
Assessing the Anthelmintic Candidates BLK127 and HBK4 for Their Efficacy on Haemonchus contortus Adults and Eggs, and Their Hepatotoxicity and Biotransformation

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Parasites and Hosts
2.3. Egg Hatch Assay
2.4. Testing the Viability of Adult H. contortus
2.5. Hepatotoxicity Testing
2.6. Biotransformation of BLK127 in H. contortus
2.7. Biotransformation of BLK127 in Ovine Liver
2.8. UHPLC-MS and HRMS Conditions
2.9. Statistical Analyses
3. Results
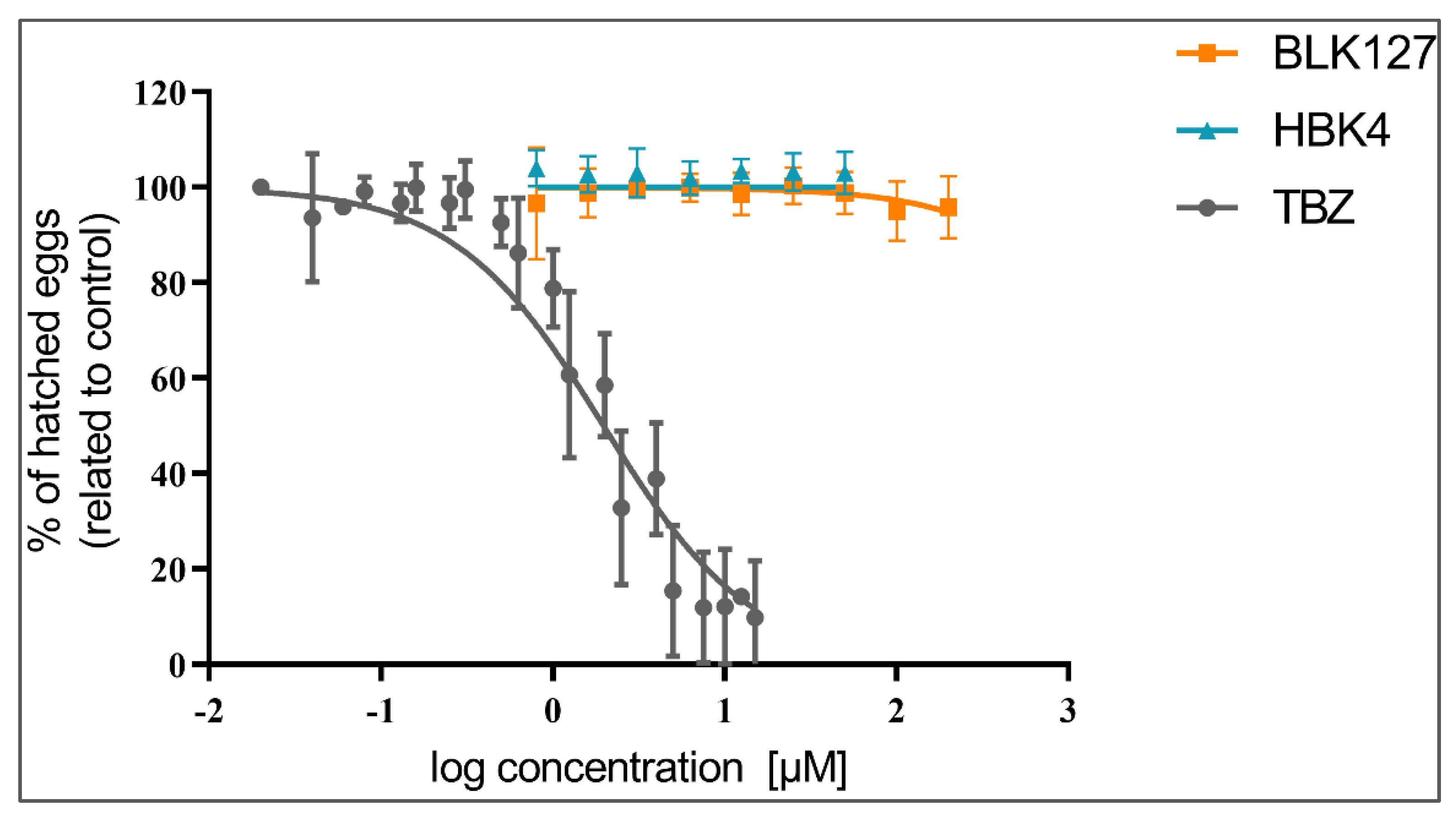
3.1. The Effect of BLK127 and HBK4 on the Hatching of H. contortus Eggs
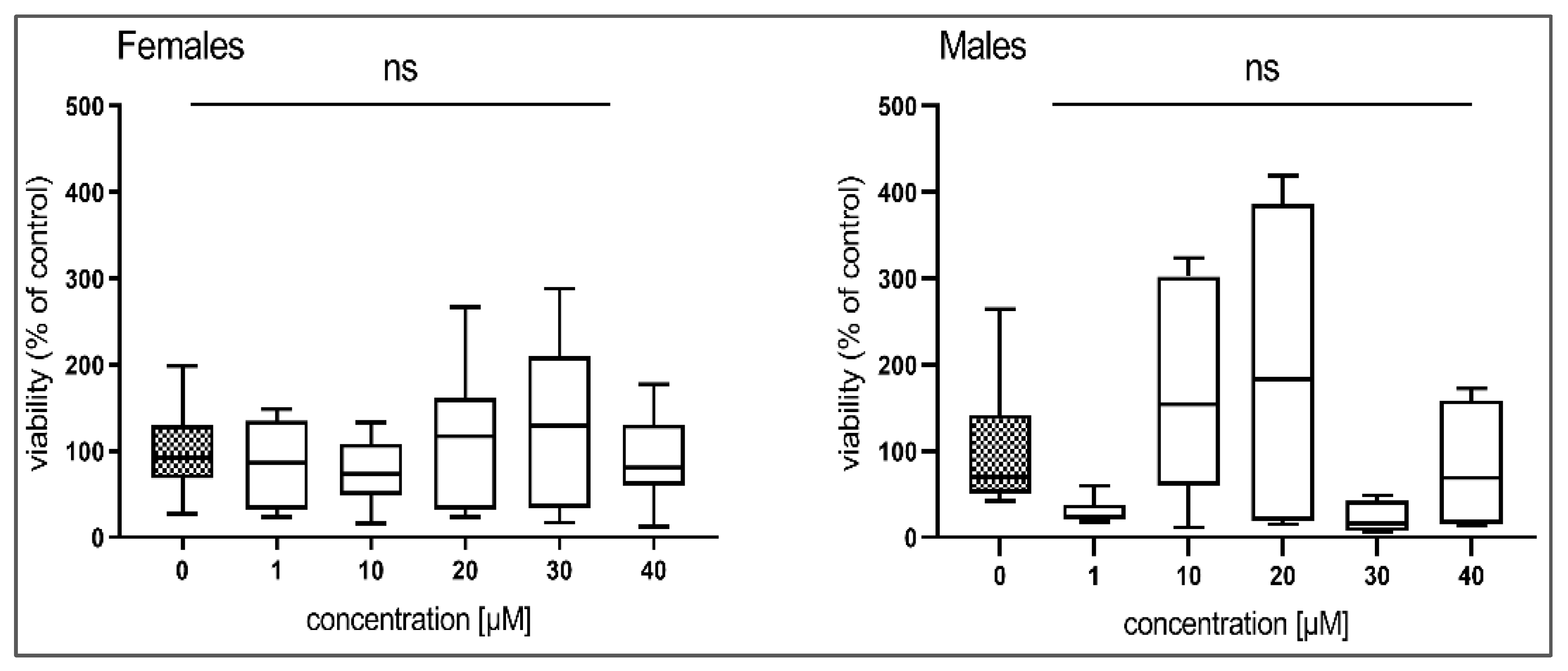
3.2. The Effect of BLK127 and HBK4 on the Viability of H. contortus Adults
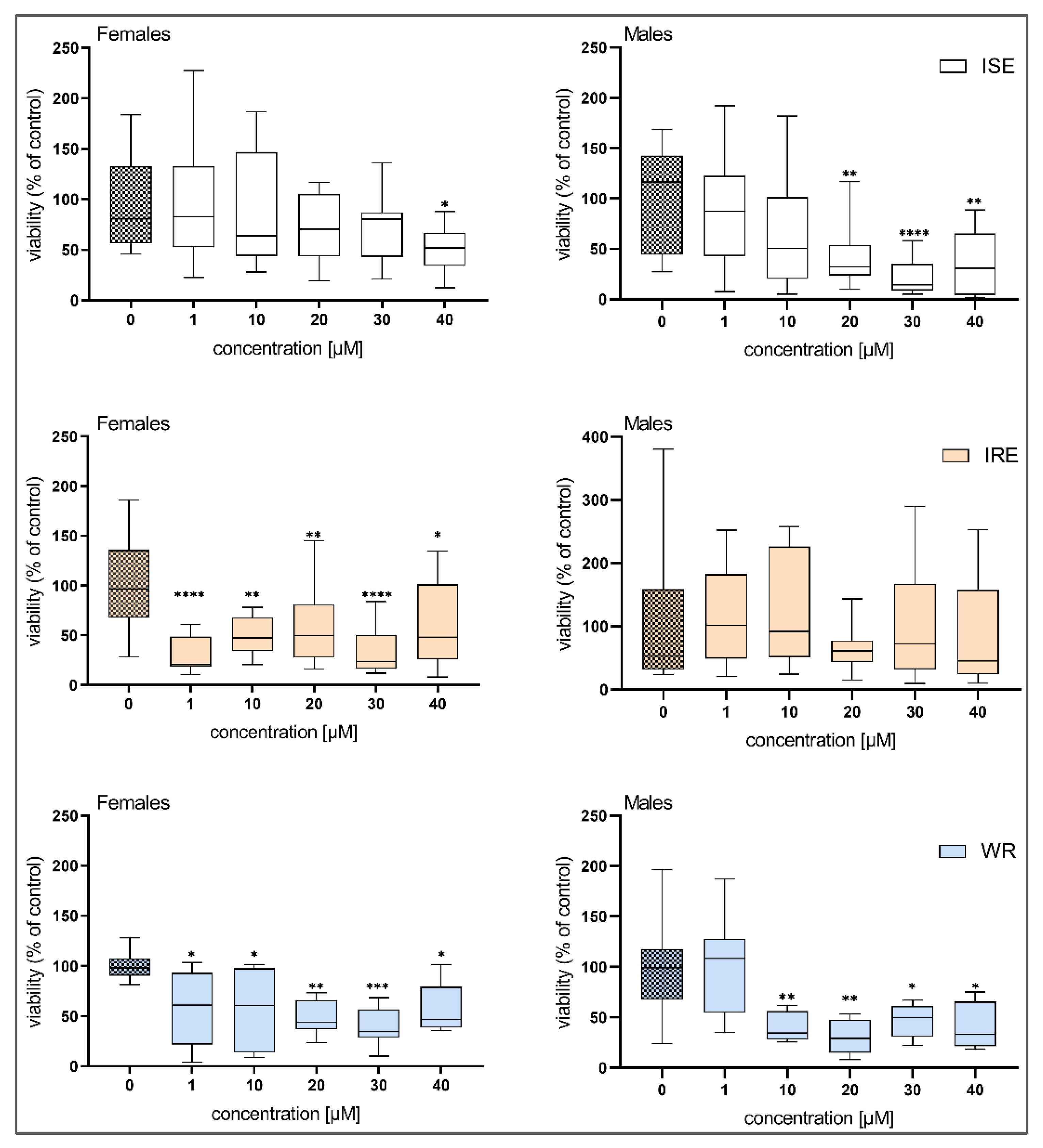
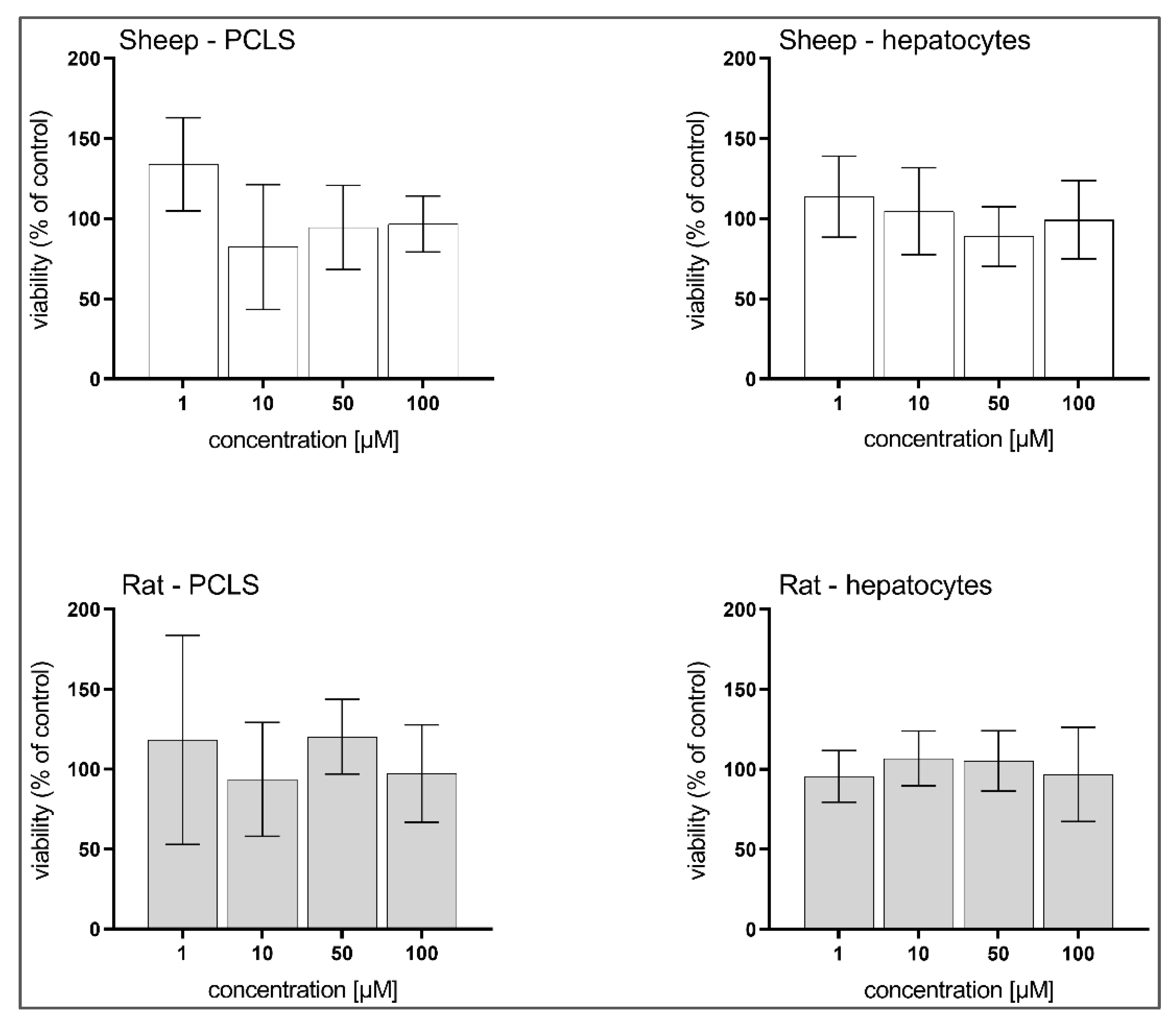
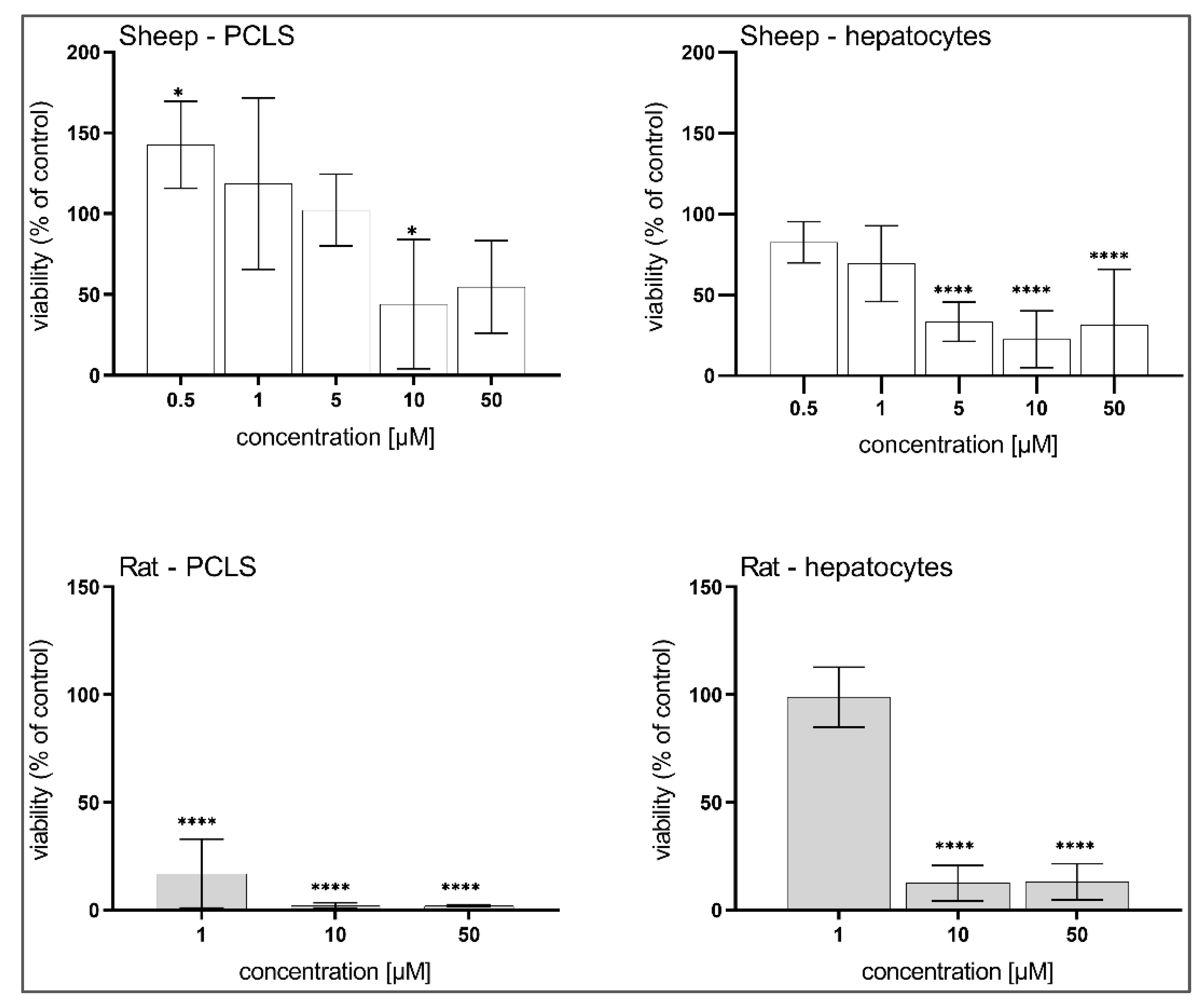
3.3. Hepatotoxicity of BLK127 and HBK4
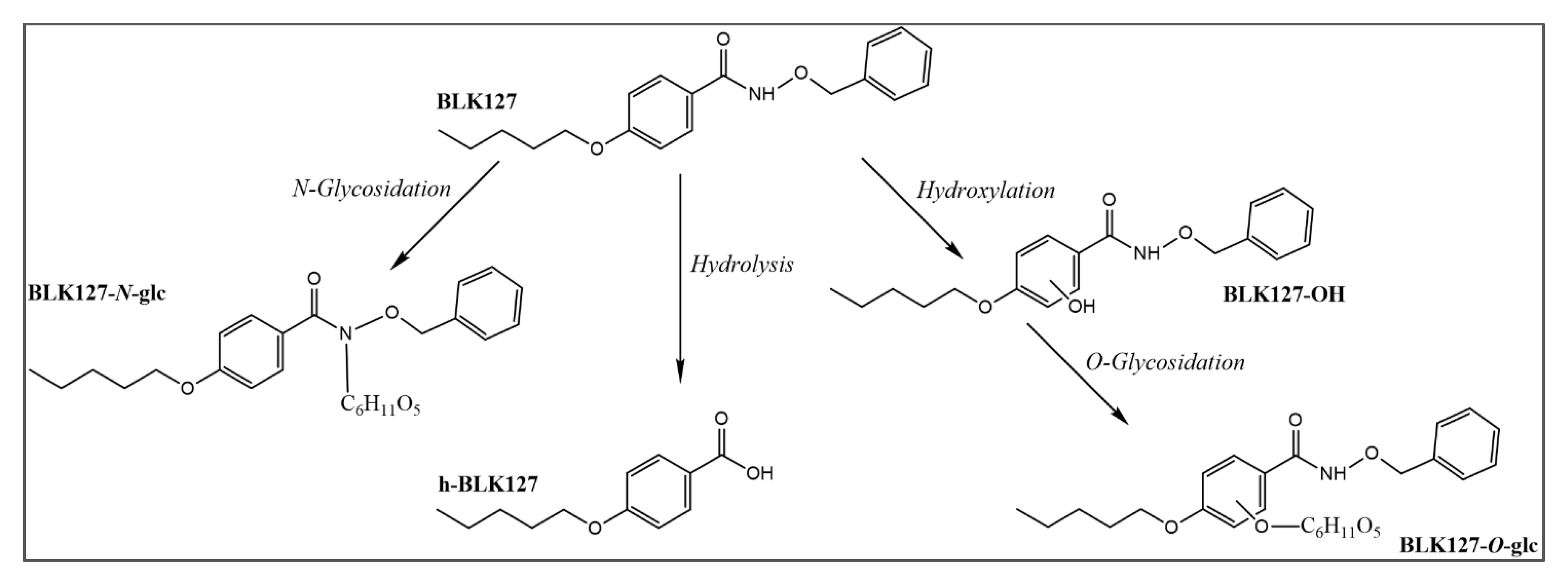
3.4. Biotransformation of BLK127 in H. contortus
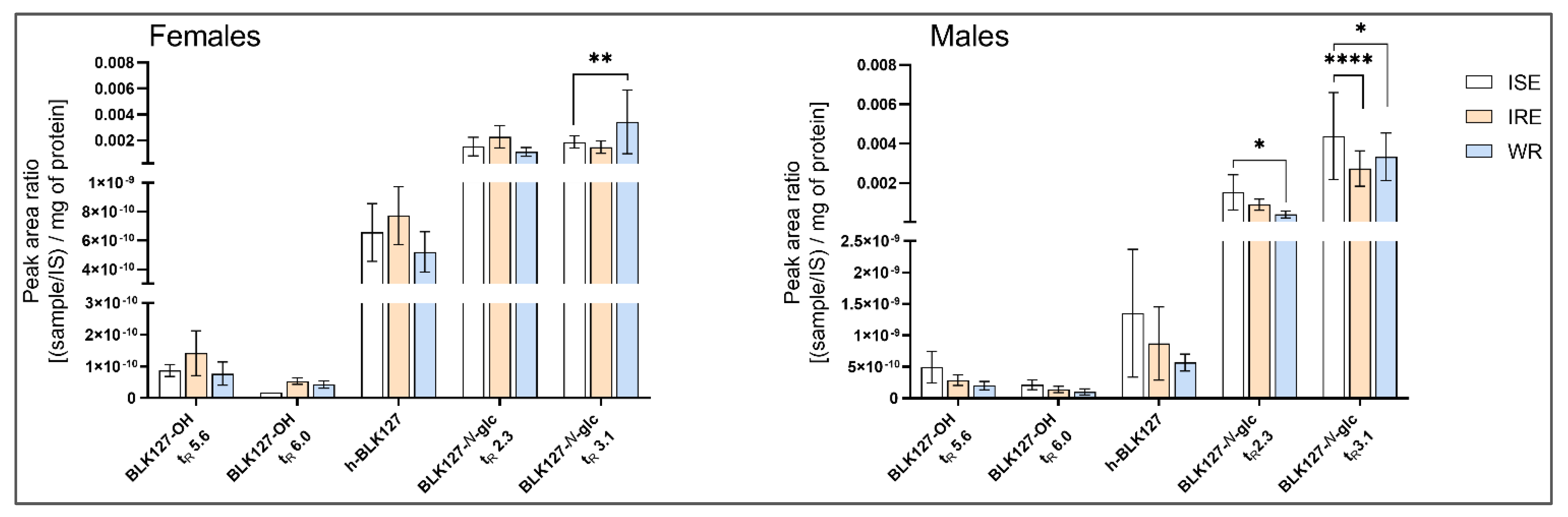
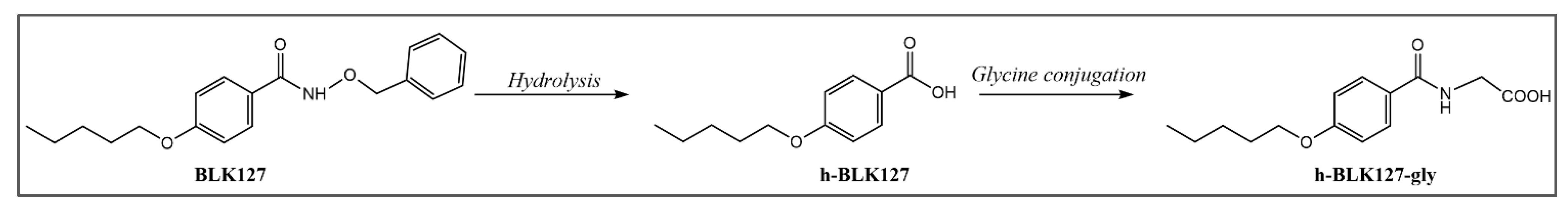
3.5. Biotransformation of BLK127 in Ovine Liver
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Formula | Designation | Lipinski’s Rule of Five | ||||
|---|---|---|---|---|---|---|
| Mw [g/mol] | LogP | tPSA | H-Bond Donors | H-Bond Acceptors | ||
 | BLK127 | 313.40 | 4.63 | 47.56 | 1 | 4 |
 | HBK4 | 504.58 | 3.68 | 116.73 | 3 | 7 |
References
- Kotze, A.C.; Prichard, R.K. Chapter Nine—Anthelmintic Resistance in Haemonchus contortus: History, Mechanisms and Diagnosis. In Advances in Parasitology; Gasser, R.B., Samson-Himmelstjerna, G.V., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 93, pp. 397–428. ISBN 9780128103951. [Google Scholar]
