
Evaluation of CTB-sLip for Targeting Lung Metastasis of Colorectal Cancer

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals and Cells
2.2. Materials
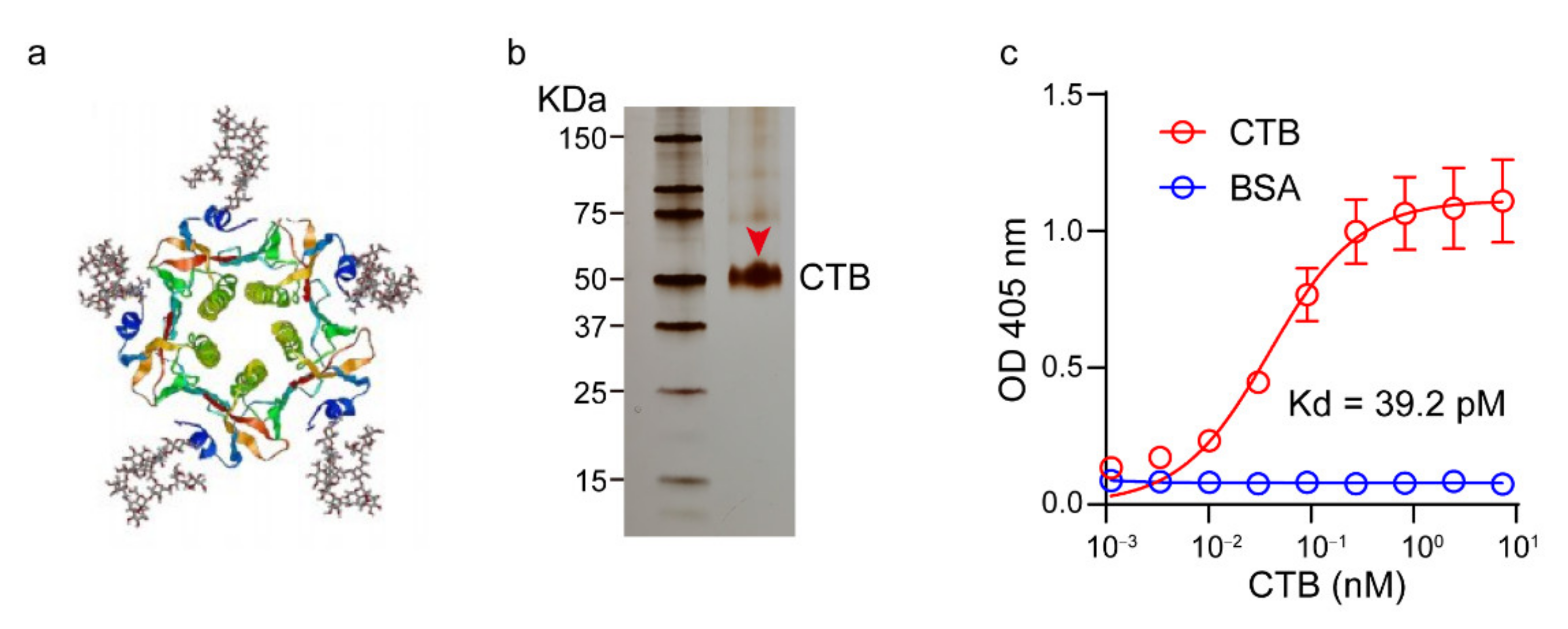
2.3. Expression and Characterization of the CTB Protein
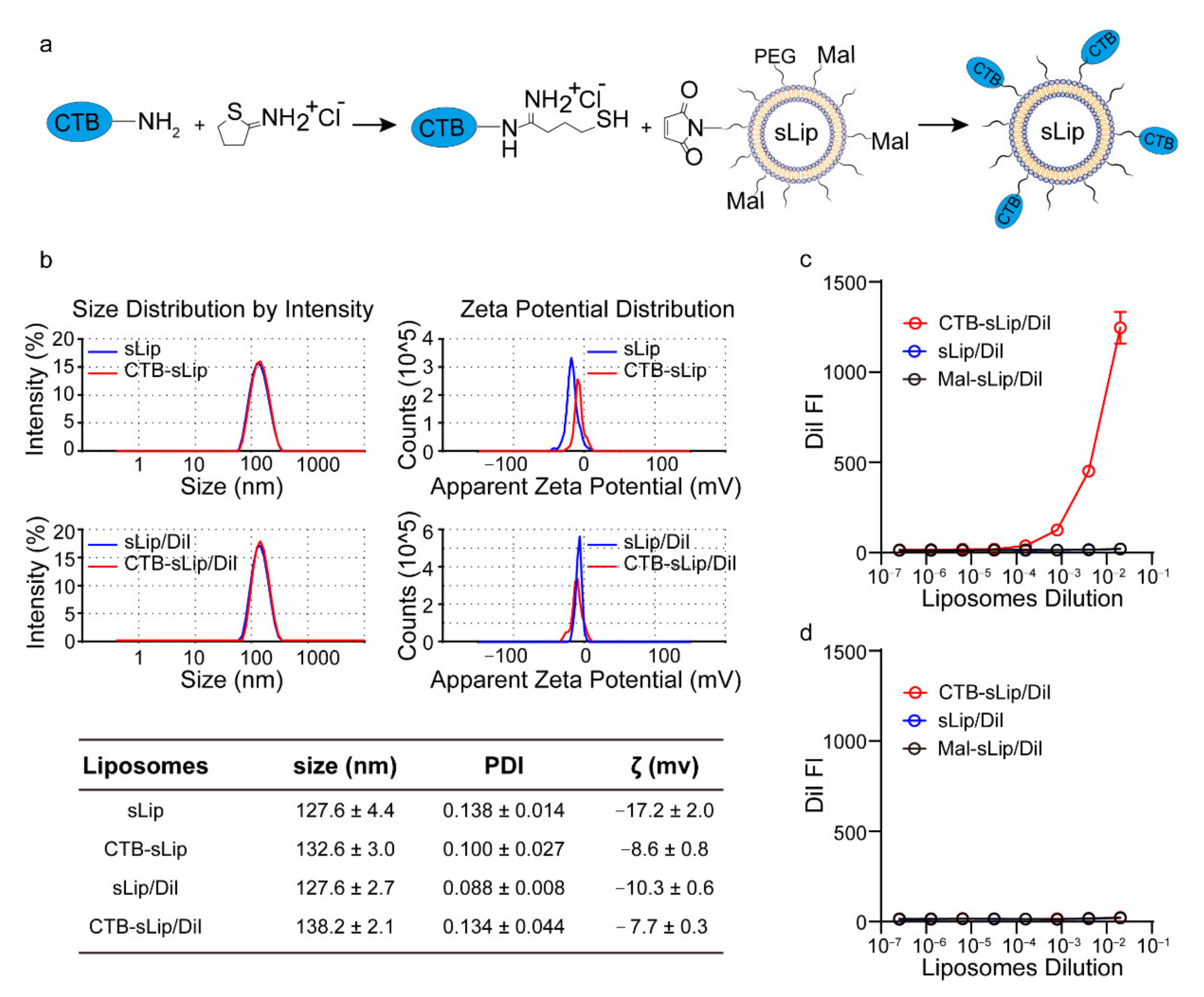
2.4. Preparation and Characterization of Liposomes
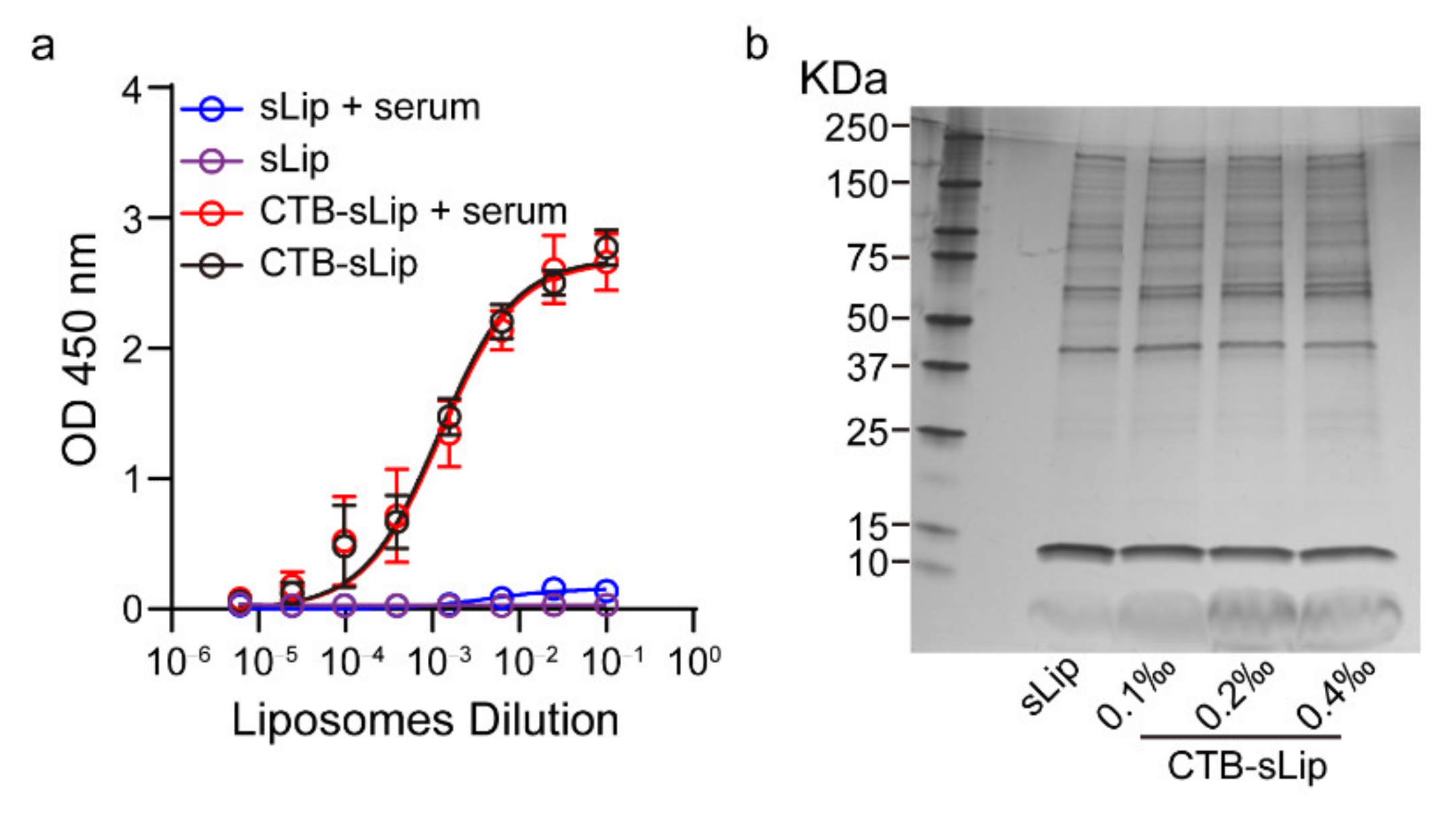
2.5. Characterization of Liposome Stability in Mouse Serum
2.6. Binding Activity of CTB-sLip with GM1 In Vitro
2.7. Protein Corona Separation and Characterization
2.8. Pharmacokinetic Profile of CTB-sLip/DiI
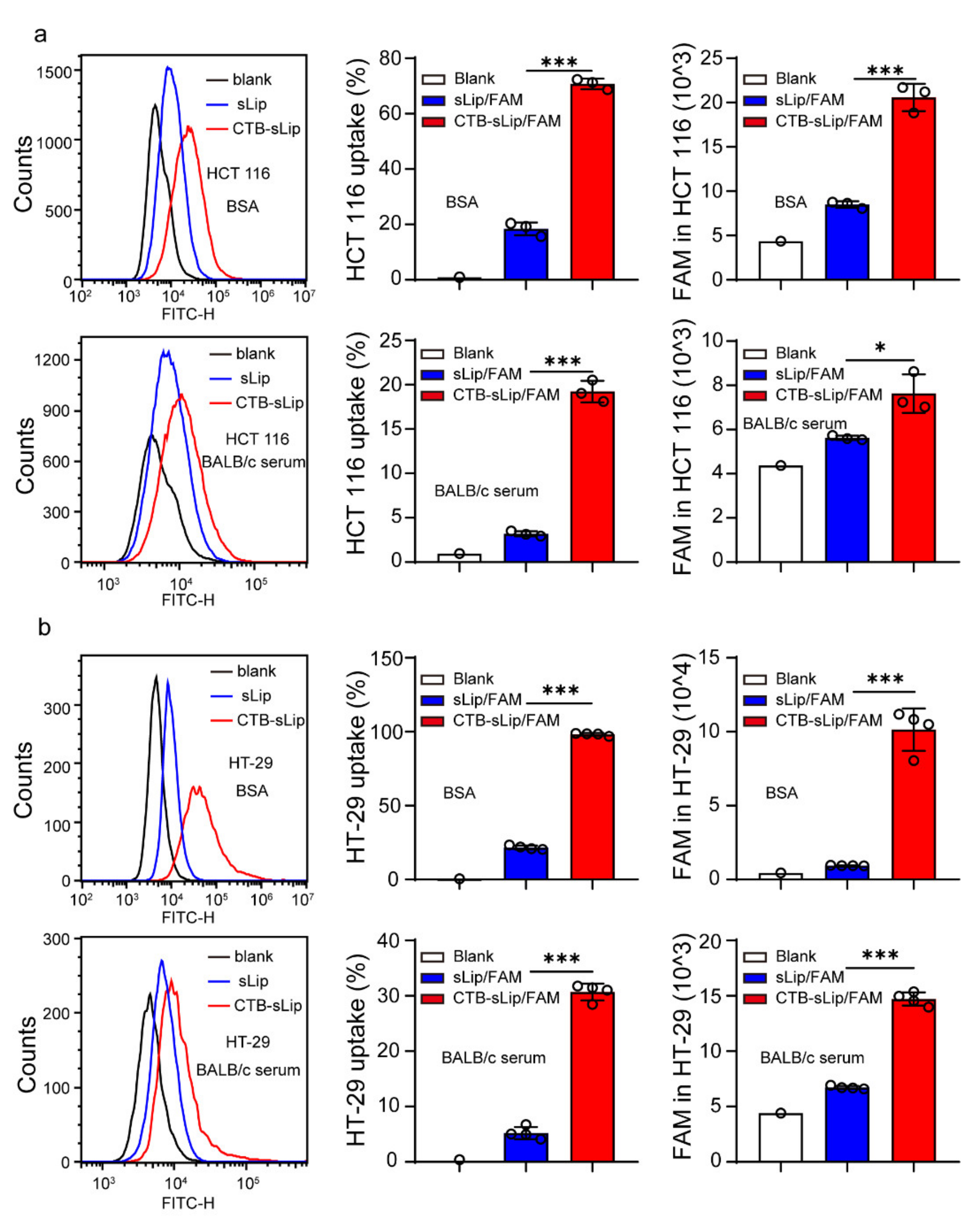
2.9. Cellular Uptake and Binding of CTB-sLip/FAM In Vitro
2.10. Uptake of CTB-FITC to Colorectal Cancer Cell Lines
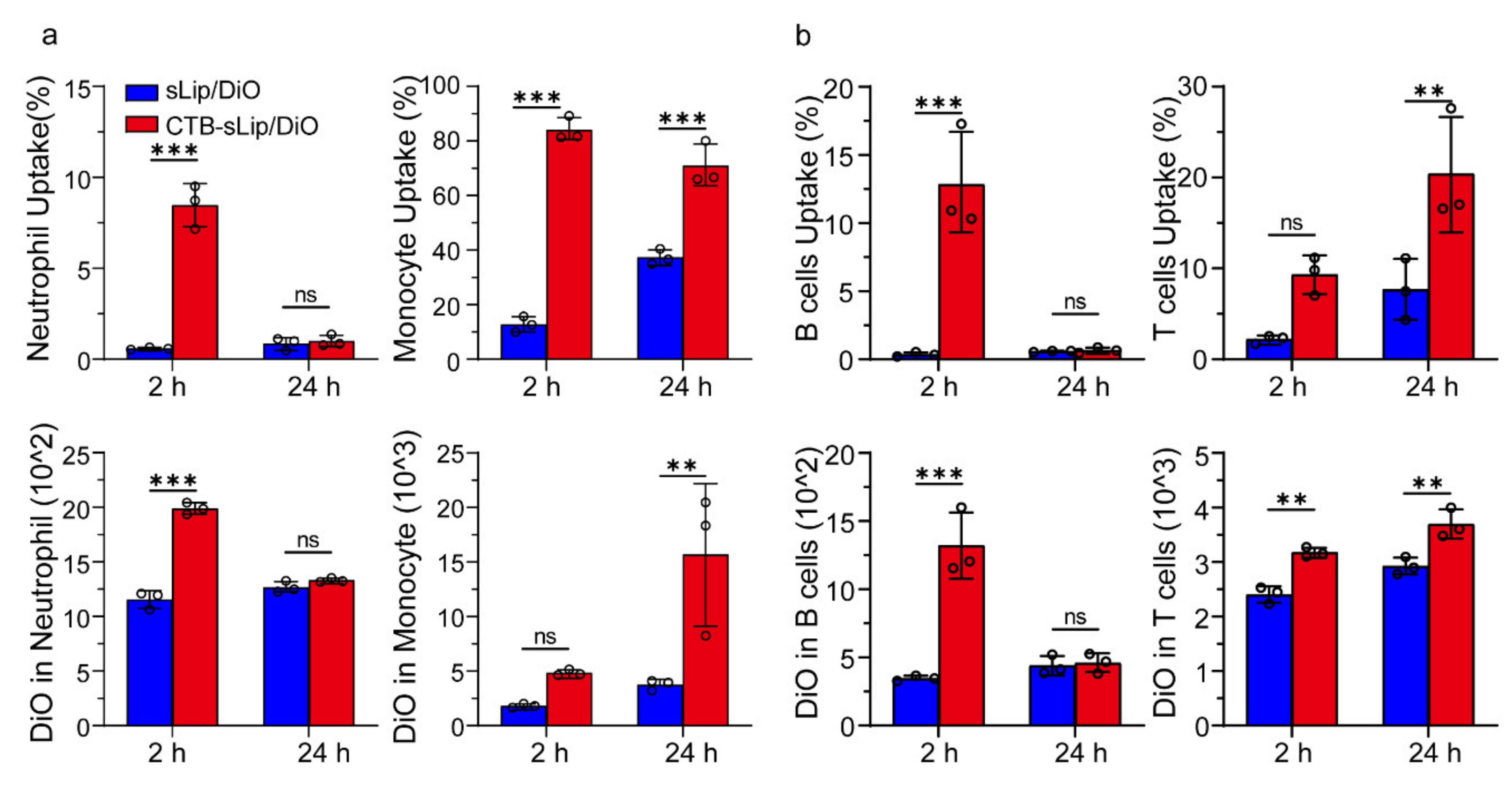
2.11. Uptake of Liposomes by Blood Cells of BALB/c Mice In Vivo
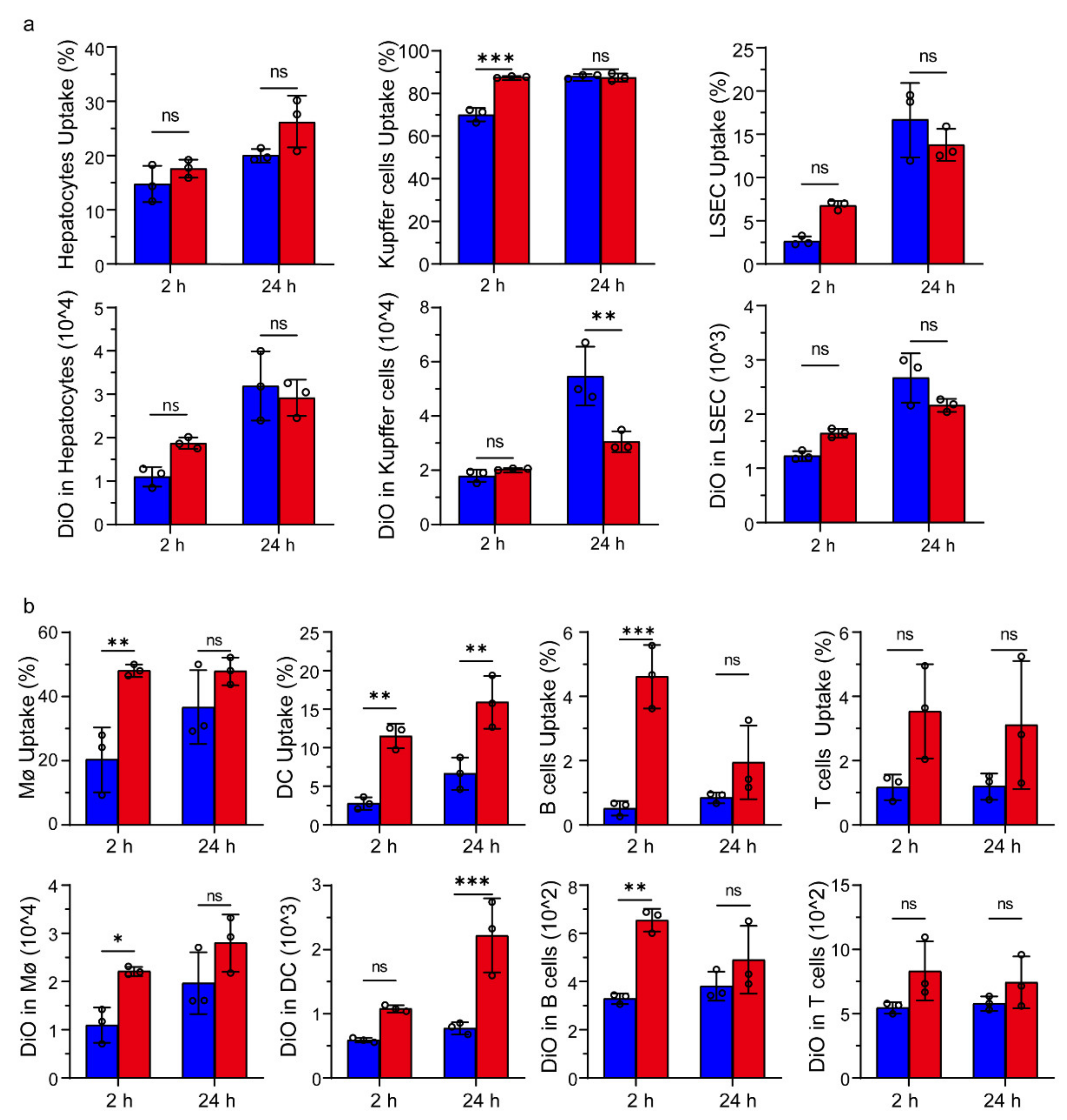
2.12. Cellular Uptake of Liposomes in Liver and Splenic Cells In Vivo
2.13. Distribution of Liposomes in BALB/c Nude Mice with Lung Metastasis of Colorectal Cancer
2.14. Immunogenicity of Liposomes
2.15. Statistical Analysis
3. Results and Discussion
