
VLA4-Enhanced Allogeneic Endothelial Progenitor Cell-Based Therapy Preserves the Aortic Valve Function in a Mouse Model of Dyslipidemia and Diabetes

 , ,
, ,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal Models
2.2. EPC Sorting
2.3. Transfection of EPCs
- -
- EPCs from dyslipidemic animals, transfected with GFP (CIT-EPC-GFP);
- -
- EPCs from diabetic dyslipidemic animals, transfected with GFP (STZ-EPC-GFP);
- -
- EPCs from dyslipidemic animals, transfected with α4 and β1 integrins (CIT-EPC-VLA4);
- -
- EPCs from diabetic dyslipidemic animals, transfected with α4 and β1 integrins (STZ-EPC-VLA4).
2.4. Animal Groups and Their Treatments
- -
- CIT-EPC-GFP in dyslipidemic animals (n = 4 males and 2 females)
- -
- STZ-EPC-GFP in diabetic dyslipidemic animals (n = 4 males and 2 females)
- -
- CIT-EPC-VLA4 in dyslipidemic animals (n = 4 males and 4 females)
- -
- STZ-EPC-VLA4 in diabetic dyslipidemic animals (n = 4 males and 4 females)
- -
- CIT-EPC-C57 in dyslipidemic animals (n = 4 males and 2 females);
- -
- STZ-EPC-C57 in diabetic dyslipidemic animals (n = 4 males and 2 females).
- -
- CIT-AFL-dyslipidemic animals (n = 2 females);
- -
- STZ-AFL-diabetic dyslipidemic animals (n = 2 females).
2.5. Biochemistry
2.6. Ecocardiography
2.7. EPC Organ Distribution
2.8. Immunohistochemistry
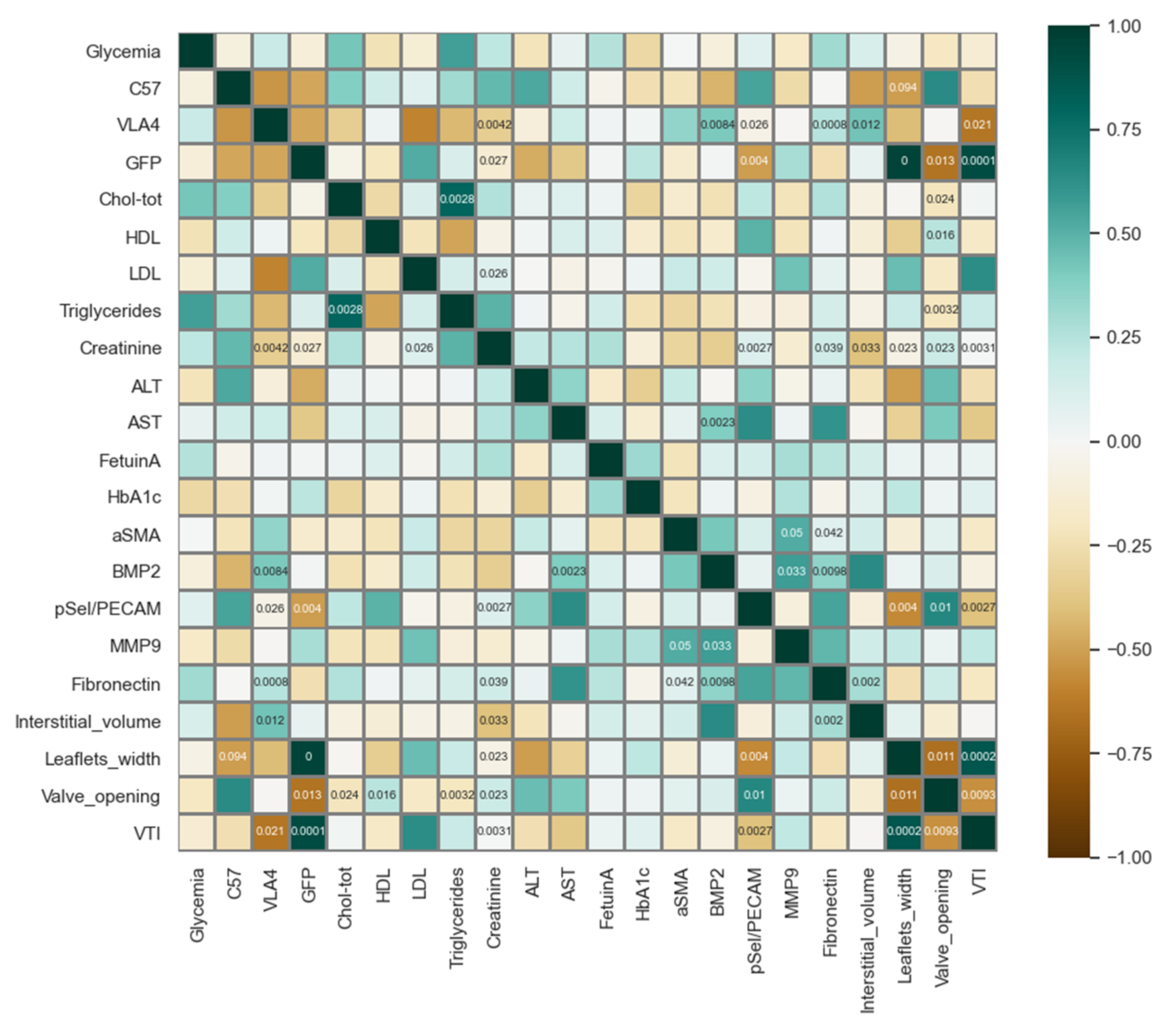
2.9. Processing and Data Analysis
3. Results
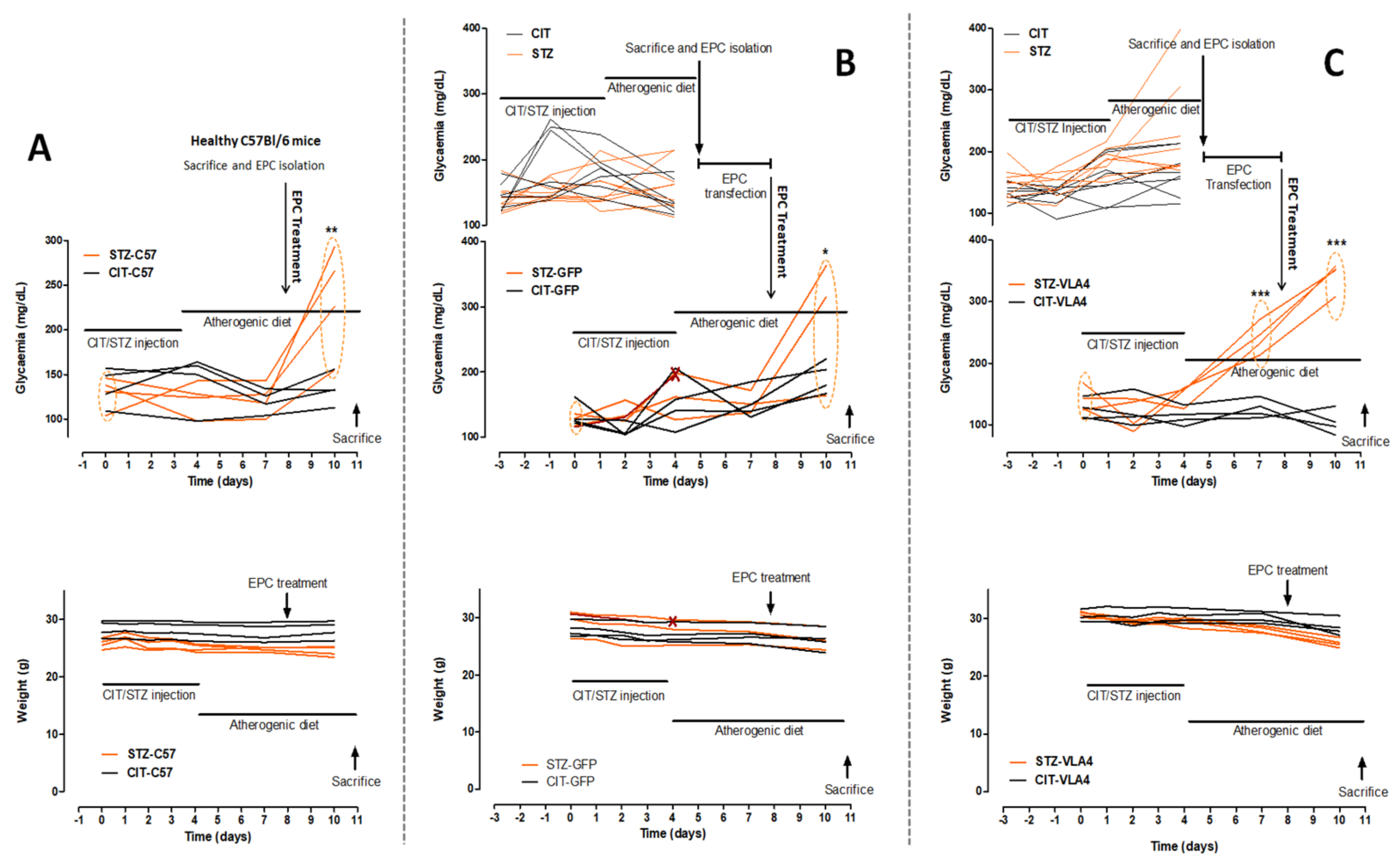
3.1. Generation of the Animal Model

Biochemical Characterization
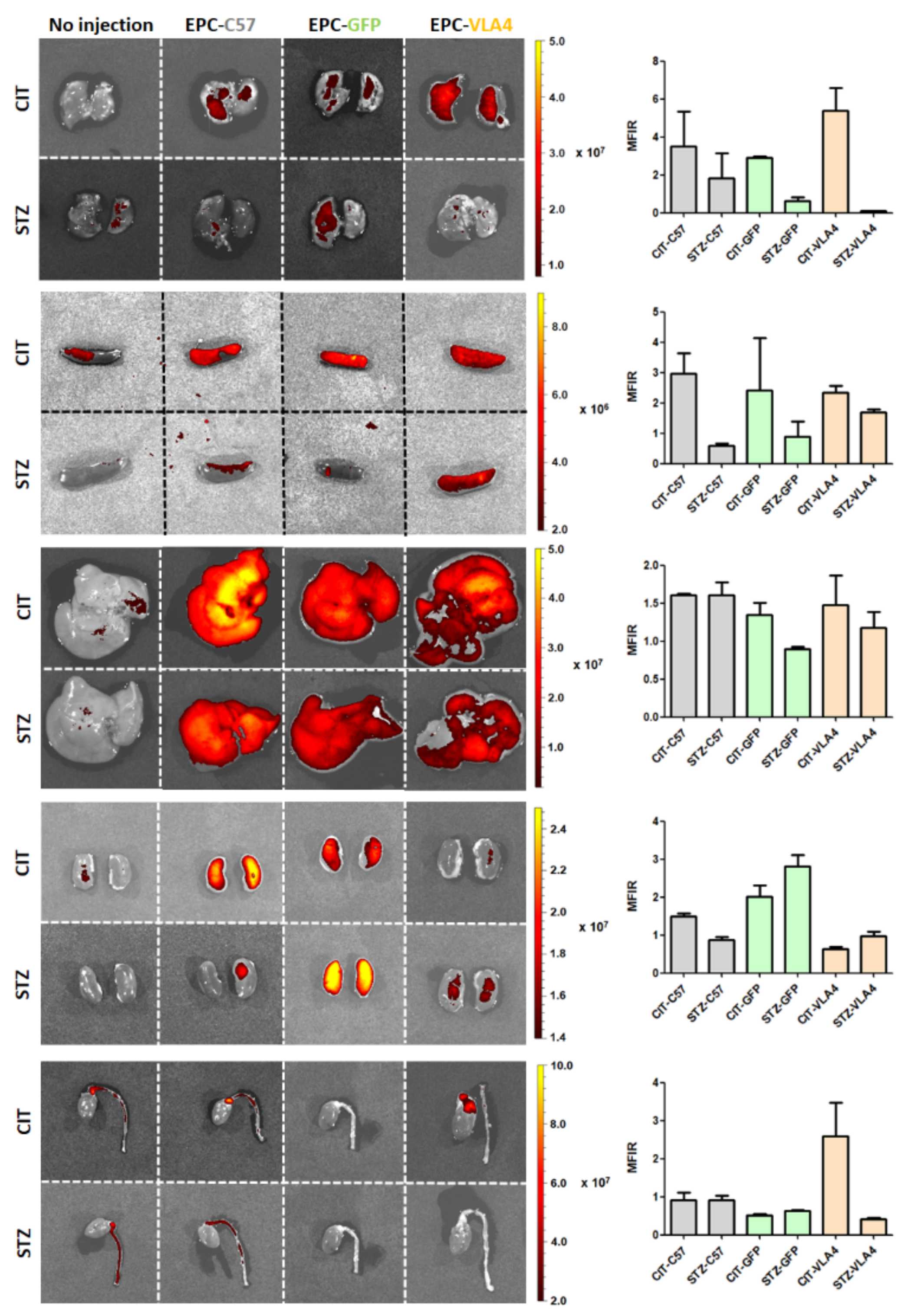
3.2. EPCs Reach Their Target Sites
3.3. VLA4 Overexpressing EPCs Protect the Valve Function as Well as Healthy EPCs
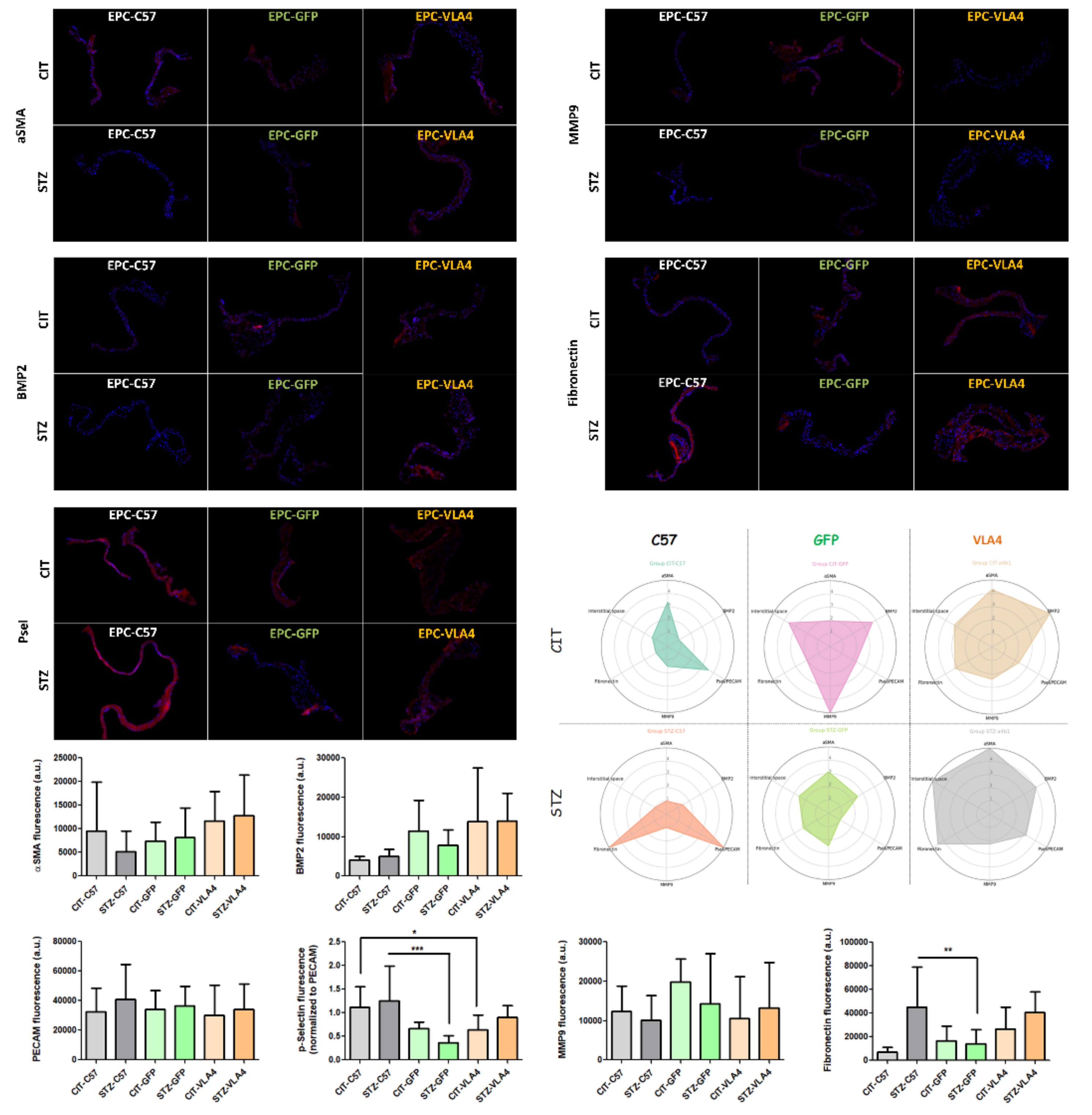
3.4. Inflammation and Remodelling Profiles Assessed by Immunochemistry
