
Investigating the Mechanisms behind the Positive Food Effect of Abiraterone Acetate: In Vitro and Rat In Situ Studies

 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Chemicals
2.2. Media
2.2.1. Addition of Undigested TG: FeSSIF*-Th and FeSSIF*-E
2.2.2. Addition of Lipid Digestion Products: FeSSIF*-FM, FeSSIF*-FMT and FeSSIF*-FMTh
2.2.3. Inclusion of Lipid Digestion: FeSSIF*-Ed
2.3. Solubility Measurements
2.4. Enzymatic Conversion Experiments
2.5. Permeability Assessment Using the AMI-System
2.6. Absorptive Flux Assessment in Rats
In Situ Intestinal Perfusion
2.7. Analysis
3. Results and Discussion
3.1. Solubility
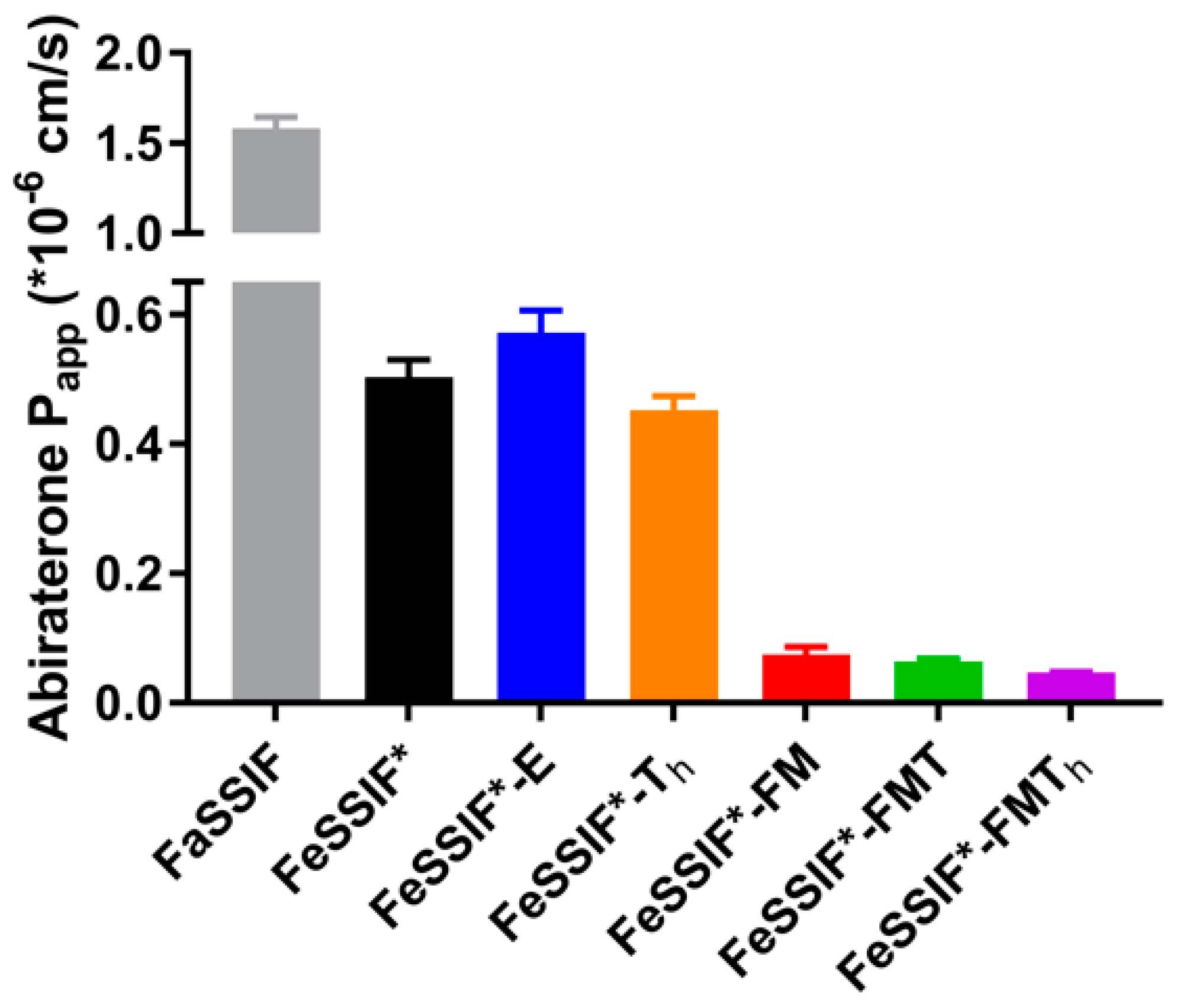
3.2. Passive Permeation (AMI-System)
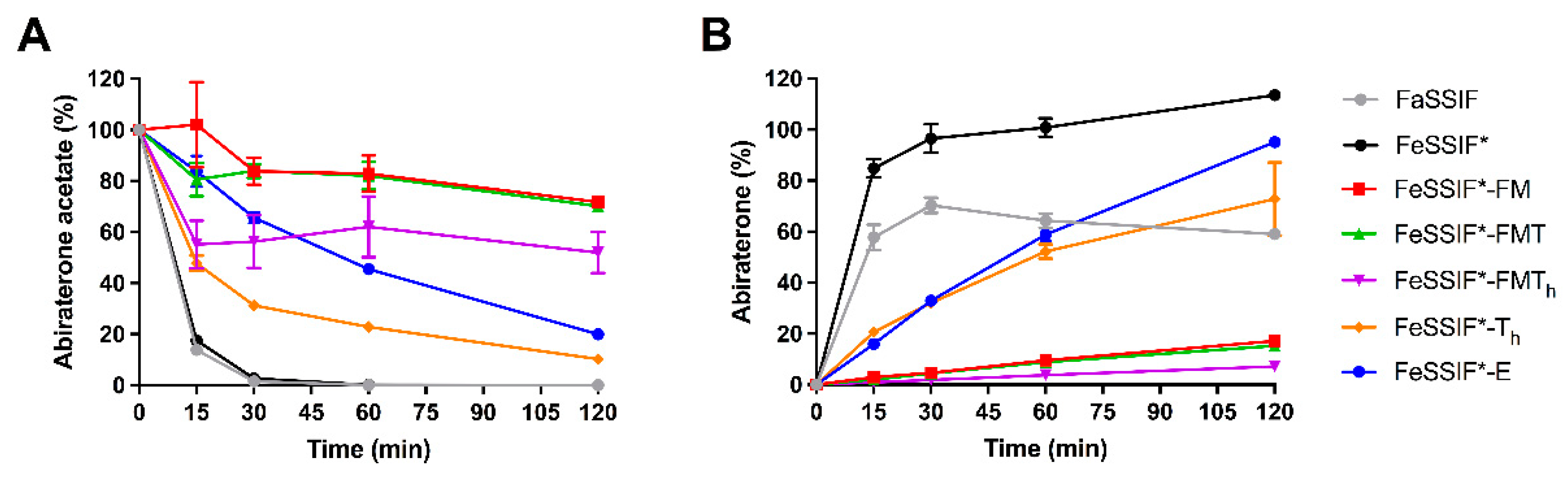
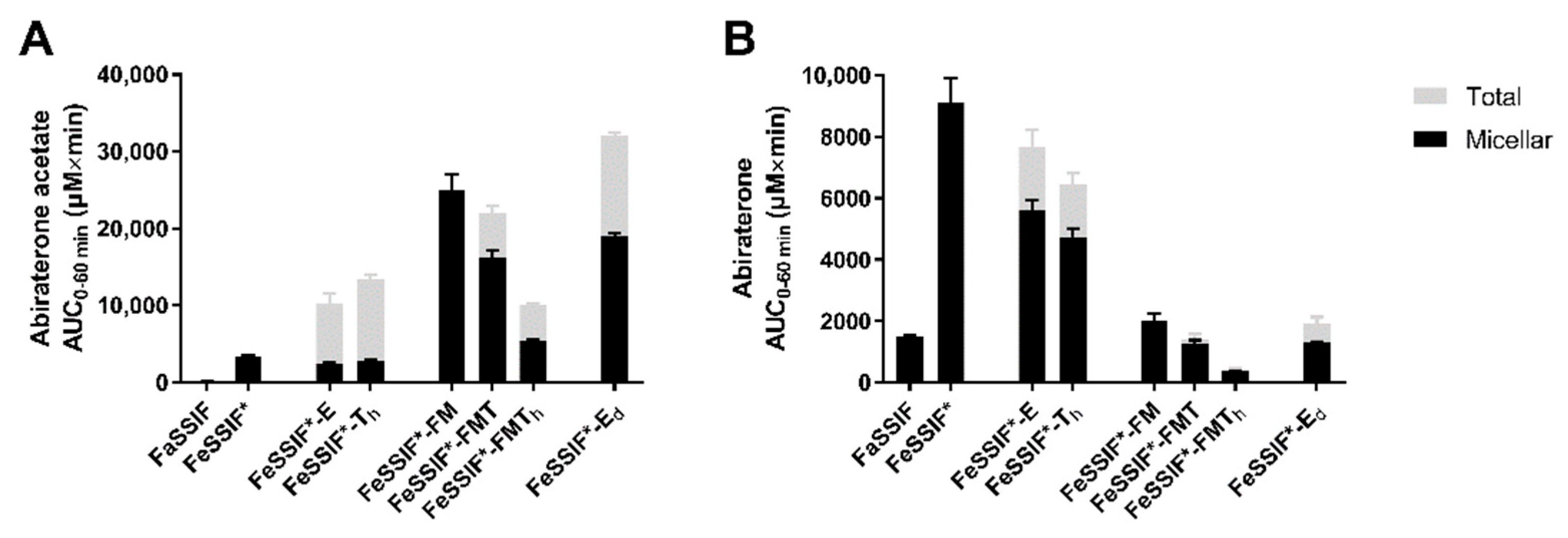
3.3. Abiraterone Acetate Hydrolysis
