
Effects of a Serine Protease Inhibitor N-p-Tosyl-L-phenylalanine Chloromethyl Ketone (TPCK) on Leishmania amazonensis and Leishmania infantum

 ,
,  ,
,
Abstract
:1. Introduction
2. Results
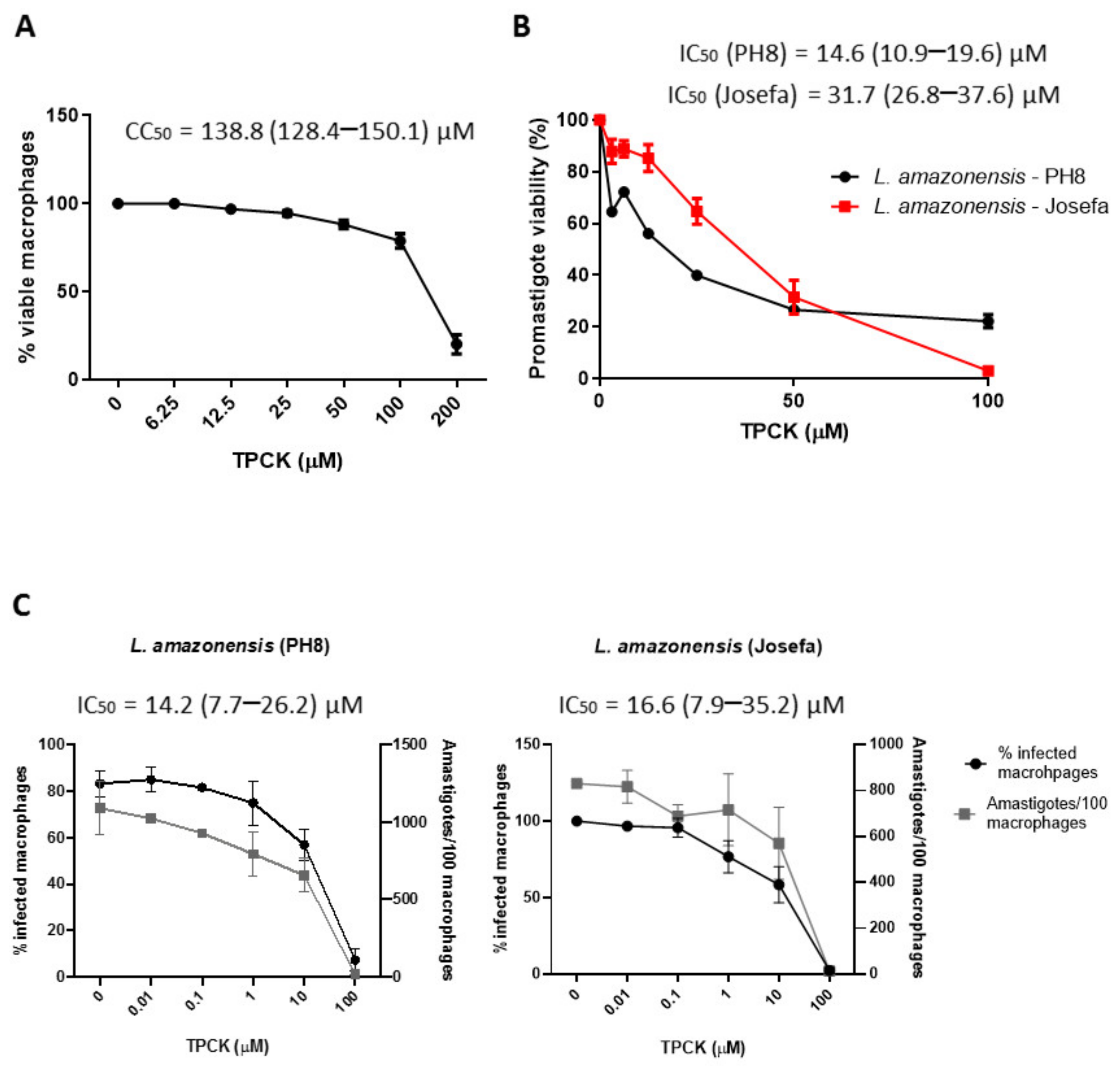
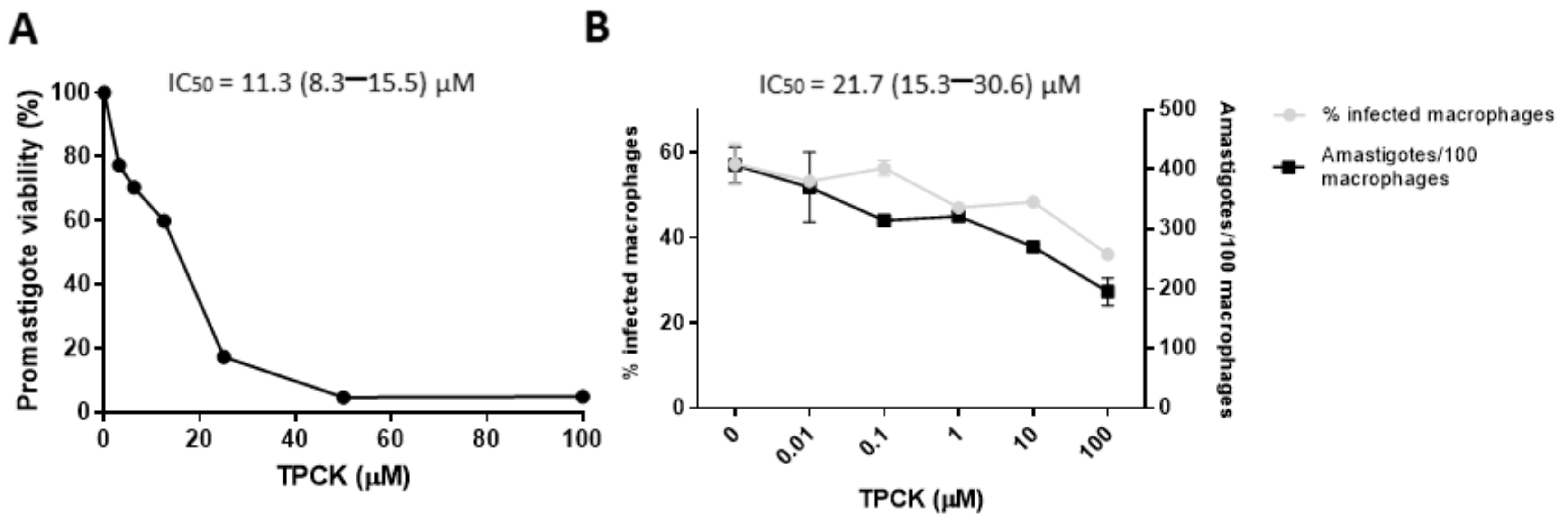
2.1. Effect of TPCK on Peritoneal Macrophages and the Leishmania Parasites, L. amazonensis and L. infantum
