
Nonclinical Pharmacokinetics and Pharmacodynamics Characterization of Anti-CD79b/CD3 T Cell-Dependent Bispecific Antibody Using a Surrogate Molecule: A Potential Therapeutic Agent for B Cell Malignancies

 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Generation of Test Materials
2.2. Human and Cyno CD79b Affinity Meaurement
2.3. Human and Cyno CD3ε Affinity Measurement
2.4. Ethics Statement
2.5. In Vitro B Cell Killing
2.6. In Vivo PK Study in SCID.Beige Mice
2.7. Single-Dose PK of Anti-CD79b/CD3 TDB, and Anti-gD/CD3-Low or -High Affinity TDBs in Cynomolgus Monkeys
2.8. Dose Ranging PKPD of the Surrogate Anti-cyCD79b/CD3 TDB in Cynomolgus Monkeys
2.9. Single- or Multiple-Dose Toxicokinetics (TK) of the Surrogate Anti-cyCD79b/CD3 TDB via IV or Sub-Cutaneous (SC) Administration in Cynomolgus Monkeys
2.10. 4-Week Repeat-Dose GLP Study of Anti-cyCD79b/CD3 TDB Administered via IV Infusion in Cynomolgus Monkeys with a 7-Week Recovery Period
2.11. Peripheral Blood Immunophenotyping
2.12. PK/TK Assay Evaluation in Mice and in Cynomolgus Monkeys
2.13. Anti-Drug Antibody (ADA) Assay
2.14. PK/TK Data Analysis
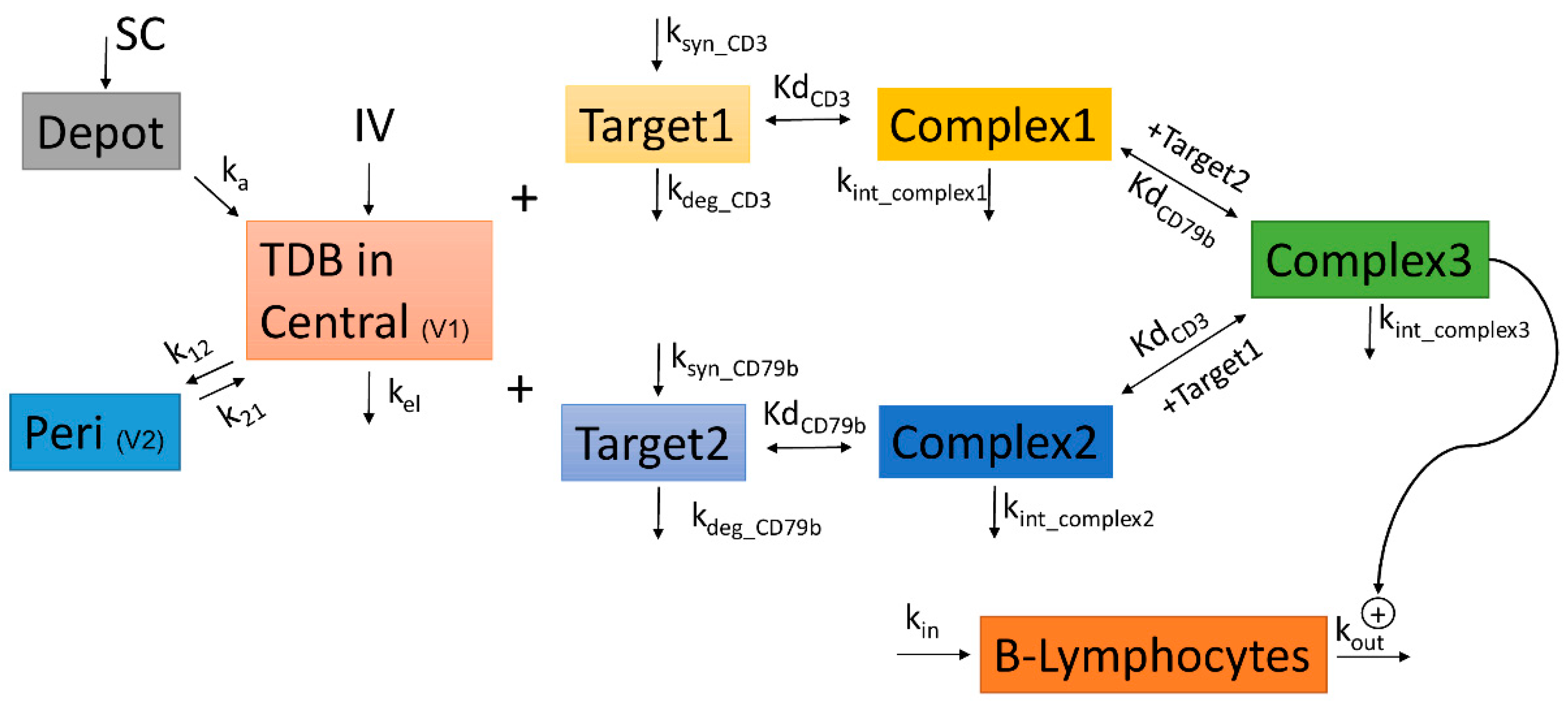
2.15. Bispecific TMDD Model Development
2.16. Estimation
3. Results
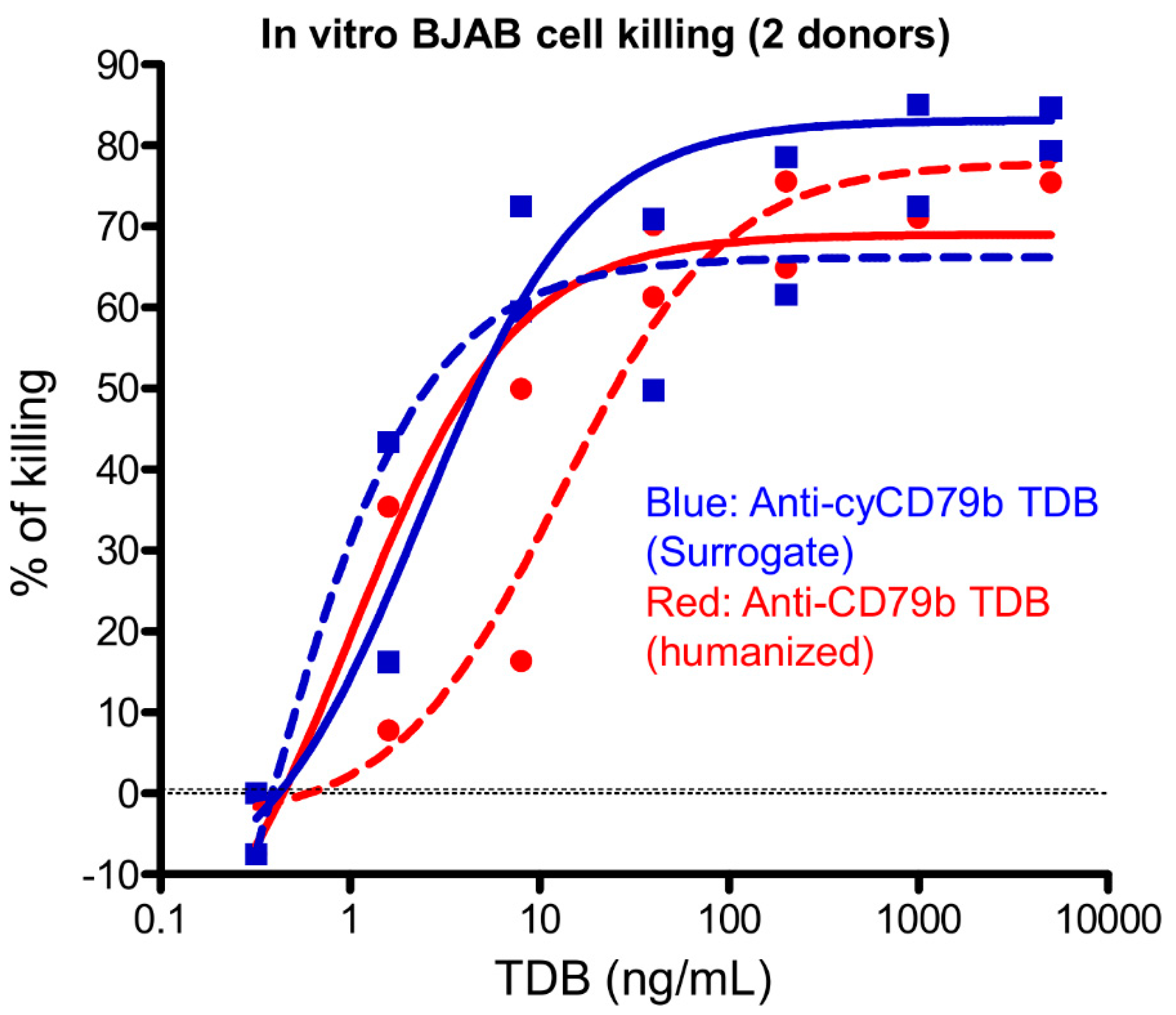
3.1. In Vitro Binding Affinity of TDBs
3.2. Anti-CD79b/CD3 and Cynomolgus Surrogate Anti-cyCD79b/CD3 TDBs Are Active against Human and Cynomolgus Monkey CD79b-Expressing B Lymphoma (BJAB) Cell Lines
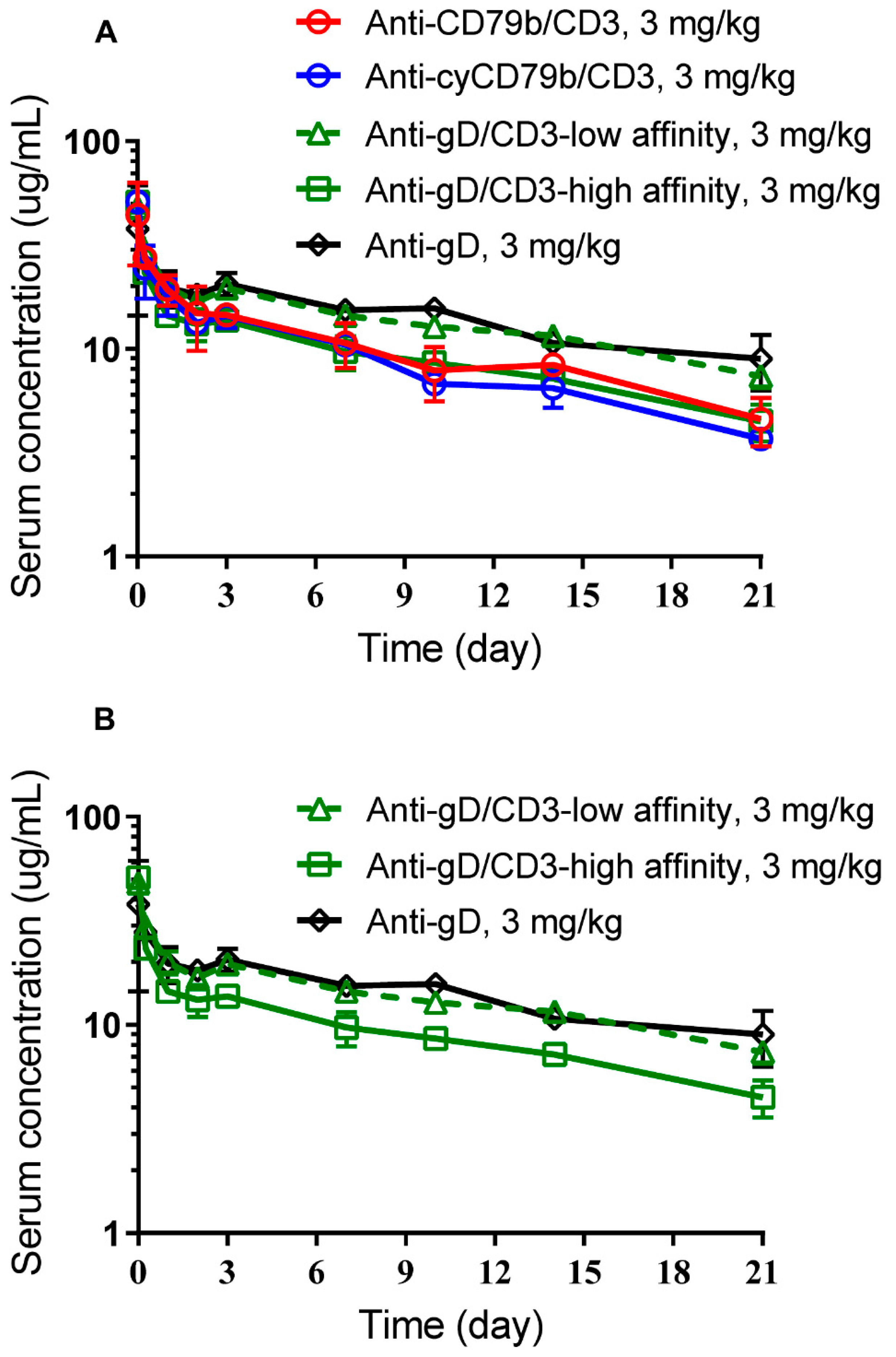
3.3. Anti-CD79b/CD3, Cynomolgus Surrogate Anti-cyCD79b/CD3, and Anti-gD/CD3-High Affinity TDBs Exhibit Comparable PK in SCID.Beige Mice
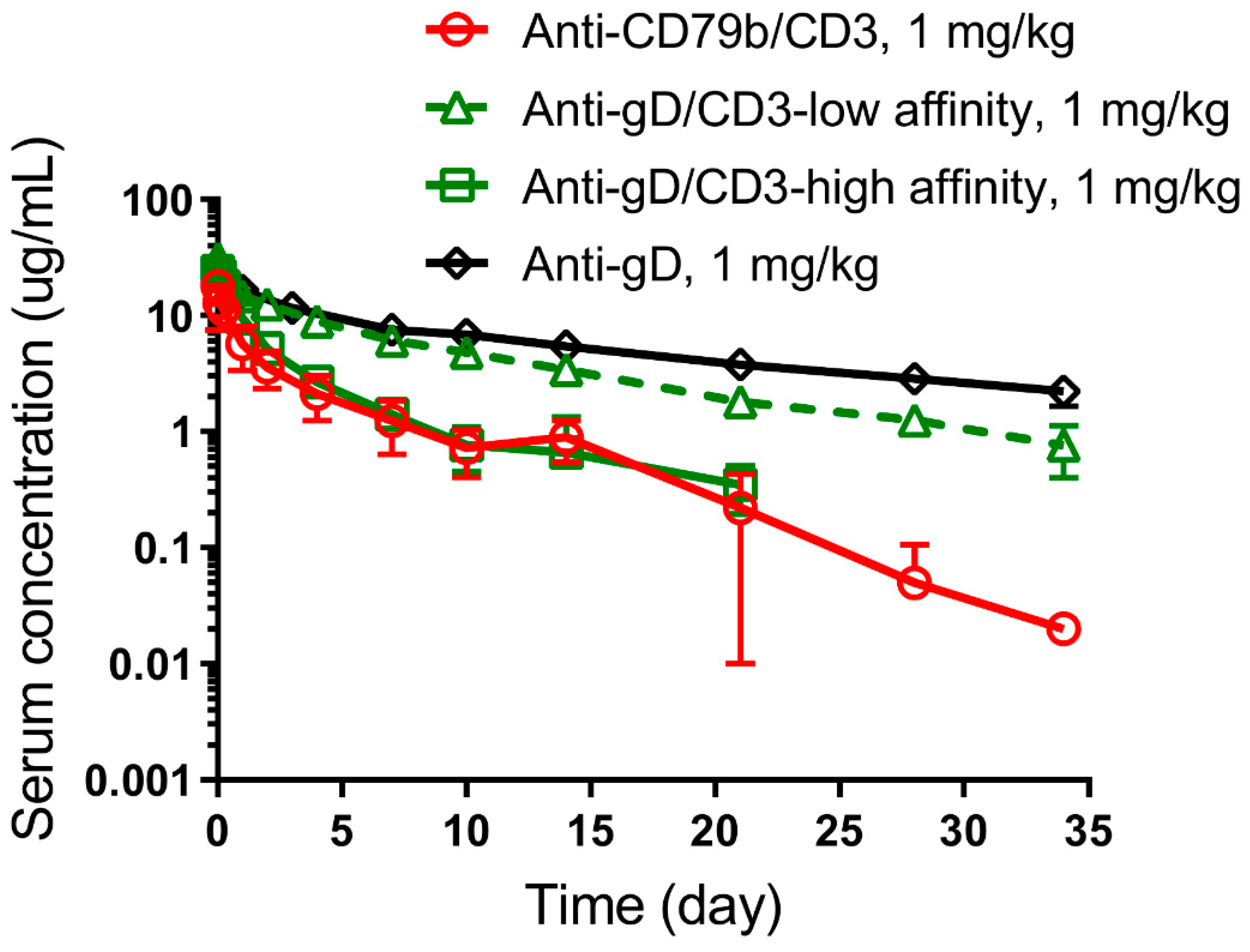
3.4. Impact of CD3 Binding Affinity on Anti-gD/CD3 TDB PK in Cynomolgus Monkeys
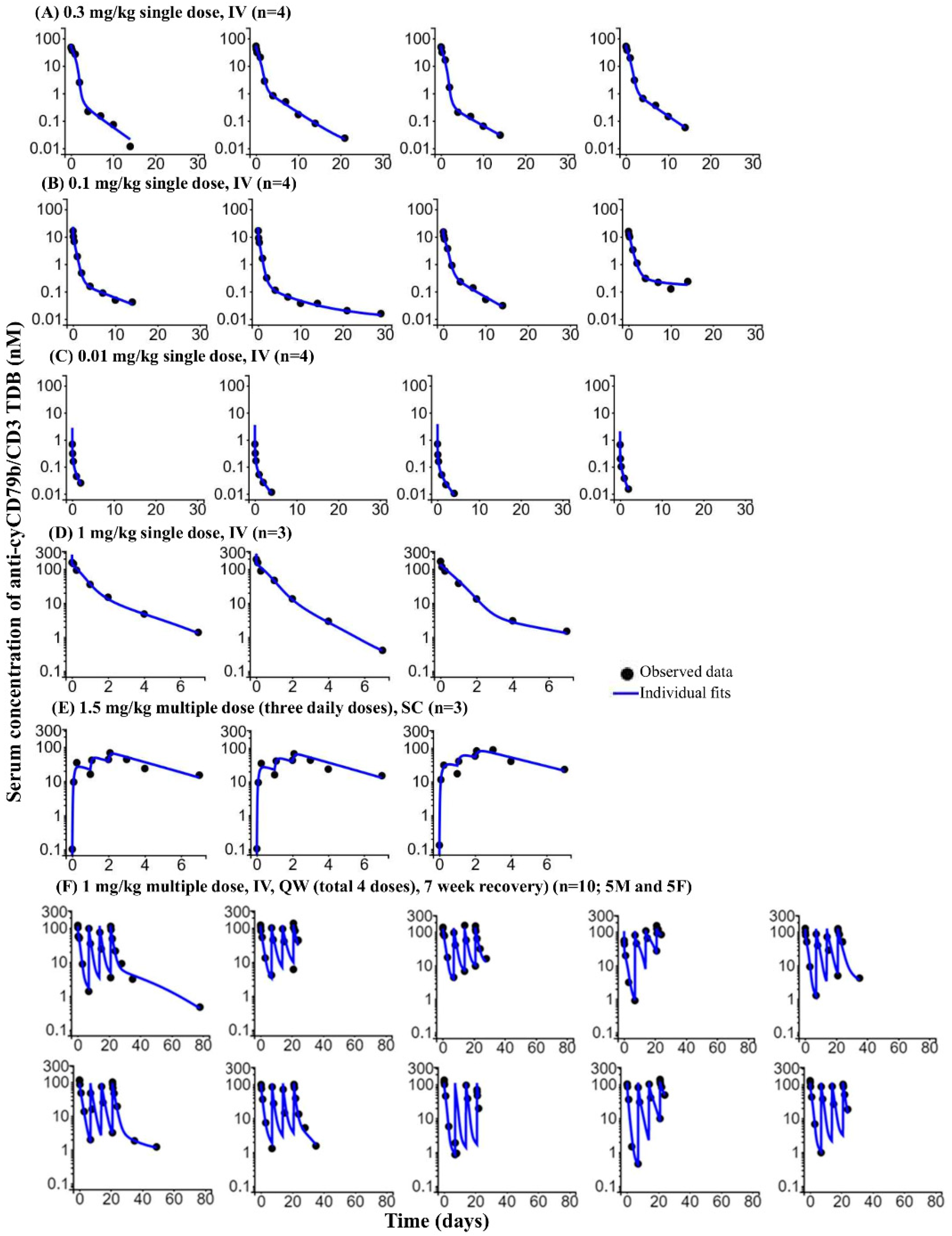
3.5. Surrogate (Anti-cyCD79b/CD3) TDB Exhibited Target-Dependent Clearance in Cynomolgus Monkey
3.6. Single- and Multiple-Dose PKPD of Anti-cyCD79b/CD3 TDB in Cynomolgus Monkeys
3.7. PKPD of Surrogate (Anti-cyCD79b/CD3) TDB in a Four-Week Repeat Dose GLP Study (with Seven-Week Recovery) in Cynomolgus Monkeys
3.8. Bispecific TMDD Model Characterized Anti-cyCD79b/CD3 Disposition Kinetics and Peripheral Blood B Cell Decrease
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Baeuerle, P.A.; Reinhardt, C. Bispecific t-cell engaging antibodies for cancer therapy. Cancer Res. 2009, 69, 4941–4944. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clevers, H.; Alarcon, B.; Wileman, T.; Terhorst, C. The t cell receptor/cd3 complex: A dynamic protein ensemble. Annu. Rev. Immunol. 1988, 6, 629–662. [Google Scholar] [CrossRef] [PubMed]
- Lejeune, M.; Köse, M.C.; Duray, E.; Einsele, H.; Beguin, Y.; Caers, J. Bispecific, t-cell-recruiting antibodies in b-cell malignancies. Front. Immunol. 2020, 11, 762. [Google Scholar] [CrossRef] [PubMed]
