]-Ifenprodil Binding Sites: Implications for Neuroprotection? Br. J. Ophthalmol. 1999, 83, 236–240. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valentino, T.L.; Lukasiewicz, P.D.; Romano, C. Immunocytochemical Localization of Polyamines in the Tiger Salamander Retina. Brain Res. 1996, 713, 278–285. [Google Scholar] [CrossRef]
- Fuwa, M.; Kageyama, M.; Ohashi, K.; Sasaoka, M.; Sato, R.; Tanaka, M.; Tashiro, K. Nafamostat and Sepimostat Identified as Novel Neuroprotective Agents via NR2B N-Methyl-D-Aspartate Receptor Antagonism Using a Rat Retinal Excitotoxicity Model. Sci. Rep. 2019, 9, 20409. [Google Scholar] [CrossRef]
- Zhang, S.; Kashii, S.; Yasuyoshi, H.; Kikuchi, M.; Honda, Y.; Kaneda, K.; Sato, S.; Akaike, A. Protective Effects of Ifenprodil against Glutamate-Induced Neurotoxicity in Cultured Retinal Neurons. Graefes Arch. Clin. Exp. Ophthalmol. Albrecht Von Graefes Arch. Klin. Exp. Ophthalmol. 2000, 238, 846–852. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ohashi, K.; Kageyama, M.; Shinomiya, K.; Fujita-Koyama, Y.; Hirai, S.-I.; Katsuta, O.; Nakamura, M. Spermidine Oxidation-Mediated Degeneration of Retinal Pigment Epithelium in Rats. Oxid. Med. Cell. Longev. 2017, 2017, 4128061. [Google Scholar] [CrossRef]
- Williams, K. Modulation and Block of Ion Channels: A New Biology of Polyamines. Cell. Signal. 1997, 9, 1–13. [Google Scholar] [CrossRef]
- Block, M.L.; Zecca, L.; Hong, J.-S. Microglia-Mediated Neurotoxicity: Uncovering the Molecular Mechanisms. Nat. Rev. Neurosci. 2007, 8, 57–69. [Google Scholar] [CrossRef]
- Brown, G.C.; Neher, J.J. Inflammatory Neurodegeneration and Mechanisms of Microglial Killing of Neurons. Mol. Neurobiol. 2010, 41, 242–247. [Google Scholar] [CrossRef]
- Scimone, C.; Donato, L.; Alibrandi, S.; Vadalà, M.; Giglia, G.; Sidoti, A.; D’Angelo, R. N-Retinylidene-N-Retinylethanolamine Adduct Induces Expression of Chronic Inflammation Cytokines in Retinal Pigment Epithelium Cells. Exp. Eye Res. 2021, 209, 108641. [Google Scholar] [CrossRef] [PubMed]
- Scozzafava, A.; Supuran, C.T.; Carta, F. Polyamines and α-Carbonic Anhydrases. Molecules 2016, 21, 1726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carta, F.; Temperini, C.; Innocenti, A.; Scozzafava, A.; Kaila, K.; Supuran, C.T. Polyamines Inhibit Carbonic Anhydrases by Anchoring to the Zinc-Coordinated Water Molecule. J. Med. Chem. 2010, 53, 5511–5522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Supuran, C.T.; Altamimi, A.S.A.; Carta, F. Carbonic Anhydrase Inhibition and the Management of Glaucoma: A Literature and Patent Review 2013–2019. Expert Opin. Ther. Pat. 2019, 29, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Withrow, C.; Ashraf, S.; O’Leary, T.; Johnson, L.R.; Fitzgerald, M.E.C.; Johnson, D.A. Effect of Polyamine Depletion on Cone Photoreceptors of the Developing Rabbit Retina. Investig. Ophthalmol. Vis. Sci. 2002, 43, 3081–3090. [Google Scholar] [PubMed]
- Slotkin, T.A.; Seidler, F.J.; Trepanier, P.A.; Whitmore, W.L.; Lerea, L.; Barnes, G.A.; Weigel, S.J.; Bartolome, J. Ornithine Decarboxylase and Polyamines in Tissues of the Neonatal Rat: Effects of Alpha-Difluoromethylornithine, a Specific, Irreversible Inhibitor of Ornithine Decarboxylase. J. Pharmacol. Exp. Ther. 1982, 222, 741–745. [Google Scholar]
- Macaione, S.; Di Giorgio, R.M.; Nicotina, P.A.; Ientile, R. Retina Maturation Following Administration of Thyroxine in Developing Rats: Effects on Polyamine Metabolism and Glutamate Decarboxylase. J. Neurochem. 1984, 43, 303–315. [Google Scholar] [CrossRef] [PubMed]
- Donato, L.; Abdalla, E.M.; Scimone, C.; Alibrandi, S.; Rinaldi, C.; Nabil, K.M.; D’Angelo, R.; Sidoti, A. Impairments of Photoreceptor Outer Segments Renewal and Phototransduction Due to a Peripherin Rare Haplotype Variant: Insights from Molecular Modeling. Int. J. Mol. Sci. 2021, 22, 3484. [Google Scholar] [CrossRef]
- Blasiak, J. Senescence in the Pathogenesis of Age-Related Macular Degeneration. Cell. Mol. Life Sci. CMLS 2020, 77, 789–805. [Google Scholar] [CrossRef]