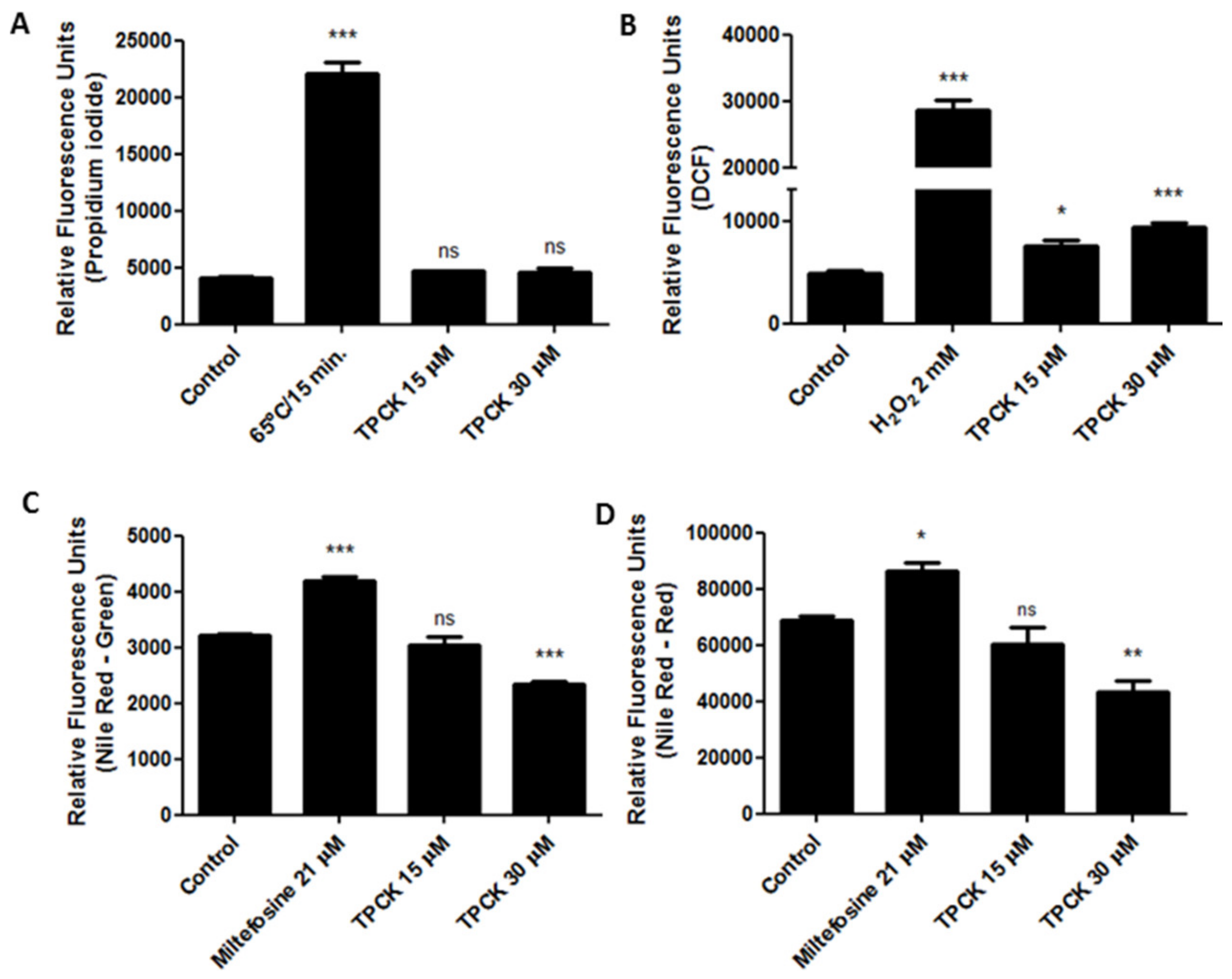
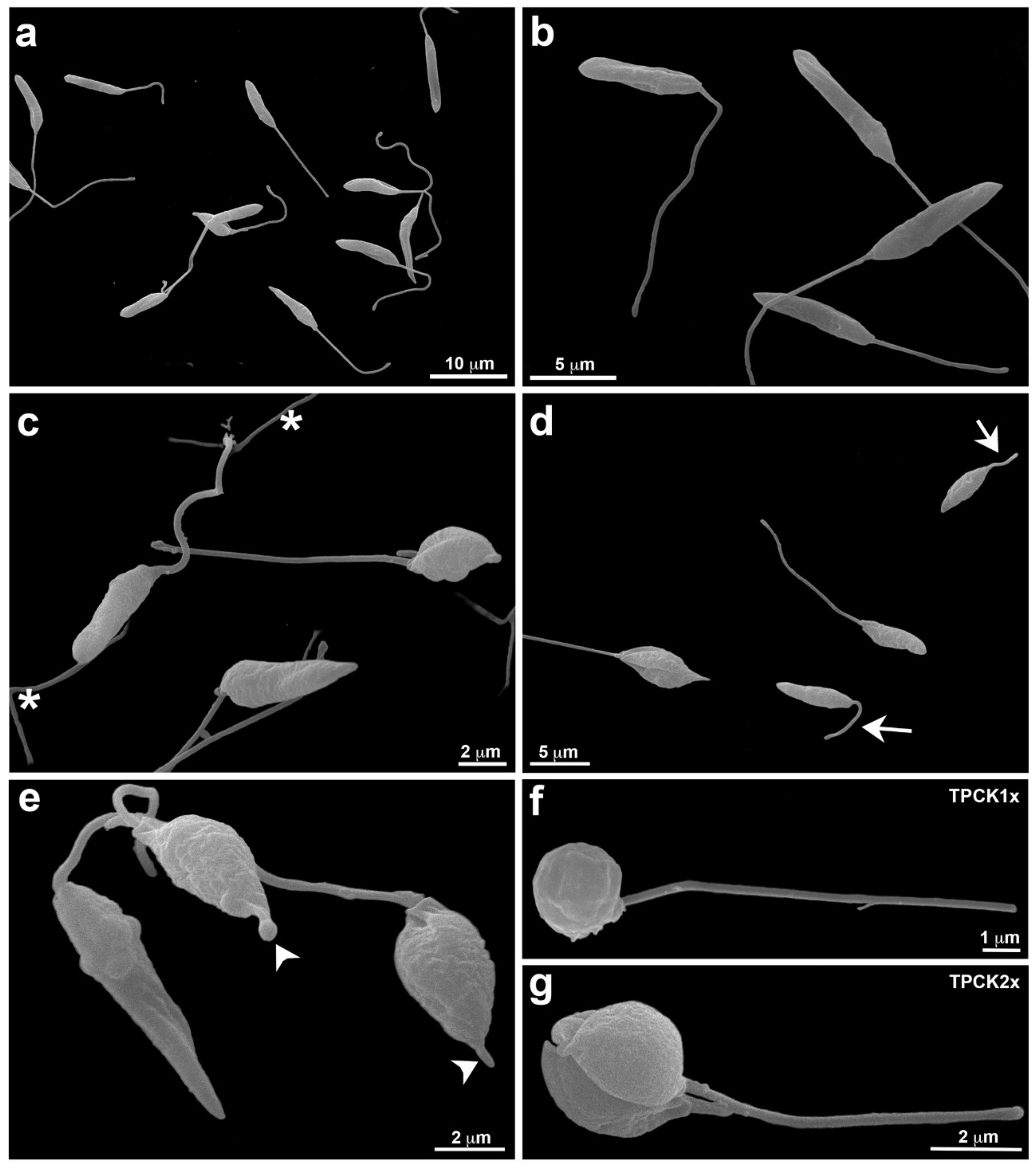
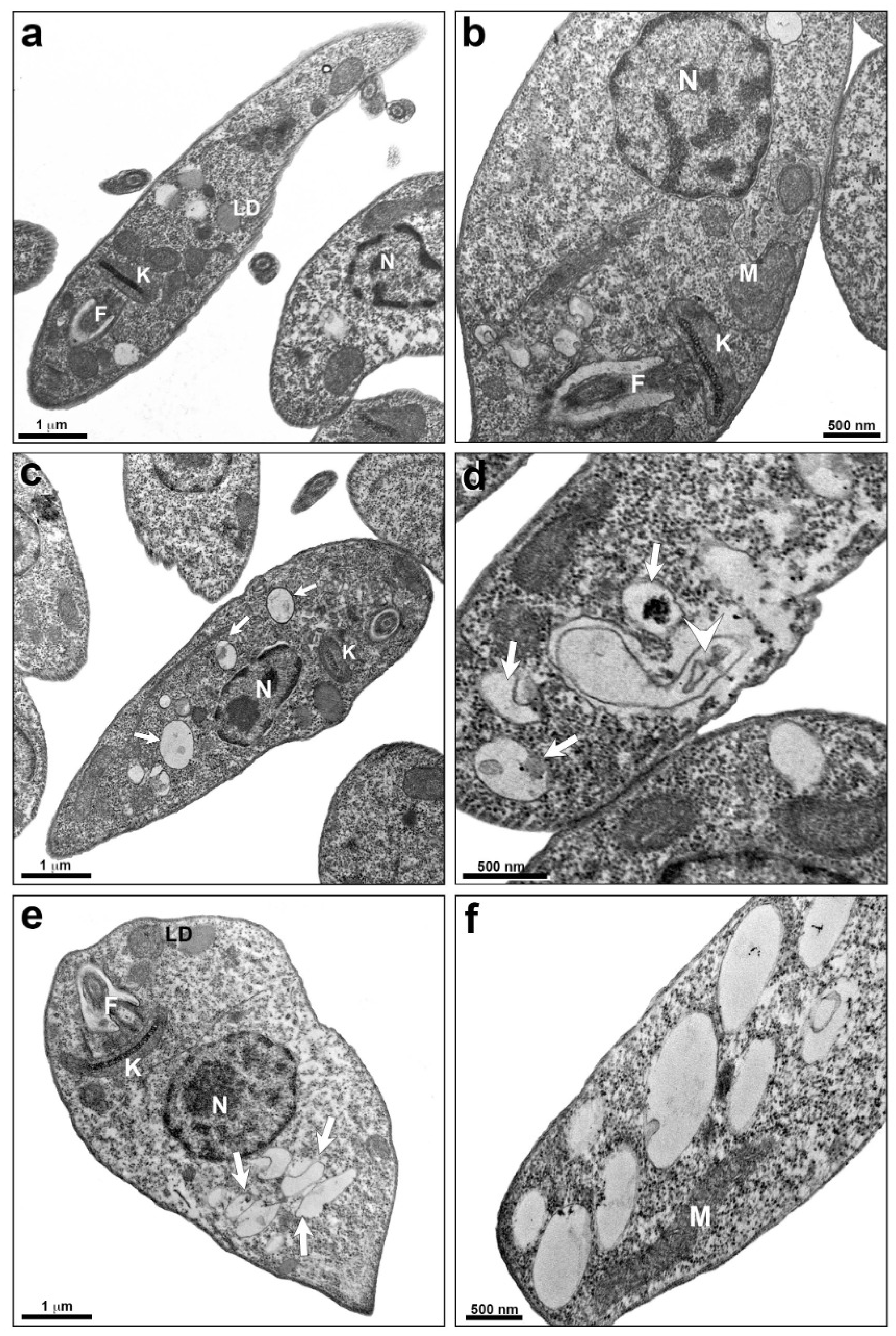
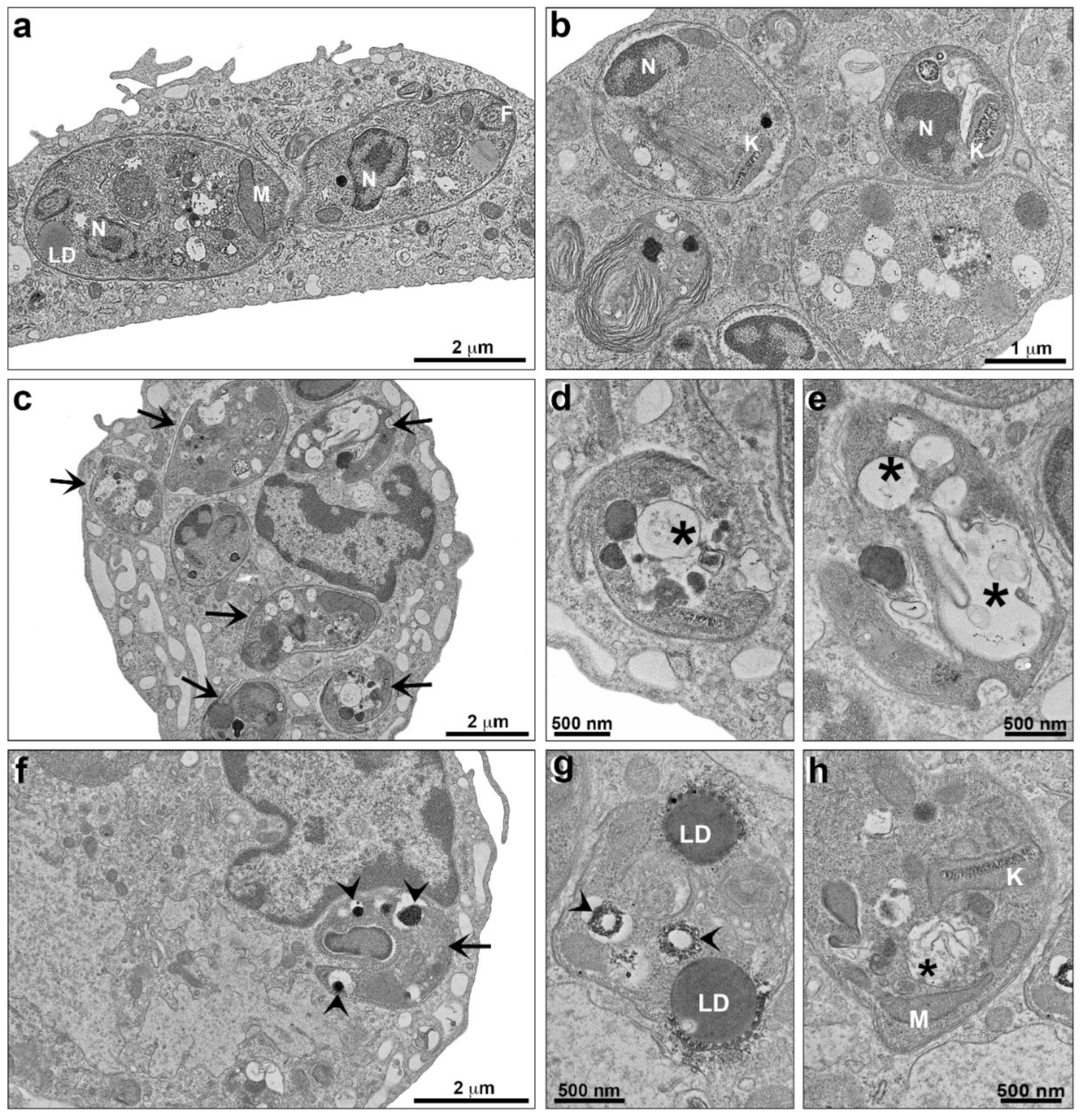
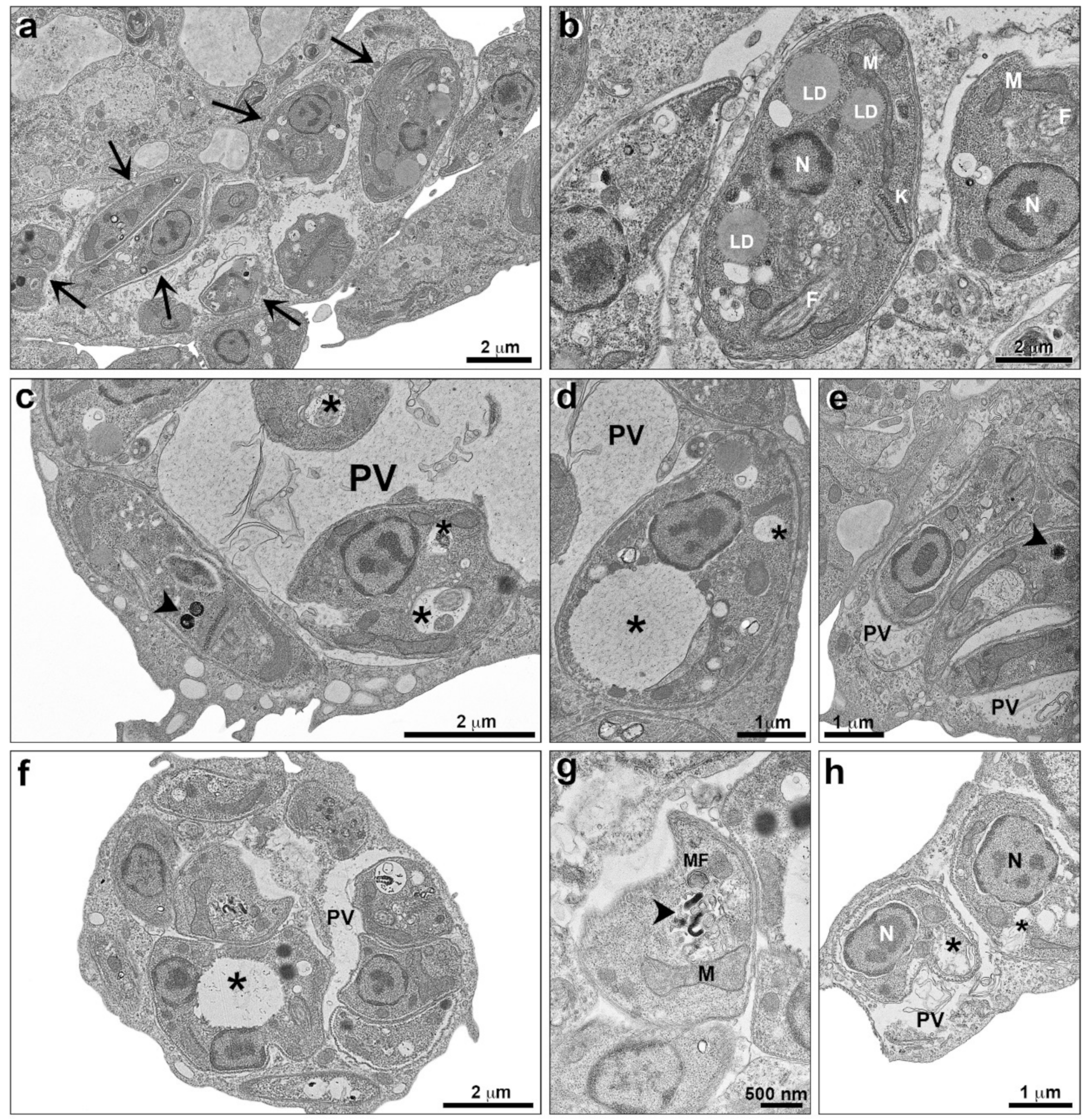
2.2. Alterations Induced by TPCK in L. amazonensis and L. infantum
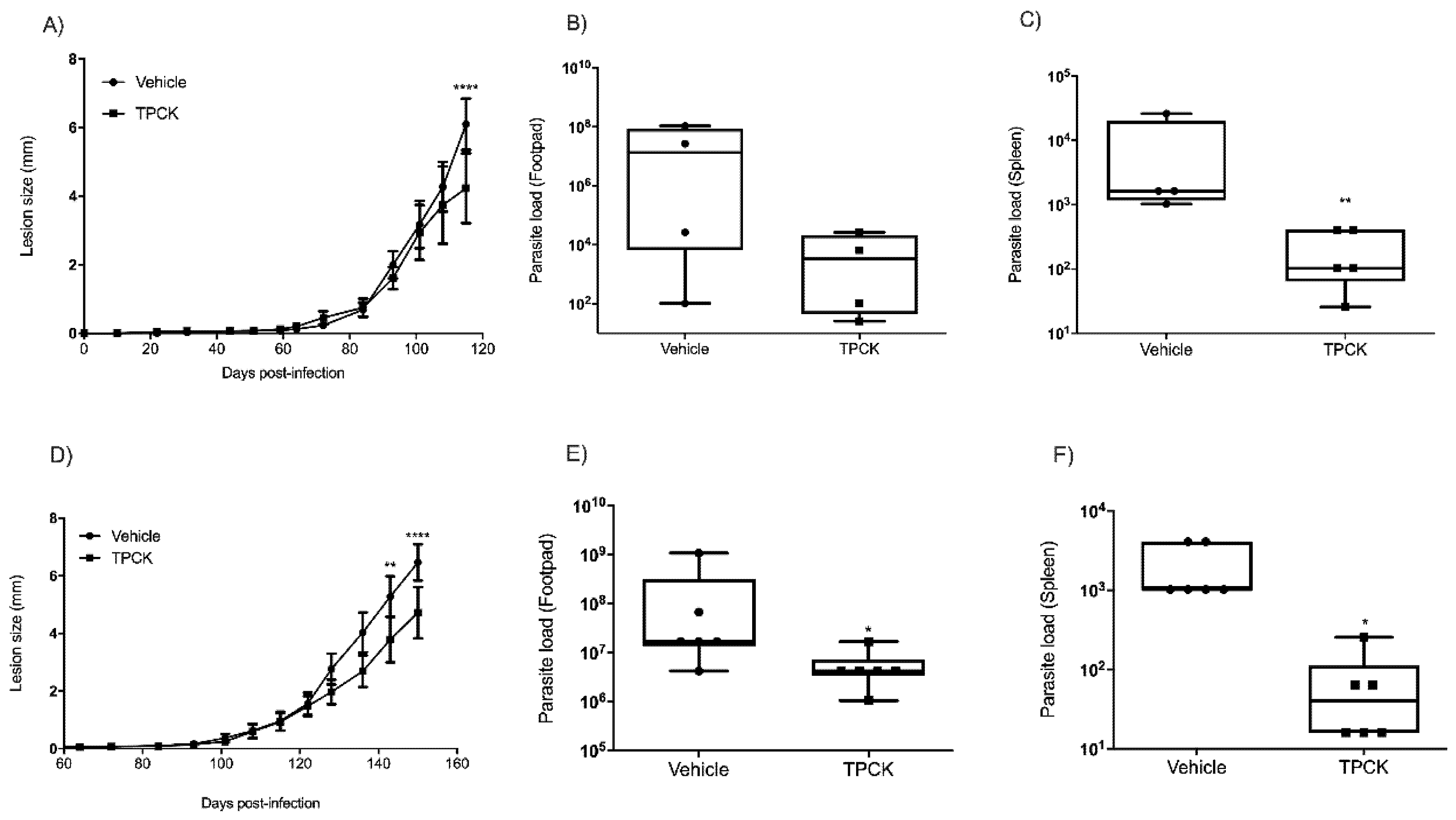
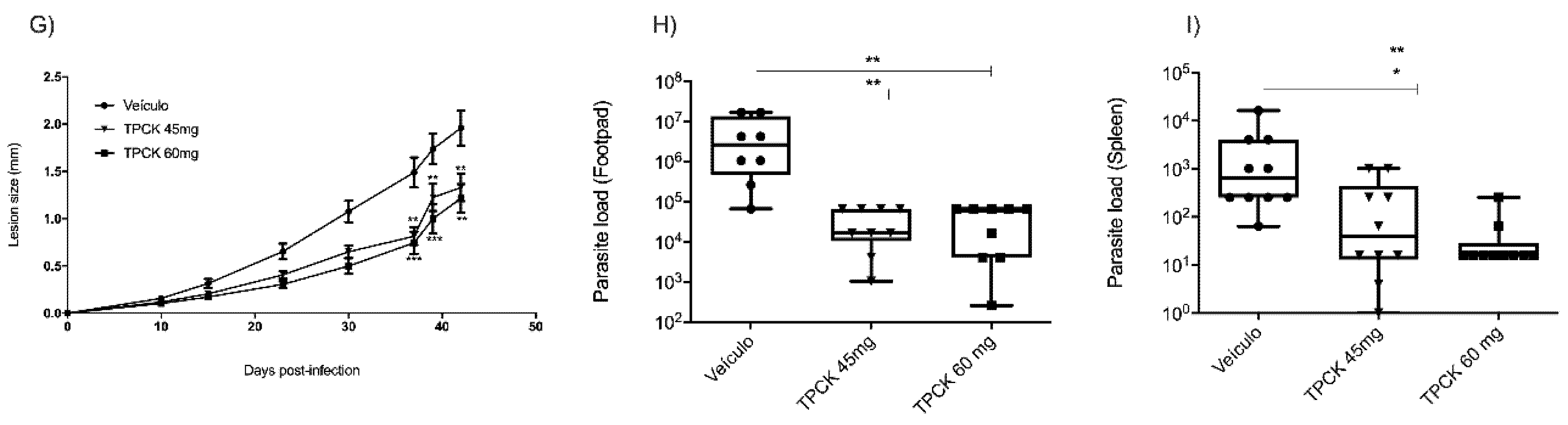