3.1. Preparation and Characterization of CTB-Conjugated Liposomes (CTB-sLip)
3.2. Effects of Serum on the Targeting Capability of CTB-sLip
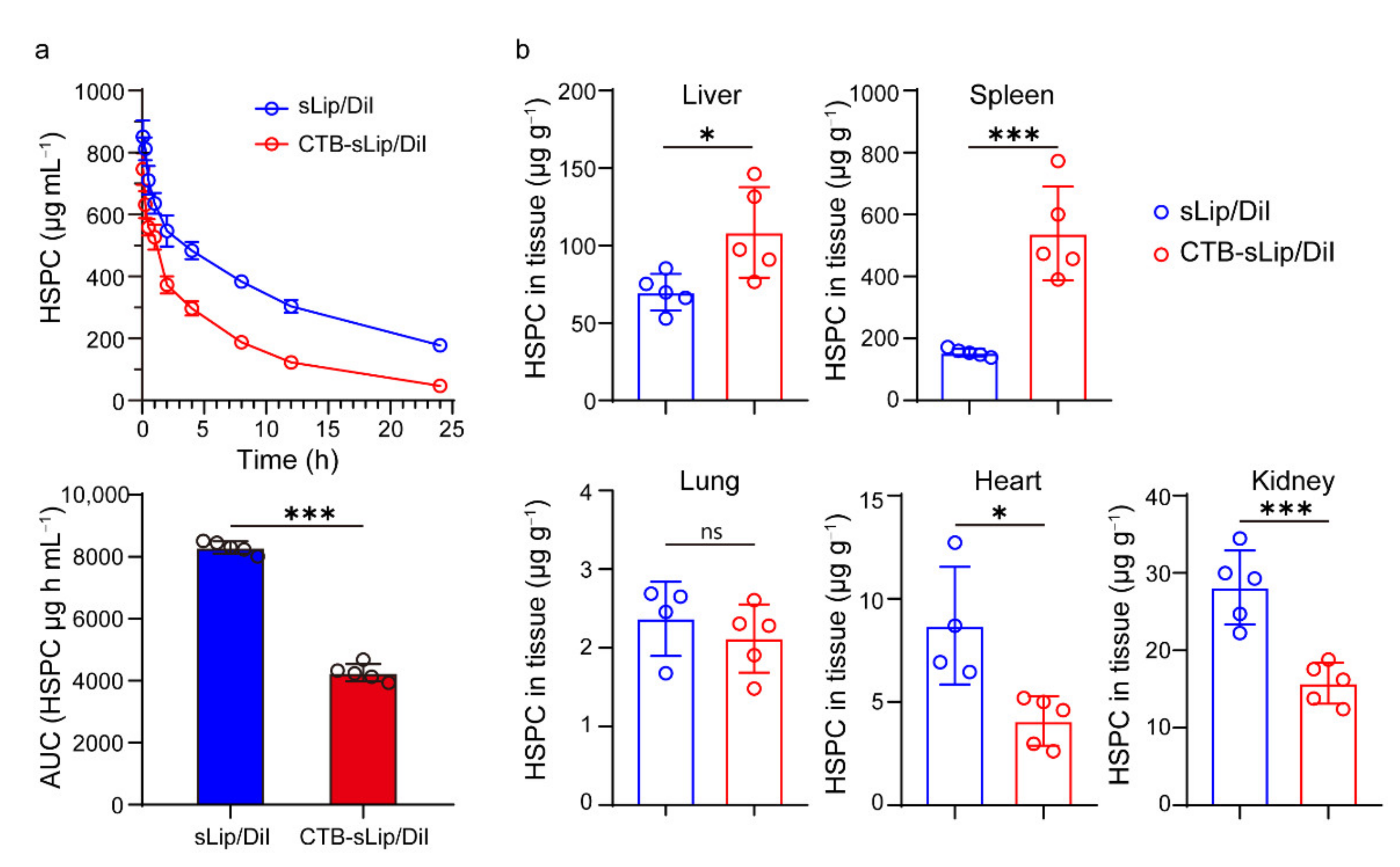
3.3. Pharmacokinetic Profiles of CTB-sLip
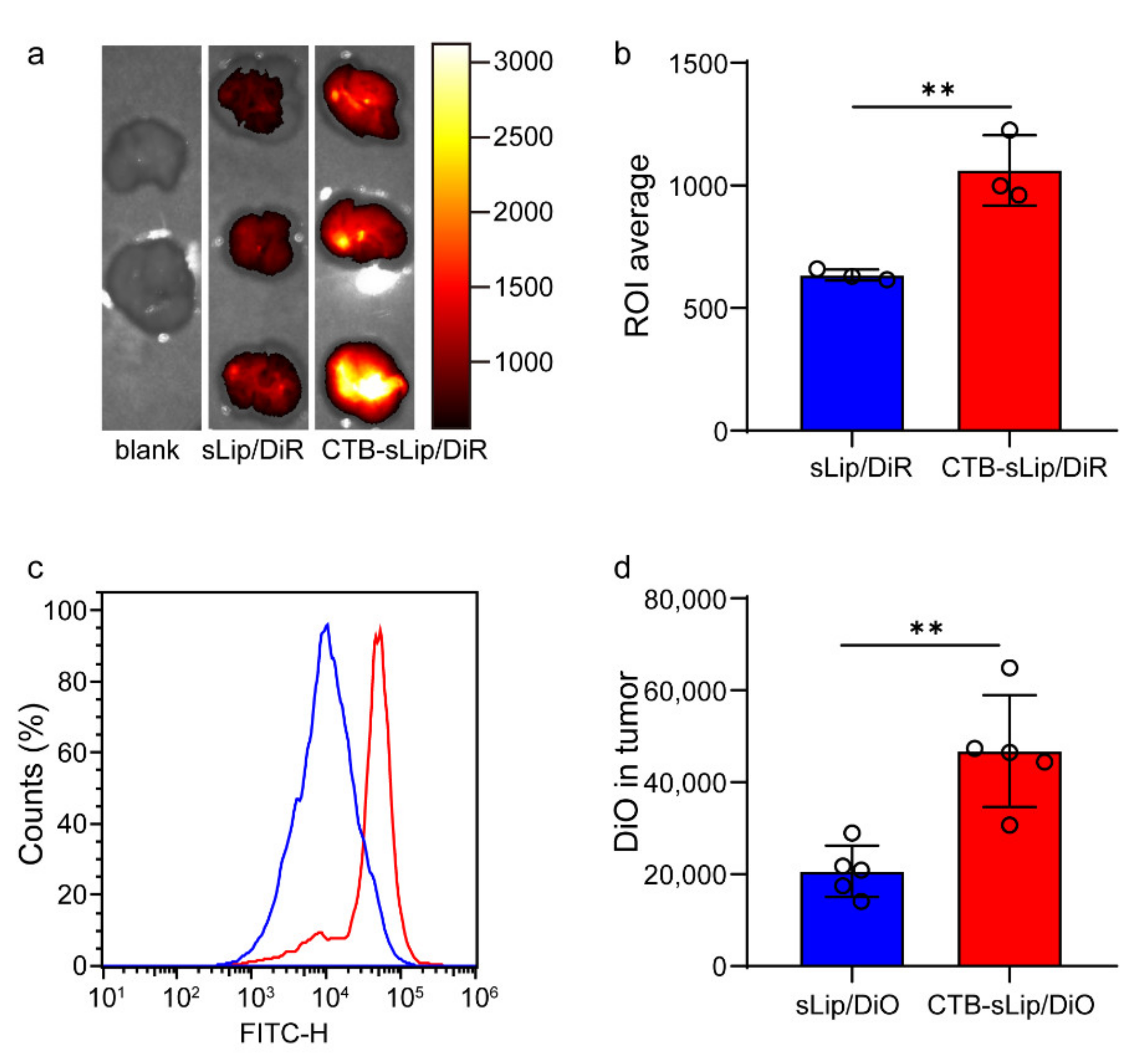
3.4. Lung Metastasis Targeting of Colorectal Cancer by CTB-sLip
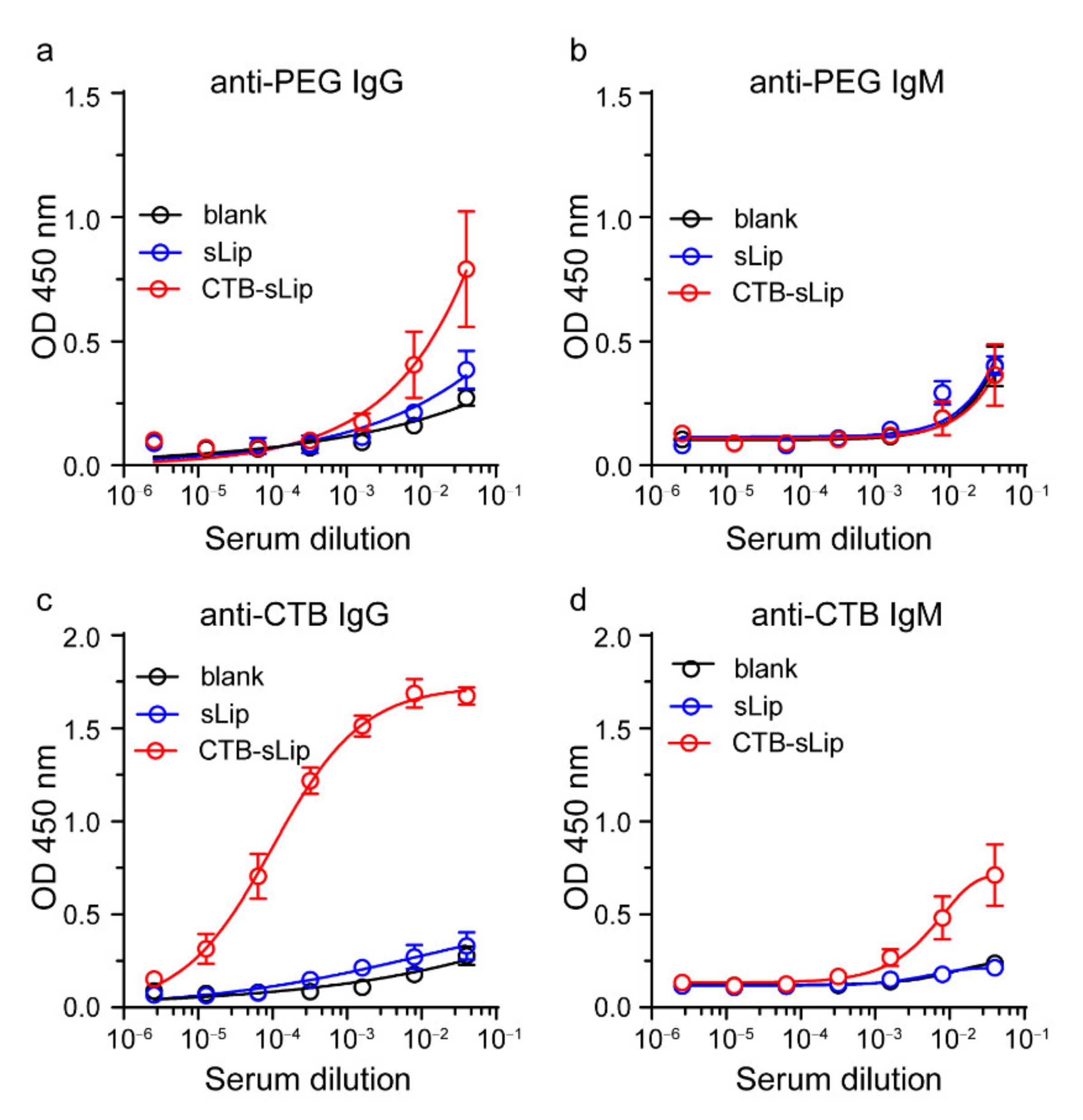
3.5. Immunogenicity of CTB-sLip
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Conflicts of Interest
References
- Siegel, R.L.; Miller, K.D.; Sauer, A.G.; Fedewa, S.A.; Butterly, L.F.; Anderson, J.C.; Cercek, A.; Smith, R.A.; Jemal, A. Colorectal cancer statistics. CA Cancer J. Clin. 2020, 70, 145–164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hiratsuka, S.; Watanabe, A.; Aburatani, H.; Maru, Y. Tumour-mediated upregulation of chemoattractants and recruitment of myeloid cells predetermines lung metastasis. Nat. Cell Biol. 2006, 8, 1369–1375. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yuan, Y.; Yang, F.; Wang, Y.; Zhu, X.; Wang, Z.; Zheng, S.; Wan, D.; He, J.; Wang, J.; et al. Expert consensus on multidisciplinary therapy of colorectal cancer with lung metastases. J. Hematol. Oncol. 