4. Discussion
5. Conclusions
6. Patents
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Cote, C.; Pibarot, P.; Despres, J.P.; Mohty, D.; Cartier, A.; Arsenault, B.J.; Couture, C.; Mathieu, P. Association between circulating oxidised low-density lipoprotein and fibrocalcific remodelling of the aortic valve in aortic stenosis. Heart 2008, 94, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Mazzone, A.; Venneri, L.; Berti, S. Aortic valve stenosis and coronary artery disease: Pathophysiological and clinical links. J. Cardiovasc. Med. 2007, 8, 983–989. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Chen, Z.; Ge, J.; Ma, J.; Chang, S.; Fan, B.; Liu, X.; Ge, L. Relationship between aortic valve calcification and the severity of coronary atherosclerotic disease. J. Heart Valve Dis. 2010, 19, 466–470. [Google Scholar] [PubMed]
- Kamalesh, M.; Ng, C.; El Masry, H.; Eckert, G.; Sawada, S. Does diabetes accelerate progression of calcific aortic stenosis? Eur. J. Echocardiogr. 2009, 10, 723–725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yan, A.T.; Koh, M.; Chan, K.K.; Guo, H.; Alter, D.A.; Austin, P.C.; Tu, J.V.; Wijeysundera, H.C.; Ko, D.T. Association between cardiovascular risk factors and aortic atenosis: The CANHEART aortic stenosis study. J. Am. Coll. Cardiol. 2017, 69, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Ferrari, S.; Pesce, M. The complex interplay of inflammation, metabolism, epigenetics, and sex in calcific disease of the aortic valve. Front. Cardiovasc. Med. 2021, 8, 791646. [Google Scholar] [CrossRef] [PubMed]