- Laing, R.; Kikuchi, T.; Martinelli, A.; Tsai, I.J.; Beech, R.N.; Redman, E.; Holroyd, N.; Bartley, D.J.; Beasley, H.; Britton, C.; et al. The genome and transcriptome of Haemonchus contortus, a key model parasite for drug and vaccine discovery. Genome Biol. 2013, 14, R88. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stear, M.J.; Doligalska, M.; Donskow-Schmelter, K. Alternatives to anthelmintics for the control of nematodes in livestock. Parasitology 2007, 134, 139–151. [Google Scholar] [CrossRef] [PubMed]
- Prichard, R. Genetic variability following selection of Haemonchus contortus with anthelmintics. Trends Parasitol. 2001, 17, 445–453. [Google Scholar] [CrossRef]
- Zajíčková, M.; Nguyen, L.T.; Skálová, L.; Stuchlíková, L.R.; Matoušková, P. Anthelmintics in the future: Current trends in the discovery and development of new drugs against gastrointestinal nematodes. Drug Discov. Today 2019, 25, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Preston, S.; Jabbar, A.; Gasser, R.B. A perspective on genomic-guided anthelmintic discovery and repurposing using Haemonchus contortus. Infect. Genet. Evol. 2016, 40, 368–373. [Google Scholar] [CrossRef] [PubMed]
- Toutain, P.-L. Pharmacokinetic/pharmacodynamic integration in drug development and dosage-regimen optimization for veterinary medicine. AAPS PharmSci 2002, 4, 160–188. [Google Scholar] [CrossRef]
- Ahmed, I. Pharmaceutical challenges in veterinary product development. Adv. Drug Deliv. Rev. 2002, 54, 871–882. [Google Scholar] [CrossRef]