- Chudasama, V.L.; Zutshi, A.; Singh, P.; Abraham, A.K.; Mager, D.E.; Harrold, J.M. Simulations of site-specific target-mediated pharmacokinetic models for guiding the development of bispecific antibodies. J. Pharmacokinet. Pharmacodyn. 2015, 42, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Campagne, O.; Delmas, A.; Fouliard, S.; Chenel, M.; Chichili, G.R.; Li, H.; Alderson, R.; Scherrmann, J.M.; Mager, D.E. Integrated pharmacokinetic/pharmacodynamic model of a bispecific cd3xcd123 dart molecule in nonhuman primates: Evaluation of activity and impact of immunogenicity. Clin. Cancer Res. Off. J. Am. Assoc. Cancer Res. 2018, 24, 2631–2641. [Google Scholar] [CrossRef] [Green Version]
- Betts, A.; Haddish-Berhane, N.; Shah, D.K.; van der Graaf, P.H.; Barletta, F.; King, L.; Clark, T.; Kamperschroer, C.; Root, A.; Hooper, A.; et al. A translational quantitative systems pharmacology model for cd3 bispecific molecules: Application to quantify t cell-mediated tumor cell killing by p-cadherin lp dart®. AAPS J. 2019, 21, 66. [Google Scholar] [CrossRef] [Green Version]
- Schropp, J.; Khot, A.; Shah, D.K.; Koch, G. Target-mediated drug disposition model for bispecific antibodies: Properties, approximation, and optimal dosing strategy. CPT Pharmacomet. Syst. Pharmacol. 2019, 8, 177–187. [Google Scholar] [CrossRef]
- Riethmüller, G. Symmetry breaking: Bispecific antibodies, the beginnings, and 50 years on. Cancer Immun. 2012, 12, 12. [Google Scholar]
- Trivedi, A.; Stienen, S.; Zhu, M.; Li, H.; Yuraszeck, T.; Gibbs, J.; Heath, T.; Loberg, R.; Kasichayanula, S. Clinical pharmacology and translational aspects of bispecific antibodies. Clin. Transl. Sci. 2017, 10, 147–162. [Google Scholar] [CrossRef]
- Krishnamurthy, A.; Jimeno, A. Bispecific antibodies for cancer therapy: A review. Pharmacol. Ther. 2018, 185, 122–134. [Google Scholar] [CrossRef]
- Labrijn, A.F.; Janmaat, M.L.; Reichert, J.M.; Parren, P. Bispecific antibodies: A mechanistic review of the pipeline. Nat. Rev. Drug Discov. 2019, 18, 585–608. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Fu, J.; Zhang, M.; Liu, D. Blinatumomab: A bispecific t cell engager (bite) antibody against cd19/cd3 for refractory acute lymphoid leukemia. J. Hematol. Oncol. 2015, 8, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goebeler, M.E.; Knop, S.; Viardot, A.; Kufer, P.; Topp, M.S.; Einsele, H.; Noppeney, R.; Hess, G.; Kallert, S.; Mackensen, A.; et al. Bispecific t-cell engager (bite) antibody construct blinatumomab for the treatment of patients with relapsed/refractory non-hodgkin lymphoma: Final results from a phase i study. J. Clin. Oncol. Off. J. Am. Soc. Clin. Oncol. 2016, 34, 1104–1111. [Google Scholar] [CrossRef]
