
VEGF Overexpression Significantly Increases Nanoparticle-Mediated siRNA Delivery and Target-Gene Downregulation

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cell Lines and Tumors
2.2. Preparation and Characterization of Chkα siRNA PEG-PEI NPs
2.3. In Vitro RNA Interference
2.4. Immunohistochemistry Staining
2.5. Optical Imaging
2.6. In Vivo 1H MR Spectroscopic Imaging and Ex Vivo High-Resolution 1H MRS
2.7. RT-PCR and Immunoblotting
2.8. Statistics Analysis
3. Results
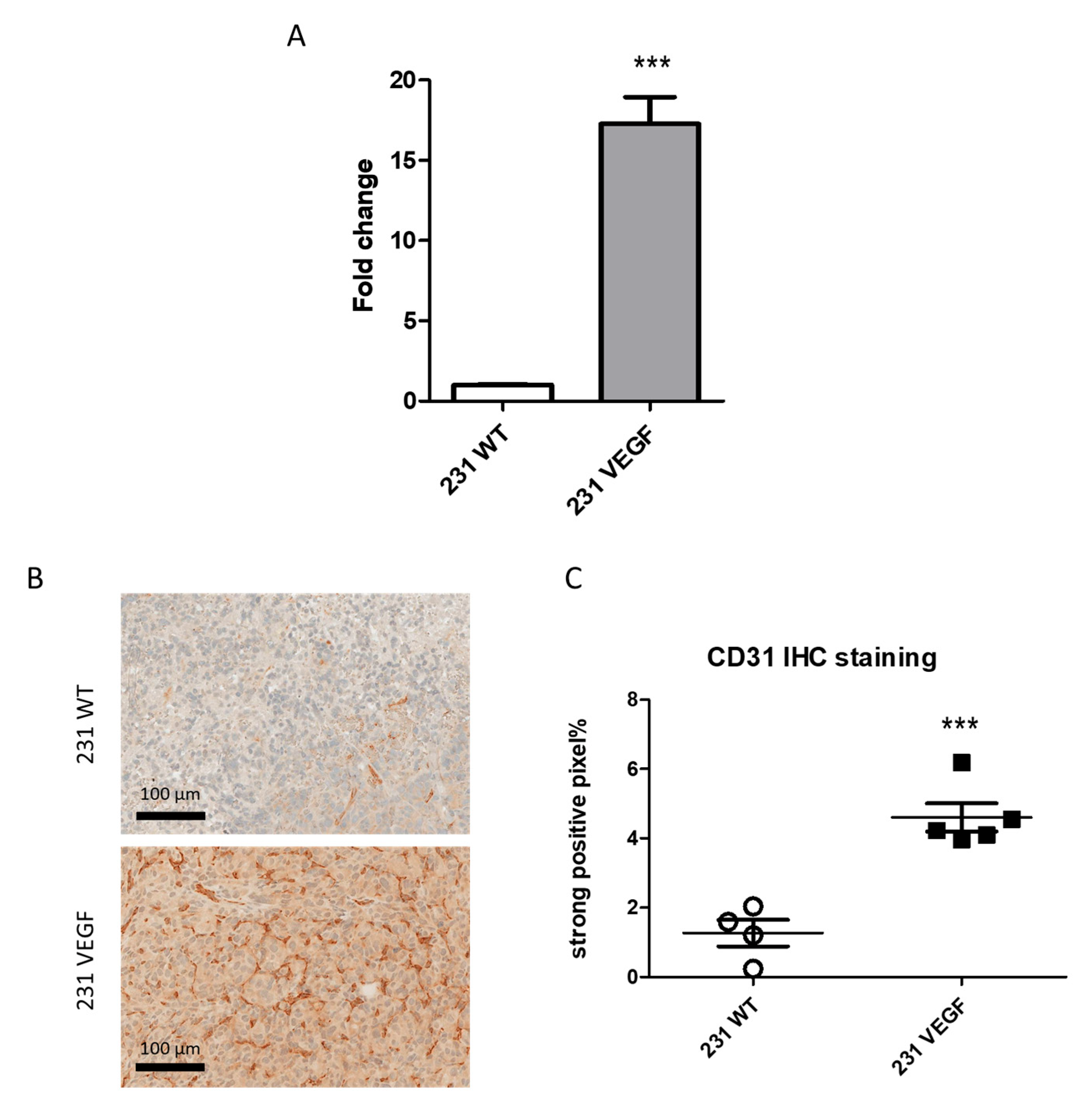
3.1. VEGF Overexpression Promotes Vascularization
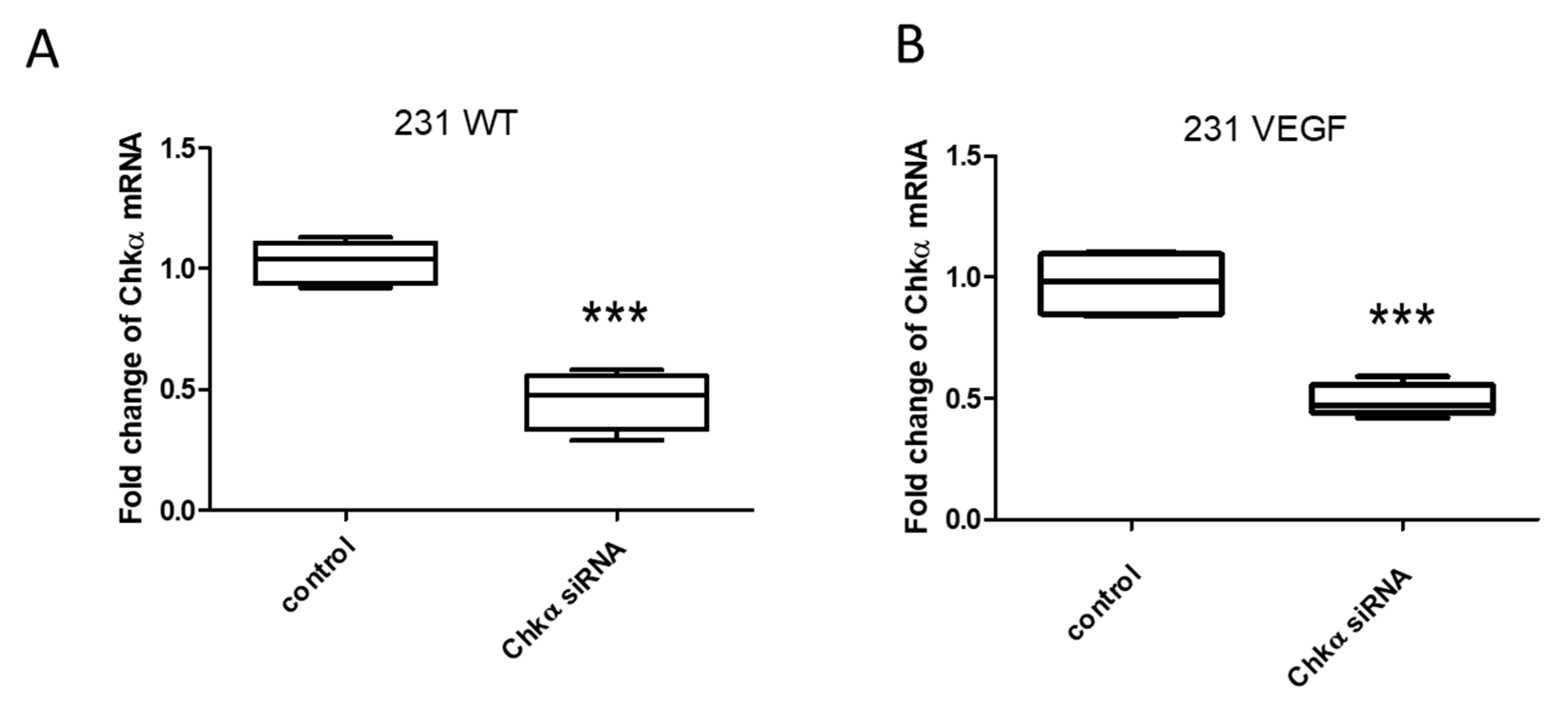
3.2. Downregulation of Chkα in Cells Following Treatment with Chkα siRNA PEG-PEI NPs
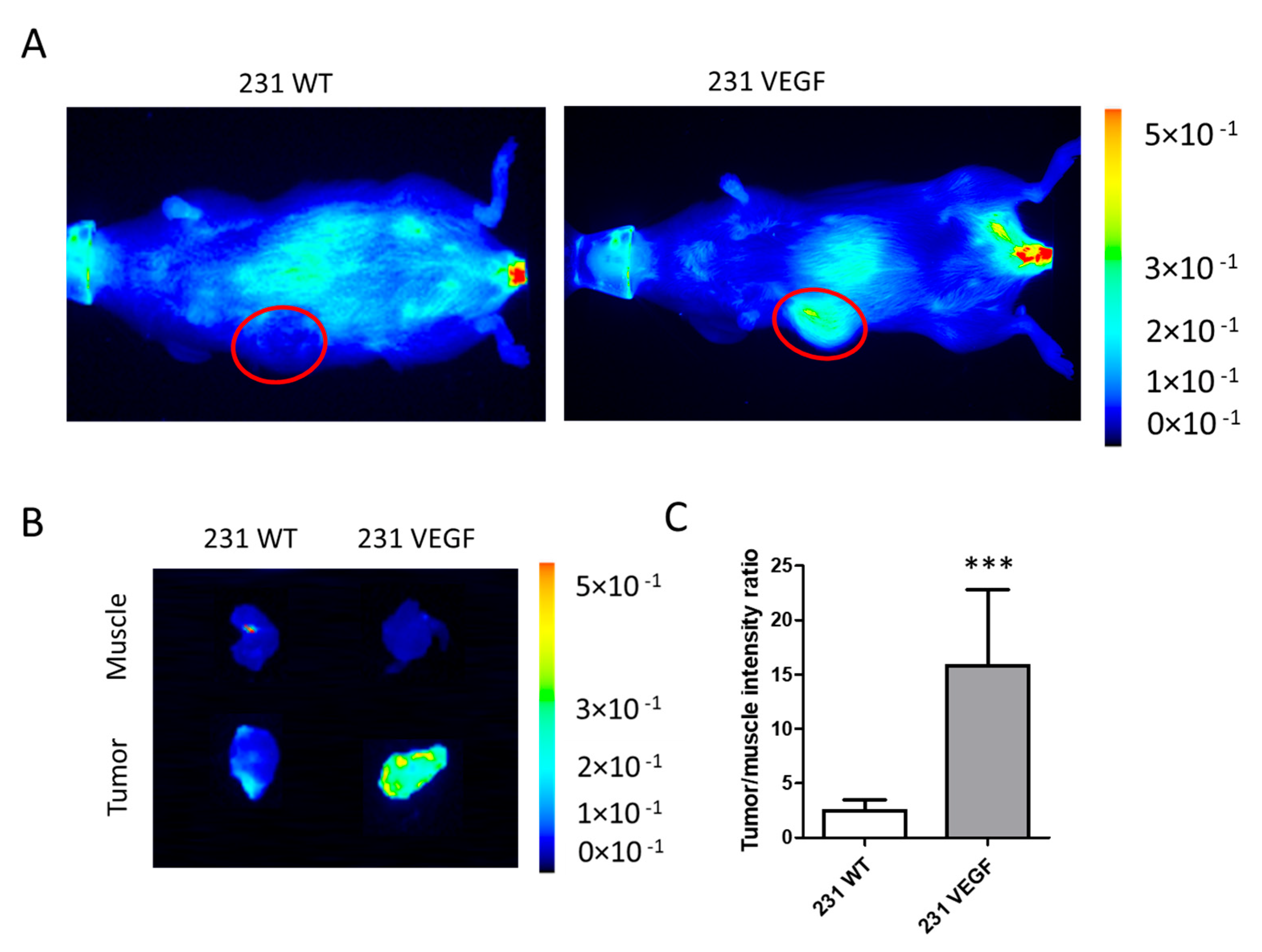
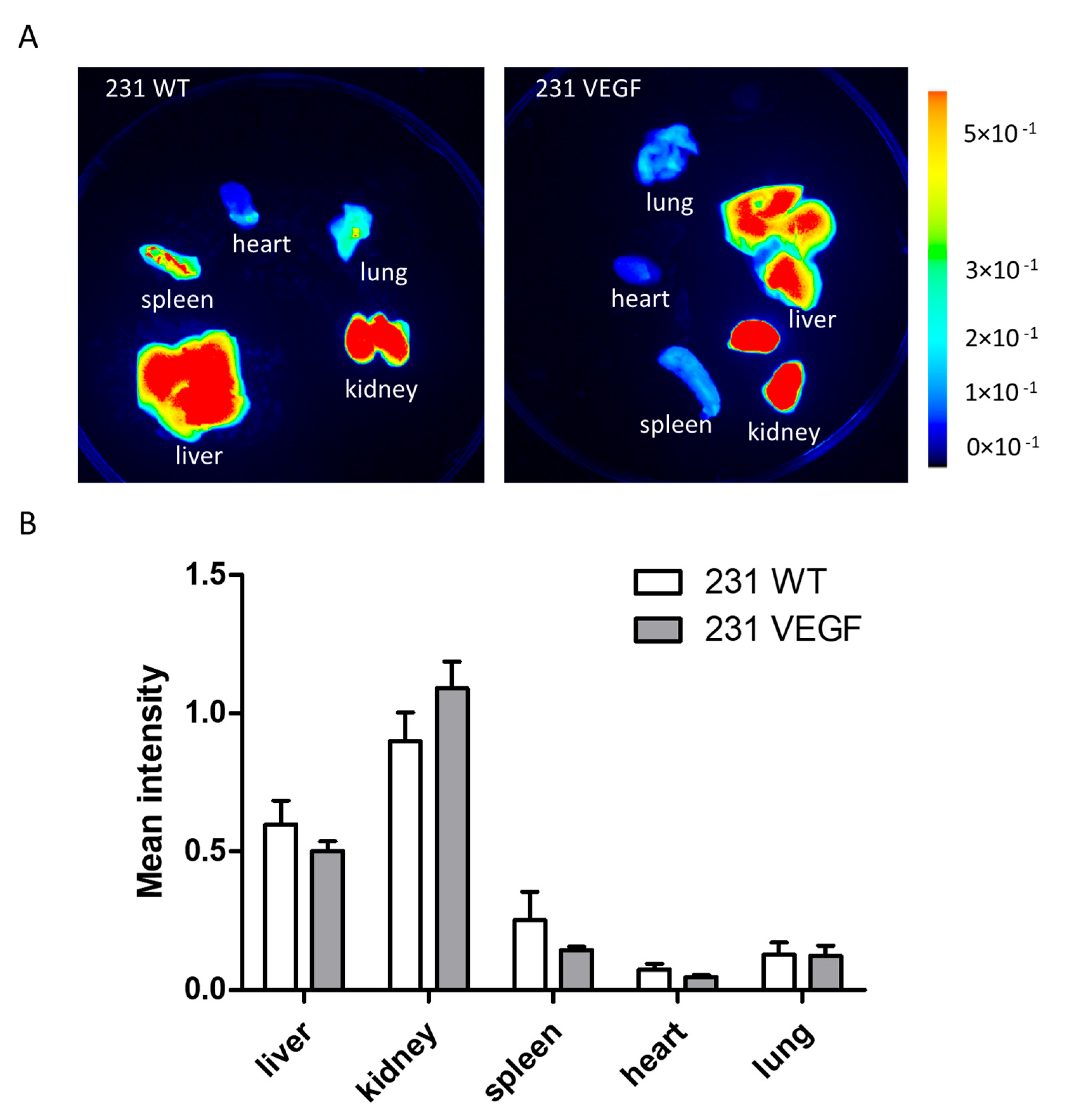
3.3. Chkα siRNA NP Biodistribution with In Vivo and Ex Vivo Optical Imaging
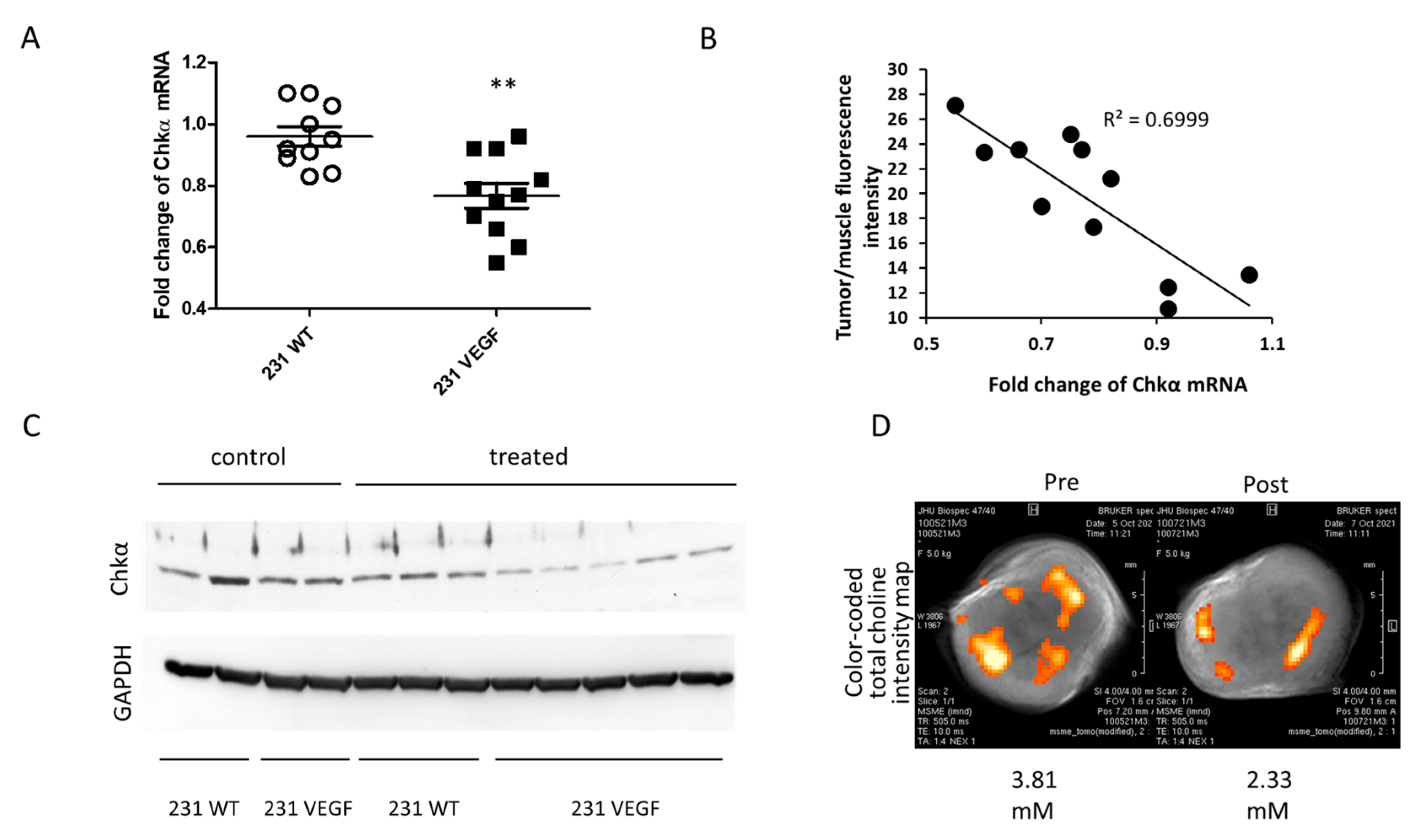
3.4. Downregulation of Chkα and Decrease in Total Choline in MDA-MB-231-VEGF Tumors
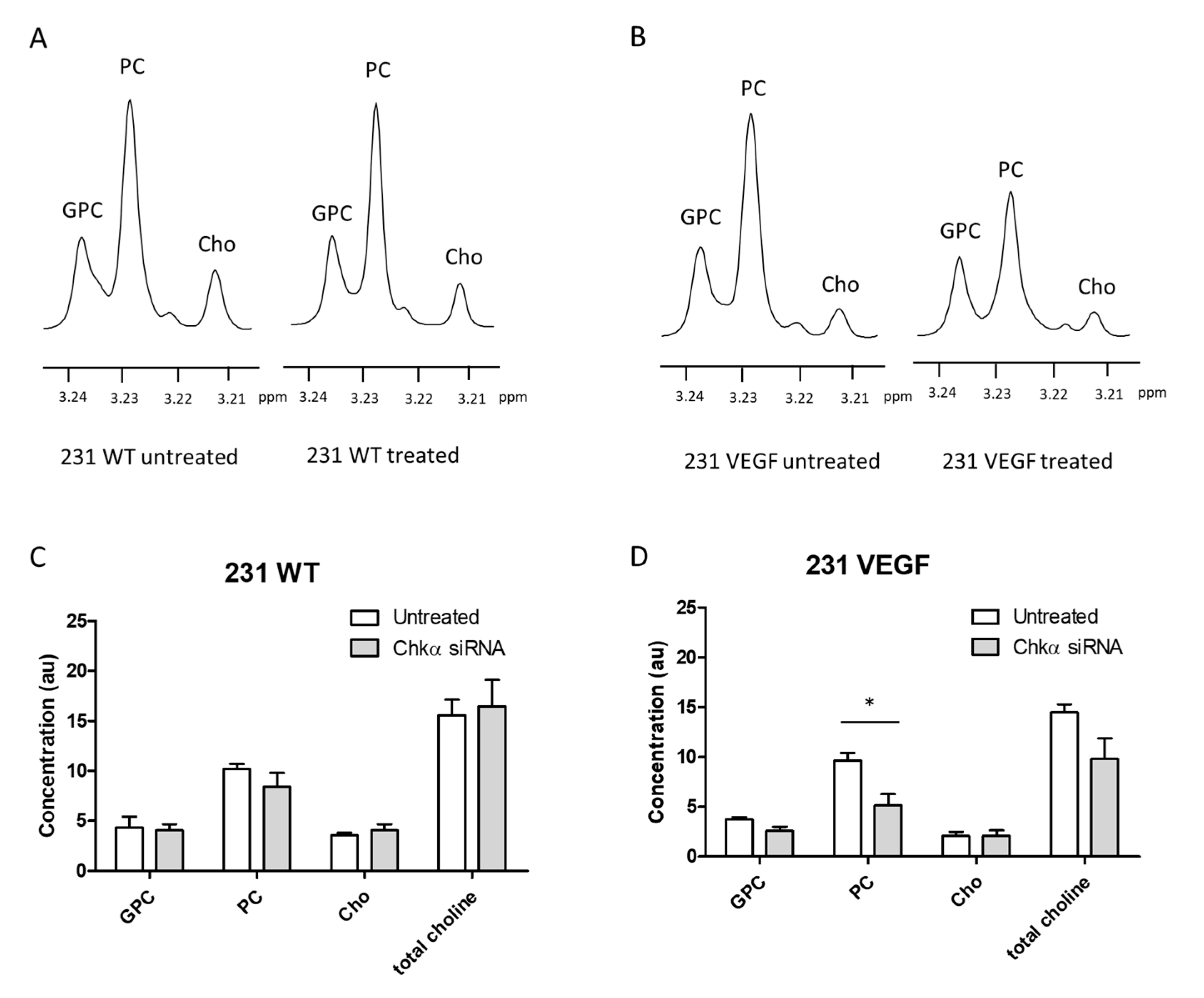
3.5. Choline Metabolite Levels in Tumor Extracts
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hu, B.; Zhong, L.; Weng, Y.; Peng, L.; Huang, Y.; Zhao, Y.; Liang, X.J. Therapeutic siRNA: State of the art. Signal Transduct. Target. Ther. 2020, 5, 101. [Google Scholar] [CrossRef] [PubMed]
- Magen, I.; Hornstein, E. Oligonucleotide-based therapy for neurodegenerative diseases. Brain Res. 2014, 1584, 116–128. [Google Scholar] [CrossRef] [PubMed]
- Danhier, F. To exploit the tumor microenvironment: Since the EPR effect fails in the clinic, what is the future of nanomedicine? J. Control. Release 2016, 244, 108–121. [Google Scholar] [CrossRef] [PubMed]
- Pacheco-Torres, J.; Penet, M.F.; Krishnamachary, B.; Mironchik, Y.; Chen, Z.; Bhujwalla, Z.M. PD-L1 siRNA Theranostics with a Dextran Nanoparticle Highlights the Importance of Nanoparticle Delivery for Effective Tumor PD-L1 Downregulation. Front. Oncol. 2020, 10, 614365. [Google Scholar] [CrossRef]