- From an Idea to the Marketplace: The Journey of an Animal Drug through the Approval Process. Available online: https://www.fda.gov/animal-veterinary/animal-health-literacy/idea-marketplace-journey-animal-drug-through-approval-process (accessed on 20 February 2022).
- Nguyen, L.T.; Kurz, T.; Preston, S.; Brueckmann, H.; Lungerich, B.; Herath, H.; Koehler, A.V.; Wang, T.; Skalova, L.; Jabbar, A.; et al. Phenotypic screening of the ‘Kurz-box’ of chemicals identifies two compounds (BLK127 and HBK4) with anthelmintic activity in vitro against parasitic larval stages of Haemonchus contortus. Parasit. Vectors 2019, 12, 191. [Google Scholar] [CrossRef]
- Jiao, Y.; Preston, S.; Garcia-Bustos, J.F.; Baell, J.B.; Ventura, S.; Le, T.; McNamara, N.; Nguyen, N.; Botteon, A.; Skinner, C.; et al. Tetrahydroquinoxalines induce a lethal evisceration phenotype in Haemonchus contortus in vitro. Int. J. Parasitol. Drugs Drug Resist. 2018, 9, 59–71. [Google Scholar] [CrossRef]
- Geary, T.G. Chapter Ten—Haemonchus contortus: Applications in Drug Discovery. In Advances in Parasitology; Gasser, R.B., Samson-Himmelstjerna, G.V., Eds.; Academic Press: Cambridge, MA, USA, 2016; Volume 93, pp. 429–463. ISBN 9780128103951. [Google Scholar]
- O’Grady, J.; Kotze, A.C. Haemonchus contortus: In vitro drug screening assays with the adult life stage. Exp. Parasitol. 2004, 106, 164–172. [Google Scholar] [CrossRef]
- Gomez-Lechon, M.J.; Lahoz, A.; Gombau, L.; Castell, J.V.; Donato, M.T. In Vitro Evaluation of Potential Hepatotoxicity Induced by Drugs. Curr. Pharm. Des. 2010, 16, 1963–1977. [Google Scholar] [CrossRef] [PubMed]