2019, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.; Ng, A.S.; Cai, S.; Li, Q.; Yang, L.; Kerr, D. Novel therapeutic strategies: Targeting epithelial–mesenchymal transition in colorectal cancer. Lancet Oncol. 2021, 22, e358–e368. [Google Scholar] [CrossRef]
- Song, L.; Li, Y.; He, B.; Gong, Y. Development of Small Molecules Targeting the Wnt Signaling Pathway in Cancer Stem Cells for the Treatment of Colorectal Cancer. Clin. Colorectal Cancer 2015, 14, 133–145. [Google Scholar] [CrossRef]
- Carvalho, M.R.; Carvalho, C.R.; Maia, F.R.; Caballero, D.; Kundu, S.C.; Reis, R.L.; Oliveira, J.M. Peptide-Modified Dendrimer Nanoparticles for Targeted Therapy of Colorectal Cancer. J. Adv. Ther. 2019, 2, 1900132. [Google Scholar] [CrossRef] [Green Version]
- Sartore-Bianchi, A.; Martini, M.; Molinari, F.; Veronese, S.; Nichelatti, M.; Artale, S.; di Nicolantonio, F.; Saletti, P.; de Dosso, S.; Mazzucchelli, L.; et al. PIK3CA mutations in colorectal cancer are associated with clinical resistance to EGFR-targeted monoclonal antibodies. Cancer Res. 2009, 69, 1851–1857. [Google Scholar] [CrossRef] [Green Version]
