
Freeze-Drying of a Capsid Virus-like Particle-Based Platform Allows Stable Storage of Vaccines at Ambient Temperature

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Expression and Purification of Catcher-cVLP and Tag-cVLP
2.2. Purification of HAstem-cVLP Vaccine
2.3. Purification of RBD-cVLP Vaccine
2.4. Freeze-Drying and Reconstitution
2.5. Stability Assessment
2.6. Dynamic light Scattering
2.7. Transmission Electron Microscopy
2.8. ACE2 Receptor Binding Assay
2.9. Mouse Immunization Studies
2.10. Serum Immunoglobulin Levels
2.11. Statistical Analysis
3. Results
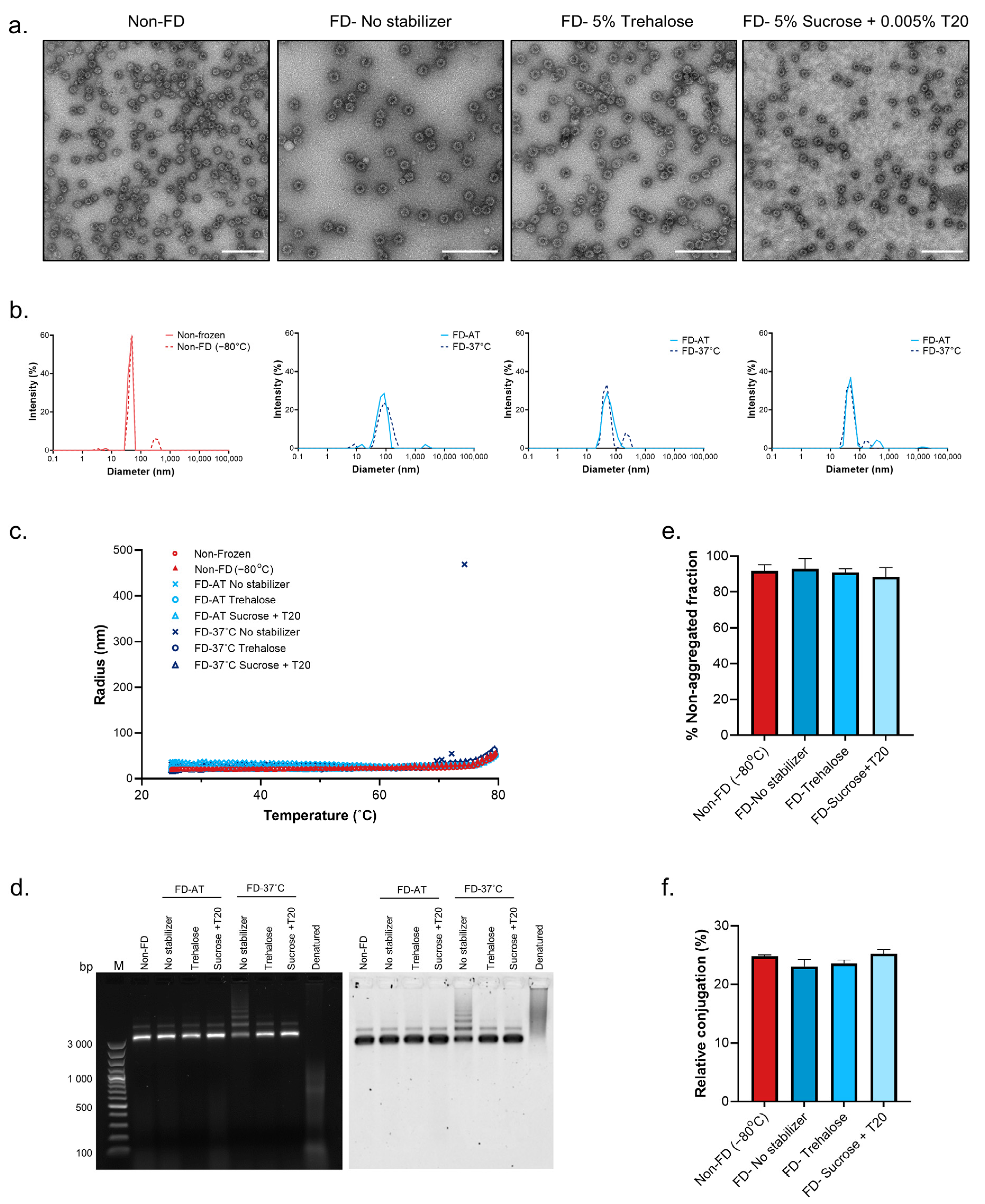
3.1. Effect of Excipient Addition on the Stability of Unconjugated Tag/Catcher-cVLPs following Freeze-Drying
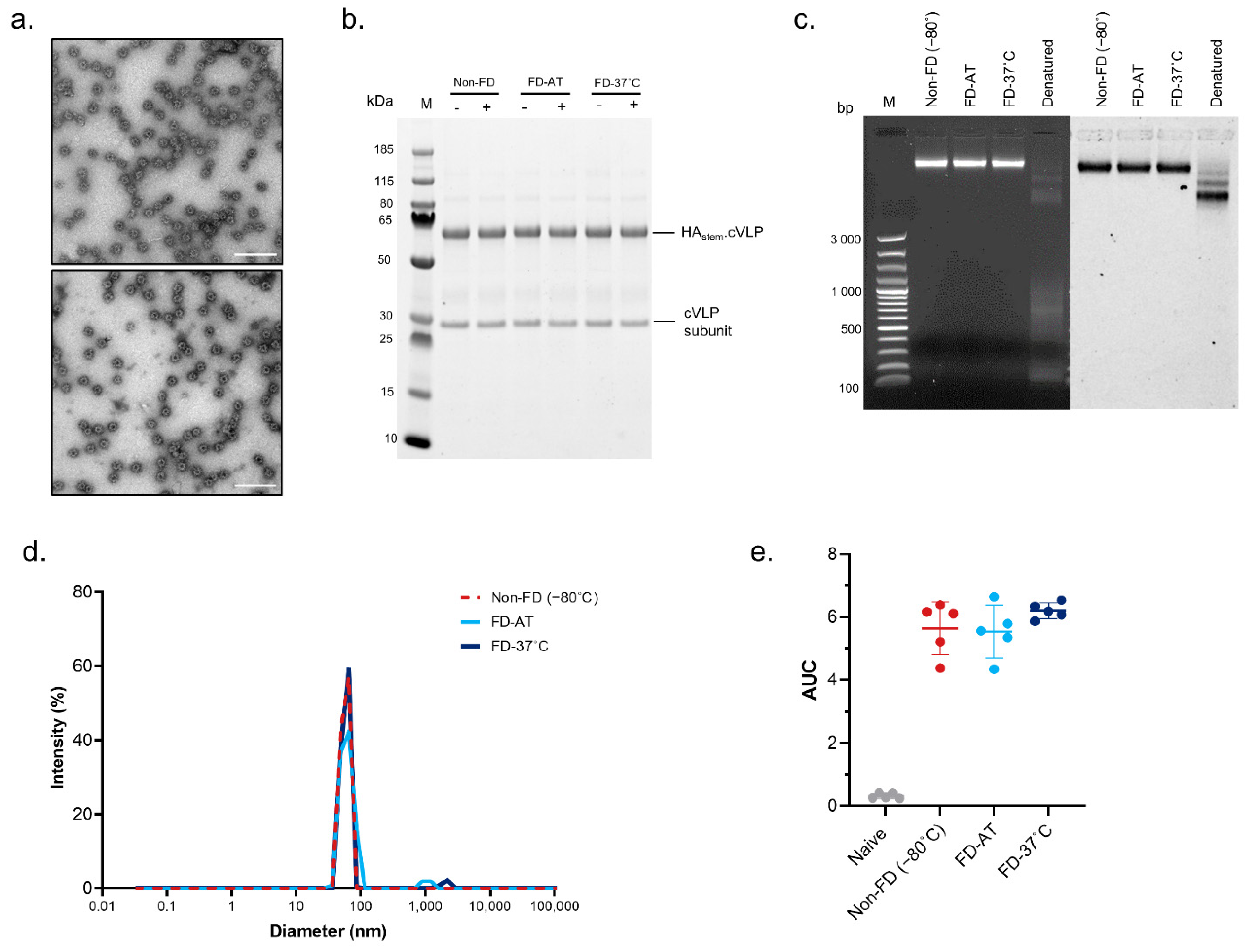
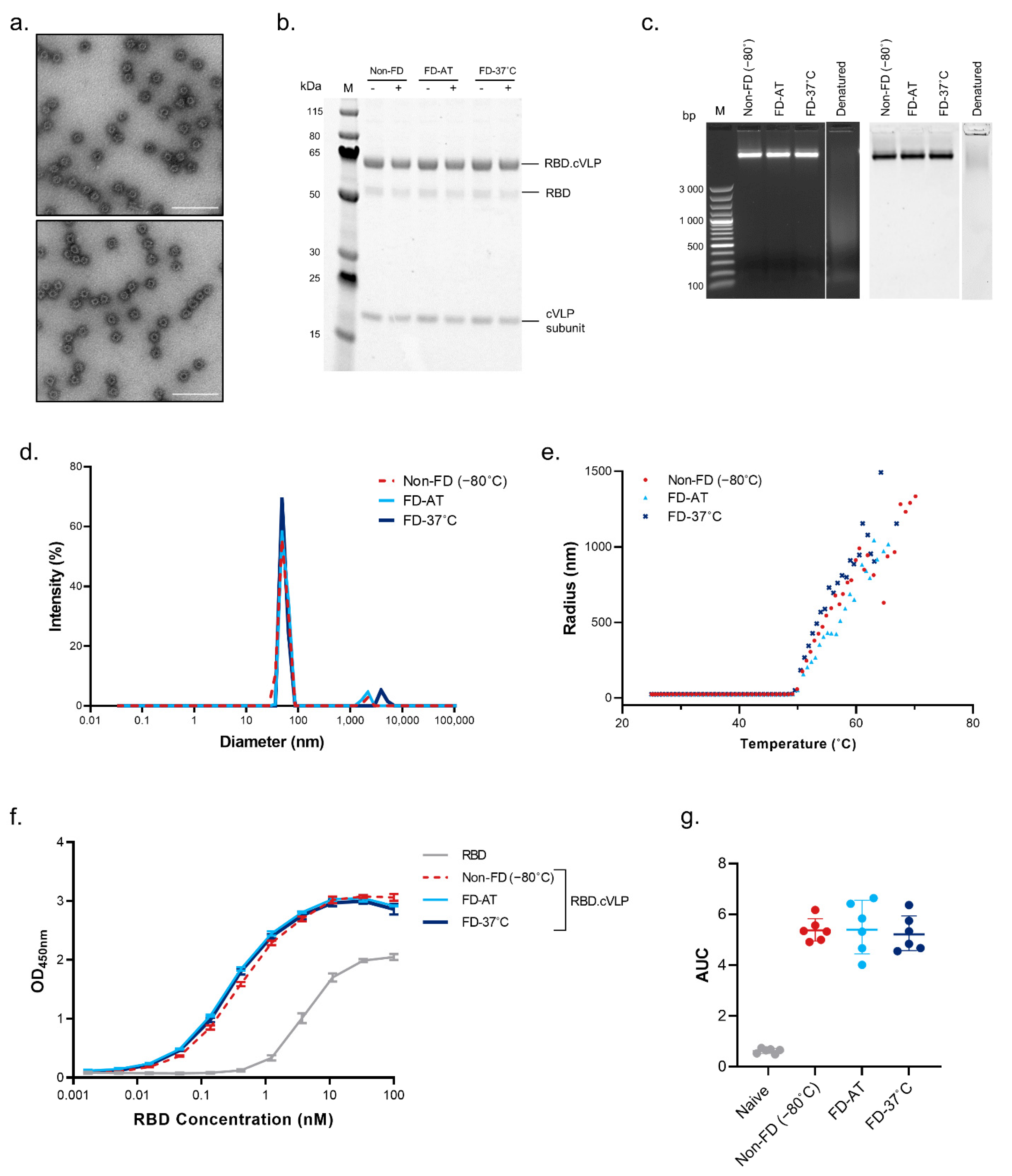
3.2. Effect of Freeze-Drying on the Structural Stability of HAstem-cVLP and RBD.cVLP Vaccines