- Ingawale, D.K.; Mandlik, S.K.; Naik, S.R. Models of hepatotoxicity and the underlying cellular, biochemical and immunological mechanism(s): A critical discussion. Environ. Toxicol. Pharmacol. 2014, 37, 118–133. [Google Scholar] [CrossRef] [PubMed]
- Olinga, P.; Schuppan, D. Precision-cut liver slices: A tool to model the liver ex vivo. J. Hepatol. 2013, 58, 1252–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Collins, S.D.; Yuen, G.; Tu, T.; Budzinska, M.A.; Spring, K.J.; Bryant, K.; Shackel, N.A. In vitro models of the liver: Disease modeling, drug discovery and clinical applications. In Hepatocellular Carcinoma; Tirnitz-Parker, J.E.E., Ed.; Codon Publications: Singapore, 2019; pp. 47–67. [Google Scholar]
- Palma, E.; Doornebal, E.J.; Chokshi, S. Precision-cut liver slices: A versatile tool to advance liver research. Hepatol. Int. 2019, 13, 51–57. [Google Scholar] [CrossRef] [Green Version]
- Prasad, B.; Garg, A.; Takwani, H.; Singh, S. Metabolite identification by liquid chromatography-mass spectrometry. Trends Analytl. Chem. 2011, 30, 360–387. [Google Scholar] [CrossRef]
- Rothwell, J.; Sangster, N. Haemonchus contortus: The uptake and metabolism of closantel. Int. J. Parasitol. 1997, 27, 313–319. [Google Scholar] [CrossRef]
- Stuchlíková, L.R.; Matoušková, P.; Vokřál, I.; Lamka, J.; Szotáková, B.; Sečkařová, A.; Dimunová, D.; Nguyen, L.T.; Várady, M.; Skálová, L. Metabolism of albendazole, ricobendazole and flubendazole in Haemonchus contortus adults: Sex differences, resistance-related differences and the identification of new metabolites. Int. J. Parasitol. Drugs Drug Resist. 2018, 8, 50–58. [Google Scholar] [CrossRef]