- Gökbuget, N.; Dombret, H.; Bonifacio, M.; Reichle, A.; Graux, C.; Faul, C.; Diedrich, H.; Topp, M.S.; Brüggemann, M.; Horst, H.A.; et al. Blinatumomab for minimal residual disease in adults with b-cell precursor acute lymphoblastic leukemia. Blood 2018, 131, 1522–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.L.; Ellerman, D.; Mathieu, M.; Hristopoulos, M.; Chen, X.; Li, Y.; Yan, X.; Clark, R.; Reyes, A.; Stefanich, E.; et al. Anti-cd20/cd3 t cell-dependent bispecific antibody for the treatment of b cell malignancies. Sci. Transl. Med. 2015, 7, 287ra270. [Google Scholar] [CrossRef] [PubMed]
- Hosseini, I.; Gadkar, K.; Stefanich, E.; Li, C.C.; Sun, L.L.; Chu, Y.W.; Ramanujan, S. Mitigating the risk of cytokine release syndrome in a phase i trial of cd20/cd3 bispecific antibody mosunetuzumab in nhl: Impact of translational system modeling. NPJ Syst. Biol. Appl. 2020, 6, 28. [Google Scholar] [CrossRef] [PubMed]
- Pfeifer, M.; Zheng, B.; Erdmann, T.; Koeppen, H.; McCord, R.; Grau, M.; Staiger, A.; Chai, A.; Sandmann, T.; Madle, H.; et al. Anti-cd22 and anti-cd79b antibody drug conjugates are active in different molecular diffuse large b-cell lymphoma subtypes. Leukemia 2015, 29, 1578–1586. [Google Scholar] [CrossRef]
- Wang, P.; Hristopoulos, M.; Clark, R.; Chen, Y.; Ellerman, D.; Mathieu, M.; Spiess, C.; Li, J.; Chalouni, C.; Sukumaran, S.; et al. T cell-dependent bispecific antibody anti-cd79b/cd3 as a potential therapy for b-cell malignancies. Cancer Res. 2017, 77, 3628. [Google Scholar]
- Spiess, C.; Merchant, M.; Huang, A.; Zheng, Z.; Yang, N.Y.; Peng, J.; Ellerman, D.; Shatz, W.; Reilly, D.; Yansura, D.G.; et al. Bispecific antibodies with natural architecture produced by co-culture of bacteria expressing two distinct half-antibodies. Nat. Biotechnol. 2013, 31, 753–758. [Google Scholar] [CrossRef]
- Ridgway, J.B.; Presta, L.G.; Carter, P. ‘Knobs-into-holes’ engineering of antibody ch3 domains for heavy chain heterodimerization. Protein Eng. 1996, 9, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Atwell, S.; Ridgway, J.B.; Wells, J.A.; Carter, P. Stable heterodimers from remodeling the domain interface of a homodimer using a phage display library. J. Mol. Biol. 1997, 270, 26–35. [Google Scholar] [CrossRef] [PubMed]
- van Steeg, T.J.; Bergmann, K.R.; Dimasi, N.; Sachsenmeier, K.F.; Agoram, B. The application of mathematical modelling to the design of bispecific monoclonal antibodies. mAbs 2016, 8, 585–592. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rhoden, J.J.; Dyas, G.L.; Wroblewski, V.J. A modeling and experimental investigation of the effects of antigen density, binding affinity, and antigen expression ratio on bispecific antibody binding to cell surface targets. J. Biol. Chem. 2016, 291, 11337–11347. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leong, S.R.; Sukumaran, S.; Hristopoulos, M.; Totpal, K.; Stainton, S.; Lu, E.; Wong, A.; Tam, L.; Newman, R.; Vuillemenot, B.R.; et al. An anti-cd3/anti-cll-1 bispecific antibody for the treatment of acute myeloid leukemia. Blood 2017, 129, 609–618. [Google Scholar] [CrossRef] [Green Version]