3.3. Effect of Freeze-Drying on the Immunogenicity of HAstem-cVLP and RBD.cVLP Vaccines
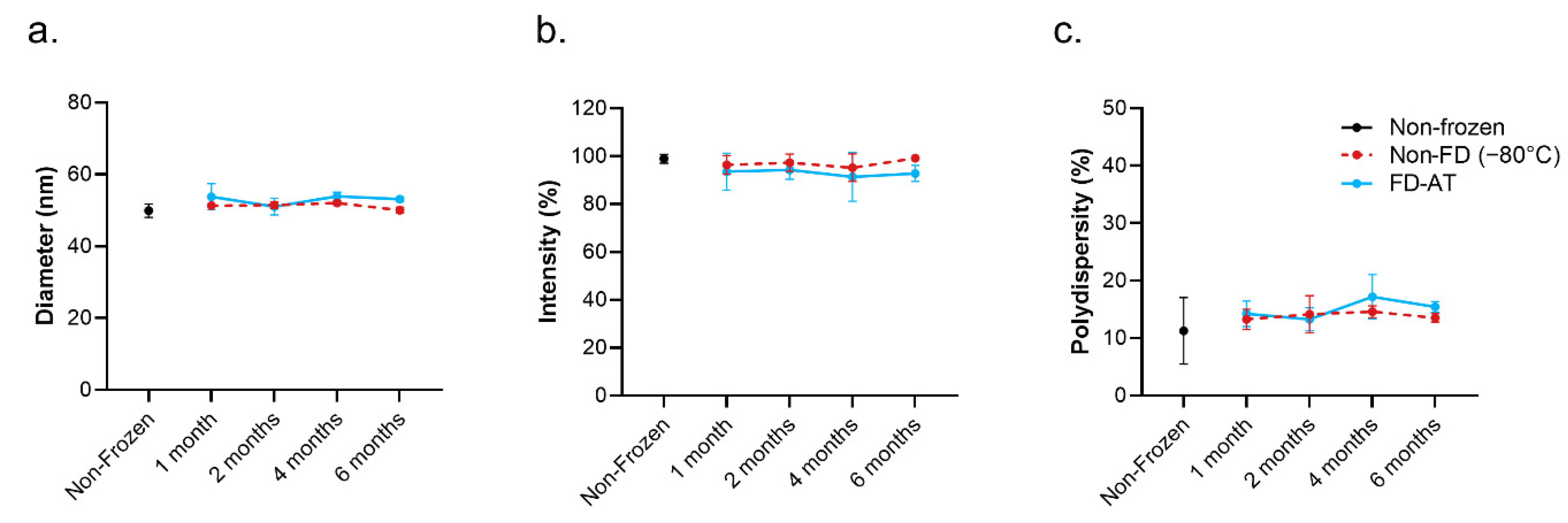
3.4. Long-Term Stability of RBD.cVLP
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kelly, H.G.; Kent, S.J.; Wheatley, A.K. Immunological Basis for Enhanced Immunity of Nanoparticle Vaccines. Expert Rev. Vaccines 2019, 18, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Bachmann, M.F.; Zinkernagel, R.M. The Influence of Virus Structure on Antibody Responses and Virus Serotype Formation. Immunol. Today 1996, 17, 553–558. [Google Scholar] [CrossRef]
- Thrane, S.; Janitzek, C.M.; Matondo, S.; Resende, M.; Gustavsson, T.; de Jongh, W.A.; Clemmensen, S.; Roeffen, W.; van de Vegte-Bolmer, M.; van Gemert, G.J.; et al. Bacterial Superglue Enables Easy Development of Efficient Virus-like Particle Based Vaccines. J. Nanobiotechnol. 