- Matoušková, P.; Vokřál, I.; Lamka, J.; Skálová, L. The Role of Xenobiotic-Metabolizing Enzymes in Anthelmintic Deactivation and Resistance in Helminths. Trends Parasitol. 2016, 32, 481–491. [Google Scholar] [CrossRef]
- Zajíčková, M.; Prchal, L.; Navrátilová, M.; Vodvárková, N.; Matoušková, P.; Vokřál, I.; Nguyen, L.T.; Skálová, L. Sertraline as a new potential anthelmintic against Haemonchus contortus: Toxicity, efficacy, and biotransformation. Vet. Res. 2021, 52, 143. [Google Scholar] [CrossRef]
- A Van Wyk, J.; Gerber, H.M.; Groeneveld, H.T. A technique for the recovery of nematodes from ruminants by migration from gastro-intestinal ingesta gelled in agar: Large-scale application. Onderstepoort J. Veter-Res. 1980, 47, 147–158. [Google Scholar]
- Zárybnický, T.; Matoušková, P.; Lancošová, B.; Šubrt, Z.; Skálová, L.; Boušová, I. Inter-Individual Variability in Acute Toxicity of R-Pulegone and R-Menthofuran in Human Liver Slices and Their Influence on miRNA Expression Changes in Comparison to Acetaminophen. Int. J. Mol. Sci. 2018, 19, 1805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kotze, A.C.; McClure, S.J. Haemonchus contortus utilises catalase in defence against exogenous hydrogen peroxide in vitro. Int. J. Parasitol. 2001, 31, 1563–1571. [Google Scholar] [CrossRef]