- Staflin, K.; Zuch de Zafra, C.L.; Schutt, L.K.; Clark, V.; Zhong, F.; Hristopoulos, M.; Clark, R.; Li, J.; Mathieu, M.; Chen, X.; et al. Target arm affinities determine preclinical efficacy and safety of anti-her2/cd3 bispecific antibody. JCI Insight 2020, 5, e133757. [Google Scholar] [CrossRef] [Green Version]
- Paborsky, L.R.; Fendly, B.M.; Fisher, K.L.; Lawn, R.M.; Marks, B.J.; McCray, G.; Tate, K.M.; Vehar, G.A.; Gorman, C.M. Mammalian cell transient expression of tissue factor for the production of antigen. Protein Eng. 1990, 3, 547–553. [Google Scholar] [CrossRef]
- Chen, X.; Dennis, M.S.; Ebens, A.J.; Junttila, T.T.; Kelley, R.F.; Mathieu, M.A.; Sun, L.L. Anti-cd3 Antibodies and Methods of Use. International Patent Application Publication WO2015095392A1, 25 June 2015. [Google Scholar]
- Ueda, O.; Wada, N.A.; Kinoshita, Y.; Hino, H.; Kakefuda, M.; Ito, T.; Fujii, E.; Noguchi, M.; Sato, K.; Morita, M.; et al. Entire cd3ε, δ, and γ humanized mouse to evaluate human cd3-mediated therapeutics. Sci. Rep. 2017, 7, 45839. [Google Scholar] [CrossRef]
- Bello, R.; Feito, M.J.; Ojeda, G.; Portolés, P.; Rojo, J.M. N-terminal negatively charged residues in cd3varepsilon chains as a phylogenetically conserved trait potentially yielding isoforms with different isoelectric points: Analysis of human cd3varepsilon chains. Immunol. Lett. 2009, 126, 8–15. [Google Scholar] [CrossRef] [Green Version]
- Zheng, B.; Fuji, R.N.; Elkins, K.; Yu, S.F.; Fuh, F.K.; Chuh, J.; Tan, C.; Hongo, J.A.; Raab, H.; Kozak, K.R.; et al. In vivo effects of targeting cd79b with antibodies and antibody-drug conjugates. Mol. Cancer Ther. 2009, 8, 2937–2946. [Google Scholar] [CrossRef] [Green Version]
- Carrasco-Triguero, M.; Davis, H.; Zhu, Y.; Coleman, D.; Nazzal, D.; Vu, P.; Kaur, S. Application of a plug-and-play immunogenicity assay in cynomolgus monkey serum for adcs at early stages of drug development. J. Immunol. Res. 2016, 2016, 2618575. [Google Scholar] [CrossRef] [Green Version]
- Cohen, S.; Chung, S.; Spiess, C.; Lundin, V.; Stefanich, E.; Laing, S.T.; Clark, V.; Brumm, J.; Zhou, Y.; Huang, C.; et al. An integrated approach for characterizing immunogenic responses toward a bispecific antibody. mAbs 2021, 13, 1944017. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.; Siradze, K.; Fischer, S.K. Characterization of robust immune responses to a bispecific antibody, a novel class of antibody therapeutics. Bioanalysis 2021, 13, 239–252. [Google Scholar] [CrossRef] [PubMed]
- Mager, D.E.; Jusko, W.J. General pharmacokinetic model for drugs exhibiting target-mediated drug disposition. J. Pharmacokinet. Pharmacodyn. 2001, 28, 507–532. [Google Scholar] [CrossRef] [PubMed]
