
Nanobiotechnology with Therapeutically Relevant Macromolecules from Animal Venoms: Venoms, Toxins, and Antimicrobial Peptides

 ,
,  , ,
, ,
Abstract
:
1. Introduction
2. Nanosystems and Nanocarriers
2.1. Nanoliposomes
2.2. Silica Nanoparticles (SiNPs)
2.3. Metallic Nanoparticles
2.4. Polymeric Nanoparticles
3. Nanobioconjugations
4. Prospects and Challenges
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Chen, N.; Xu, S.; Zhang, Y.; Wang, F. Animal protein toxins: Origins and therapeutic applications. Biophys. Rep. 2018, 4, 233–242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Perumal Samy, R.; Stiles, B.G.; Franco, O.L.; Sethi, G.; Lim, L.H.K. Animal venoms as antimicrobial agents. Biochem. Pharmacol. 2017, 134, 127–138. [Google Scholar] [CrossRef] [PubMed]
- Roque-Borda, C.A.; Bento, P.; da Silva, P.B.; Rodrigues, M.C.; Azevedo, R.B.; Di Filippo, L.; Duarte, J.L.; Chorilli, M.; Festozo Vicente, E.; Pavan, F.R. Challenge in the Discovery of New Drugs: Antimicrobial Peptides against WHO-List of Critical and High-Priority Bacteria. Pharmaceutics 2021, 13, 773. [Google Scholar] [CrossRef]
- Andrei, S.; Droc, G.; Stefan, G. FDA approved antibacterial drugs: 2018–2019. Discoveries 2019, 7, e102. [Google Scholar] [CrossRef]
- Fry, B.G.; Scheib, H.; van der Weerd, L.; Young, B.; McNaughtan, J.; Ryan Ramjan, S.F.; Vidal, N.; Poelmann, R.E.; Norman, J.A. Evolution of an arsenal: Structural and functional diversification of the venom system in the advanced snakes (Caenophidia). Mol. Cell. Proteom. 2008, 7, 215–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvete, J.J.; Borges, A.; Segura, Á.; Flores-Díaz, M.; Alape-Girón, A.; Gutiérrez, J.M.; Diez, N.; De Sousa, L.; Kiriakos, D.; Sánchez, E.; et al. Snake venomics and antivenomics of Bothrops colombiensis, a medically important pitviper of the Bothrops atrox-asper complex endemic to Venezuela: Contributing to its taxonomy and snakebite management. J. Proteom. 2009, 72, 227–240. [Google Scholar] [CrossRef]
- Lomonte, B.; Angulo, Y.; Moreno, E. Synthetic Peptides Derived from the C-Terminal Region of Lys49 Phospholipase A2 Homologues from Viperidae Snake Venoms: Biomimetic Activities and Potential Applications. Curr. Pharm. Des. 2010, 16, 3224–3230. [Google Scholar] [CrossRef]
- De Bordon, K.C.F.; Cologna, C.T.; Fornari-Baldo, E.C.; Pinheiro-Júnior, E.L.; Cerni, F.A.; Amorim, F.G.; Anjolette, F.A.P.; Cordeiro, F.A.; Wiezel, G.A.; Cardoso, I.A.; et al. From Animal Poisons and Venoms to Medicines: Achievements, Challenges and Perspectives in Drug Discovery. Front. Pharmacol. 2020, 11, 1132. [Google Scholar] [CrossRef]
- Wang, K.R.; Zhang, B.Z.; Zhang, W.; Yan, J.X.; Li, J.; Wang, R. Antitumor effects, cell selectivity and structure–activity relationship of a novel antimicrobial peptide polybia-MPI. Peptides 2008, 29, 963–968. [Google Scholar] [CrossRef]
- Yamane, E.S.; Bizerra, F.C.; Oliveira, E.B.; Moreira, J.T.; Rajabi, M.; Nunes, G.L.C.; De Souza, A.O.; Da Silva, I.D.C.G.; Yamane, T.; Karpel, R.L.; et al. Unraveling the antifungal activity of a South American rattlesnake toxin crotamine. Biochimie 2013, 95, 231–240. [Google Scholar] [CrossRef] [Green Version]
- Sartim, M.A.; Menaldo, D.L.; Sampaio, S.V. Immunotherapeutic potential of Crotoxin: Anti-inflammatory and immunosuppressive properties. J. Venom. Anim. Toxins Incl. Trop. Dis. 2018, 24, 39. [Google Scholar] [CrossRef] [Green Version]
- Roque-Borda, C.A.; Silva, H.R.L.; Crusca Junior, E.; Serafim, J.A.; Meneguin, A.B.; Chorilli, M.; Macedo, W.C.; Teixeira, S.R.; Guastalli, E.A.L.; Soares, N.M.; et al. Alginate-based microparticles coated with HPMCP/AS cellulose-derivatives enable the Ctx(Ile21)-Ha antimicrobial peptide application as a feed additive. Int. J. Biol. Macromol. 2021, 183, 1236–1247. [Google Scholar] [CrossRef] [PubMed]
- Severino, P.; da Silva, C.F.; Andrade, L.N.; de Lima Oliveira, D.; Campos, J.; Souto, E.B. Alginate Nanoparticles for Drug Delivery and Targeting. Curr. Pharm. Des. 2019, 25, 1312–1334. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Shi, L.; Su, L.; Van der Mei, H.C.; Jutte, P.C.; Ren, Y.; Busscher, H.J. Nanotechnology-based antimicrobials and delivery systems for biofilm-infection control. Chem. Soc. Rev. 2019, 48, 428–446. [Google Scholar] [CrossRef]
- Li, R.; Lyu, Y.; Luo, S.; Wang, H.; Zheng, X.; Li, L.; Ao, N.; Zha, Z. Fabrication of a multi-level drug release platform with liposomes, chitooligosaccharides, phospholipids and injectable chitosan hydrogel to enhance anti-tumor effectiveness. Carbohydr. Polym. 2021, 269, 118322. [Google Scholar] [CrossRef] [PubMed]
- Walsh, G. Pharmaceutical Biotechnology: Concepts and Applications; John Wiley & Sons: Hoboken, NJ, USA, 2013. [Google Scholar]
- Reddy, M.; Murthy, K.; Srilakshmi, A.; Rao, K.S.; Pullaiah, T. Phytosynthesis of eco-friendly silver nanoparticles and biological applications –A novel concept in nanobiotechnology. Afr. J. Biotechnol. 2015, 14, 222–247. [Google Scholar] [CrossRef]
- Tasima, L.J.; Serino-Silva, C.; Hatakeyama, D.M.; Nishiduka, E.S.; Tashima, A.K.; Sant’Anna, S.S.; Grego, K.F.; de Morais-Zani, K.; Tanaka-Azevedo, A.M. Crotamine in Crotalus durissus: Distribution according to subspecies and geographic origin, in captivity or nature. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26, e20190053. [Google Scholar] [CrossRef]
- Misra, S.K.; Ye, M.; Kim, S.; Pan, D. Defined nanoscale chemistry influences delivery of peptido-toxins for cancer therapy. PLoS ONE 2015, 10, e0125908. [Google Scholar] [CrossRef] [Green Version]
- Sant’Anna, M.B.; Giardini, A.C.; Ribeiro, M.A.C.; Lopes, F.S.R.; Teixeira, N.B.; Kimura, L.F.; Bufalo, M.C.; Ribeiro, O.G.; Borrego, A.; Cabrera, W.H.K.; et al. The Crotoxin:SBA-15 Complex Down-Regulates the Incidence and Intensity of Experimental Autoimmune Encephalomyelitis Through Peripheral and Central Actions. Front. Immunol. 2020, 11, 591563. [Google Scholar] [CrossRef]
- Al-Rabia, M.W.; Alhakamy, N.A.; Ahmed, O.A.A.; Eljaaly, K.; Aloafi, A.L.; Mostafa, A.; Asfour, H.Z.; Aldarmahi, A.A.; Darwish, K.M.; Ibrahim, T.S.; et al. Repurposing of sitagliptin-melittin optimized nanoformula against SARS-CoV-2: Antiviral screening and molecular docking studies. Pharmaceutics 2021, 13, 307. [Google Scholar] [CrossRef]