- DeSantis, C.E.; Ma, J.; Gaudet, M.M.; Newman, L.A.; Miller, K.D.; Goding Sauer, A.; Jemal, A.; Siegel, R.L. Breast cancer statistics, 2019. CA Cancer J. Clin. 2019, 69, 438–451. [Google Scholar] [CrossRef]
- Dent, R.; Trudeau, M.; Pritchard, K.I.; Hanna, W.M.; Kahn, H.K.; Sawka, C.A.; Lickley, L.A.; Rawlinson, E.; Sun, P.; Narod, S.A. Triple-negative breast cancer: Clinical features and patterns of recurrence. Clin. Cancer Res. 2007, 13, 4429–4434. [Google Scholar] [CrossRef] [Green Version]
- Nishimura, R.; Arima, N. Is triple negative a prognostic factor in breast cancer? Breast Cancer 2008, 15, 303–308. [Google Scholar] [CrossRef]
- Li, X.; Yang, J.; Peng, L.; Sahin, A.A.; Huo, L.; Ward, K.C.; O’Regan, R.; Torres, M.A.; Meisel, J.L. Triple-negative breast cancer has worse overall survival and cause-specific survival than non-triple-negative breast cancer. Breast Cancer Res. Treat. 2017, 161, 279–287. [Google Scholar] [CrossRef]
- Dent, R.; Hanna, W.M.; Trudeau, M.; Rawlinson, E.; Sun, P.; Narod, S.A. Pattern of metastatic spread in triple-negative breast cancer. Breast Cancer Res. Treat. 2009, 115, 423–428. [Google Scholar] [CrossRef]
- Glunde, K.; Bhujwalla, Z.M.; Ronen, S.M. Choline metabolism in malignant transformation. Nat. Rev. Cancer 2011, 11, 835–848. [Google Scholar] [CrossRef] [Green Version]
- Ramirez de Molina, A.; Gutierrez, R.; Ramos, M.A.; Silva, J.M.; Silva, J.; Bonilla, F.; Sanchez, J.J.; Lacal, J.C. Increased choline kinase activity in human breast carcinomas: Clinical evidence for a potential novel antitumor strategy. Oncogene 2002, 21, 4317–4322. [Google Scholar] [CrossRef] [Green Version]
- Rodriguez-Gonzalez, A.; Ramirez de Molina, A.; Fernandez, F.; Lacal, J.C. Choline kinase inhibition induces the increase in ceramides resulting in a highly specific and selective cytotoxic antitumoral strategy as a potential mechanism of action. Oncogene 2004, 23, 8247–8259. [Google Scholar] [CrossRef] [Green Version]
- Banez-Coronel, M.; Ramirez de Molina, A.; Rodriguez-Gonzalez, A.; Sarmentero, J.; Ramos, M.A.; Garcia-Cabezas, M.A.; Garcia-Oroz, L.; Lacal, J.C. Choline kinase alpha depletion selectively kills tumoral cells. Curr. Cancer Drug Targets 2008, 8, 709–719. [Google Scholar] [CrossRef]