3.3.1. Hydrolysis in FaSSIF and Simulated Fed State Media
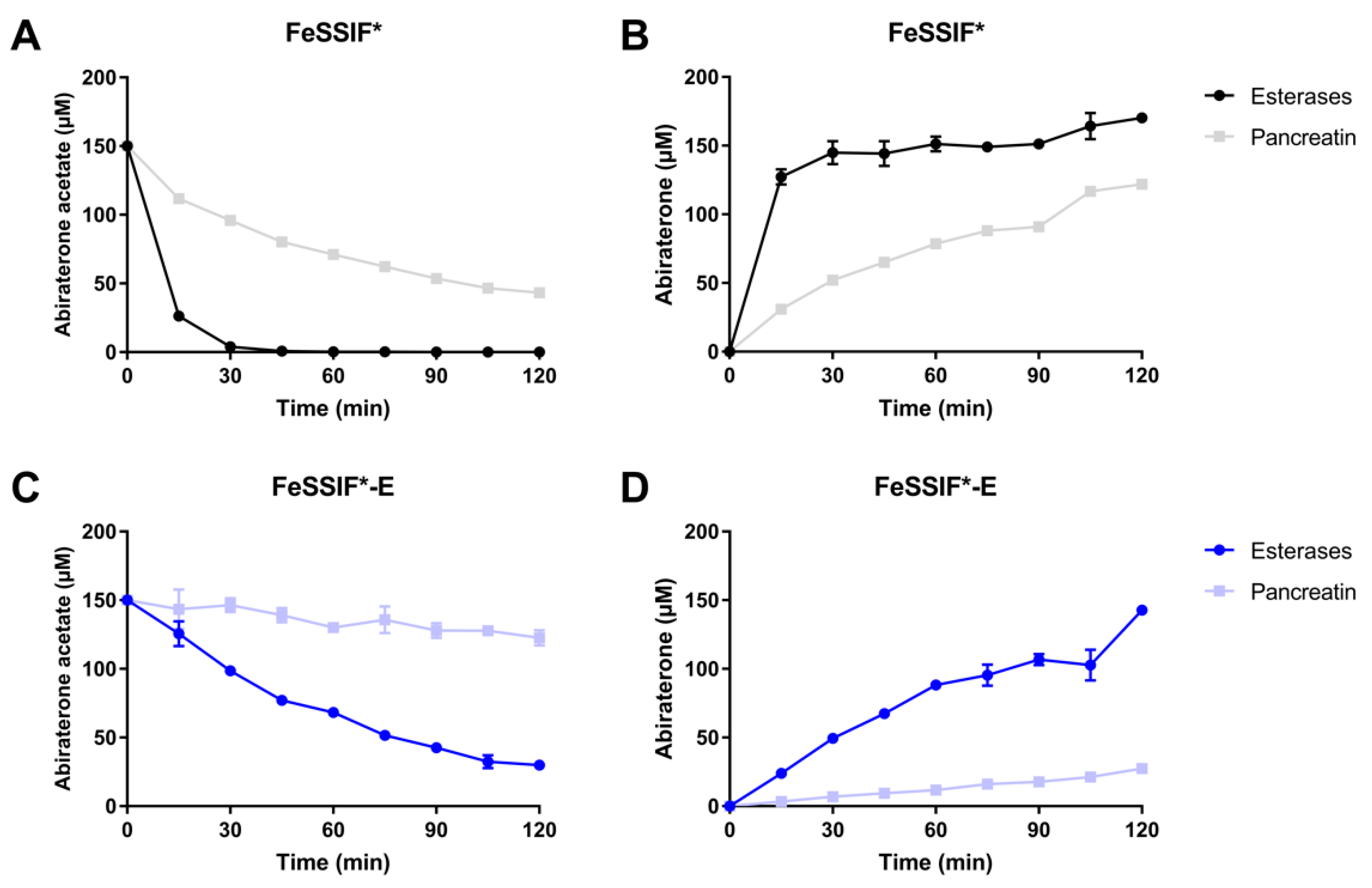
3.3.2. Pancreatin vs. Esterases
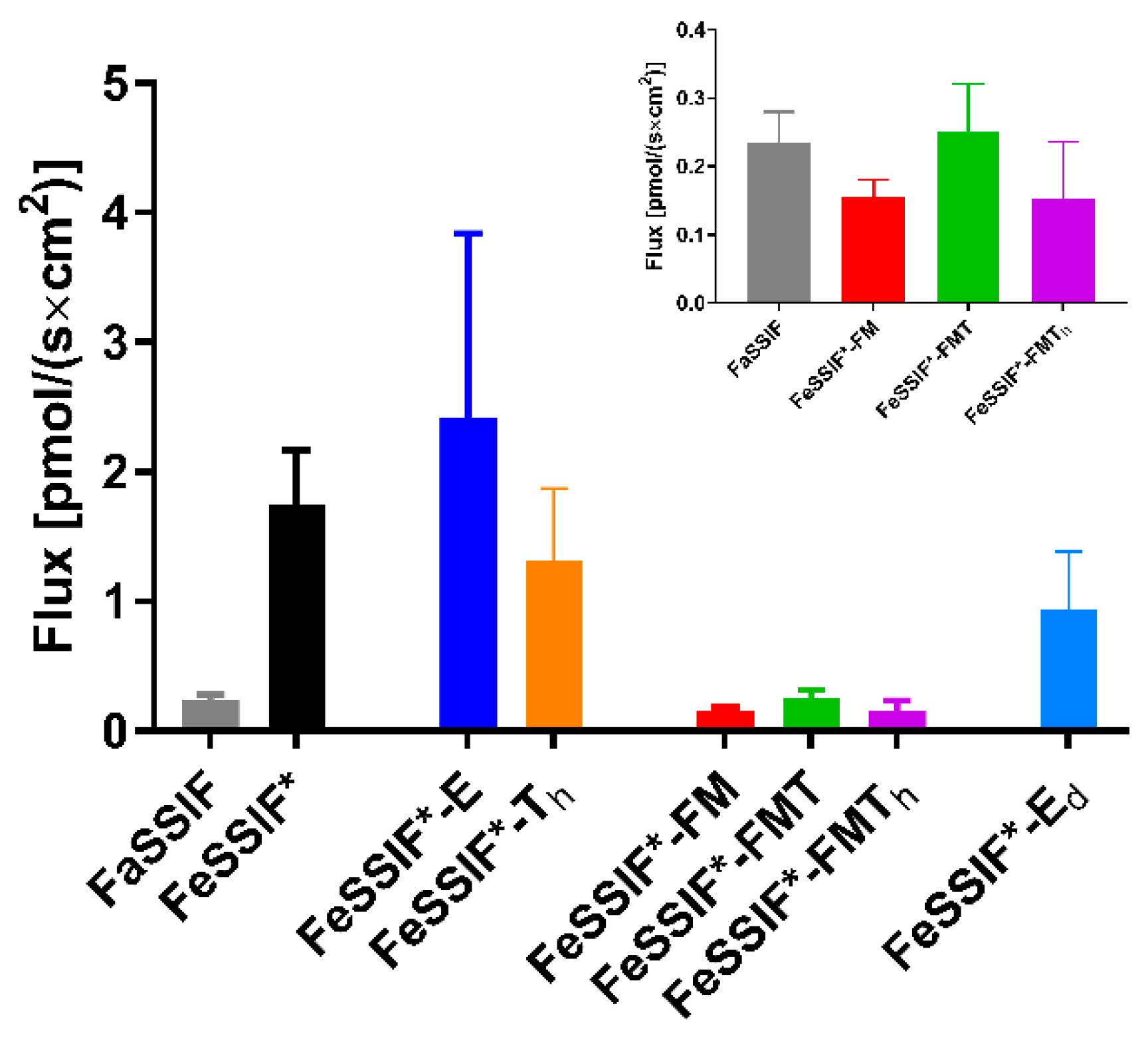
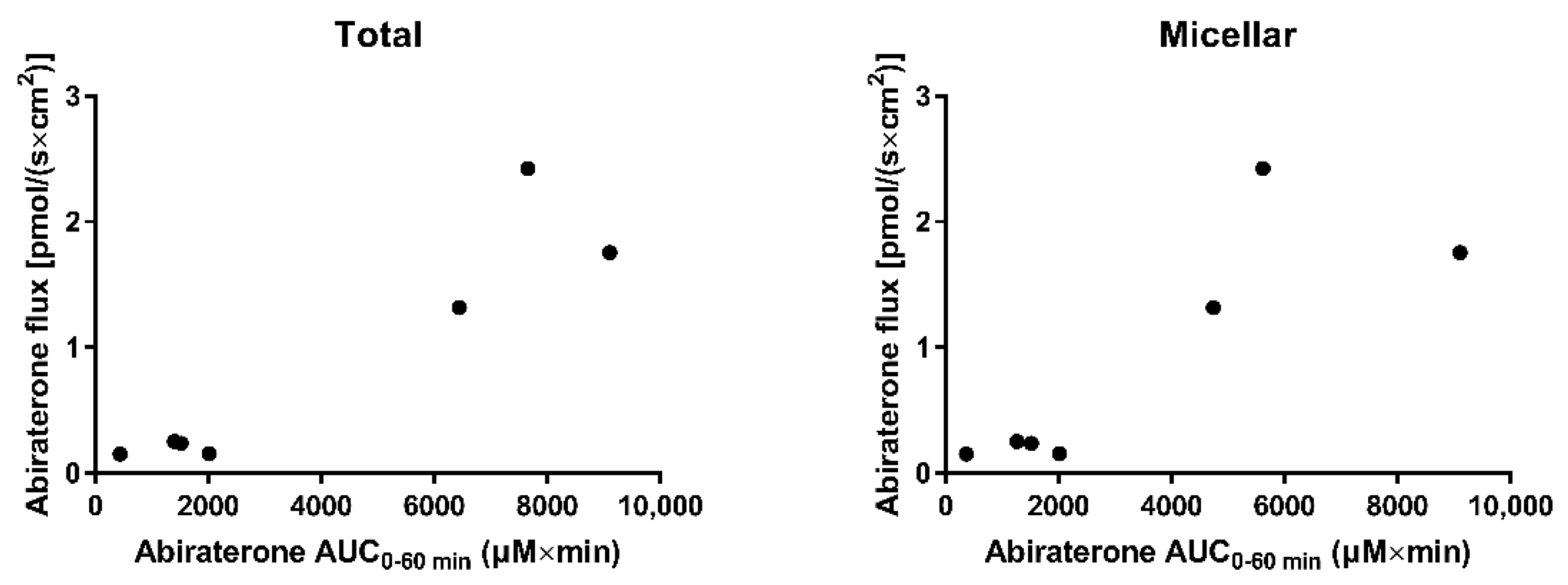
3.4. In Situ Rat Perfusion
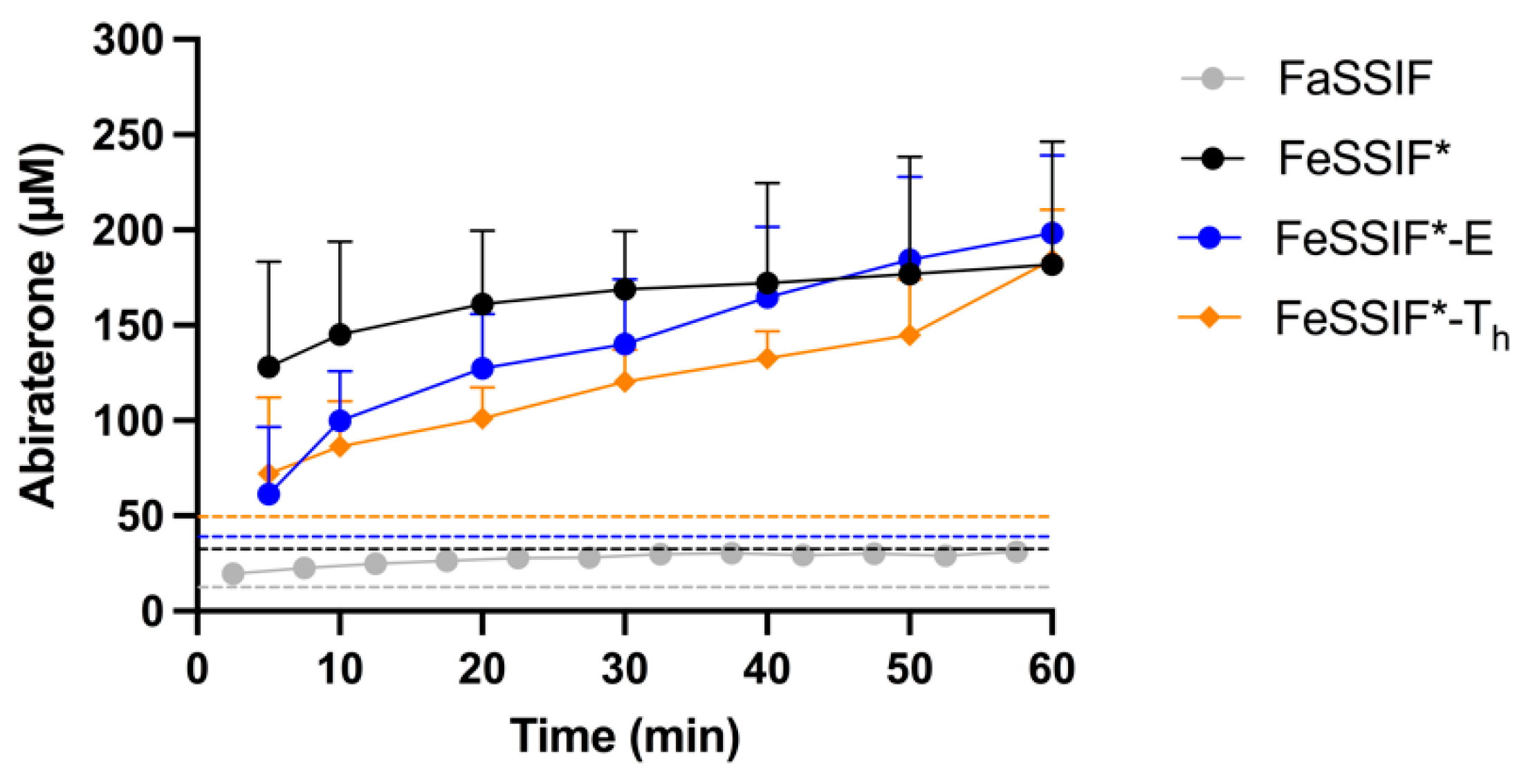
3.4.1. FaSSIF vs. FeSSIF*
3.4.2. The Effect of Undigested TG
3.4.3. The Effect of Lipid Digestion Products
3.4.4. The Effect of Ongoing Lipolysis
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Koziolek, M.; Alcaro, S.; Augustijns, P.; Basit, A.W.; Grimm, M.; Hens, B.; Hoad, C.L.; Jedamzik, P.; Madla, C.M.; Maliepaard, M.; et al. The mechanisms of pharmacokinetic food-drug interactions—A perspective from the UNGAP group. Eur. J. Pharm. Sci. 2019, 134, 31–59. [Google Scholar] [CrossRef] [PubMed]