- Santos-Filho, N.A.; Lorenzon, E.N.; Ramos, M.A.S.; Santos, C.T.; Piccoli, J.P.; Bauab, T.M.; Fusco-Almeida, A.M.; Cilli, E.M. Synthesis and characterization of an antibacterial and non-toxic dimeric peptide derived from the C-terminal region of Bothropstoxin-I. Toxicon 2015, 103, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Swenson, S.D.; Silva-Hirschberg, C.; Markland, F.S. Methods for Evaluation of a Snake Venom-Derived Disintegrin in Animal Models of Human Cancer. Methods Mol. Biol. 2020, 2068, 185–204. [Google Scholar] [CrossRef] [PubMed]
- Minea, R.O.; Helchowski, C.M.; Zidovetzki, S.J.; Costa, F.K.; Swenson, S.D.; Markland, F.S., Jr. Vicrostatin–An Anti-Invasive Multi-Integrin Targeting Chimeric Disintegrin with Tumor Anti-Angiogenic and Pro-Apoptotic Activities. PLoS ONE 2010, 5, e10929. [Google Scholar] [CrossRef] [PubMed]
- Mao, J.; Liu, S.; Ai, M.; Wang, Z.; Wang, D.; Li, X.; Hu, K.; Gao, X.; Yang, Y. A novel melittin nano-liposome exerted excellent anti-hepatocellular carcinoma efficacy with better biological safety. J. Hematol. Oncol. 2017, 10, 4–7. [Google Scholar] [CrossRef] [Green Version]
- Yu, X.; Chen, L.; Liu, J.; Dai, B.; Xu, G.; Shen, G.; Luo, Q.; Zhang, Z. Immune modulation of liver sinusoidal endothelial cells by melittin nanoparticles suppresses liver metastasis. Nat. Commun. 2019, 10, 574. [Google Scholar] [CrossRef]
- Yu, X.; Dai, Y.; Zhao, Y.; Qi, S.; Liu, L.; Lu, L.; Luo, Q.; Zhang, Z. Melittin-lipid nanoparticles target to lymph nodes and elicit a systemic anti-tumor immune response. Nat. Commun. 2020, 11, 1110. [Google Scholar] [CrossRef] [Green Version]
- Cantor, S.; Vargas, L.; Rojas, A.O.; Yarce, C.; Salamanca, C.; Oñate-Garzón, J. Evaluation of the Antimicrobial Activity of Cationic Peptides Loaded in Surface-Modified Nanoliposomes against Foodborne Bacteria. Int. J. Mol. Sci. 2019, 20, 680. [Google Scholar] [CrossRef] [Green Version]
- Chen, C.; Zheng, H.; Xu, J.; Shi, X.; Li, F.; Wang, X. Sustained-release study on Exenatide loaded into mesoporous silica nanoparticles: In vitro characterization and in vivo evaluation. DARU J. Pharm. Sci. 2017, 25, 20. [Google Scholar] [CrossRef]
- Ferreira Junior, R.S.; Anderlini, R.P.; Pimenta, D.C.; De Oliveira Orsi, R.; Barraviera, B.; Sant’Anna, O.A. New Nanostructured Silica Adjuvant (SBA-15) Employed to Produce Antivenom in Young Sheep Using Crotalus durissus terrificus and Apis mellifera Venoms Detoxified by Cobalt-60. J. Toxicol. Environ. Health Part A 2010, 73, 926–933. [Google Scholar] [CrossRef]
- Baudou, F.G.; Fusco, L.; Giorgi, E.; Diaz, E.; Municoy, S.; Desimone, M.F.; Leiva, L.; De Marzi, M.C. Physicochemical and biological characterization of nanovenoms, a new tool formed by silica nanoparticles and Crotalus durissus terrificus venom. Colloids Surf. B Biointerfaces 2020, 193, 111128. [Google Scholar] [CrossRef]
- Mercuri, L.P.; Carvalho, L.V.; Lima, F.A.; Quayle, C.; Fantini, M.C.A.; Tanaka, G.S.; Cabrera, W.H.; Furtado, M.F.D.; Tambourgi, D.V.; do Matos, J.R.; et al. Ordered Mesoporous Silica SBA-15: A New Effective Adjuvant to Induce Antibody Response. Small 2006, 2, 254–256. [Google Scholar] [CrossRef] [PubMed]
- Sayed, D.; Al-Sadoon, M.K.; Badr, G. Silica nanoparticles sensitize human multiple myeloma cells to snake (Walterinnesia aegyptia) venom-induced apoptosis and growth arrest. Oxid. Med. Cell. Longev. 2012, 2012, 386286. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Badr, G.; Al-Sadoon, M.K.; Rabah, D.M.; Sayed, D. Snake (Walterinnesia aegyptia) venom-loaded silica nanoparticles induce apoptosis and growth arrest in human prostate cancer cells. Apoptosis 2013, 18, 300–314. [Google Scholar] [CrossRef]
- Picolo, G. Crotoxin induces analgesic and immunomodulatory effects on chronic pain models that are potentiated by nanostructured silica SBA-15. Toxicon 2020, 177, S20. [Google Scholar] [CrossRef]
- Sant’Anna, M.B.; Lopes, F.S.R.; Kimura, L.F.; Giardini, A.C.; Sant’Anna, O.A.; Picolo, G. Crotoxin conjugated to SBA-15 nanostructured mesoporous silica induces long-last analgesic effect in the neuropathic pain model in mice. Toxins 2019, 11, 679. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Zhang, Y.; Shan, Y.; Wang, J.; Liu, F.; Liu, H.; Xing, G.; Lei, J.; Zhou, J. A pH-dependent Antibacterial Peptide Release Nano-system Blocks Tumor Growth in vivo without Toxicity. Sci. Rep. 2017, 7, 11242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karpel, R.L.; da Silva Liberato, M.; Campeiro, J.D.; Bergeon, L.; Szychowski, B.; Butler, A.; Marino, G.; Cusic, J.F.; de Oliveira, L.C.G.; Oliveira, E.B.; et al. Design and characterization of crotamine-functionalized gold nanoparticles. Colloids Surf. B Biointerfaces 2018, 163, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, S.; Dasgupta, S.C.; Dasgupta, A.K.; Gomes, A.; Gomes, A. Gold Nanoparticles (AuNPs) Conjugated with Andrographolide Ameliorated Viper (Daboia russellii russellii) Venom-Induced Toxicities in Animal Model. J. Nanosci. Nanotechnol. 2020, 20, 3404–3414. [Google Scholar] [CrossRef]
- Oliveira, I.C.F.; de Paula, M.O.; Lastra, H.C.B.; de Alves, B.B.; Moreno, D.A.N.; Yoshida, E.H.; Amaral Filho, J.; Cogo, J.C.; Varanda, E.A.; Rai, M.; et al. Activity of silver nanoparticles on prokaryotic cells and Bothrops jararacussu snake venom. Drug Chem. Toxicol. 2019, 42, 60–64. [Google Scholar] [CrossRef]
- Jalaei, J.; Layeghi-Ghalehsoukhteh, S.; Hosseini, A.; Fazeli, M. Antibacterial effects of gold nanoparticles functionalized with the extracted peptide from Vespa orientalis wasp venom. J. Pept. Sci. 2018, 24, e3124. [Google Scholar] [CrossRef]
- Shi, Y.; Sun, X.; Zhang, L.; Sun, K.; Li, K.; Li, Y.; Zhang, Q. Fc-modified exenatide-loaded nanoparticles for oral delivery to improve hypoglycemic effects in mice. Sci. Rep. 2018, 8, 726. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamzaoui, A.; Laraba-Djebari, F. Development and evaluation of polymeric nanoparticles as a delivery system for snake envenoming prevention. Biologicals 2021, 70, 44–52. [Google Scholar] [CrossRef] [PubMed]