- Mager, D.E.; Krzyzanski, W. Quasi-equilibrium pharmacokinetic model for drugs exhibiting target-mediated drug disposition. Pharm. Res. 2005, 22, 1589–1596. [Google Scholar] [CrossRef]
- Jiang, X.; Chen, X.; Carpenter, T.J.; Wang, J.; Zhou, R.; Davis, H.M.; Heald, D.L.; Wang, W. Development of a target cell-biologics-effector cell (tbe) complex-based cell killing model to characterize target cell depletion by t cell redirecting bispecific agents. mAbs 2018, 10, 876–889. [Google Scholar] [CrossRef]
- Pan, S.; Yu, H.; Surti, A.; Cheng, I.; Marks, S.D.; Brogan, P.A.; Eleftheriou, D.; Standing, J.F. Pharmacodynamics of rituximab on b lymphocytes in paediatric patients with autoimmune diseases. Br. J. Clin. Pharmacol. 2019, 85, 1790–1797. [Google Scholar] [CrossRef]
- Wang, B.; Liang, M.; Yao, Z.; Vainshtein, I.; Lee, R.; Schneider, A.; Zusmanovich, M.; Jin, F.; O’Connor, K.; Donato-Weinstein, B.; et al. Pharmacokinetic and pharmacodynamic comparability study of moxetumomab pasudotox, an immunotoxin targeting cd22, in cynomolgus monkeys. J. Pharm. Sci. 2013, 102, 250–261. [Google Scholar] [CrossRef]
- Li, D.; Lee, D.; Dere, R.C.; Zheng, B.; Yu, S.F.; Fuh, F.K.; Kozak, K.R.; Chung, S.; Bumbaca Yadav, D.; Nazzal, D.; et al. Evaluation and use of an anti-cynomolgus monkey cd79b surrogate antibody-drug conjugate to enable clinical development of polatuzumab vedotin. Br. J. Pharmacol. 2019, 176, 3805–3818. [Google Scholar] [CrossRef] [Green Version]
- Ovacik, A.M.; Li, J.; Lemper, M.; Danilenko, D.; Stagg, N.; Mathieu, M.; Ellerman, D.; Gupta, V.; Kalia, N.; Nguy, T.; et al. Single cell-produced and in vitro-assembled anti-fcrh5/cd3 t-cell dependent bispecific antibodies have similar in vitro and in vivo properties. mAbs 2019, 11, 422–433. [Google Scholar] [CrossRef] [Green Version]
- Diehl, K.H.; Hull, R.; Morton, D.; Pfister, R.; Rabemampianina, Y.; Smith, D.; Vidal, J.M.; van de Vorstenbosch, C. A good practice guide to the administration of substances and removal of blood, including routes and volumes. J. Appl. Toxicol. 2001, 21, 15–23. [Google Scholar] [CrossRef]
- Hötzel, I.; Theil, F.P.; Bernstein, L.J.; Prabhu, S.; Deng, R.; Quintana, L.; Lutman, J.; Sibia, R.; Chan, P.; Bumbaca, D.; et al. A strategy for risk mitigation of antibodies with fast clearance. mAbs 2012, 4, 753–760. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ryan, A.M.; Sokolowski, S.A.; Ng, C.K.; Shirai, N.; Collinge, M.; Shen, A.C.; Arrington, J.; Radi, Z.; Cummings, T.R.; Ploch, S.A.; et al. Comparative nonclinical assessments of the proposed biosimilar pf-05280586 and rituximab (mabthera®). Toxicol. Pathol. 2014, 42, 1069–1081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, L.; Xue, J.; Zhang, J.; Li, S.; Liu, D.; Zhou, T. Mechanistic prediction of first-in-human dose for bispecific cd3/epcam t-cell engager antibody m701, using an integrated pk/pd modeling method. Eur. J. Pharm. Sci. Off. J. Eur. Fed. Pharm. Sci. 2021, 158, 105584. [Google Scholar] [CrossRef] [PubMed]