- Ciortan, L.; Macarie, R.D.; Cecoltan, S.; Vadana, M.; Tucureanu, M.M.; Mihaila, A.C.; Droc, I.; Butoi, E.; Manduteanu, I. Chronic high glucose concentration induces inflammatory and remodeling changes in valvular endothelial cells and valvular interstitial cells in a gelatin methacrylate 3D model of the human aortic valve. Polymers 2020, 12, 2786. [Google Scholar] [CrossRef] [PubMed]
- Tucureanu, M.M.; Filippi, A.; Alexandru, N.; Ana Constantinescu, C.; Ciortan, L.; Macarie, R.; Vadana, M.; Voicu, G.; Frunza, S.; Nistor, D.; et al. Diabetes-induced early molecular and functional changes in aortic heart valves in a murine model of atherosclerosis. Diabetes Vasc. Dis. Res. 2019, 16, 562–576. [Google Scholar] [CrossRef] [Green Version]
- Teo, K.K.; Corsi, D.J.; Tam, J.W.; Dumesnil, J.G.; Chan, K.L. Lipid lowering on progression of mild to moderate aortic stenosis: Meta-analysis of the randomized placebo-controlled clinical trials on 2344 patients. Can. J. Cardiol. 2011, 27, 800–808. [Google Scholar] [CrossRef]
- Andersson, C.; Abdulla, J. Is the use of renin-angiotensin system inhibitors in patients with aortic valve stenosis safe and of prognostic benefit? A systematic review and meta-analysis. Eur. Heart J. Cardiovasc. Pharmacother. 2017, 3, 21–27. [Google Scholar] [CrossRef] [Green Version]
- Gyongyosi, M.; Haller, P.M.; Blake, D.J.; Martin Rendon, E. Meta-Analysis of Cell Therapy Studies in Heart Failure and Acute Myocardial Infarction. Circ. Res 2018, 123, 301–308. [Google Scholar] [CrossRef] [PubMed]
- Sacks, M.S.; Schoen, F.J.; Mayer, J.E. Bioengineering challenges for heart valve tissue engineering. Annu. Rev. Biomed. Eng. 2009, 11, 289–313. [Google Scholar] [CrossRef] [PubMed]
- Movileanu, I.; Nistor, D.; Sierad, L.; Brinzaniuc, K.; Oprita, C.; Harpa, M.; Preda, T.; Cotoi, O.; Al Hussein, H.; Al Hussein, H.; et al. Developing the tissue engineered heart valve-a descriptive hemodynamic and ultrasound in vitro characterization study of heart valves in a bioreactor. Rom. J. Cardiol. 2021, 31, 555–563. [Google Scholar] [CrossRef]