- Müllertz, A.; Fatouros, D.G.; Smith, J.R.; Vertzoni, M.; Reppas, C. Insights into intermediate phases of human intestinal fluids visualized by atomic force microscopy and Cryo-transmission electron microscopy ex vivo. Mol. Pharm. 2012, 9, 237–247. [Google Scholar] [CrossRef] [PubMed]
- Riethorst, D.; Baatsen, P.; Remijn, C.; Mitra, A.; Tack, J.; Brouwers, J.; Augustijns, P. An In-Depth View into Human Intestinal Fluid Colloids: Intersubject Variability in Relation to Composition. Mol. Pharm. 2016, 13, 3484–3493. [Google Scholar] [CrossRef] [PubMed]
- Stappaerts, J.; Geboers, S.; Snoeys, J.; Brouwers, J.; Tack, J.; Annaert, P.; Augustijns, P. Rapid conversion of the ester prodrug abiraterone acetate results in intestinal supersaturation and enhanced absorption of abiraterone: In vitro, rat in situ and human in vivo studies. Eur. J. Pharm. Biopharm. 2015, 90, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Geboers, S.; Stappaerts, J.; Mols, R.; Snoeys, J.; Tack, J.; Annaert, P.; Augustijns, P. The Effect of Food on the Intraluminal Behavior of Abiraterone Acetate in Man. J. Pharm. Sci. 2016, 105, 2974–2981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schultz, H.B.; Meola, T.R.; Thomas, N.; Prestidge, C.A. Oral formulation strategies to improve the bioavailability and mitigate the food effect of abiraterone acetate. Int. J. Pharm. 2020, 577, 119069. [Google Scholar] [CrossRef] [PubMed]
- Acharya, M.; Gonzalez, M.; Mannens, G.; De Vries, R.; Lopez, C.; Griffin, T.; Tran, N. A phase I, open-label, single-dose, mass balance study of 14C-labeled abiraterone acetate in healthy male subjects. Xenobiotica 2012, 43, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Riethorst, D.; Mitra, A.; Kesisoglou, F.; Xu, W.; Tack, J.; Brouwers, J.; Augustijns, P. Human intestinal fluid layer separation: The effect on colloidal structures & solubility of lipophilic compounds. Eur. J. Pharm. Biopharm. 