- Ferl, G.Z.; Reyes, A.; Sun, L.L.; Cheu, M.; Oldendorp, A.; Ramanujan, S.; Stefanich, E.G. A preclinical population pharmacokinetic model for anti-cd20/cd3 t-cell-dependent bispecific antibodies. Clin. Transl. Sci. 2018, 11, 296–304. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Treacy, G. Using an analogous monoclonal antibody to evaluate the reproductive and chronic toxicity potential for a humanized anti-tnfalpha monoclonal antibody. Hum. Exp. Toxicol. 2000, 19, 226–228. [Google Scholar] [CrossRef]
- Green, J.D.; Terrell, T.G. Utilization of homologous proteins to evaluate the safety of recombinant human proteins--case study: Recombinant human interferon-gamma (rhifn-gamma). Toxicol. Lett. 1992, 64–65, 321–327. [Google Scholar] [CrossRef]
- Clarke, J.; Leach, W.; Pippig, S.; Joshi, A.; Wu, B.; House, R.; Beyer, J. Evaluation of a surrogate antibody for preclinical safety testing of an anti-cd11a monoclonal antibody. Regul. Toxicol. Pharmacol. 2004, 40, 219–226. [Google Scholar] [CrossRef]
- Bussiere, J.L.; Martin, P.; Horner, M.; Couch, J.; Flaherty, M.; Andrews, L.; Beyer, J.; Horvath, C. Alternative strategies for toxicity testing of species-specific biopharmaceuticals. Int. J. Toxicol. 2009, 28, 230–253. [Google Scholar] [CrossRef]
- Bornstein, G.G.; Klakamp, S.L.; Andrews, L.; Boyle, W.J.; Tabrizi, M. Surrogate approaches in development of monoclonal antibodies. Drug Discov. Today 2009, 14, 1159–1165. [Google Scholar] [CrossRef]
- Bumbaca Yadav, D.; Sharma, V.K.; Boswell, C.A.; Hotzel, I.; Tesar, D.; Shang, Y.; Ying, Y.; Fischer, S.K.; Grogan, J.L.; Chiang, E.Y.; et al. Evaluating the use of antibody variable region (fv) charge as a risk assessment tool for predicting typical cynomolgus monkey pharmacokinetics. J. Biol. Chem. 2015, 290, 29732–29741. [Google Scholar] [CrossRef] [Green Version]
- Li, J.; Stagg, N.J.; Johnston, J.; Harris, M.J.; Menzies, S.A.; DiCara, D.; Clark, V.; Hristopoulos, M.; Cook, R.; Slaga, D.; et al. Membrane-proximal epitope facilitates efficient t cell synapse formation by anti-fcrh5/cd3 and is a requirement for myeloma cell killing. Cancer Cell 2017, 31, 383–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dostalek, M.; Gardner, I.; Gurbaxani, B.M.; Rose, R.H.; Chetty, M. Pharmacokinetics, pharmacodynamics and physiologically-based pharmacokinetic modelling of monoclonal antibodies. Clin. Pharmacokinet. 2013, 52, 83–124. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.A.; Zhang, W.; Rainey, G.J.; Burke, S.; Li, H.; Huang, L.; Gorlatov, S.; Veri, M.C.; Aggarwal, S.; Yang, Y.; et al. Application of dual affinity retargeting molecules to achieve optimal redirected t-cell killing of b-cell lymphoma. Blood 2011, 117, 4542–4551. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, R.H. B cells in health and disease. Mayo Clin. Proc. 2006, 81, 377–384. [Google Scholar] [CrossRef] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Monovalent Affinity | CD79b #, KD (nM) | CD3ε [25], KD (nM) | ||