- Caplan, A.I. Mesenchymal Stem Cells: Time to Change the Name! Stem Cells Transl. Med. 2017, 6, 1445–1451. [Google Scholar] [CrossRef] [Green Version]
- Jana, S.; Tranquillo, R.T.; Lerman, A. Cells for tissue engineering of cardiac valves. J. Tissue Eng. Regen. Med. 2016, 10, 804–824. [Google Scholar] [CrossRef]
- Loomans, C.J.; de Koning, E.J.; Staal, F.J.; Rookmaaker, M.B.; Verseyden, C.; de Boer, H.C.; Verhaar, M.C.; Braam, B.; Rabelink, T.J.; van Zonneveld, A.J. Endothelial progenitor cell dysfunction: A novel concept in the pathogenesis of vascular complications of type 1 diabetes. Diabetes 2004, 53, 195–199. [Google Scholar] [CrossRef] [Green Version]
- Sibal, L.; Aldibbiat, A.; Agarwal, S.C.; Mitchell, G.; Oates, C.; Razvi, S.; Weaver, J.U.; Shaw, J.A.; Home, P.D. Circulating endothelial progenitor cells, endothelial function, carotid intima-media thickness and circulating markers of endothelial dysfunction in people with type 1 diabetes without macrovascular disease or microalbuminuria. Diabetologia 2009, 52, 1464–1473. [Google Scholar] [CrossRef] [Green Version]
- Hortenhuber, T.; Rami-Mehar, B.; Satler, M.; Nagl, K.; Hobaus, C.; Hollerl, F.; Koppensteiner, R.; Schernthaner, G.; Schober, E.; Schernthaner, G.H. Endothelial progenitor cells are related to glycemic control in children with type 1 diabetes over time. Diabetes Care 2013, 36, 1647–1653. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Dong, J.; Lobe, C.G.; Gong, P.; Liu, J.; Liao, L. CCR5 facilitates endothelial progenitor cell recruitment and promotes the stabilization of atherosclerotic plaques in ApoE−/− mice. Stem Cell Res. Ther. 2015, 6, 36. [Google Scholar] [CrossRef] [Green Version]