2018, 129, 104–110. [Google Scholar] [CrossRef] [PubMed]
- Berben, P.; Bauer-Brandl, A.; Brandl, M.; Faller, B.; Flaten, G.E.; Jacobsen, A.C.; Brouwers, J.; Augustijns, P. Drug permeability profiling using cell-free permeation tools: Overview and applications. Eur. J. Pharm. Sci. 2018, 119, 219–233. [Google Scholar] [CrossRef] [PubMed]
- Stappaerts, J.; Brouwers, J.; Annaert, P.; Augustijns, P. In situ perfusion in rodents to explore intestinal drug absorption: Challenges and opportunities. Int. J. Pharm. 2015, 478, 665–681. [Google Scholar] [CrossRef] [PubMed]
- Riethorst, D.; Mols, R.; Duchateau, G.; Tack, J.; Brouwers, J.; Augustijns, P. Characterization of Human Duodenal Fluids in Fasted and Fed State Conditions. J. Pharm. Sci. 2016, 105, 673–681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vertzoni, M.; Alsenz, J.; Augustijns, P.; Bauer-Brandl, A.; Bergström, C.A.S.; Brouwers, J.; Müllerz, A.; Perlovich, G.; Saal, C.; Sugano, K.; et al. UNGAP best practice for improving solubility data quality of orally administered drugs. Eur. J. Pharm. Sci. 2022, 168, 106043. [Google Scholar] [CrossRef] [PubMed]
- Berben, P.; Brouwers, J.; Augustijns, P. Assessment of Passive Intestinal Permeability Using an Artificial Membrane Insert System. J. Pharm. Sci. 2018, 107, 250–256. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Medium | FaSSIF/FeSSIF/FaSSGF Powder (mg/mL) | FFA (mg/mL) | MG (mg/mL) | TG (mg/mL) | Enzymes (mg/mL) | |
|---|---|---|---|---|---|---|
| Sodium Oleate | Glycerol Monooleate | Cream 1 | Ensure Plus 2 | Pancreatin | ||
| FaSSIF | 2.24 | |||||