- Yanagihara, N.; Moriwaki, M.; Shiraki, K.; Miki, T.; Otani, S. The Involvement of Polyamines in the Proliferation of Cultured Retinal Pigment Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 1996, 37, 1975–1983. [Google Scholar]
- Li, J.; Qiu, C.; Wei, Y.; Yuan, W.; Liu, J.; Cui, W.; Zhou, J.; Qiu, C.; Guo, L.; Huang, L.; et al. Human Amniotic Epithelial Stem Cell-Derived Retinal Pigment Epithelium Cells Repair Retinal Degeneration. Front. Cell Dev. Biol. 2021, 9, 737242. [Google Scholar] [CrossRef] [PubMed]
- Kauppinen, A.; Paterno, J.J.; Blasiak, J.; Salminen, A.; Kaarniranta, K. Inflammation and Its Role in Age-Related Macular Degeneration. Cell. Mol. Life Sci. CMLS 2016, 73, 1765–1786. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaneko, S.; Ueda-Yamada, M.; Ando, A.; Matsumura, S.; Okuda-Ashitaka, E.; Matsumura, M.; Uyama, M.; Ito, S. Cytotoxic Effect of Spermine on Retinal Pigment Epithelial Cells. Investig. Ophthalmol. Vis. Sci. 2007, 48, 455–463. [Google Scholar] [CrossRef]
- Jiang, D.; Jiang, Y.; Long, S.; Chen, Z.; Li, Y.; Mo, G.; Bai, L.; Hao, X.; Yan, Y.; Li, L.; et al. Spermidine at Supraphysiological Doses Induces Oxidative Stress and Granulosa Cell Apoptosis in Mouse Ovaries. Theriogenology 2021, 168, 25–32. [Google Scholar] [CrossRef] [PubMed]
- Hou, W.-K.; Zhang, B.-Z.; Fu, S.-J.; Guan, G.-J.; Lai, H. Alteration of serum polyamines and its clinical significance in type 2daibetic patients. Acta Acad. Med. Shandong 2001, 39, 231–233. (In Chinese) [Google Scholar] [CrossRef]
- Seghieri, G.; Gironi, A.; Mammini, P.; Alviggi, L.; De Giorgio, L.A.; Bartolomei, G.; Ignesti, G.; Franconi, F. Erythrocyte Spermidine Levels in IDDM Patients. Diabetes Care 1992, 15, 543–545. [Google Scholar] [CrossRef] [PubMed]
- Nicoletti, R.; Venza, I.; Ceci, G.; Visalli, M.; Teti, D.; Reibaldi, A. Vitreous Polyamines Spermidine, Putrescine, and Spermine in Human Proliferative Disorders of the Retina. Br. J. Ophthalmol. 2003, 87, 1038–1042. [Google Scholar] [CrossRef] [Green Version]
- Yang, Q.-Q.; Ma, Q.-X.; Shi, J.-J.; Fu, D.-T.; Zheng, C.-W.; Zhou, J.; Chen, Y. Establishment and pathological characteristics of a new rat model of diabetic retinopathy. Chin. J. Diabetes 2021, 13, 490–497. (In Chinese) [Google Scholar] [CrossRef]
- Sen, P.; Wu, W.-C.; Chandra, P.; Vinekar, A.; Manchegowda, P.T.; Bhende, P. Retinopathy of Prematurity Treatment: Asian Perspectives. Eye Lond. Engl. 2020, 34, 632–642. [Google Scholar] [CrossRef]
- Grałek, M.; Fogel, W.A. Polyamine system in experimental retinopathy of prematurity. Klin. Ocz. 1995, 97, 182–184. [Google Scholar]
- Gralek, M.; Sasiak, K.; Wojcik, A.; Gardiner, T.A.; Fogel, W.A. Polyamine System in Developing Rat Eye and an Animal Model of Retinopathy of Prematurity. Graefes Arch. Clin. Exp. Ophthalmol. Albrecht Von Graefes Arch. Klin. Exp. Ophthalmol. 1999, 237, 781–786. [Google Scholar] [CrossRef] [PubMed]
- Igarashi, K.; Ueda, S.; Yoshida, K.; Kashiwagi, K. Polyamines in Renal Failure. Amino Acids 2006, 31, 477–483. [Google Scholar] [CrossRef] [PubMed]
- Zou, D.; Zhao, Z.; Li, L.; Min, Y.; Zhang, D.; Ji, A.; Jiang, C.; Wei, X.; Wu, X. A Comprehensive Review of Spermidine: Safety, Health Effects, Absorption and Metabolism, Food Materials Evaluation, Physical and Chemical Processing, and Bioprocessing. Compr. Rev. Food Sci. Food Saf. 2022, 21, 2820–2842. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Diseases | Spermidine Variation | Samples | References |
|---|---|---|---|
| Glaucoma | decreased | Serum, plasma, and aqueous humor (human) | [91,92,93] |
| Cataract | decreased | Lens (rabbit) | [19,82] |
| KC | decreased | Corneal fibroblasts (human) | [78] |
| Uveitis | Decreased in 48 h then increased | Aqueous humor (rabbit) | [86,87] |
| PDR | increased | Serum, erythrocyte, and vitreous (human) | [124,125,126] |
| PVR | increased | Vitreous (human) | [126] |
| ROP | Decreased (first 14 days) | Retina and lens (rat) | [129] |
| Study Type | Dose (Route) | Task | Findings | References |