- Georgescu, A.; Alexandru, N.; Constantinescu, A.; Titorencu, I.; Popov, D. The promise of EPC-based therapies on vascular dysfunction in diabetes. Eur. J. Pharmacol. 2011, 669, 1–6. [Google Scholar] [CrossRef]
- Sen, S.; McDonald, S.P.; Coates, P.T.; Bonder, C.S. Endothelial progenitor cells: Novel biomarker and promising cell therapy for cardiovascular disease. Clin. Sci. 2011, 120, 263–283. [Google Scholar] [CrossRef] [Green Version]
- Georgescu, A.; Alexandru, N.; Andrei, E.; Dragan, E.; Cochior, D.; Dias, S. Effects of transplanted circulating endothelial progenitor cells and platelet microparticles in atherosclerosis development. Biol. Cell. 2016, 108, 219–243. [Google Scholar] [CrossRef]
- Filippi, A.; Constantin, A.; Alexandru, N.; Voicu, G.; Constantinescu, C.A.; Rebleanu, D.; Fenyo, M.; Simionescu, D.; Simionescu, A.; Manduteanu, I.; et al. Integrins α4β1 and αVβ3 are reduced in endothelial progenitor cells from diabetic dyslipidemic mice and may represent new targets for therapy in aortic valve disease. Cell Transpl. 2020, 29, 963689720946277. [Google Scholar] [CrossRef]
- Abplanalp, W.T.; Conklin, D.J.; Cantor, J.M.; Ginsberg, M.H.; Wysoczynski, M.; Bhatnagar, A.; O’Toole, T.E. Enhanced integrin alpha4beta1-mediated adhesion contributes to a mobilization defect of endothelial progenitor cells in diabetes. Diabetes 2016, 65, 3505–3515. [Google Scholar] [CrossRef] [Green Version]
- Georgescu, A.; Alexandru, N.; Andrei, E.; Titorencu, I.; Dragan, E.; Tarziu, C.; Ghiorghe, S.; Badila, E.; Bartos, D.; Popov, D. Circulating microparticles and endothelial progenitor cells in atherosclerosis: Pharmacological effects of irbesartan. J. Thromb. Haemost. 2012, 10, 680–691. [Google Scholar] [CrossRef]