- Zhao, J.; Han, F.; Zhang, W.; Yang, Y.; You, D.; Li, L. Toward improved wound dressings: Effects of polydopamine-decorated poly(lactic-co-glycolic acid) electrospinning incorporating basic fibroblast growth factor and ponericin G1. RSC Adv. 2019, 9, 33038–33051. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, S.; Mohamed, M.S.; Mizuki, T.; Maekawa, T.; Kumar, D.S. Chlorotoxin modified morusin–PLGA nanoparticles for targeted glioblastoma therapy. J. Mater. Chem. B 2019, 7, 5896–5919. [Google Scholar] [CrossRef]
- Lee, J.; Kim, Y.M.; Kim, J.H.; Cho, C.W.; Jeon, J.W.; Park, J.K.; Lee, S.H.; Jung, B.G.; Lee, B.J. Nasal delivery of chitosan/alginate nanoparticle encapsulated bee (Apis mellifera) venom promotes antibody production and viral clearance during porcine reproductive and respiratory syndrome virus infection by modulating T cell related responses. Vet. Immunol. Immunopathol. 2018, 200, 40–51. [Google Scholar] [CrossRef]
- El-Dek, S.; Hassan, M.; Mohamed, A.; Abdel Wahab, A.W. Apoptotic and necrotic effects of chitosan nanoparticles loaded with the honeybee, Apis mellifera venom on different cancer cell lines. J. Egypt. Soc. Parasitol. 2019, 49, 115–122. [Google Scholar] [CrossRef]
- Alalawy, A.I.; El Rabey, H.A.; Almutairi, F.M.; Tayel, A.A.; Al-Duais, M.A.; Zidan, N.S.; Sakran, M.I. Effectual Anticancer Potentiality of Loaded Bee Venom onto Fungal Chitosan Nanoparticles. Int. J. Polym. Sci. 2020, 2020, 2785304. [Google Scholar] [CrossRef]
- Nait Mohamed, F.A.; Nouri, A.; Laraba-Djebari, F. Reactogenicity and safety assessment of an attenuated nanovaccine against scorpion envenomation: Preclinical study. Vaccine 2017, 35, 6657–6663. [Google Scholar] [CrossRef]
- Nait Mohamed, F.A.; Laraba-Djebari, F. Development and characterization of a new carrier for vaccine delivery based on calcium-alginate nanoparticles: Safe immunoprotective approach against scorpion envenoming. Vaccine 2016, 34, 2692–2699. [Google Scholar] [CrossRef]
- Rebbouh, F.; Martin-Eauclaire, M.F.; Laraba-Djebari, F. Chitosan nanoparticles as a delivery platform for neurotoxin II from Androctonus australis hector scorpion venom: Assessment of toxicity and immunogenicity. Acta Trop. 2020, 205, 105353. [Google Scholar] [CrossRef]
- Piras, A.M.; Maisetta, G.; Sandreschi, S.; Gazzarri, M.; Bartoli, C.; Grassi, L.; Esin, S.; Chiellini, F.; Batoni, G. Chitosan nanoparticles loaded with the antimicrobial peptide temporin B exert a long-term antibacterial activity in vitro against clinical isolates of Staphylococcus epidermidis. Front. Microbiol. 2015, 6, 372. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Torres-Rêgo, M.; Gláucia-Silva, F.; Rocha Soares, K.S.; de Souza, L.B.F.C.; Damasceno, I.Z.; dos Santos-Silva, E.; Lacerda, A.F.; Chaves, G.M.; da Silva-Júnior, A.A.; de Fernandes-Pedrosa, M.F. Biodegradable cross-linked chitosan nanoparticles improve anti-Candida and anti-biofilm activity of TistH, a peptide identified in the venom gland of the Tityus stigmurus scorpion. Mater. Sci. Eng. C 2019, 103, 109830. [Google Scholar] [CrossRef] [PubMed]
- Da Silva, F.G.; Rocha Soares, K.S.; Ribeiro Bastos De França, A.C.; Do Rêgo, M.T.; Ribeiro Bastos De França, A.C.; Da Silva, D.P.; De Sousa Ferreira, S.; Dos Santos Silva, A.M.; Da Silva Júnior, A.A.; De Freitas Fernandes Pedrosa, M. Serum production against Crotalus durissus cascavella snake venom using a biotechnological approach as immunoadjuvant. Toxicon 2019, 168, S38. [Google Scholar] [CrossRef]
- Mohammadpourdounighi, N.; Behfar, A.; Ezabadi, A.; Zolfagharian, H.; Heydari, M. Preparation of chitosan nanoparticles containing Naja naja oxiana snake venom. Nanomed. Nanotechnol. Biol. Med. 2010, 6, 137–143. [Google Scholar] [CrossRef]
- Nakamoto, M.; Zhao, D.; Benice, O.R.; Lee, S.H.; Shea, K.J. Abiotic Mimic of Endogenous Tissue Inhibitors of Metalloproteinases: Engineering Synthetic Polymer Nanoparticles for Use as a Broad-Spectrum Metalloproteinase Inhibitor. J. Am. Chem. Soc. 2020, 142, 2338–2345. [Google Scholar] [CrossRef]
- Attarde, S.S.; Pandit, S.V. Anticancer potential of nanogold conjugated toxin GNP-NN-32 from Naja naja venom. J. Venom. Anim. Toxins Incl. Trop. Dis. 2020, 26. [Google Scholar] [CrossRef]
- Tan, H.; Huang, Y.; Xu, J.; Chen, B.; Zhang, P.; Ye, Z.; Liang, S.; Xiao, L.; Liu, Z. Spider toxin peptide lycosin-I functionalized gold nanoparticles for in vivo tumor targeting and therapy. Theranostics 2017, 7, 3168–3178. [Google Scholar] [CrossRef] [Green Version]
- Khamehchian, S.; Nikkhah, M.; Madani, R.; Hosseinkhani, S. Enhanced and selective permeability of gold nanoparticles functionalized with cell penetrating peptide derived from maurocalcine animal toxin. J. Biomed. Mater. Res. Part A 2016, 104, 2693–2700. [Google Scholar] [CrossRef]
- Di Filippo, L.D.; Duarte, J.; Fonseca-Santos, B.; Tavares Júnior, A.G.; Araújo, V.; Roque Borda, C.A.; Vicente, E.F.; Chorilli, M. Mucoadhesive Nanosystems for Nose-to-Brain Drug Delivery in the Treatment of Central Nervous System Diseases. Curr. Med. Chem. 2021, 28, 3079–3110. [Google Scholar] [CrossRef]
- Semenova, M.; Antipova, A.; Martirosova, E.; Zelikina, D.; Palmina, N.; Chebotarev, S. Essential contributions of food hydrocolloids and phospholipid liposomes to the formation of carriers for controlled delivery of biologically active substances via the gastrointestinal tract. Food Hydrocoll. 2021, 120, 106890. [Google Scholar] [CrossRef]
- da Silva Malheiros, P.; Daroit, D.J.; Brandelli, A. Food applications of liposome-encapsulated antimicrobial peptides. Trends Food Sci. Technol. 2010, 21, 284–292. [Google Scholar] [CrossRef]
- Jesorka, A.; Orwar, O. Liposomes: Technologies and analytical applications. Annu. Rev. Anal. Chem. 2008, 1, 801–832. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, M.L.; dos Santos, J.; Silveira, N.P.; Brandelli, A. Phospholipid nanovesicles containing a bacteriocin-like substance for control of Listeria monocytogenes. Innov. Food Sci. Emerg. Technol. 2008, 9, 49–53. [Google Scholar] [CrossRef]
- Colas, J.C.; Shi, W.; Rao, V.S.N.M.; Omri, A.; Mozafari, M.R.; Singh, H. Microscopical investigations of nisin-loaded nanoliposomes prepared by Mozafari method and their bacterial targeting. Micron 2007, 38, 841–847. [Google Scholar] [CrossRef]
- Taylor, T.M.; Bruce, B.D.; Weiss, J.; Davidson, P.M. Listeria monocytogenes and Escherichia coli O157:H7 inhibition in vitro by liposome-encapsulated nisin and ethylene diaminetetraacetic acid. J. Food Saf. 2008, 28, 183–197. [Google Scholar] [CrossRef]