- Falcon, S.C.; Hudson, H.C.; Huang, Y.; Mortimore, M.; Golec, J.M.; Charlton, P.A.; Weber, P.; Sundaram, H. A non-catalytic role of choline kinase alpha is important in promoting cancer cell survival. Oncogenesis 2013, 2, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Mori, N.; Wildes, F.; Kakkad, S.; Jacob, D.; Solaiyappan, M.; Glunde, K.; Bhujwalla, Z.M. Choline kinase-alpha protein and phosphatidylcholine but not phosphocholine are required for breast cancer cell survival. NMR Biomed. 2015, 28, 1697–1706. [Google Scholar] [CrossRef]
- Cao, Y.; Guangqi, E.; Wang, E.; Pal, K.; Dutta, S.K.; Bar-Sagi, D.; Mukhopadhyay, D. VEGF exerts an angiogenesis-independent function in cancer cells to promote their malignant progression. Cancer Res. 2012, 72, 3912–3918. [Google Scholar] [CrossRef] [Green Version]
- Rice, A.; Quinn, C.M. Angiogenesis, thrombospondin, and ductal carcinoma in situ of the breast. J. Clin. Pathol. 2002, 55, 569–574. [Google Scholar] [CrossRef] [Green Version]
- Kostopoulos, I.; Arapantoni-Dadioti, P.; Gogas, H.; Papadopoulos, S.; Malamou-Mitsi, V.; Scopa, C.D.; Markaki, S.; Karagianni, E.; Kyriakou, V.; Margariti, A.; et al. Evaluation of the prognostic value of HER-2 and VEGF in breast cancer patients participating in a randomized study with dose-dense sequential adjuvant chemotherapy. Breast Cancer Res. Treat. 2006, 96, 251–261. [Google Scholar] [CrossRef]
- Fuckar, D.; Dekanic, A.; Stifter, S.; Mustac, E.; Krstulja, M.; Dobrila, F.; Jonjic, N. VEGF expression is associated with negative estrogen receptor status in patients with breast cancer. Int. J. Surg. Pathol. 2006, 14, 49–55. [Google Scholar] [CrossRef]
- Pham, I.; Uchida, T.; Planes, C.; Ware, L.B.; Kaner, R.; Matthay, M.A.; Clerici, C. Hypoxia upregulates VEGF expression in alveolar epithelial cells in vitro and in vivo. Am. J. Physiol. Lung Cell Mol. Physiol. 2002, 283, L1133–L1142. [Google Scholar] [CrossRef] [Green Version]