2016, 14, 30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aves, K.-L.; Goksøyr, L.; Sander, A.F. Advantages and Prospects of Tag/Catcher Mediated Antigen Display on Capsid-Like Particle-Based Vaccines. Viruses 2020, 12, 185. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palladini, A.; Thrane, S.; Janitzek, C.M.; Pihl, J.; Clemmensen, S.B.; de Jongh, W.A.; Clausen, T.M.; Nicoletti, G.; Landuzzi, L.; Penichet, M.L.; et al. Virus-like Particle Display of HER2 Induces Potent Anti-Cancer Responses. Oncoimmunology 2018, 7, e1408749. [Google Scholar] [CrossRef] [Green Version]
- Janitzek, C.M.; Peabody, J.; Thrane, S.; Carlsen, P.H.R.; Theander, T.G.; Salanti, A.; Chackerian, B.; Nielsen, M.A.; Sander, A.F. A Proof-of-Concept Study for the Design of a VLP-Based Combinatorial HPV and Placental Malaria Vaccine. Sci. Rep. 2019, 9, 5260. [Google Scholar] [CrossRef] [Green Version]
- Escolano, A.; Gristick, H.B.; Abernathy, M.E.; Merkenschlager, J.; Gautam, R.; Oliveira, T.Y.; Pai, J.; West, J.P., Jr.; Barnes, C.O.; Cohen, A.A.; et al. Immunization Expands B Cells Specific to HIV-1 V3 Glycan in Mice and Macaques. Nature 2019, 570, 468–473. [Google Scholar] [CrossRef]
- Singh, S.K.; Thrane, S.; Janitzek, C.M.; Nielsen, M.A.; Theander, T.G.; Theisen, M.; Salanti, A.; Sander, A.F. Improving the Malaria Transmission-Blocking Activity of a Plasmodium Falciparum 48/45 Based Vaccine Antigen by SpyTag/SpyCatcher Mediated Virus-like Display. Vaccine 2017, 35, 3726–3732. [Google Scholar] [CrossRef]
- Thrane, S.; Aves, K.-L.; Uddbäck, I.E.M.; Janitzek, C.M.; Han, J.; Yang, Y.R.; Ward, A.B.; Theander, T.G.; Nielsen, M.A.; Salanti, A.; et al. A Vaccine Displaying a Trimeric Influenza-A HA Stem Protein on Capsid-Like Particles Elicits Potent and Long-Lasting Protection in Mice. Vaccines 2020, 8, 389. [Google Scholar] [CrossRef]
- Fougeroux, C.; Goksøyr, L.; Idorn, M.; Soroka, V.; Myeni, S.K.; Dagil, R.; Janitzek, C.M.; Søgaard, M.; Aves, K.-L.; Horsted, E.W.; et al. Capsid-like Particles Decorated with the SARS-CoV-2 Receptor-Binding Domain Elicit Strong Virus Neutralization Activity. Nat. Commun. 2021, 12, 324. [Google Scholar] [CrossRef]
- Volkmann, A.; Koopman, G.; Mooij, P.; Verschoor, E.J.; Verstrepen, B.E.; Bogers, W.M.J.M.; Idorn, M.; Paludan, S.R.; Vang, S.; Nielsen, M.A.; et al. A Capsid Virus-like Particle-Based SARS-CoV-2 Vaccine Induces High Levels of Antibodies and Protects Rhesus Macaques. Front. Immunol. 2022, 13, 1379. [Google Scholar] [CrossRef] [PubMed]
- AdaptVac. ABNCoV2 Phase I/II Shows High-Level and Broad In Vitro Efficacy with Low Reactogenicity [Press Release]. Available online: https://www.adaptvac.com/news (accessed on 7 April 2022).