- Go, G.; Lee, C.-S.; Yoon, Y.M.; Lim, J.H.; Kim, T.H.; Lee, S.H. PrPC aptamer conjugated–gold nanoparticles for targeted delivery of doxorubicin to colorectal cancer cells. Int. J. Mol. Sci. 2021, 22, 1976. [Google Scholar] [CrossRef]
- Yang, H.; Fu, Y.; Jang, M.-S.; Li, Y.; Lee, J.H.; Chae, H.; Lee, D.S. Interfaces, Multifunctional Polymer Ligand Interface CdZnSeS/ZnS Quantum Dot/Cy3-Labeled Protein Pairs as Sensitive FRET Sensors. J. ACS Appl. Mater. 2016, 8, 35021–35032. [Google Scholar] [CrossRef]
- Morales-Cruz, M.; Delgado, Y.; Castillo, B.; Figueroa, C.M.; Molina, A.M.; Torres, A.; Milian, M.; Griebenow, K. Smart Targeting To Improve Cancer Therapeutics. Drug Des. Dev. Ther. 2019, 13, 3753–3772. [Google Scholar] [CrossRef] [Green Version]
- Zhang, J.; Wei, K.; Shi, J.; Zhu, Y.; Guan, M.; Fu, X.; Zhang, Z. Biomimetic Nanoscale Erythrocyte Delivery System for Enhancing Chemotherapy via Overcoming Biological Barriers. ACS Biomater. Sci. Eng. 2021, 7, 1496–1505. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Guan, J.; Jiang, Z.; Yang, Y.; Liu, J.; Hua, W.; Mao, Y.; Li, C.; Lu, W.; Qian, J.; et al. Brain-targeted drug delivery by manipulating protein corona functions. Nat. Commun. 