- Laridi, R.; Kheadr, E.E.; Benech, R.O.; Vuillemard, J.C.; Lacroix, C.; Fliss, I. Liposome encapsulated nisin Z: Optimization, stability and release during milk fermentation. Int. Dairy J. 2003, 13, 325–336. [Google Scholar] [CrossRef]
- Were, L.M.; Bruce, B.; Davidson, P.M.; Weiss, J. Encapsulation of nisin and lysozyme in liposomes enhances efficacy against Listeria monocytogenes. J. Food Prot. 2004, 67, 922–927. [Google Scholar] [CrossRef]
- Gan, Z.R.; Gould, R.J.; Jacobs, J.W.; Friedman, P.A.; Polokoff, M.A. Echistatin. A potent platelet aggregation inhibitor from the venom of the viper, Echis carinatus. J. Biol. Chem. 1988, 263, 19827–19832. [Google Scholar] [CrossRef]
- Trikha, M.; De Clerck, Y.A.; Markland, F.S. Contortrostatin, a Snake Venom Disintegrin, Inhibits β1 Integrin-mediated Human Metastatic Melanoma Cell Adhesion and Blocks Experimental Metastasis. Cancer Res. 1994, 54, 4993–4998. [Google Scholar]
- Schönthal, A.H.; Swenson, S.D.; Chen, T.C.; Markland, F.S. Preclinical studies of a novel snake venom-derived recombinant disintegrin with antitumor activity: A review. Biochem. Pharmacol. 2020, 181, 114149. [Google Scholar] [CrossRef]
- Al-Asmari, A.K.; Ullah, Z.; Al Balowi, A.; Islam, M. In vitro determination of the efficacy of scorpion venoms as anti-cancer agents against colorectal cancer cells: A nano-liposomal delivery approach. Int. J. Nanomed. 2017, 12, 559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zimmermann, R.; Mollay, C. Import of honeybee prepromelittin into the endoplasmic reticulum. Requirements for membrane insertion, processing, and sequestration. J. Biol. Chem. 1986, 261, 12889–12895. [Google Scholar] [CrossRef]
- Müller, G.; Zimmermann, R. Import of honeybee prepromelittin into the endoplasmic reticulum: Structural basis for independence of SRP and docking protein. EMBO J. 1987, 6, 2099–2107. [Google Scholar] [CrossRef] [PubMed]
- Ladokhin, A.S.; White, S.H. Folding of amphipathic α-helices on membranes: Energetics of helix formation by melittin. J. Mol. Biol. 1999, 285, 1363–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guha, S.; Ferrie, R.P.; Ghimire, J.; Ventura, C.R.; Wu, E.; Sun, L.; Kim, S.Y.; Wiedman, G.R.; Hristova, K.; Wimley, W.C. Applications and evolution of melittin, the quintessential membrane active peptide. Biochem. Pharmacol. 2021, 193, 114769. [Google Scholar] [CrossRef] [PubMed]
- Dempsey, C.E.; Bazzo, R.; Harvey, T.S.; Syperek, I.; Boheim, G.; Campbell, I.D. Contribution of proline-14 to the structure and actions of melittin. FEBS Lett. 1991, 281, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Bazzo, R.; Tappin, M.J.; Pastore, A.; Harvey, T.S.; Carver, J.A.; Campbell, I.D. The structure of melittin. A 1H-NMR study in methanol. Eur. J. Biochem. 1988, 173, 139–146. [Google Scholar] [CrossRef]
- Terwilliger, T.C.; Eisenberg, D. The structure of melittin. II. Interpretation of the structure. J. Biol. Chem. 1982, 257, 6016–6022. [Google Scholar] [CrossRef]
- Rex, S. A Pro→Ala substitution in melittin affects self-association, membrane binding and pore-formation kinetics due to changes in structural and electrostatic properties. Biophys. Chem. 2000, 85, 209–228. [Google Scholar] [CrossRef]
- Ceremuga, M.; Stela, M.; Janik, E.; Gorniak, L.; Synowiec, E.; Sliwinski, T.; Sitarek, P.; Saluk-Bijak, J.; Bijak, M. Melittin—A Natural Peptide from Bee Venom Which Induces Apoptosis in Human Leukaemia Cells. Biomolecules 2020, 10, 247. [Google Scholar] [CrossRef] [Green Version]
- Socarras, K.; Theophilus, P.; Torres, J.; Gupta, K.; Sapi, E. Antimicrobial Activity of Bee Venom and Melittin against Borrelia burgdorferi. Antibiotics 2017, 6, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Conlon, J.M.; Ahmed, E.; Pal, T.; Sonnevend, A. Potent and rapid bactericidal action of alyteserin-1c and its [E4K] analog against multidrug-resistant strains of Acinetobacter baumannii. Peptides 2010, 31, 1806–1810. [Google Scholar] [CrossRef]
- Subasinghage, A.P.; O’Flynn, D.; Conlon, J.M.; Hewage, C.M. Conformational and membrane interaction studies of the antimicrobial peptide alyteserin-1c and its analogue [E4K]alyteserin-1c. Biochim. Biophys. Acta-Biomembr. 2011, 1808, 1975–1984. [Google Scholar] [CrossRef] [Green Version]
- Aragón-Muriel, A.; Ausili, A.; Sánchez, K.; Rojas A., O.E.; Mosquera, J.L.; Polo-Cerón, D.; Oñate-Garzón, J. Studies on the Interaction of Alyteserin 1c Peptide and Its Cationic Analogue with Model Membranes Imitating Mammalian and Bacterial Membranes. Biomolecules 2019, 9, 527. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selvarajan, V.; Obuobi, S.; Ee, P.L.R. Silica Nanoparticles—A Versatile Tool for the Treatment of Bacterial Infections. Front. Chem. 2020, 8, 602. [Google Scholar] [CrossRef] [PubMed]
- Carvalho, G.C.; Sábio, R.M.; de Cássia Ribeiro, T.; Monteiro, A.S.; Pereira, D.V.; Ribeiro, S.J.L.; Chorilli, M. Highlights in Mesoporous Silica Nanoparticles as a Multifunctional Controlled Drug Delivery Nanoplatform for Infectious Diseases Treatment. Pharm. Res. 2020, 37, 191. [Google Scholar] [CrossRef] [PubMed]
- Sábio, R.M.; Meneguin, A.B.; Ribeiro, T.C.; Silva, R.R.; Chorilli, M. New insights towards mesoporous silica nanoparticles as a technological platform for chemotherapeutic drugs delivery. Int. J. Pharm. 2019, 564, 379–409. [Google Scholar] [CrossRef]
- Schlossbauer, A.; Schaffert, D.; Kecht, J.; Wagner, E.; Bein, T. Click Chemistry for High-Density Biofunctionalization of Mesoporous Silica. J. Am. Chem. Soc. 2008, 130, 12558–12559. [Google Scholar] [CrossRef]
- Carvalho, L.V.; Ruiz, R.D.C.; Scaramuzzi, K.; Marengo, E.B.; Matos, J.R.; Tambourgi, D.V.; Fantini, M.C.A.; Anna, O.A. Immunological parameters related to the adjuvant effect of the ordered mesoporous silica SBA-15. Vaccine 2010, 28, 7829–7836. [Google Scholar] [CrossRef]
- Sábio, R.M.; Meneguin, A.B.; Martins dos Santos, A.; Monteiro, A.S.; Chorilli, M. Exploiting mesoporous silica nanoparticles as versatile drug carriers for several routes of administration. Microporous Mesoporous Mater. 2021, 312, 110774. [Google Scholar] [CrossRef]
- Schlossbauer, A.; Dohmen, C.; Schaffert, D.; Wagner, E.; Bein, T. pH-Responsive Release of Acetal-Linked Melittin from SBA-15 Mesoporous Silica. Angew. Chem. Int. Ed. 2011, 50, 6828–6830. [Google Scholar] [CrossRef] [PubMed]