- Pathak, A.P.; McNutt, S.; Shah, T.; Wildes, F.; Raman, V.; Bhujwalla, Z.M. In vivo “MRI phenotyping” reveals changes in extracellular matrix transport and vascularization that mediate VEGF-driven increase in breast cancer metastasis. PloS ONE 2013, 8, e63146. [Google Scholar] [CrossRef] [Green Version]
- Gilkes, D.M.; Semenza, G.L.; Wirtz, D. Hypoxia and the extracellular matrix: Drivers of tumour metastasis. Nat. Rev. Cancer 2014, 14, 430–439. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Penet, M.F.; Wildes, F.; Takagi, T.; Chen, Z.; Winnard, P.T.; Artemov, D.; Bhujwalla, Z.M. Nanoplex delivery of siRNA and prodrug enzyme for multimodality image-guided molecular pathway targeted cancer therapy. ACS Nano 2010, 4, 6707–6716. [Google Scholar] [CrossRef] [Green Version]
- Merkel, O.M.; Librizzi, D.; Pfestroff, A.; Schurrat, T.; Buyens, K.; Sanders, N.N.; De Smedt, S.C.; Behe, M.; Kissel, T. Stability of siRNA polyplexes from poly(ethylenimine) and poly(ethylenimine)-g-poly(ethylene glycol) under in vivo conditions: Effects on pharmacokinetics and biodistribution measured by Fluorescence Fluctuation Spectroscopy and Single Photon Emission Computed Tomography (SPECT) imaging. J. Control. Release 2009, 138, 148–159. [Google Scholar]
- Glunde, K.; Raman, V.; Mori, N.; Bhujwalla, Z.M. RNA interference-mediated choline kinase suppression in breast cancer cells induces differentiation and reduces proliferation. Cancer Res. 2005, 65, 11034–11043. [Google Scholar] [CrossRef] [Green Version]
- Penet, M.F.; Pathak, A.P.; Raman, V.; Ballesteros, P.; Artemov, D.; Bhujwalla, Z.M. Noninvasive multiparametric imaging of metastasis-permissive microenvironments in a human prostate cancer xenograft. Cancer Res. 2009, 69, 8822–8829. [Google Scholar] [CrossRef] [Green Version]
- Penet, M.F.; Shah, T.; Bharti, S.; Krishnamachary, B.; Artemov, D.; Mironchik, Y.; Wildes, F.; Maitra, A.; Bhujwalla, Z.M. Metabolic imaging of pancreatic ductal adenocarcinoma detects altered choline metabolism. Clin. Cancer Res. 2015, 21, 386–395. [Google Scholar] [CrossRef] [Green Version]