2019, 10, 3561. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guan, J.; Shen, Q.; Zhang, Z.; Jiang, Z.; Yang, Y.; Lou, M.; Qian, J.; Lu, W.; Zhan, C. Enhanced immunocompatibility of ligand-targeted liposomes by attenuating natural IgM absorption. Nat. Commun. 2018, 9, 2982. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; Wu, T.; Yu, W.; Ruan, S.; He, Q.; Gao, H. Ligand Size and Conformation Affect the Behavior of Nanoparticles Coated with in Vitro and in Vivo Protein Corona. ACS Appl. Mater. Interfaces 2018, 10, 9094–9103. [Google Scholar] [CrossRef]
- Cai, R.; Ren, J.; Ji, Y.; Wang, Y.; Liu, Y.; Chen, Z.; Sabet, Z.F.; Wu, X.; Lynch, I.; Chen, C. Corona of Thorns: The Surface Chemistry-Mediated Protein Corona Perturbs the Recognition and Immune Response of Macrophages. ACS Appl. Mater. Interfaces 2020, 12, 1997–2008. [Google Scholar] [CrossRef]
- Yoo, J.; Park, C.; Yi, G.; Lee, D.; Koo, H. Active Targeting Strategies Using Biological Ligands for Nanoparticle Drug Delivery Systems. Cancers 2019, 11, 640. [Google Scholar] [CrossRef] [Green Version]