|---|---|---|---|---|
| Human | Cynomolgus | Human | Cynomolgus | |
| Anti-CD79b | 17 * | - | 14.4 | 12.8 |
| Anti-cyCD79b | - | 1 * | 14.4 | 12.8 |
| Anti-CD3-low affinity | - | - | 446 | 387 |
| Anti-CD3-high affinity | - | - | 14.4 | 12.8 |
| SCID.bg Mice | |||||
|---|---|---|---|---|---|
| PK Parameter | Anti-CD79b/CD3 | Anti-cyCD79b/CD3 | Anti-gD/CD3-Low Affinity | Anti-gD/CD3-High Affinity | Anti-gD |
| (3 mg/kg) | (3 mg/kg) | (3 mg/kg) | (3 mg/kg) | (3 mg/kg) | |
| Cmax a (μg/mL) | 44.2 ± 10.9 | 51.2 ± 2.2 | 47.9 ± 1.1 | 50.9 ± 2.0 | 37.9 ± 13.5 |
| AUC0–last (μg·day/mL) | 215.1 | 193.4 | 289.2 | 197.6 | 304.9 |
| AUC0–∞ (μg·day/mL) | 292.5 | 246.0 | 432.5 | 274.8 | 518.4 |
| CL (mL/kg/day) | 10.3 | 12.2 | 6.93 | 10.9 | 5.79 |
| Vss (mL/kg) | 161 | 162.4 | 132.2 | 180 | 136.1 |
| PK Parameter | Anti-CD79b/CD3 | Anti-gD/CD3-Low Affinity | Anti-gD/CD3-High Affinity | Anti-gD |
|---|---|---|---|---|
| Cmax (μg/mL) | 17.8 ± 5.3 | 31.1 ± 4.7 | 24.0 ± 1.05 | 27.6 ± 3.9 |
| AUC0–7 (μg·day/mL) | 25.1 ± 8.6 | 78.2 ± 4.9 | 36.9 ± 2.1 | 86.7 ± 2.2 |
| AUC0–last (μg·day/mL) | 36.2 ± 14.0 | 146 ± 18.1 | 46.1 ± 7.5 | 206 ± 17.8 |
| AUC0–∞ (μg·day/mL) | 36.3 ± 14.1 | 157 ± 24.9 | 48.1 ± 8.6 | 260 ± 42.4 |
| CL (mL/kg/day) | 30.9 ± 13.0 | 6.46 ± 0.94 | 21.3 ± 4.2 | 3.9 ± 0.62 |
| Vss (mL/kg) | 154 ± 34.0 | 72.5 ± 8.1 | 94.5 ± 32.8 | 80.1 ± 4.7 |
| PK Parameter | Anti-cyCD79b/CD3 | ||||
|---|---|---|---|---|---|
| Dose Ranging PK (Single Dose) | Single (IV) or Multiple (SC) Dose 1-Week TK | ||||
| (0.01 mg/kg) | (0.1 mg/kg) | (0.3 mg/kg) | (1 mg/kg) | (1.5 mg/kg, daily, SC × 3) | |
| Cmax (μg/mL) | 0.10 ± 0.0029 | 2.4 ± 0.11 | 7.6 ±0.30 | 25.8 ± 2.8 | 15.4 ± 6.6 |
| AUC0–7 (μg·day/mL) | 0.030 ± 0.007 | 1.5 ± 0.38 | 7.1 ± 0.80 | 19.7 ± 0.95 | 58.6 ± 34.5 |
| AUC0–last (μg·day/mL) | 0.030 ± 0.0068 | 1.5 ± 0.38 | 7.2 ± 0.82 | 19.7 ± 0.95 | 58.6 ± 34.5 |
| AUC0–∞ (μg·day/mL) | 0.033 ± 0.0074 | 1.6 ± 0.41 | 7.3 ± 0.81 | NA | NA |
| CL (mL/kg/day) | 315 ± 80.9 | 66.2 ± 16.6 | 41.8 ± 5.1 | 50.0 ± 2.3 | NA |
| Vss (mL/kg) | 284 ± 52.4 | 165 ± 105 | 43.3 ± 10.1 | 57.2 ± 9.3 | NA |
| Tmax (day) | NA | NA | NA | NA | 3.0 ± 0.96 |
| Cmax Day 0 (µg/mL) | Cmax Day 21 (µg/mL) | AUC0–7 (μg·day/mL) | AUC21–28 (μg·day/mL) | AUC0–last (μg·day/mL) | Accumulation Ratio | |
|---|---|---|---|---|---|---|
| Male | 16.9 ± 4.9 | 20.5 ± 2.8 | 23.7 ± 8.96 | 55.5 ± 22.5 | 148 ± 30.2 | 2.34 |
| Female | 17.6 ± 2.4 | 15.5 ± 3.8 | 21.4 ± 3.5 | 29.3 ± 16.9 | 107 ± 23.1 | 1.37 |
| Combined | 17.2 ± 3.6 | 18.1 ± 4.1 | 22.5 ±6.5 | 42.4 ± 123.3 | 128 ± 33.5 | 1.88 |
| Parameter | Definition | Unit | Estimate (%RSE) | Source |
|---|---|---|---|---|
| CL | Clearance | mL/day/kg | 20.0 (47.3) | Model estimated |