2.3. In Vivo Treatment with TPCK
3. Discussion
4. Material and Methods
4.1. Parasites
4.2. Mice
4.3. Chemicals
4.4. Cytotoxicity Assay
4.5. Anti-Promastigote Activity
4.6. Anti-Amastigote Activity
4.7. Mitochondrial Membrane Potential (ΔΨ𝑚) Determination
4.8. Mitochondrial Superoxide Production Determination
4.9. Plasma Membrane Permeability Assessments
4.10. Intracellular Reactive Oxygen Species (ROS) Level Determinations
4.11. Lipid Accumulation Assessments
4.12. Scanning Electron Microscopy (SEM)
4.13. Transmission Electron Microscopy (TEM)
4.14. In Vivo Assay and Treatment
4.15. Limiting Dilution Assay (LDA)
4.16. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Serafim, T.D.; Coutinho-Abreu, I.V.; Oliveira, F.; Meneses, C.; Kamhawi, S.; Valenzuela, J.G. Sequential blood meals promote Leishmania replication and reverse metacyclogenesis augmenting vector infectivity. Nat. Microbiol. 2018, 3, 548–555. [Google Scholar] [CrossRef]
- WHO. Leishmaniasis. World Health Organization. 2020. Available online: https://www.who.int/news-room/fact-sheets/detail/leishmaniasis (accessed on 31 March 2022).