- Jansig, E.; Geissler, S.; Rieckmann, V.; Kuenemund, A.; Hietel, B.; Schenk, M.; Wussow, S.; Kreideweiss, P.; Panzner, S.; Reinsch, C.; et al. Viromers as carriers for mRNA-mediated expression of therapeutic molecules under inflammatory conditions. Sci. Rep. 2020, 10, 15090. [Google Scholar] [CrossRef]
- Vlad, M.L.; Manea, S.A.; Lazar, A.G.; Raicu, M.; Muresian, H.; Simionescu, M.; Manea, A. Histone acetyltransferase-dependent pathways mediate upregulation of NADPH oxidase 5 in human macrophages under inflammatory conditions: A potential mechanism of reactive oxygen species overproduction in atherosclerosis. Oxid. Med. Cell Longev. 2019, 2019, 3201062. [Google Scholar] [CrossRef] [Green Version]
- Urban, P.; Rabajdova, M.; Spakova, I.; Sabol, F.; Mickova, H.; Lakatosova, K.; Zavacka, M. Molecular recognition of aortic valve stenosis and regurgitation. Eur. Rev. Med. Pharmacol. Sci. 2019, 23, 10996–11003. [Google Scholar] [CrossRef]
- Ladhoff, J.; Fleischer, B.; Hara, Y.; Volk, H.D.; Seifert, M. Immune privilege of endothelial cells differentiated from endothelial progenitor cells. Cardiovasc. Res. 2010, 88, 121–129. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Animal Groups | Strain | Diabetic | Dyslipidemic | Injection with EPCs from C57BL or ApoE−/− mice | EPCs Transfected with GFP or α4β1 | Animal Number |
|---|---|---|---|---|---|---|
| CIT-EPC-C57 | ApoE−/− | No | YES | C57BL—Healthy | - | 4 |
| STZ-EPC-C57 | ApoE−/− | YES | YES | C57BL—Healthy | - | 4 |
| CIT-EPC-GFP | ApoE−/− | No | YES | ApoE−/−—Dyslipidemic | GFP | 4 |
| STZ-EPC-GFP | ApoE−/− | YES | YES | ApoE−/−—Dyslipidemic, Diabetic | GFP | 4 (3) |
| CIT-EPC-VLA4 | ApoE−/− | No | YES | ApoE−/−—Dyslipidemic | α4β1 | 4 |
| STZ-EPC-VLA4 | ApoE−/− | YES | YES | ApoE−/−—Dyslipidemic, Diabetic | α4β1 | 4 |