- Galllwitz, B. Exenatide in type 2 diabetes: Treatment effects in clinical studies and animal study data. Int. J. Clin. Pract. 2006, 60, 1654–1661. [Google Scholar] [CrossRef] [PubMed]
- Alves, P.L.; Abdalla, F.M.F.; Alponti, R.F.; Silveira, P.F. Anti-obesogenic and hypolipidemic effects of a glucagon-like peptide-1 receptor agonist derived from the saliva of the Gila monster. Toxicon 2017, 135, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Kuniyoshi, A.K.; Kodama, R.T.; Cajado-Carvalho, D.; Iwai, L.K.; Kitano, E.; da Silva, C.C.F.; Duzzi, B.; Dias da Silva, W.; Portaro, F.C. Experimental antivenom against serine proteases from the Bothrops jararaca venom obtained in mice, and its comparison with the antibothropic serum from the Butantan Institute. Toxicon 2019, 169, 59–67. [Google Scholar] [CrossRef]
- Badr, G.; Al-Sadoon, M.K.; Abdel-Maksoud, M.A.; Rabah, D.M.; El-Toni, A.M. Cellular and Molecular Mechanisms Underlie the Anti-Tumor Activities Exerted by Walterinnesia aegyptia Venom Combined with Silica Nanoparticles against Multiple Myeloma Cancer Cell Types. PLoS ONE 2012, 7, e51661. [Google Scholar] [CrossRef] [Green Version]
- Badr, G.; Al-Sadoon, M.K.; Rabah, D.M. Therapeutic efficacy and molecular mechanisms of snake (Walterinnesia aegyptia) venom-loaded silica nanoparticles in the treatment of breast cancer- and prostate cancer-bearing experimental mouse models. Free Radic. Biol. Med. 2013, 65, 175–189. [Google Scholar] [CrossRef]
- de Almeida, C.S.; Andrade-Oliveira, V.; Câmara, N.O.S.; Jacysyn, J.F.; Faquim-Mauro, E.L. Crotoxin from Crotalus durissus terrificus Is Able to Down-Modulate the Acute Intestinal Inflammation in Mice. PLoS ONE 2015, 10, e0121427. [Google Scholar] [CrossRef] [Green Version]
- de Andrade, C.M.; Rey, F.M.; Bianchini, F.J.; Sampaio, S.V.; Torqueti, M.R. Crotoxin, a neurotoxin from Crotalus durissus terrificus snake venom, as a potential tool against thrombosis development. Int. J. Biol. Macromol. 2019, 134, 653–659. [Google Scholar] [CrossRef]
- Muller, S.P.; Silva, V.A.O.; Silvestrini, A.V.P.; de Macedo, L.H.; Caetano, G.F.; Reis, R.M.; Mazzi, M.V. Crotoxin from Crotalus durissus terrificus venom: In vitro cytotoxic activity of a heterodimeric phospholipase A2 on human cancer-derived cell lines. Toxicon 2018, 156, 13–22. [Google Scholar] [CrossRef] [Green Version]
- Sant’Anna, M.B.; Giardini, A.C.; Lopes, F.S.; Kimura, L.F.; Chacur, M.; Pagano, R.L.; Zambelli, V.O.; Sant’Anna, O.A.; Picolo, G. Rattlesnake venom components in the control of experimental autoimmune encephalomyelitis: Immunomodulatory properties potentiated by silica SBA-15. Toxicon 2020, 182, S22. [Google Scholar] [CrossRef]
- Vicente, E.F.; Basso, L.G.M.; Crusca Junior, E.; Roque-Borda, C.A.; Costa-Filho, A.J.; Cilli, E.M. Biophysical Studies of TOAC Analogs of the Ctx(Ile21)-Ha Antimicrobial Peptide Using Liposomes. Brazilian J. Phys. 2022, 52, 71. [Google Scholar] [CrossRef]
- Roque-Borda, C.A.; Pereira, L.P.; Guastalli, E.A.L.; Soares, N.M.; Mac-Lean, P.A.B.; Salgado, D.D.; Meneguin, A.B.; Chorilli, M.; Vicente, E.F. HPMCP-Coated Microcapsules Containing the Ctx(Ile21)-Ha Antimicrobial Peptide Reduce the Mortality Rate Caused by Resistant Salmonella Enteritidis in Laying Hens. Antibiotics 2021, 10, 616. [Google Scholar] [CrossRef] [PubMed]
- Lorenzón, E.N.; Cespedes, G.F.; Vicente, E.F.; Nogueira, L.G.; Bauab, T.M.; Castro, M.S.; Cilli, E.M. Effects of Dimerization on the Structure and Biological Activity of Antimicrobial Peptide Ctx-Ha. Antimicrob. Agents Chemother. 2012, 56, 3004–3010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Roque-Borda, C.A.; de Souza Saraiva, M.; Monte, D.F.M.; Rodrigues Alves, L.B.; de Almeida, A.M.; Ferreira, T.S.; de Lima, T.S.; Benevides, V.P.; Cabrera, J.M.; Claire, S.; et al. HPMCAS-Coated Alginate Microparticles Loaded with Ctx(Ile 21 )-Ha as a Promising Antimicrobial Agent against Salmonella Enteritidis in a Chicken Infection Model. ACS Infect. Dis. 2022, 8, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Roque-Borda, C.; de Saraiva, M.M.S.; Macedo, W.C.; Montesinos, J.M.; Meneguin, A.; de Almeida, A.; Teixeira, S.; Chorilli, M.; Pavan, F.; Junior, A.B.; et al. Development of an oral feed additive against intestinal infections based on microparticles’ systems loaded with Ctx(Ile21)-Ha peptide coated with HPMCAS/Chitosan. In Proceedings of the 7th International Electronic Conference on Medicinal Chemistry, Online, 1–30 November 2021; p. 11574. [Google Scholar] [CrossRef]
- Zhang, N.; Zhi, X.; Zhao, J.; Wei, J.; Li, J.; Yang, H. Mesoporous silica induces hippocampal neurons cell autophagy through AMPK/mTOR/P70S6K signaling pathway. Environ. Toxicol. 2020, 35, 176–187. [Google Scholar] [CrossRef]
- Zhao, J.; Bu, D.; Zhang, N.; Tian, D.; Ma, L.; Yang, H. Cytotoxicity of mesoporous silica modified by amino and carboxyl groups on vascular endothelial cells. Environ. Toxicol. 2021, 36, 1422–1433. [Google Scholar] [CrossRef]
- Salem, S.S.; Fouda, A. Green Synthesis of Metallic Nanoparticles and Their Prospective Biotechnological Applications: An Overview. Biol. Trace Elem. Res. 2021, 199, 344–370. [Google Scholar] [CrossRef]
- Goncalves, J.M.; Polson, A. The electrophoretic analysis of snake venoms. Arch. Biochem. 1947, 13, 253–259. [Google Scholar]
- Laure, C.J. The primary structure of crotamine (author’s transl). Hoppe. Seylers. Z. Physiol. Chem. 1975, 356, 213–215. [Google Scholar]
- Beltran, J.R.; Mascarenhas, Y.P.; Craievich, A.F.; Laure, C.J. SAXS study of the snake toxin α-crotamine. Eur. Biophys. J. 1990, 17, 325–329. [Google Scholar] [CrossRef]
- Nicastro, G.; Franzoni, L.; de Chiara, C.; Mancin, A.C.; Giglio, J.R.; Spisni, A. Solution structure of crotamine, a Na+ channel affecting toxin from Crotalus durissus terrificus venom. Eur. J. Biochem. 2003, 270, 1969–1979. [Google Scholar] [CrossRef] [PubMed]
- Fadel, V.; Bettendorff, P.; Herrmann, T.; De Azevedo, W.F.; Oliveira, E.B.; Yamane, T.; Wüthrich, K. Automated NMR structure determination and disulfide bond identification of the myotoxin crotamine from Crotalus durissus terrificus. Toxicon 2005, 46, 759–767. [Google Scholar] [CrossRef] [PubMed]