- PAHO. Pan American Health Organization—LEISHMANIASES Epidemiological Report of the Americas. In Report No. 8 (Pan American Health Organization/World Health Organization, 2019); PAHO: Washington, DC, USA, 2019. [Google Scholar]
- Valdivia, H.O.; Almeida, L.V.; Roatt, B.M.; Reis-Cunha, J.L.; Pereira, A.A.; Gontijo, C.; Fujiwara, R.T.; Reis, A.B.; Sanders, M.J.; Cotton, J.A.; et al. Comparative genomics of canine-isolated Leishmania (Leishmania) amazonensis from an endemic focus of visceral leishmaniasis in Governador Valadares, southeastern Brazil. Sci. Rep. 2017, 7, 40804. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barral, A.; Pedral-Sampaio, D.; Grimaldi Junior, G.; Momen, H.; McMahon-Pratt, D.; Ribeiro de Jesus, A.; Almeida, R.; Badaro, R.; Barral-Netto, M.; Carvalho, E.M.; et al. Leishmaniasis in Bahia, Brazil: Evidence that Leishmania amazonensis produces a wide spectrum of clinical disease. Am. J. Trop. Med. Hyg. 1991, 44, 536–546. [Google Scholar] [CrossRef] [PubMed]
- De Oliveira, J.P.; Fernandes, F.; Cruz, A.K.; Trombela, V.; Monteiro, E.; Camargo, A.A.; Barral, A.; de Oliveira, C.I. Genetic diversity of Leishmania amazonensis strains isolated in northeastern Brazil as revealed by DNA sequencing, PCR-based analyses and molecular karyotyping. Kinetoplastid Biol. Dis. 2007, 6, 5. [Google Scholar] [CrossRef]
- Serafim, T.D.; Iniguez, E.; Oliveira, F. Leishmania infantum. Trends Parasitol. 2020, 36, 80–81. [Google Scholar] [CrossRef]
- Morizot, G.; Jouffroy, R.; Faye, A.; Chabert, P.; Belhouari, K.; Calin, R.; Charlier, C.; Miailhes, P.; Siriez, J.Y.; Mouri, O.; et al. Antimony to Cure Visceral Leishmaniasis Unresponsive to Liposomal Amphotericin B. PLoS Negl. Trop. Dis. 2016, 10, e0004304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kato, K.C.; Morais-Teixeira, E.; Reis, P.G.; Silva-Barcellos, N.M.; Salaun, P.; Campos, P.P.; Dias Correa-Junior, J.; Rabello, A.; Demicheli, C.; Frezard, F. Hepatotoxicity of pentavalent antimonial drug: Possible role of residual Sb(III) and protective effect of ascorbic acid. Antimicrob. Agents Chemother. 2014, 58, 481–488. [Google Scholar] [CrossRef] [Green Version]
- McKerrow, J.H.; Rosenthal, P.J.; Swenerton, R.; Doyle, P. Development of protease inhibitors for protozoan infections. Curr. Opin. Infect. Dis. 2008, 21, 668–672. [Google Scholar] [CrossRef] [PubMed]
- Gandra, R.M.; Silva, L.N.; Souto, X.M.; Sangenito, L.S.; Cruz, L.P.S.; Braga-Silva, L.A.; Goncalves, D.S.; Seabra, S.H.; Branquinha, M.H.; Santos, A.L.S. The serine peptidase inhibitor TPCK induces several morphophysiological changes in the opportunistic fungal pathogen Candida parapsilosis sensu stricto. Med. Mycol. 2019, 57, 1024–1037. [Google Scholar] [CrossRef]
- Conseil, V.; Soete, M.; Dubremetz, J.F. Serine protease inhibitors block invasion of host cells by Toxoplasma gondii. Antimicrob. Agents Chemother. 1999, 43, 1358–1361. [Google Scholar] [CrossRef] [Green Version]
- Silva-Lopez, R.E.; Morgado-Diaz, J.A.; Chavez, M.A.; Giovanni-De-Simone, S. Effects of serine protease inhibitors on viability and morphology of Leishmania (Leishmania) amazonensis promastigotes. Parasitol. Res. 2007, 101, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Guo, R.X.; Mo, L.Q.; Liao, X.X.; Li, W.; Zhi, J.L.; Sun, S.N.; Wang, Y.L.; Cui, Y.; Liu, W.; et al. Nuclear factor-kappaB mediates cytoprotection of hydrogen peroxide preconditioning against apoptosis induced by oxidative stress in PC12 cells. Clin. Exp. Pharmacol. Physiol. 2009, 36, 304–311. [Google Scholar] [CrossRef] [PubMed]
- Xue, J.H.; Yuan, Z.; Wu, Y.; Liu, Y.; Zhao, Y.; Zhang, W.P.; Tian, Y.L.; Liu, W.M.; Liu, Y.; Kishimoto, C. High glucose promotes intracellular lipid accumulation in vascular smooth muscle cells by impairing cholesterol influx and efflux balance. Cardiovasc. Res. 2010, 86, 141–150. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dosch, S.F.; Mahajan, S.D.; Collins, A.R. SARS coronavirus spike protein-induced innate immune response occurs via activation of the NF-kappaB pathway in human monocyte macrophages in vitro. Virus Res. 2009, 142, 19–27. [Google Scholar] [CrossRef]
- Jin, Y.J.; Zhang, X.; Cai, C.Y.; Burakoff, S.J. Alkylating HIV-1 Nef—A potential way of HIV intervention. AIDS Res. Ther. 2010, 7, 26. [Google Scholar] [CrossRef] [Green Version]
- Chakraborti, T.; Das, P.; Choudhury, R.; De, T. Effect of different serine protease inhibitors in validating the 115 kDa Leishmania donovani secretory serine protease as chemotherapeutic target. Indian J. Biochem. Biophys. 2015, 52, 14–22. [Google Scholar]