- Holcapek, M.; Kolárová, L.; Nobilis, M. High-performance liquid chromatography-tandem mass spectrometry in the identification and determination of phase I and phase II drug metabolites. Anal. Bioanal. Chem. 2008, 391, 59–78. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vokřál, I.; Jirásko, R.; Stuchlíková, L.; Bártíková, H.; Szotáková, B.; Lamka, J.; Várady, M.; Skálová, L. Biotransformation of albendazole and activities of selected detoxification enzymes in Haemonchus contortus strains susceptible and resistant to anthelmintics. Vet. Parasitol. 2013, 196, 373–381. [Google Scholar] [CrossRef]
- Preston, S.; Jabbar, A.; Nowell, C.; Joachim, A.; Ruttkowski, B.; Baell, J.; Cardno, T.; Korhonen, P.K.; Piedrafita, D.; Ansell, B.R.E.; et al. Low cost whole-organism screening of compounds for anthelmintic activity. Int. J. Parasitol. 2015, 45, 333–343. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Zajíčková, M.; Mašátová, E.; Matoušková, P.; Skálová, L. The ATP bioluminescence assay: A new application and optimization for viability testing in the parasitic nematode Haemonchus contortus. Vet. Res. 2021, 52, 124. [Google Scholar] [CrossRef]
- Laing, R.; Bartley, D.J.; Morrison, A.A.; Rezansoff, A.; Martinelli, A.; Laing, S.T.; Gilleard, J.S. The cytochrome P450 family in the parasitic nematode Haemonchus contortus. Int. J. Parasitol. 2015, 45, 243–251. [Google Scholar] [CrossRef] [Green Version]
- Huckle, K.R.; Hutson, D.H.; Millburn, P. Species differences in the metabolism of 3-phenoxybenzoic acid. Drug Metab. Dispos. 1981, 9, 352. [Google Scholar]
- Parkinson, A. Biotransformation of xenobiotics. In Casarett and Doull’s Toxicology: The Basic Science of Poisons, 8th ed.; CD, K., Ed.; McGraw-Hill: New York, NY, USA, 2013; pp. 133–224. [Google Scholar]