- Subedi, P.; Schneider, M.; Philipp, J.; Azimzadeh, O.; Metzger, F.; Moertl, S.; Atkinson, M.J.; Tapio, S. Comparison of methods to isolate proteins from extracellular vesicles for mass spectrometry-based proteomic analyses. Anal. Biochem. 2019, 584, 113390. [Google Scholar] [CrossRef]
- Weis, S.M.; Cheresh, D.A. Pathophysiological consequences of VEGF-induced vascular permeability. Nature 2005, 437, 497–504. [Google Scholar] [CrossRef]
- Chen, Z.; Krishnamachary, B.; Pachecho-Torres, J.; Penet, M.F.; Bhujwalla, Z.M. Theranostic small interfering RNA nanoparticles in cancer precision nanomedicine. Wiley Interdiscip. Rev. Nanomed. Nanobiotechnol. 2020, 12, e1595. [Google Scholar] [CrossRef]
- Whitehead, K.A.; Langer, R.; Anderson, D.G. Knocking down barriers: Advances in siRNA delivery. Nature Rev. Drug Discov. 2009, 8, 129–138. [Google Scholar] [CrossRef] [PubMed]
- Mao, S.; Sun, W.; Kissel, T. Chitosan-based formulations for delivery of DNA and siRNA. Adv. Drug Deliv. Rev. 2010, 62, 12–27. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhao, B.; Jiang, H.; Wang, B.; Ma, B. Cationic lipids and polymers mediated vectors for delivery of siRNA. J. Control. Release 2007, 123, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Tagami, T.; Suzuki, T.; Matsunaga, M.; Nakamura, K.; Moriyoshi, N.; Ishida, T.; Kiwada, H. Anti-angiogenic therapy via cationic liposome-mediated systemic siRNA delivery. Int. J. Pharm. 2012, 422, 280–289. [Google Scholar] [CrossRef]
- Pittella, F.; Zhang, M.; Lee, Y.; Kim, H.J.; Tockary, T.; Osada, K.; Ishii, T.; Miyata, K.; Nishiyama, N.; Kataoka, K. Enhanced endosomal escape of siRNA-incorporating hybrid nanoparticles from calcium phosphate and PEG-block charge-conversional polymer for efficient gene knockdown with negligible cytotoxicity. Biomaterials 2011, 32, 3106–3114. [Google Scholar] [CrossRef]
- David, S.; Resnier, P.; Guillot, A.; Pitard, B.; Benoit, J.P.; Passirani, C. siRNA LNCs—A novel platform of lipid nanocapsules for systemic siRNA administration. Eur. J. Pharm. Biopharm. 2012, 81, 448–452. [Google Scholar] [CrossRef] [Green Version]