- Rokhlin, O.W.; Guseva, N.V.; Taghiyev, A.F.; Glover, R.A.; Cohen, M.B. Multiple effects of N-alpha-tosyl-L-phenylalanyl chloromethyl ketone (TPCK) on apoptotic pathways in human prostatic carcinoma cell lines. Cancer Biol. Ther. 2004, 3, 761–768. [Google Scholar] [CrossRef] [Green Version]
- Silva-Lopez, R.E. Proteases de Leishmania: Novos alvos para o desenvolvimento racional de fármacos—Leishmania proteases: New targets for rational drug development. Química Nova 2010, 33, 1541–1548. [Google Scholar] [CrossRef] [Green Version]
- Lasse, C.; Azevedo, C.S.; de Araujo, C.N.; Motta, F.N.; Andrade, M.A.; Rocha, A.P.; Sampaio, I.; Charneau, S.; Geze, M.; Grellier, P.; et al. Prolyl Oligopeptidase From Leishmania infantum: Biochemical Characterization and Involvement in Macrophage Infection. Front. Microbiol. 2020, 11, 1060. [Google Scholar] [CrossRef]
- Da Silva-Lopez, R.E.; Giovanni-De-Simone, S. Leishmania (Leishmania) amazonensis: Purification and characterization of a promastigote serine protease. Exp. Parasitol. 2004, 107, 173–182. [Google Scholar] [CrossRef]
- Morgado-Diaz, J.A.; Silva-Lopez, R.E.; Alves, C.R.; Soares, M.J.; Corte-Real, S.; De Simone, S.G. Subcellular localization of an intracellular serine protease of 68 kDa in Leishmania (Leishmania) amazonensis promastigotes. Mem. Inst. Oswaldo Cruz 2005, 100, 377–383. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guedes, H.L.; Rezende, J.M.; Fonseca, M.A.; Salles, C.M.; Rossi-Bergmann, B.; De-Simone, S.G. Identification of serine proteases from Leishmania braziliensis. Z. Nat. C J. Biosci. 2007, 62, 373–381. [Google Scholar] [CrossRef] [PubMed]
- Fidalgo, L.M.; Gille, L. Mitochondria and trypanosomatids: Targets and drugs. Pharm. Res. 2011, 28, 2758–2770. [Google Scholar] [CrossRef] [PubMed]
- Horman, S.; Del Bino, G.; Fokan, D.; Mosselmans, R.; Galand, P. Effect of the serine protease inhibitor N-tosyl-l-phenylalanine-chloromethyl ketone (TPCK) on MCF-7 mammary tumour cells growth and differentiation. Cell Biol. Int. 2000, 24, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Zhu, H.; Dinsdale, D.; Alnemri, E.S.; Cohen, G.M. Apoptosis in human monocytic THP.1 cells involves several distinct targets of N-tosyl-L-phenylalanyl chloromethyl ketone (TPCK). Cell Death Differ. 1997, 4, 590–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frydrych, I.; Mlejnek, P. Serine protease inhibitors N-alpha-tosyl-L-lysinyl-chloromethylketone (TLCK) and N-tosyl-L-phenylalaninyl-chloromethylketone (TPCK) are potent inhibitors of activated caspase proteases. J. Cell Biochem. 2008, 103, 1646–1656. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Tolle, D.P.; Barrett, A.J. Evolutionary families of peptidase inhibitors. Biochem. J. 2004, 378, 705–716. [Google Scholar] [CrossRef]
- Trivedi, J.; Ghosh, P.; Mitra, D. N-p-Tosyl-L-phenylalanine chloromethyl ketone (TPCK) inhibits HIV-1 by suppressing the activity of viral protease. Biochem. Biophys. Res. Commun. 2020, 527, 167–172. [Google Scholar] [CrossRef]
- Cuervo, P.; Mesquita-Rodrigues, C.; d’Avila Levy, C.M.; Britto, C.; Pires, F.A.; Gredilha, R.; Alves, C.R.; Jesus, J.B. Serine protease activities in Oxysarcodexia thornax (Walker) (Diptera: Sarcophagidae) first instar larva. Mem. Inst. Oswaldo Cruz 2008, 103, 504–506. [Google Scholar] [CrossRef] [Green Version]
- Alam, A. Serine Proteases of Malaria Parasite Plasmodium falciparum: Potential as Antimalarial Drug Targets. Interdiscip. Perspect. Infect. Dis. 2014, 2014, 453186. [Google Scholar] [CrossRef]
- Verma, S.; Das, S.; Mandal, A.; Ansari, M.Y.; Kumari, S.; Mansuri, R.; Kumar, A.; Singh, R.; Saini, S.; Abhishek, K.; et al. Role of inhibitors of serine peptidases in protecting Leishmania donovani against the hydrolytic peptidases of sand fly midgut. Parasit. Vectors 2017, 10, 303. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Da Silva Lopez, R.E.; De Simone, S.G. A serine protease from a detergent-soluble extract of Leishmania (Leishmania) amazonensis. Z. Nat. C J. Biosci. 2004, 59, 590–598. [Google Scholar] [CrossRef] [PubMed]
- Machado, P.A.; Gomes, P.S.; Midlej, V.; Coimbra, E.S.; de Matos Guedes, H.L. PF-429242, a Subtilisin Inhibitor, Is Effective in vitro Against Leishmania infantum. Front. Microbiol. 2021, 12, 583834. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Farhoudi, R. Leishmaniasis in humans: Drug or vaccine therapy? Drug Des. Devel. Ther. 2018, 12, 25–40. [Google Scholar] [CrossRef] [Green Version]
- Shadyro, O.; Lisovskaya, A. ROS-induced lipid transformations without oxygen participation. Chem. Phys. Lipids 2019, 221, 176–183. [Google Scholar] [CrossRef]