- Vital Brazil, O.; Fontana, M.D. Toxins as tools in the study of sodium channel distribution in the muscle fibre membrane. Toxicon 1993, 31, 1085–1098. [Google Scholar] [CrossRef]
- Peigneur, S.; Orts, D.J.B.; Prieto Da Silva, A.R.; Oguiura, N.; Boni-Mitake, M.; De Oliveira, E.B.; Zaharenko, A.J.; De Freitas, J.C.; Tytgat, J. Crotamine pharmacology revisited: Novel insights based on the inhibition of K v channels. Mol. Pharmacol. 2012, 82, 90–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hayashi, M.A.F.; Nascimento, F.D.; Kerkis, A.; Oliveira, V.; Oliveira, E.B.; Pereira, A.; Rádis-Baptista, G.; Nader, H.B.; Yamane, T.; Kerkis, I.; et al. Cytotoxic effects of crotamine are mediated through lysosomal membrane permeabilization. Toxicon 2008, 52, 508–517. [Google Scholar] [CrossRef] [PubMed]
- Campeiro, J.D.; Marinovic, M.P.; Carapeto, F.C.; Dal Mas, C.; Monte, G.G.; Carvalho Porta, L.; Nering, M.B.; Oliveira, E.B.; Hayashi, M.A.F. Oral treatment with a rattlesnake native polypeptide crotamine efficiently inhibits the tumor growth with no potential toxicity for the host animal and with suggestive positive effects on animal metabolic profile. Amino Acids 2018, 50, 267–278. [Google Scholar] [CrossRef] [PubMed]
- Macedo, S.R.A.; De Barros, N.B.; Ferreira, A.S.; Moreira-Dill, L.S.; Calderon, L.A.; Soares, A.M.; Nicolete, R. Biodegradable Microparticles Containing Crotamine Isolated from Crotalus durissus terrificus Display Antileishmanial Activity in vitro. Pharmacology 2015, 95, 78–86. [Google Scholar] [CrossRef] [PubMed]
- Passero, L.F.D.; Tomokane, T.Y.; Corbett, C.E.P.; Laurenti, M.D.; Toyama, M.H. Comparative studies of the anti-leishmanial activity of three Crotalus durissus ssp. venoms. Parasitol. Res. 2007, 101, 1365–1371. [Google Scholar] [CrossRef]
- Dal Mas, C.; Moreira, J.T.; Pinto, S.; Monte, G.G.; Nering, M.B.; Oliveira, E.B.; Gazarini, M.L.; Mori, M.A.; Hayashi, M.A.F. Anthelmintic effects of a cationic toxin from a South American rattlesnake venom. Toxicon 2016, 116, 49–55. [Google Scholar] [CrossRef]
- El Chamy Maluf, S.; Dal Mas, C.; Oliveira, E.B.; Melo, P.M.; Carmona, A.K.; Gazarini, M.L.; Hayashi, M.A.F. Inhibition of malaria parasite Plasmodium falciparum development by crotamine, a cell penetrating peptide from the snake venom. Peptides 2016, 78, 11–16. [Google Scholar] [CrossRef]
- Mohammed, M.A.; Syeda, J.T.M.; Wasan, K.M.; Wasan, E.K. An Overview of Chitosan Nanoparticles and Its Application in Non-Parenteral Drug Delivery. Pharmaceutics 2017, 9, 53. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielinska, A.; Carreiró, F.; Oliveira, A.M.; Neves, A.; Pires, B.; Venkatesk, D.N.; Durazzo, A.; Lucarin, M.; Eder, P.; Silva, A.M.; et al. Polymeric Nanoparticles: Production, Characterization, Toxicology and Ecotoxicology. Molecules 2020, 25, 3731–3751. [Google Scholar] [CrossRef] [PubMed]
- De Freitas, L.M.; Calixto, G.M.F.; Chorilli, M.; Giusti, J.S.M.; Bagnato, V.S.; Soukos, N.S.; Amiji, M.M.; Fontana, C.R. Polymeric Nanoparticle-Based Photodynamic Therapy for Chronic Periodontitis in Vivo. Int. J. Mol. Sci. 2016, 17, 769. [Google Scholar] [CrossRef] [PubMed]
- Bellich, B.; D’Agostino, I.; Semeraro, S.; Gamini, A.; Cesàro, A. “The Good, the Bad and the Ugly” of Chitosans. Mar. Drugs 2016, 14, 99. [Google Scholar] [CrossRef] [Green Version]
- Lu, B.; Lv, X.; Le, Y. Chitosan-Modified PLGA Nanoparticles for Control-Released Drug Delivery. Polymers 2019, 11, 304. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Deng, C.; Meng, F.; Zhang, J.; Sun, H.; Zhong, Z. Hyaluronic acid coated PLGA nanoparticulate docetaxel effectively targets and suppresses orthotopic human lung cancer. J. Control. Release 2017, 259, 76–82. [Google Scholar] [CrossRef]
- El-Hammadi, M.M.; Delgado, Á.V.; Melguizo, C.; Prados, J.C.; Arias, J.L. Folic acid-decorated and PEGylated PLGA nanoparticles for improving the antitumour activity of 5-fluorouracil. Int. J. Pharm. 2017, 516, 61–70. [Google Scholar] [CrossRef]
- Orivel, J.; Redeker, V.; Le Caer, J.P.; Krier, F.; Revol-Junelles, A.M.; Longeon, A.; Chaffotte, A.; Dejean, A.; Rossier, J. Ponericins, New Antibacterial and Insecticidal Peptides from the Venom of the Ant Pachycondyla goeldii. J. Biol. Chem. 2001, 276, 17823–17829. [Google Scholar] [CrossRef] [Green Version]
- Shukla, A.; Fleming, K.E.; Chuang, H.F.; Chau, T.M.; Loose, C.R.; Stephanopoulos, G.N.; Hammond, P.T. Controlling the release of peptide antimicrobial agents from surfaces. Biomaterials 2010, 31, 2348–2357. [Google Scholar] [CrossRef]
- Zhang, H.; Jiao, J.; Jin, H. Degradable poly-l-lysine-modified PLGA cell microcarriers with excellent antibacterial and osteogenic activity. Int. Mater. Rev. 2021, 66, 77–113. [Google Scholar] [CrossRef] [Green Version]
- DeBin, J.A.; Maggio, J.E.; Strichartz, G.R. Purification and characterization of chlorotoxin, a chloride channel ligand from the venom of the scorpion. Am. J. Physiol.-Cell Physiol. 1993, 264, C361–C369. [Google Scholar] [CrossRef] [PubMed]
- Sawaya, R.E.; Yamamoto, M.; Gokaslan, Z.L.; Wang, S.W.; Mohanam, S.; Fuller, G.N.; McCutcheon, I.E.; Stetler-Stevenson, W.G.; Nicolson, G.L.; Rao, J.S. Expression and localization of 72 kDa type IV collagenase (MMP-2) in human malignant gliomas in vivo. Clin. Exp. Metastasis 1996, 14, 35–42. [Google Scholar] [CrossRef] [PubMed]
- Deshane, J.; Garner, C.C.; Sontheimer, H. Chlorotoxin Inhibits Glioma Cell Invasion via Matrix Metalloproteinase-2. J. Biol. Chem. 2003, 278, 4135–4144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zahoor, A.; Sharma, S.; Khuller, G.K. Inhalable alginate nanoparticles as antitubercular drug carriers against experimental tuberculosis. Int. J. Antimicrob. Agents 2005, 26, 298–303. [Google Scholar] [CrossRef] [PubMed]
- de Vos, P.; Faas, M.M.; Spasojevic, M.; Sikkema, J. Encapsulation for preservation of functionality and targeted delivery of bioactive food components. Int. Dairy J. 2010, 20, 292–302. [Google Scholar] [CrossRef]
- Khalil, A.; Elesawy, B.H.; Ali, T.M.; Ahmed, O.M. Bee Venom: From Venom to Drug. Molecules 2021, 26, 4941. [Google Scholar] [CrossRef]