- Wei, X.; Zhan, C.; Shen, Q.; Fu, W.; Xie, C.; Gao, J.; Peng, C.; Zheng, P.; Lu, W. A D-peptide ligand of nicotine acetylcholine receptors for brain-targeted drug delivery. Angew. Chem. 2015, 54, 3023–3027. [Google Scholar] [CrossRef]
- Ledeen, R.W.; Wu, G. The multi-tasked life of GM1 ganglioside, a true factotum of nature. Trends Biochem. Sci. 2015, 40, 407–418. [Google Scholar] [CrossRef]
- Ruhlman, T.; Ahangari, R.; Devine, A.; Samsam, M.; Daniell, H. Expression of cholera toxin B-proinsulin fusion protein in lettuce and tobacco chloroplasts—Oral administration protects against development of insulitis in non-obese diabetic mice. Plant Biotechnol. J. 2007, 5, 495–510. [Google Scholar] [CrossRef] [Green Version]
- Guan, J.; Zhang, Z.; Hu, X.; Yang, Y.; Chai, Z.; Liu, X.; Liu, J.; Gao, B.; Lu, W.; Qian, J.J.A.h.m. Cholera Toxin Subunit B Enabled Multifunctional Glioma-Targeted Drug Delivery. Adv. Healthc. Mater. 2017, 6, 1700709. [Google Scholar] [CrossRef]
- Guan, J.; Qian, J.; Zhan, C. Preparation of Cholera Toxin Subunit B Functionalized Nanoparticles for Targeted Therapy of Glioblastoma. Methods Mol. Biol. 2020, 2059, 207–212. [Google Scholar] [PubMed]