- Lipinski, C.A.; Lombardo, F.; Dominy, B.W.; Feeney, P.J. Experimental and computational approaches to estimate solubility and permeability in drug discovery and development settings. Adv. Drug Deliv. Rev. 2001, 46, 3–26. [Google Scholar] [CrossRef]








| m/z Precursor Ions [M+H] + | tR LC-MS [min] | SRM [M+H] + | CE |
|---|---|---|---|
| 314 | 5.0 1; 8.9 2 | 190.95 | −11.0 |
| 90.95 | −25.0 | ||
| 121 | −26.0 | ||
| 322 | 4.8 1; 8.5 2 | 198 | −13 |
| 91 | −12 | ||
| 191 | −22 | ||
| 209 | 2.8 1 | 121 | −20 |
| 139 | −20 | ||
| 330 | 5.6 2 | 207 | −20 |
| 6.0 2 | 121 | −20 | |
| 6.2 2 | 91 | −20 | |
| 476 | 2.3 1 | 191 | −20 |
| 3.1 1 | 121 | −20 | |
| 91 | −20 |
| Compound | Elemental Composition | Designation | tR LC-HRMS [min] | m/z Precursor Ions [M+H]+ [M−H]− | m/z Product Ions [M+H]+ [M−H]− | CE | Occurrence |
|---|---|---|---|---|---|---|---|
| BLK127 | C19H23NO3 | - | 15.64 1; 14.22 2 | 314.1751 | 191.1063 | 20 | - |
| 121.0285 | |||||||
| 91.0547 | |||||||
| Internal standard | C20H19NO3 | IS | 15.24 1; 14.11 2 | 322.1431 | 199.0752 | 20 | - |
| 171.0802 | |||||||
| 127.0549 | |||||||
| 91.0548 | |||||||
| BLK127 hydrolysis | C12H16O3 | h-BLK127 | 13.971 | 209.1169 | 191.1064 | 20 | H. contortus |
| 139.0388 | |||||||
| 121.0284 | Ovine liver | ||||||
| 95.0496 | |||||||
| Hydroxylated BLK127 | C19H23NO4 | BLK127-OH | 11.24 1 | 330.1702 | 207.1015 | 25 | H. contortus |
| 11.78 1 | 121.0285 | ||||||
| 12.05 1 | 91.0548 | ||||||
| BLK127 N-glycosidation | C25H33NO8 | BLK127-N-glc | 10.99 1 | 476.2279 | 268.1176 | 25 | H. contortus |