- Bavarian Nodic. Bavarian Nordic Reports Positive Topline Phase 2 Results for Its COVID-19 Vaccine Candidate [Press Release]. Available online: https://www.bavarian-nordic.com/investor/news/news.aspx?news=6440 (accessed on 7 April 2022).
- Mvundura, M.; Lorenson, K.; Chweya, A.; Kigadye, R.; Bartholomew, K.; Makame, M.; Lennon, T.P.; Mwangi, S.; Kirika, L.; Kamau, P.; et al. Estimating the Costs of the Vaccine Supply Chain and Service Delivery for Selected Districts in Kenya and Tanzania. Vaccine 2015, 33, 2697–2703. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Y.B.; Briggs, K.T.; Taraban, M.B.; Brinson, R.G.; Marino, J.P. Grand Challenges in Pharmaceutical Research Series: Ridding the Cold Chain for Biologics. Pharm. Res. 2021, 38, 3–7. [Google Scholar] [CrossRef] [PubMed]
- Guignard, A.; Praet, N.; Jusot, V.; Bakker, M.; Baril, L. Introducing New Vaccines in Low- and Middle-Income Countries: Challenges and Approaches. Expert Rev. Vaccines 2019, 18, 119–131. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Monitoring Vaccine Wastage at Country Level; World Health Organization: Geneva, Switzerland, 2005. [Google Scholar]
- Thielmann, A.; Puth, M.T.; Kersting, C.; Porz, J.; Weltermann, B. Vaccine Cold Chain in General Practices: A Prospective Study in 75 Refrigerators (Keep Cool Study). PLoS ONE 2019, 14, e0224972. [Google Scholar] [CrossRef] [PubMed]
- Payton, N.M.; Lodaya, R.N.; Padilla, A.M. Lyophilized Vaccine Development. In Practical Aspects of Vaccine Development; Kolhe, P., Ohtake, S., Eds.; Academic Press, Elsevier Inc.: London, UK, 2022; pp. 297–327. [Google Scholar]
- Izutsu, K.I. Applications of Freezing and Freeze-Drying in Pharmaceutical Formulations. In Advances in Experimental Medicine and Biology; Iwaya-Inoue, M., Ed.; Springer Nature Singapore Pte Ltd.: Singapore, 2018; Volume 1081, pp. 371–383. [Google Scholar]
- Liu, B.; Zhou, X. Freeze-Drying of Proteins. In Cryopreservation and Freeze-Drying Protocals, Methods in Molecular Biology; Wolkers, W.F., Oldenhof, H., Eds.; SpringerScience and Business Media: New York, NY, USA, 2015; Volume 1257, pp. 459–476. [Google Scholar]
- Lang, R.; Winter, G.; Vogt, L.; Zürcher, A.; Dorigo, B.; Schimmele, B. Rational Design of a Stable, Freeze-Dried Virus-like Particle-Based Vaccine Formulation. Drug Dev. Ind. Pharm. 2009, 35, 83–97. [Google Scholar] [CrossRef] [PubMed]
- Impagliazzo, A.; Milder, F.; Kuipers, H.; Wagner, M.V.; Zhu, X.; Hoffman, R.M.B.; Van Meersbergen, R.; Huizingh, J.; Wanningen, P.; Verspuij, J.; et al. A Stable Trimeric Influenza Hemagglutinin Stem as a Broadly Protective Immunogen. Science 2015, 349, 1301–1306. [Google Scholar] [CrossRef] [Green Version]
- Kumar, R.; Srivastava, V.; Baindara, P.; Ahmad, A. Thermostable Vaccines: An Innovative Concept in Vaccine Development. Expert Rev. Vaccines 2022, 1, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Uddin, M.N.; Roni, M.A. Challenges of Storage and Stability of MRNA-Based COVID-19 Vaccines. Vaccines 2021, 9, 1033. [Google Scholar] [CrossRef]