- Holmgren, J.; Lonnroth, I.; Mansson, J.; Svennerholm, L. Interaction of cholera toxin and membrane GM1 ganglioside of small intestine. Proc. Natl. Acad. Sci. USA 1975, 72, 2520–2524. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, J.; Holmgren, J. Cholera toxin—A foe & a friend. Indian J. Med. Res. 2011, 133, 153. [Google Scholar] [PubMed]
- Tang, W.; Zhang, Z.; Li, C.; Chu, Y.; Qian, J.; Ying, T.; Lu, W.; Zhan, C. Facile Separation of PEGylated Liposomes Enabled by Anti-PEG scFv. Nano Lett. 2021, 21, 10107–10113. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Chu, Y.; Li, C.; Tang, W.; Qian, J.; Wei, X.; Lu, W.; Ying, T.; Zhan, C. Anti-PEG scFv corona ameliorates accelerated blood clearance phenomenon of PEGylated nanomedicines. J. Control. Release 2021, 330, 493–501. [Google Scholar] [CrossRef]
- Lamichhane, N.; Udayakumar, T.S.; D’Souza, W.D.; Simone, C.B., 2nd; Raghavan, S.R.; Polf, J.; Mahmood, J. Liposomes: Clinical Applications and Potential for Image-Guided Drug Delivery. Molecules 2018, 23, 288. [Google Scholar] [CrossRef] [Green Version]