|---|---|---|---|---|
| in vitro | 1–30 μM (cc) | Reverse apoptosis of RPEs treated with H2O2 | Blocked the increase of intracellular Ca2+, exhibiting anti-oxidative effect. Concentrations over 20 μM were found to be toxic. | [14] |
| in vitro | 10 μM (cc) | Assess RPEs migration | Promoted the migration of RPEs. | [13] |
| in vitro | 10 μM (cc) | Assess its role in RPEs proliferation | Being indispensable in RPEs proliferation. | [119] |
| in vitro | 10 mM (cc) | Assess cytotoxic effect of spermine and spermidine on RPE cells | 10mM spermidine caused slight RPEs shrinkage and reduced cell density; while spermine exhibited significant cytotoxicity. | [122] |
| in vitro | 100 μM (cc) | Counter the effect of NMDAr inhibitor in retinal neuronal cells | Reversed the neuro-protective effect of nafamostat, a NMDAr inhibitor, in the cell viability analysis. | [105] |
| in vitro | 100–500 μM (cc) | Mediate oxidative degeneration of RPEs | Induced RPEs death in vitro. | [106] |
| in vivo | 10 μL, 30 nmol/eye (i.v.i) | Induced RPEs and photoreceptor cell degeneration in rats. | ||
| in vivo | 10 μL, 20 nmol/eye (i.v.i) | Induced reversible RPEs and photoreceptor cell degeneration in rats. | ||
| in vivo | 10 μL, 10 nmol/eye (i.v.i) | No significant harmful effects were witnessed. | ||
| in vivo | 5 µL, 50 nmol/eye (i.v.i) | Counter the effect of NMDAr inhibitor in retina | Reversed the neuro-protective effect of nafamostat, a NMDAr inhibitor, after the application of neurotoxic NMDA. | [104] |
| in vivo | 10 μL, 20 nmol/eye (i.v.i) | Establishment of PDR murine model | Contributed to the establishment of PDR murine model. | [127] |
| in vivo | 10 μL, 20 pmol/eye (i.v.i) | Assess optic nerve regeneration | Promoted optic nerve regeneration after injury. | [20] |
| 10 μL, 40 pmol/eye (i.v.i) | No significant effects were observed. | |||
| in vitro | 20 μM (cc) | Assess the dose and the effect of overcoming inhibition by MAG and myelin | Could effectively reverse the inhibition of MAG and myelin. | |
| 100 μM (cc) | Diminished effects. | |||
| 500 μM (cc) | No effects. | |||
| in vivo | 10 mg/kg (s.c.) | Prevent monosodium glutamate neurotoxicity in the rat retina | Prevented neurodegeneration and neurotoxicity of monosodium glutamate in retina. | [98] |
| 1 mg/kg (s.c.) | No effects. | |||
| in vitro | 20 µL, 0.2/0.5% (Gutt) | Ameliorate dry eye syndrome induced by PM2.5 | Alleviated the symptom of dry eye murine model. Exhibited anti-inflammatory effects on ocular surface. | [17] |
| in vivo | 30 mM in drinking water (p.o.) | Assess its protective role in optic neuritis and retinal neurons. | Alleviated optic neuritis and improved visual function. | [96] |
| in vitro | 4 mM (cc) | Protected RGCs from H2O2 induced oxidative stress. | ||
| in vivo | 30 mM in drinking water (p.o.) | Investigate its protective role after optic nerve injury | Prevented RGCs death, suppressed retinal degeneration, and enhanced optic nerve regeneration. | [97] |
| in vivo | 30 mM in drinking water (p.o.) | Investigate its protective role in normal tension glaucoma murine model | Suppressed retinal degeneration, ameliorated visual impairment, and reduced oxidative stress level in retina. | [16] |
| in vitro | 100 mM (tissue incubation) | Assess its role in delaying lens opacification. | Ameliorate lens opacification induced by Ca2+. | [19,82] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Han, W.; Li, H.; Chen, B. Research Progress and Potential Applications of Spermidine in Ocular Diseases. Pharmaceutics 2022, 14, 1500. https://doi.org/10.3390/pharmaceutics14071500
Han W, Li H, Chen B. Research Progress and Potential Applications of Spermidine in Ocular Diseases. Pharmaceutics. 2022; 14(7):1500. https://doi.org/10.3390/pharmaceutics14071500
Chicago/Turabian StyleHan, Wentao, Haoyu Li, and Baihua Chen. 2022. "Research Progress and Potential Applications of Spermidine in Ocular Diseases" Pharmaceutics 14, no. 7: 1500. https://doi.org/10.3390/pharmaceutics14071500