- Zhou, J.; Wan, C.; Cheng, J.; Huang, H.; Lovell, J.F.; Jin, H. Delivery Strategies for Melittin-Based Cancer Therapy. ACS Appl. Mater. Interfaces 2021, 13, 17158–17173. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Canofeni, S.; Miele, R.; Mangoni, M.L.; Barra, D. Temporins, Antimicrobial Peptides from the European Red Frog Rana temporaria. Eur. J. Biochem. 1996, 242, 788–792. [Google Scholar] [CrossRef]
- Argiolas, A.; Pisano, J.J. Isolation and characterization of two new peptides, mastoparan C and crabrolin, from the venom of the European hornet, Vespa crabro. J. Biol. Chem. 1984, 259, 10106–10111. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Rinaldi, A.C.; Di Giulio, A.; Mignogna, G.; Bozzi, A.; Barra, D.; Simmaco, M. Structure–function relationships of temporins, small antimicrobialpeptides from amphibian skin. Eur. J. Biochem. 2000, 267, 1447–1454. [Google Scholar] [CrossRef] [Green Version]
- Mangoni, M.L.; Shai, Y. Temporins and their synergism against Gram-negative bacteria and in lipopolysaccharide detoxification. Biochim. Biophys. Acta-Biomembr. 2009, 1788, 1610–1619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mahalka, A.K.; Kinnunen, P.K.J. Binding of amphipathic α-helical antimicrobial peptides to lipid membranes: Lessons from temporins B and L. Biochim. Biophys. Acta-Biomembr. 2009, 1788, 1600–1609. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kakar, A.; Holzknecht, J.; Dubrac, S.; Gelmi, M.L.; Romanelli, A.; Marx, F. New perspectives in the antimicrobial activity of the amphibian temporin b: Peptide analogs are effective inhibitors of Candida albicans growth. J. Fungi 2021, 7, 457. [Google Scholar] [CrossRef]
- Marcocci, M.E.; Amatore, D.; Villa, S.; Casciaro, B.; Aimola, P.; Franci, G.; Grieco, P.; Galdiero, M.; Palamara, A.T.; Mangoni, M.L.; et al. The amphibian antimicrobial peptide temporin b inhibits in vitro herpes simplex virus 1 infection. Antimicrob. Agents Chemother. 2018, 62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribeiro Bastos De França, A.C.; Rocha Soares, K.S.; Da Silva, F.G.; Da Silva, A.D.; Do Rêgo, M.T.; Da Silva, D.P.; De Sousa Ferreira, S.; Furtado, A.A.; Da Silva Júnior, A.A.; De Freitas Fernandes Pedrosa, M. Evaluation of self-assembled Bothrops jararacussu snake venom proteins cross-linked chitosan nanoparticles activity for use as a potential antibacterial. Toxicon 2019, 168, S32–S33. [Google Scholar] [CrossRef]
- Soares, K.; Gláucia-Silva, F.; Daniele-Silva, A.; Torres-Rêgo, M.; Araújo, N.; Menezes, Y.; Damasceno, I.; Tambourgi, D.; da Silva-Júnior, A.; Fernandes-Pedrosa, M. Antivenom Production against Bothrops jararaca and Bothrops erythromelas Snake Venoms Using Cross-Linked Chitosan Nanoparticles as an Immunoadjuvant. Toxins 2018, 10, 158. [Google Scholar] [CrossRef] [Green Version]
- Gláucia-Silva, F.; Torres-Rêgo, M.; Rocha Soares, K.S.; Damasceno, I.Z.; Tambourgi, D.V.; da Silva-Júnior, A.A.; de Freitas Fernandes-Pedrosa, M. A biotechnological approach to immunotherapy: Antivenom against Crotalus durissus cascavella snake venom produced from biodegradable nanoparticles. Int. J. Biol. Macromol. 2018, 120, 1917–1924. [Google Scholar] [CrossRef]
- Attarde, S.S.; Pandit, S.V. In Vivo Toxicity Profile of NN-32 and Nanogold Conjugated GNP-NN-32 from Indian Spectacled Cobra Venom. Curr. Pharm. Biotechnol. 2020, 21, 1479–1488. [Google Scholar] [CrossRef]
- de Pontes, J.T.C.; Toledo Borges, A.B.; Roque-Borda, C.A.; Pavan, F.R. Antimicrobial Peptides as an Alternative for the Eradication of Bacterial Biofilms of Multi-Drug Resistant Bacteria. Pharmaceutics 2022, 14, 642. [Google Scholar] [CrossRef]
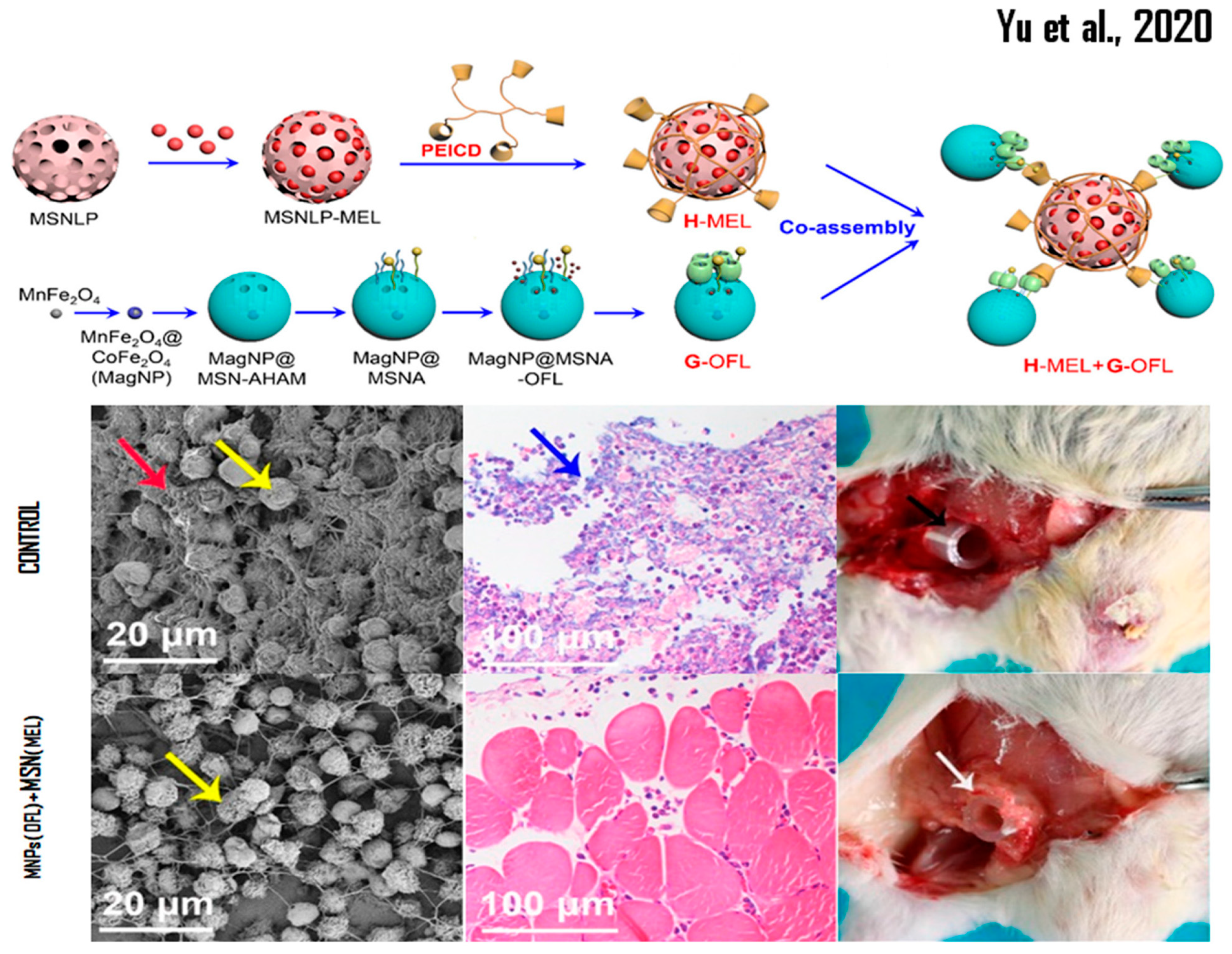
- Yu, Q.; Deng, T.; Lin, F.C.; Zhang, B.; Zink, J.I. Supramolecular Assemblies of Heterogeneous Mesoporous Silica Nanoparticles to Co-deliver Antimicrobial Peptides and Antibiotics for Synergistic Eradication of Pathogenic Biofilms. ACS Nano 2020, 14, 5926–5937. [Google Scholar] [CrossRef]
- Santos-Filho, N.A.; Fernandes, R.S.; Sgardioli, B.F.; Ramos, M.A.S.; Piccoli, J.P.; Camargo, I.L.B.C.; Bauab, T.M.; Cilli, E.M. Antibacterial Activity of the Non-Cytotoxic Peptide (p-BthTX-I)2 and Its Serum Degradation Product against Multidrug-Resistant Bacteria. Mol. A J. Synth. Chem. Nat. Prod. Chem. 2017, 22, 1898. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Baelen, A.-C.; Robin, P.; Kessler, P.; Maïga, A.; Gilles, N.; Servent, D. Structural and Functional Diversity of Animal Toxins Interacting With GPCRs. Front. Mol. Biosci. 2022, 9, 811365. [Google Scholar] [CrossRef] [PubMed]
- Gilles, N.; Servent, D. The European FP7 Venomics Project. Future Med. Chem. 2014, 6, 1611–1612. [Google Scholar] [CrossRef] [Green Version]
- Hunter, H.N.; Jing, W.; Schibli, D.J.; Trinh, T.; Park, I.Y.; Kim, S.C.; Vogel, H.J. The interactions of antimicrobial peptides derived from lysozyme with model membrane systems. Biochim. Biophys. Acta-Biomembr. 2005, 1668, 175–189. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dathe, M.; Nikolenko, H.; Klose, J.; Bienert, M. Cyclization increases the antimicrobial activity and selectivity of arginine- and tryptophan-containing hexapeptides. Biochemistry 2004, 43, 9140–9150. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.; Schibli, D.; Vogel, H. Structural studies and model membrane interactions of two peptides derived from bovine lactoferricin. J. Pept. Sci. 2005, 11, 379–389. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.T.; Chau, J.K.; Perry, N.A.; de Boer, L.; Zaat, S.A.J.; Vogel, H.J. Serum Stabilities of Short Tryptophan- and Arginine-Rich Antimicrobial Peptide Analogs. PLoS ONE 2010, 5, e12684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorenzon, E.N.; Piccoli, J.P.; Santos-Filho, N.A.; Cilli, E.M. Dimerization of Antimicrobial Peptides: A Promising Strategy to Enhance Antimicrobial Peptide Activity. Protein Pept. Lett. 2019, 26, 98. [Google Scholar] [CrossRef]
- Giribaldi, J.; Smith, J.J.; Schroeder, C.I. Recent developments in animal venom peptide nanotherapeutics with improved selectivity for cancer cells. Biotechnol. Adv. 2021, 50, 107769. [Google Scholar] [CrossRef]
- Sadauskas, E.; Wallin, H.; Stoltenberg, M.; Vogel, U.; Doering, P.; Larsen, A.; Danscher, G. Kupffer cells are central in the removal of nanoparticles from the organism. Part. Fibre Toxicol. 2007, 4, 10. [Google Scholar] [CrossRef] [Green Version]