| FeSSIF* | 11.2 | |||||
| FeSSIF*-FM | 11.2 | 8.8 | 3.8 | |||
| FeSSIF*-FMT | 11.2 | 8.8 | 3.8 | 0.91 | ||
| FeSSIF*-FMTh | 11.2 | 8.8 | 3.8 | 8.2 | ||
| FeSSIF*-Th | 11.2 | 8.2 | ||||
| FeSSIF*-E | 9.33 | 8.2 | ||||
| FeSSIF*-Ed | 9.33 | 8.2 | 0.83 3 | |||
| Medium | Abiraterone Acetate (µM) | Abiraterone (µM) | ||
|---|---|---|---|---|
| Micellar | Total | Micellar | Total | |
| FaSSIF | 64.6 (±5.2) 1 | n/a | 12.7 (±4.2) 1 | n/a |
| FeSSIF* | 148 (±15.7) | n/a | 32.7 (±1.34) | n/a |
| FeSSIF*-FM | 2111 (±63) | n/a | 176 (±8.63) | n/a |
| FeSSIF*-FMT | 2018 (±114) | 2284 (±103) | 161 (±7.83) | 199 (±13.6) |
| FeSSIF*-FMTh | 1283 (±53.4) | 4615 (±421) | 158 (±13.0) | 221 (±13.5) |
| FeSSIF*-Th | 330 (±18.0) | 947 (±144) | 31 (±2.55) | 50 (±2.01) |
| FeSSIF*-E | 225 (±42.3) | 892 (±92.8) | 22 (±3.24) | 39 (±10.8) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Braeckmans, M.; Augustijns, P.; Mols, R.; Servais, C.; Brouwers, J. Investigating the Mechanisms behind the Positive Food Effect of Abiraterone Acetate: In Vitro and Rat In Situ Studies. Pharmaceutics 2022, 14, 952. https://doi.org/10.3390/pharmaceutics14050952
Braeckmans M, Augustijns P, Mols R, Servais C, Brouwers J. Investigating the Mechanisms behind the Positive Food Effect of Abiraterone Acetate: In Vitro and Rat In Situ Studies. Pharmaceutics. 2022; 14(5):952. https://doi.org/10.3390/pharmaceutics14050952
Chicago/Turabian StyleBraeckmans, Marlies, Patrick Augustijns, Raf Mols, Cécile Servais, and Joachim Brouwers. 2022. "Investigating the Mechanisms behind the Positive Food Effect of Abiraterone Acetate: In Vitro and Rat In Situ Studies" Pharmaceutics 14, no. 5: 952. https://doi.org/10.3390/pharmaceutics14050952