- Kerwin, B.A.; Heller, M.C.; Levin, S.H.; Randolph, T.W. Effects of Tween 80 and Sucrose on Acute Short-Term Stability and Long-Term Storage at −20 °C of a Recombinant Hemoglobin. J. Pharm. Sci. 1998, 87, 1062–1068. [Google Scholar] [CrossRef]
- Izutsu, K.I.; Yoshioka, S.; Terao, T. Decreased Protein-Stabilizing Effects of Cryoprotectants Due to Crystallization. Pharm. Res. 1993, 10, 1232–1237. [Google Scholar] [CrossRef] [PubMed]
- Izutsu, K.I.; Yoshioka, S.; Kojima, S. Physical Stability and Protein Stability of Freeze-Dried Cakes during Storage at Elevated Temperatures. Pharm. Res. 1994, 11, 995–999. [Google Scholar] [CrossRef] [PubMed]
- Pitoiset, F.; Vazquez, T.; Levacher, B.; Nehar-Belaid, D.; Dérian, N.; Vigneron, J.; Klatzmann, D.; Bellier, B. Retrovirus-Based Virus-Like Particle Immunogenicity and Its Modulation by Toll-Like Receptor Activation. J. Virol. 2017, 91, e01230-17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krueger, C.C.; Thoms, F.; Keller, E.; Leoratti, F.M.S.; Vogel, M.; Bachmann, M.F. RNA and Toll-like Receptor 7 License the Generation of Superior Secondary Plasma Cells at Multiple Levels in a B Cell Intrinsic Fashion. Front. Immunol. 2019, 10, 736. [Google Scholar] [CrossRef]
- Gomes, A.C.; Roesti, E.S.; El-Turabi, A.; Bachmann, M.F. Type of RNA Packed in VLPs Impacts IgG Class Switching—Implications for an Influenza Vaccine Design. Vaccines 2019, 7, 47. [Google Scholar] [CrossRef] [Green Version]
- Zheng, Y.; Lee, P.W.; Wang, C.; Thomas, L.D.; Stewart, P.L.; Steinmetz, N.F.; Pokorski, J.K. Freeze-Drying to Produce Efficacious CPMV Virus-like Particles. Nano Lett. 2019, 19, 2099–2105. [Google Scholar] [CrossRef]
- Stratta, L.; Capozzi, L.C.; Franzino, S.; Pisano, R. Economic Analysis of a Freeze-Drying Cycle. Processes 2020, 8, 1399. [Google Scholar] [CrossRef]
- Wang, W. Lyophilization and Development of Solid Protein Pharmaceuticals. Int. J. Pharm. 2000, 203, 1–60. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aves, K.-L.; Janitzek, C.M.; Fougeroux, C.E.; Theander, T.G.; Sander, A.F. Freeze-Drying of a Capsid Virus-like Particle-Based Platform Allows Stable Storage of Vaccines at Ambient Temperature. Pharmaceutics 2022, 14, 1301. https://doi.org/10.3390/pharmaceutics14061301
Aves K-L, Janitzek CM, Fougeroux CE, Theander TG, Sander AF. Freeze-Drying of a Capsid Virus-like Particle-Based Platform Allows Stable Storage of Vaccines at Ambient Temperature. Pharmaceutics. 2022; 14(6):1301. https://doi.org/10.3390/pharmaceutics14061301
Chicago/Turabian StyleAves, Kara-Lee, Christoph M. Janitzek, Cyrielle E. Fougeroux, Thor G. Theander, and Adam F. Sander. 2022. "Freeze-Drying of a Capsid Virus-like Particle-Based Platform Allows Stable Storage of Vaccines at Ambient Temperature" Pharmaceutics 14, no. 6: 1301. https://doi.org/10.3390/pharmaceutics14061301