| CLd | Distribution clearance | mL/day/kg | 22.0 (16.0) | Model estimated |
| V1 | Central volume of distribution | mL/kg | 51.0 (6.59) | Model estimated |
| V2 | Peripheral volume of distribution | mL/kg | 90.0 (fixed) [5] | |
| kon_CD3 | CD3 association constant | 1/nM/day | 4.45 | In vitro data |
| Kd_CD3 | CD3 binding affinity (koff/kon) | nM | 12.8 | In vitro data (Table 1) |
| kon_CD79b | CD79b associate constant | 1/nM/day | 2.96 | In vitro data |
| Kd_CD79b | CD79b binding affinity (koff/kon) | nM | 1.0 | In vitro data (Table 1) |
| ln(2)/kdeg_CD3 | Degradation half-life of CD3 | day | 0.74 (32.8) | Model estimated |
| ln(2)/kdeg_CD79b | Degradation half-life of CD79b | day | 0.79 (31.7) | Model estimated |
| ln(2)/kint_complex1 | elimination half-life for complex1 | day | 346.5 (34.2) | Model estimated |
| ln(2)/kint_complex2 | elimination half-life for complex2 | day | 7.79 (63.6) | Model estimated |
| ln(2)/kint_complex3 | elimination half-life for complex3 | day | 5.77 (12.9) | Model estimated |
| ka | SC absorption rate constant | 1/day | 0.31 (14.7) | Model estimated |
| F | SC bioavailability | Fraction | 0.84 (fixed) | [45] |
| Emax | Maximum stimulation factor | unitless | 221 (49.3) | Model estimated |
| EC50 | Ternary complex concentration inducing 50% of kout | nM | 1.20 (21.0) | Model estimated |
| BSL_B-lymphocyte | Baseline of CD20+ B-lymphocyte | counts/µL | 1223.3 (14.0) | Model estimated |
| ln(2)/kout_B-lymphocyte | Degradation half-life of B-lymphocyte | day | 55 (fixed) | [38] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yadav, R.; Sukumaran, S.; Zabka, T.S.; Li, J.; Oldendorp, A.; Morrow, G.; Reyes, A.; Cheu, M.; Li, J.; Wallin, J.J.; et al. Nonclinical Pharmacokinetics and Pharmacodynamics Characterization of Anti-CD79b/CD3 T Cell-Dependent Bispecific Antibody Using a Surrogate Molecule: A Potential Therapeutic Agent for B Cell Malignancies. Pharmaceutics 2022, 14, 970. https://doi.org/10.3390/pharmaceutics14050970
Yadav R, Sukumaran S, Zabka TS, Li J, Oldendorp A, Morrow G, Reyes A, Cheu M, Li J, Wallin JJ, et al. Nonclinical Pharmacokinetics and Pharmacodynamics Characterization of Anti-CD79b/CD3 T Cell-Dependent Bispecific Antibody Using a Surrogate Molecule: A Potential Therapeutic Agent for B Cell Malignancies. Pharmaceutics. 2022; 14(5):970. https://doi.org/10.3390/pharmaceutics14050970
Chicago/Turabian StyleYadav, Rajbharan, Siddharth Sukumaran, Tanja S. Zabka, Jinze Li, Amy Oldendorp, Gary Morrow, Arthur Reyes, Melissa Cheu, Jessica Li, Jeffrey J. Wallin, and et al. 2022. "Nonclinical Pharmacokinetics and Pharmacodynamics Characterization of Anti-CD79b/CD3 T Cell-Dependent Bispecific Antibody Using a Surrogate Molecule: A Potential Therapeutic Agent for B Cell Malignancies" Pharmaceutics 14, no. 5: 970. https://doi.org/10.3390/pharmaceutics14050970