- Mu, P.; Nagahara, S.; Makita, N.; Tarumi, Y.; Kadomatsu, K.; Takei, Y. Systemic delivery of siRNA specific to tumor mediated by atelocollagen: Combined therapy using siRNA targeting Bcl-xL and cisplatin against prostate cancer. Int. J. Cancer 2009, 125, 2978–2990. [Google Scholar] [CrossRef]
- Werth, S.; Urban-Klein, B.; Dai, L.; Hobel, S.; Grzelinski, M.; Bakowsky, U.; Czubayko, F.; Aigner, A. A low molecular weight fraction of polyethylenimine (PEI) displays increased transfection efficiency of DNA and siRNA in fresh or lyophilized complexes. J. Control. Release 2006, 112, 257–270. [Google Scholar] [CrossRef]
- Aigner, A.; Fischer, D.; Merdan, T.; Brus, C.; Kissel, T.; Czubayko, F. Delivery of unmodified bioactive ribozymes by an RNA-stabilizing polyethylenimine (LMW-PEI) efficiently down-regulates gene expression. Gene Ther. 2002, 9, 1700–1707. [Google Scholar] [CrossRef] [Green Version]
- Urban-Klein, B.; Werth, S.; Abuharbeid, S.; Czubayko, F.; Aigner, A. RNAi-mediated gene-targeting through systemic application of polyethylenimine (PEI)-complexed siRNA in vivo. Gene Ther. 2005, 12, 461–466. [Google Scholar] [CrossRef]
- Ovais, M.; Nethi, S.K.; Ullah, S.; Ahmad, I.; Mukherjee, S.; Chen, C. Recent advances in the analysis of nanoparticle-protein coronas. Nanomedicine 2020, 15, 1037–1061. [Google Scholar] [CrossRef]
- Malek, A.; Merkel, O.; Fink, L.; Czubayko, F.; Kissel, T.; Aigner, A. In vivo pharmacokinetics, tissue distribution and underlying mechanisms of various PEI(-PEG)/siRNA complexes. Toxicol. Appl. Pharmacol. 2009, 236, 97–108. [Google Scholar] [CrossRef]
- Poon, W.; Zhang, Y.N.; Ouyang, B.; Kingston, B.R.; Wu, J.L.; Wilhelm, S.; Chan, W.C. Elimination Pathways of Nanoparticles. ACS Nano 2019, 13, 5785–5798. [Google Scholar] [CrossRef]
- Burri, P.H.; Hlushchuk, R.; Djonov, V. Intussusceptive angiogenesis: Its emergence, its characteristics, and its significance. Dev. Dyn. 2004, 231, 474–488. [Google Scholar] [CrossRef]