- De Paula, J.C.; Bakoshi, A.B.K.; Lazarin-Bidoia, D.; Ud Din, Z.; Rodrigues-Filho, E.; Ueda-Nakamura, T.; Nakamura, C.V. Antiproliferative activity of the dibenzylideneacetone derivate (E)-3-ethyl-4-(4-nitrophenyl)but3-en-2-one in Trypanosoma cruzi. Acta Trop. 2020, 211, 105653. [Google Scholar] [CrossRef]
- Mallick, S.; Dey, S.; Mandal, S.; Dutta, A.; Mukherjee, D.; Biswas, G.; Chatterjee, S.; Mallick, S.; Lai, T.K.; Acharya, K.; et al. A novel triterpene from Astraeus hygrometricus induces reactive oxygen species leading to death in Leishmania donovani. Future Microbiol. 2015, 10, 763–789. [Google Scholar] [CrossRef]
- Pramanik, P.K.; Chakraborti, S.; Bagchi, A.; Chakraborti, T. Bioassay-based Corchorus capsularis L. leaf-derived beta-sitosterol exerts antileishmanial effects against Leishmania donovani by targeting trypanothione reductase. Sci. Rep. 2020, 10, 20440. [Google Scholar] [CrossRef]
- Nouri, K.; Feng, Y.; Schimmer, A.D. Mitochondrial ClpP serine protease-biological function and emerging target for cancer therapy. Cell Death Dis. 2020, 11, 841. [Google Scholar] [CrossRef]
- Bulteau, A.L.; Bayot, A. Mitochondrial proteases and cancer. Biochim. Biophys. Acta 2011, 1807, 595–601. [Google Scholar] [CrossRef] [Green Version]
- Wu, L.; Li, X.; Li, Z.; Cheng, Y.; Wu, F.; Lv, C.; Zhang, W.; Tang, W. HtrA serine proteases in cancers: A target of interest for cancer therapy. Biomed. Pharmacother. 2021, 139, 111603. [Google Scholar] [CrossRef] [PubMed]
- Swenerton, R.K.; Knudsen, G.M.; Sajid, M.; Kelly, B.L.; McKerrow, J.H. Leishmania subtilisin is a maturase for the trypanothione reductase system and contributes to disease pathology. J. Biol. Chem. 2010, 285, 31120–31129. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- LeBlanc, M.H.; Feng, Y.; Fratkin, J.D. N-tosyl-L-phenylalanyl-chloromethylketone reduces hypoxic-ischemic brain injury in rat pups. Eur. J. Pharmacol. 2000, 390, 249–256. [Google Scholar] [CrossRef]
- Hara, A.; Niwa, M.; Nakashima, M.; Iwai, T.; Uematsu, T.; Yoshimi, N.; Mori, H. Protective effect of apoptosis-inhibitory agent, N-tosyl-L-phenylalanyl chloromethyl ketone against ischemia-induced hippocampal neuronal damage. J. Cereb. Blood Flow Metab. 1998, 18, 819–823. [Google Scholar] [CrossRef] [Green Version]
- Choi, J.; Ha, K.H.; Byun, M.S.; Min, S.Y.; Park, M.J.; Park, H.S.; Oh, H.J.; Ju, J.H.; Kim, H.Y.; Jue, D.M. Treatment with N-tosyl-l-phenylalanine chloromethyl ketone after the onset of collagen-induced arthritis reduces joint erosion and NF-kappaB activation. Eur. J. Pharmacol. 2008, 595, 108–113. [Google Scholar] [CrossRef]
- Machado, P.A.; Carneiro, M.P.D.; Sousa-Batista, A.J.; Lopes, F.J.P.; Lima, A.; Chaves, S.P.; Sodero, A.C.R.; de Matos Guedes, H.L. Leishmanicidal therapy targeted to parasite proteases. Life Sci. 2019, 219, 163–181. [Google Scholar] [CrossRef]
- Von Stebut, E. Leishmaniasis. J. Dtsch. Dermatol. Ges. 2015, 13, 191–200, quiz 201. [Google Scholar] [CrossRef]
- Layoun, A.; Samba, M.; Santos, M.M. Isolation of murine peritoneal macrophages to carry out gene expression analysis upon Toll-like receptors stimulation. J. Vis. Exp. 2015, 98, e52749. [Google Scholar] [CrossRef] [Green Version]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| IC50 a/CC50 b (µM) | ||||
|---|---|---|---|---|
| Promastigotes | Intracellular Amastigotes | |||
| L. amazonensis (IC50) | PH8 | Josefa | PH8 | Josefa |
| 14.6 (10.9–19.6) | 31.7 (26.8–37.6) | 14.2 (7.7–26.2) | 16.6 (7.9–35.2) | |
| L. infantum (IC50) | 11.3 (8.3–15.5) | 21.7 (15.3–30.6) | ||
| Peritoneal macrophages (CC50) | 138.8 (128.4–150.1) | |||
| Selectivity Index (SI) c | ||||
| L. amazonensis | PH8 | Josefa | PH8 | Josefa |
| 9.5 | 4.4 | 9.8 | 8.3 | |
| L. infantum | 12.2 | 6.4 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Machado, P.d.A.; Gomes, P.S.; Carneiro, M.P.D.; Midlej, V.; Coimbra, E.S.; de Matos Guedes, H.L. Effects of a Serine Protease Inhibitor N-p-Tosyl-L-phenylalanine Chloromethyl Ketone (TPCK) on Leishmania amazonensis and Leishmania infantum. Pharmaceutics 2022, 14, 1373. https://doi.org/10.3390/pharmaceutics14071373
Machado PdA, Gomes PS, Carneiro MPD, Midlej V, Coimbra ES, de Matos Guedes HL. Effects of a Serine Protease Inhibitor N-p-Tosyl-L-phenylalanine Chloromethyl Ketone (TPCK) on Leishmania amazonensis and Leishmania infantum. Pharmaceutics. 2022; 14(7):1373. https://doi.org/10.3390/pharmaceutics14071373
Chicago/Turabian StyleMachado, Patrícia de A., Pollyanna S. Gomes, Monique P. D. Carneiro, Victor Midlej, Elaine S. Coimbra, and Herbert L. de Matos Guedes. 2022. "Effects of a Serine Protease Inhibitor N-p-Tosyl-L-phenylalanine Chloromethyl Ketone (TPCK) on Leishmania amazonensis and Leishmania infantum" Pharmaceutics 14, no. 7: 1373. https://doi.org/10.3390/pharmaceutics14071373