| CIT-EPC-C57 | STZ-EPC-C57 | CIT-EPC-GFP | STZ-EPC-GFP | CIT-EPC-VLA4 | STZ-EPC-VLA4 | Statistical Significance # | |
|---|---|---|---|---|---|---|---|
| Glycaemia (mg/dL) | 133.5 ± 17.6 | 235 ± 60.02 | 192.8 ± 23.77 | 280.7 ± 103.8 | 103.5 ± 19.71 | 342.5 ± 23.19 | CIT-EPC-C57 vs. STZ-EPC-C57, * p < 0.05; STZ-EPC-C57 vs. STZ-EPC-VLA4, * p < 0.05; CIT-EPC-VLA4 vs. STZ-EPC-VLA4, ** p < 0.01 |
| HbA1c (%) | 3.84 ± 0.968 | 3.028 ± 0.565 | 4.164 ± 1.228 | 3.704 ± 0.879 | 4.035 ± 0.259 | 3.404 ± 0.338 | |
| Fetuin A (µg/mL) | 72.62 ± 11.28 | 89.54 ± 29.07 | 87.58 ± 36.37 | 76.29 ± 15.07 | 75.61 ± 13.63 | 90.53 ± 7.68 | |
| Triglycerides (mg/dL) | 214.8 ± 100.1 | 380.8 ± 55.44 | 158.2 ± 64.8 | 409 ± 132 | 83.84 ± 23.47 | 244 ± 102.7 | CIT-EPC-GFP vs. STZ-EPC-GFP, * p < 0.05 |
| Total cholesterol (mg/dL) | 1224 ± 339.2 | 1516 ± 245.8 | 1124 ± 80.25 | 1345 ± 136 | 961 ± 170.9 | 1282 ± 106 | |
| HDL Cholesterol (mg/dL) | 58.18 ± 9.535 | 65.19 ± 7.167 | 68.48 ± 12.21 | 37.84 ± 2.119 | 58.87 ± 5.395 | 60.51 ± 11.21 | CIT-EPC-GFP vs. STZ-EPC-GFP, ** p < 0.01; STZ-EPC-GFP vs. STZ-EPC-VLA4, * p < 0.05; |
| LDL Cholesterol (mg/dL) | 390.4 ± 15.3 | 414.4 ± 9.522 | 449.9 ± 77.25 | 450.3 ± 7.774 | 349.4 ± 59.59 | 322.8 ± 110 | |
| ALT (U/L) | 22.1 ± 11.85 | 23.83 ± 6.203 | 8.87 ± 3.782 | 11.92 ± 6.915 | 19.39 ± 9.38 | 12.04 ± 2.238 | |
| AST (U/L) | 42.81 ± 13.21 | 49.15 ± 26.77 | 28.42 ± 6.9 | 36.61 ± 7.452 | 49.29 ± 24.67 | 43.01 ± 11.5 | |
| Creatinine (mg/dL) | 1.093 ± 0.83 | 1.827 ± 1.125 | 0.347 ± 0.463 | 1.396 ± 0.248 | 0.474 ± 0.268 | 0.717 ± 0.508 |
| Animal Groups | Strain | Diabetic | Dyslipidemic | Injection with EPCs from C57BL or ApoE−/− Mice | EPCs Transfected with GFP or α4β1 | Animal Number |
|---|---|---|---|---|---|---|