| 12.37 1 | 232.0977 | ||||||
| 209.117 | |||||||
| 191.1065 | |||||||
| 121.0285 | |||||||
| 91.0548 | |||||||
| BLK127 O-glycosidation | C25H33NO9 | BLK127-O-glc | 9.20 1 | 492.2224 | - | 15; 25 | H. contortus |
| 9.60 1 | |||||||
| 9.83 1 | |||||||
| Hydrolysed BLK127-glycine | C14H19NO4 | h-BLK127-gly | 10.572 | 264.1246 | 220.1335 | 20 | Ovine liver |
| 93.03333 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zajíčková, M.; Prchal, L.; Vokřál, I.; Nguyen, L.T.; Kurz, T.; Gasser, R.; Bednářová, K.; Mičundová, M.; Lungerich, B.; Michel, O.; et al. Assessing the Anthelmintic Candidates BLK127 and HBK4 for Their Efficacy on Haemonchus contortus Adults and Eggs, and Their Hepatotoxicity and Biotransformation. Pharmaceutics 2022, 14, 754. https://doi.org/10.3390/pharmaceutics14040754
Zajíčková M, Prchal L, Vokřál I, Nguyen LT, Kurz T, Gasser R, Bednářová K, Mičundová M, Lungerich B, Michel O, et al. Assessing the Anthelmintic Candidates BLK127 and HBK4 for Their Efficacy on Haemonchus contortus Adults and Eggs, and Their Hepatotoxicity and Biotransformation. Pharmaceutics. 2022; 14(4):754. https://doi.org/10.3390/pharmaceutics14040754
Chicago/Turabian StyleZajíčková, Markéta, Lukáš Prchal, Ivan Vokřál, Linh Thuy Nguyen, Thomas Kurz, Robin Gasser, Klára Bednářová, Magdalena Mičundová, Beate Lungerich, Oliver Michel, and et al. 2022. "Assessing the Anthelmintic Candidates BLK127 and HBK4 for Their Efficacy on Haemonchus contortus Adults and Eggs, and Their Hepatotoxicity and Biotransformation" Pharmaceutics 14, no. 4: 754. https://doi.org/10.3390/pharmaceutics14040754