- Bozzuto, G.; Molinari, A. Liposomes as nanomedical devices. Int. J. Nanomed. 2015, 10, 975–999. [Google Scholar] [CrossRef] [Green Version]
- Aparicio-Vergara, M.; Tencerova, M.; Morgantini, C.; Barreby, E.; Aouadi, M. Isolation of Kupffer Cells and Hepatocytes from a Single Mouse Liver. Methods Mol. Biol. 2017, 1639, 161–171. [Google Scholar]









Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, X.; Tang, W.; Wen, H.; Wu, E.; Ding, T.; Gu, J.; Lv, Z.; Zhan, C. Evaluation of CTB-sLip for Targeting Lung Metastasis of Colorectal Cancer. Pharmaceutics 2022, 14, 868. https://doi.org/10.3390/pharmaceutics14040868
Zhang X, Tang W, Wen H, Wu E, Ding T, Gu J, Lv Z, Zhan C. Evaluation of CTB-sLip for Targeting Lung Metastasis of Colorectal Cancer. Pharmaceutics. 2022; 14(4):868. https://doi.org/10.3390/pharmaceutics14040868
Chicago/Turabian StyleZhang, Xiaoying, Wenjing Tang, Haoyu Wen, Ercan Wu, Tianhao Ding, Jie Gu, Zhongwei Lv, and Changyou Zhan. 2022. "Evaluation of CTB-sLip for Targeting Lung Metastasis of Colorectal Cancer" Pharmaceutics 14, no. 4: 868. https://doi.org/10.3390/pharmaceutics14040868