| CIT-AFL | ApoE−/− | No | YES | - | - | 2 |
| STZ-AFL | ApoE−/− | YES | YES | - | - | 2 |
| CIT-EPC-C57 | ApoE−/− | No | YES | C57BL—Healthy | - | 2 |
| STZ-EPC-C57 | ApoE−/− | YES | YES | C57BL—Healthy | - | 2 |
| CIT-EPC-GFP | ApoE−/− | No | YES | ApoE−/−—Dyslipidemic | GFP | 2 |
| STZ-EPC-GFP | ApoE−/− | YES | YES | ApoE−/−—Dyslipidemic, Diabetic | GFP | 2 |
| CIT-EPC-VLA4 | ApoE−/− | No | YES | ApoE−/−—Dyslipidemic | α4β1 | 4 |
| STZ-EPC-VLA4 | ApoE−/− | YES | YES | ApoE−/−—Dyslipidemic, Diabetic | α4β1 | 4 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Filippi, A.; Constantin, A.; Alexandru, N.; Mocanu, C.A.; Vlad, M.L.; Fenyo, I.M.; Simionescu, A.; Simionescu, D.T.; Manduteanu, I.; Georgescu, A. VLA4-Enhanced Allogeneic Endothelial Progenitor Cell-Based Therapy Preserves the Aortic Valve Function in a Mouse Model of Dyslipidemia and Diabetes. Pharmaceutics 2022, 14, 1077. https://doi.org/10.3390/pharmaceutics14051077
Filippi A, Constantin A, Alexandru N, Mocanu CA, Vlad ML, Fenyo IM, Simionescu A, Simionescu DT, Manduteanu I, Georgescu A. VLA4-Enhanced Allogeneic Endothelial Progenitor Cell-Based Therapy Preserves the Aortic Valve Function in a Mouse Model of Dyslipidemia and Diabetes. Pharmaceutics. 2022; 14(5):1077. https://doi.org/10.3390/pharmaceutics14051077
Chicago/Turabian StyleFilippi, Alexandru, Alina Constantin, Nicoleta Alexandru, Cristina Ana Mocanu, Mihaela Loredana Vlad, Ioana Madalina Fenyo, Agneta Simionescu, Dan Teodor Simionescu, Ileana Manduteanu, and Adriana Georgescu. 2022. "VLA4-Enhanced Allogeneic Endothelial Progenitor Cell-Based Therapy Preserves the Aortic Valve Function in a Mouse Model of Dyslipidemia and Diabetes" Pharmaceutics 14, no. 5: 1077. https://doi.org/10.3390/pharmaceutics14051077