- Jin, S.W.; Patterson, C. The opening act: Vasculogenesis and the origins of circulation. Arterioscler. Thromb. Vasc. Biol. 2009, 29, 623–629. [Google Scholar] [CrossRef]
- Lancrin, C.; Sroczynska, P.; Stephenson, C.; Allen, T.; Kouskoff, V.; Lacaud, G. The haemangioblast generates haematopoietic cells through a haemogenic endothelium stage. Nature 2009, 457, 892–895. [Google Scholar] [CrossRef] [Green Version]
- Claesson-Welsh, L.; Welsh, M. VEGFA and tumour angiogenesis. J. Intern. Med. 2013, 273, 114–127. [Google Scholar] [CrossRef]
- Semenza, G.L. HIF-1 and mechanisms of hypoxia sensing. Curr. Opin. Cell Biol. 2001, 13, 167–171. [Google Scholar] [CrossRef]
- Imoukhuede, P.I.; Popel, A.S. Quantification and cell-to-cell variation of vascular endothelial growth factor receptors. Exp. Cell Res. 2011, 317, 955–965. [Google Scholar] [CrossRef] [Green Version]
- Vempati, P.; Popel, A.S.; Mac Gabhann, F. Extracellular regulation of VEGF: Isoforms, proteolysis, and vascular patterning. Cytokine Growth Factor Rev. 2014, 25, 1–19. [Google Scholar] [CrossRef] [Green Version]
- Dudley, A.C. Tumor endothelial cells. Cold Spring Harb. Perspect. Med. 2012, 2, a006536. [Google Scholar] [CrossRef]
- Raman, V.; Artemov, D.; Pathak, A.P.; Winnard, P.T., Jr.; McNutt, S.; Yudina, A.; Bogdanov, A., Jr.; Bhujwalla, Z.M. Characterizing vascular parameters in hypoxic regions: A combined magnetic resonance and optical imaging study of a human prostate cancer model. Cancer Res. 2006, 66, 9929–9936. [Google Scholar] [CrossRef] [Green Version]
- Janardhan, S.; Srivani, P.; Sastry, G.N. Choline kinase: An important target for cancer. Curr. Med. Chem. 2006, 13, 1169–1186. [Google Scholar] [CrossRef]
- De la Cueva, A.; Ramirez de Molina, A.; Alvarez-Ayerza, N.; Ramos, M.A.; Cebrian, A.; Del Pulgar, T.G.; Lacal, J.C. Combined 5-FU and ChoKalpha inhibitors as a new alternative therapy of colorectal cancer: Evidence in human tumor-derived cell lines and mouse xenografts. PloS ONE 2013, 8, e64961. [Google Scholar] [CrossRef] [Green Version]