- Poon, W.; Zhang, Y.-N.; Ouyang, B.; Kingston, B.R.; Wu, J.L.Y.; Wilhelm, S.; Chan, W.C.W. Elimination Pathways of Nanoparticles. ACS Nano 2019, 13, 5785–5798. [Google Scholar] [CrossRef] [PubMed]
- Fox, J.; Serrano, S. Approaching the Golden Age of Natural Product Pharmaceuticals from Venom Libraries: An Overview of Toxins and Toxin-Derivatives Currently Involved in Therapeutic or Diagnostic Applications. Curr. Pharm. Des. 2007, 13, 2927–2934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smallwood, T.B.; Clark, R.J. Advances in venom peptide drug discovery: Where are we at and where are we heading? Expert Opin. Drug Discov. 2021, 16, 1163–1173. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
| Toxin | Classification/Source | Nanosystem | Tested Activities | Refs. |
|---|---|---|---|---|
| Vicrostatin | Quimeric peptide Echistatin (Echis carinatus) and contortrostatin (Agkistrodon contortrix contortrix) | Liposomes | Anticancer (mouse breast cancer, human ovarian cancer, glioma, and prostate cancer) | [23,24] |
| Melittin | Peptide from Apis mellifera venom | Liposomes | Anti-hepatocellular carcinoma; Prevention of metastatic lesions | [25,26,27] |
| Alyteserin-1c | Peptide from Alytes obstetricans venom | Polymer-coated liposomes | Antibacterial activity against Listeria monocytogenes | [28] |
| Exenatide | Peptide from Heloderma suspectum | Mesoporous Silica NPs | Glycemic modulation | [29] |
| Detoxified venom | South American rattlesnake (Crotalus durissus terrificus) venom | Mesoporous Silica NPs Silica NPs | Modulation and stimulation of antibodies | [30,31] |
| Detoxified venom | Apis mellifera venom | Mesoporous Silica NPs | Modulation and stimulation of antibodies | [30] |
| Crude venom | Micrurus ibiboboca snake | Mesoporous Silica NPs | Modulation and stimulation of antibodies | [32] |
| Crude venom | Walterinnesia aegyptia venom | Mesoporous Silica NPs | Anti-cancer potential against breast cancer, human multiple myeloma, and prostate cancer cells | [33,34] |
| Crotoxin (CTX) | Beta-neurotoxin (PLA2) from C. durissus terrificus snake venom | Mesoporous Silica NPs | Potential activity against autoimmune, inflammatory diseases and cancer. | [35] |
| Crotoxin (CTX) | Beta-neurotoxin (PLA2) from C. durissus terrificus snake venom | Mesoporous Silica NPs | Potential contraceptive activity | [36] |
| Crotalphine | Peptide from C. durissus terrificus snake venom | Mesoporous Silica NPs | Potential activity against autoimmune and inflammatory diseases | [20] |
| Hylin a1 | Peptide from South American tree frogs’ skin | Mesoporous Silica NPs | Anti-tumor activity and reduced hemolytic activity | [37] |
| Crotamine | Myotoxin from C. durissus terrificus snake venom | Gold NPs (PEG linker) | Anticancer and cellular imaging | [38] |
| Crude venom | Daboia russellii russellii venom | Gold NPs | Prevention of envenomation symptoms | [39] |
| Crude venom | Bothrops jararacussu, Daboia russelii venom and Naja kaouthia venom | TiO2 NPs | Prevention of envenomation symptoms | [40] |
| Peptide INLKAIAALVKKV | Peptide from wasp venom (Vespa orientalis) | Gold NPs | Antibacterial activity | [41] |
| Exenatide | Peptide from Heloderma suspectum | PEG/PLGA NPs | Glycemic modulation | [42] |
| Crude venom | Cerastes cerastes (horned viper) and Vipera lebetina | PLGA NPs | Development of an intranasal vaccine against envenomation | [43] |
| Ponericin-G1 | Peptide from Pachycondyla goeldii ant venom | PDA-PLGA nanofibers | Antimicrobial activity | [44] |
| Chlorotoxin | Peptide from Leiurus quinquestriatus scorpion venom | Morusin-PLGA NPs | Anticancer activity | [45] |
| Crude-lyophilized venom | Apis melifera venom | Alginate/chitosan NPs | Antiviral activity and vaccine adjuvant | [46] |
| Crude-lyophilized venom | Apis melifera venom | Chitosan NPs | Activity against cervical carcinoma, larynx, and breast cancer cells | [47,48] |
| Attenuated venom | Androctonus australis hector scorpion venom | Alginate NPs | Development of a vaccine against envenomation | [49,50] |
| Aah II toxin | Peptide from Androctonus australis hector scorpion venom | Chitosan NP | Development of a vaccine against envenomation | [51] |
| Temporin B | Peptide from the skin secretion of Rana temporaria | Chitosan NPs | Antibacterial activity | [52] |
| Hypotensin | Peptide from Tityus stigmurus scorpion venom | Chitosan NPs | Antibacterial and antifungal activity | [53] |
| Crude-lyophilized venom | Bothrops jararacussu venom | Chitosan NPs | Antibacterial activity against Gram-positive bacterias | [54] |
| Crude-lyophilized venom | Naja naja oxiana, Bothrops jararaca and Bothrops erythromelas | Chitosan NPs | Prevention of envenmation symptoms | [55] |
| Venom proteins | Crotalus durissus cascavella venom | Chitosan NPs | Application as ant-venom | [54] |
| Crude venom | Crotalus atrox, Bitis arietans, Bitis gabonica, Echis ocellatus, and Echis carinatus | Polymeric NPs | Application as anti-venom, deactivation of metalloproteinases | [56] |
| NN-32 | Peptide from the Indian cobra Naja naja venom | Functionalization of gold NPs | Antitumor activity | [57] |
| Lycosin-I | Peptide from Lycosa singorensis spider | Functionalization of gold NPs | Anticancer activity | [58] |
| Maurocalcine | Peptide from Scorpio maurus palmatus | Functionalization of gold NPs | Anticancer activity | [59] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Roque-Borda, C.A.; Gualque, M.W.d.L.; da Fonseca, F.H.; Pavan, F.R.; Santos-Filho, N.A. Nanobiotechnology with Therapeutically Relevant Macromolecules from Animal Venoms: Venoms, Toxins, and Antimicrobial Peptides. Pharmaceutics 2022, 14, 891. https://doi.org/10.3390/pharmaceutics14050891
Roque-Borda CA, Gualque MWdL, da Fonseca FH, Pavan FR, Santos-Filho NA. Nanobiotechnology with Therapeutically Relevant Macromolecules from Animal Venoms: Venoms, Toxins, and Antimicrobial Peptides. Pharmaceutics. 2022; 14(5):891. https://doi.org/10.3390/pharmaceutics14050891
Chicago/Turabian StyleRoque-Borda, Cesar Augusto, Marcos William de Lima Gualque, Fauller Henrique da Fonseca, Fernando Rogério Pavan, and Norival Alves Santos-Filho. 2022. "Nanobiotechnology with Therapeutically Relevant Macromolecules from Animal Venoms: Venoms, Toxins, and Antimicrobial Peptides" Pharmaceutics 14, no. 5: 891. https://doi.org/10.3390/pharmaceutics14050891