- Asim, M.; Massie, C.E.; Orafidiya, F.; Pertega-Gomes, N.; Warren, A.Y.; Esmaeili, M.; Selth, L.A.; Zecchini, H.I.; Luko, K.; Qureshi, A.; et al. Choline Kinase Alpha as an Androgen Receptor Chaperone and Prostate Cancer Therapeutic Target. J. Natl. Cancer Inst. 2016, 108, djv371. [Google Scholar] [CrossRef] [Green Version]
- Miyake, T.; Parsons, S.J. Functional interactions between Choline kinase alpha, epidermal growth factor receptor and c-Src in breast cancer cell proliferation. Oncogene 2012, 31, 1431–1441. [Google Scholar] [CrossRef] [Green Version]
- Krishnamachary, B.; Glunde, K.; Wildes, F.; Mori, N.; Takagi, T.; Raman, V.; Bhujwalla, Z.M. Noninvasive detection of lentiviral-mediated choline kinase targeting in a human breast cancer xenograft. Cancer Res. 2009, 69, 3464–3471. [Google Scholar] [CrossRef] [Green Version]
- Hirata, H.; Nagakura, T.; Tsujii, M.; Morita, A.; Fujisawa, K.; Uchida, A. The relationship of VEGF and PGE2 expression to extracellular matrix remodelling of the tenosynovium in the carpal tunnel syndrome. J. Pathol. 2004, 204, 605–612. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tan, S.; Chen, Z.; Mironchik, Y.; Mori, N.; Penet, M.-F.; Si, G.; Krishnamachary, B.; Bhujwalla, Z.M. VEGF Overexpression Significantly Increases Nanoparticle-Mediated siRNA Delivery and Target-Gene Downregulation. Pharmaceutics 2022, 14, 1260. https://doi.org/10.3390/pharmaceutics14061260
Tan S, Chen Z, Mironchik Y, Mori N, Penet M-F, Si G, Krishnamachary B, Bhujwalla ZM. VEGF Overexpression Significantly Increases Nanoparticle-Mediated siRNA Delivery and Target-Gene Downregulation. Pharmaceutics. 2022; 14(6):1260. https://doi.org/10.3390/pharmaceutics14061260
Chicago/Turabian StyleTan, Shanshan, Zhihang Chen, Yelena Mironchik, Noriko Mori, Marie-France Penet, Ge Si, Balaji Krishnamachary, and Zaver M. Bhujwalla. 2022. "VEGF Overexpression Significantly Increases Nanoparticle-Mediated siRNA Delivery and Target-Gene Downregulation" Pharmaceutics 14, no. 6: 1260. https://doi.org/10.3390/pharmaceutics14061260