
Study of Viral Photoinactivation by UV-C Light and Photosensitizer Using a Pseudotyped Model
_Sadraeian.png)
 ,
,  ,
,  , , ,
, , ,
Abstract
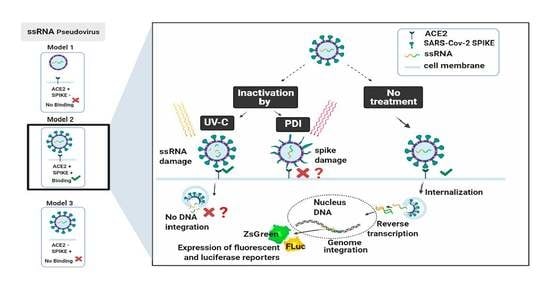
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Materials and Methods
2.1. Chemical Reagents
2.2. Cells and Viruses
2.3. Plasmids
2.4. UV-Vis Spectroscopy
2.5. Dynamic Light Scattering
2.6. Generation of Pseudovirus with SARS-CoV-2 Spike and Naked Control
2.7. Viral Inactivation Using UV-C Irradiation
2.8. Photosensitizer-Based Photodynamic Inactivation
2.9. Quantification of Viral RNA and Proviral DNA
2.10. Flow Cytometry
2.11. Luciferase Assay
2.12. Titration of Pseudovirus
2.13. Confocal Microscopy
2.14. Statistical Analyses
3. Results and Discussion
3.1. Generation of Pseudovirus with SARS-CoV-2 Spike and Naked Control
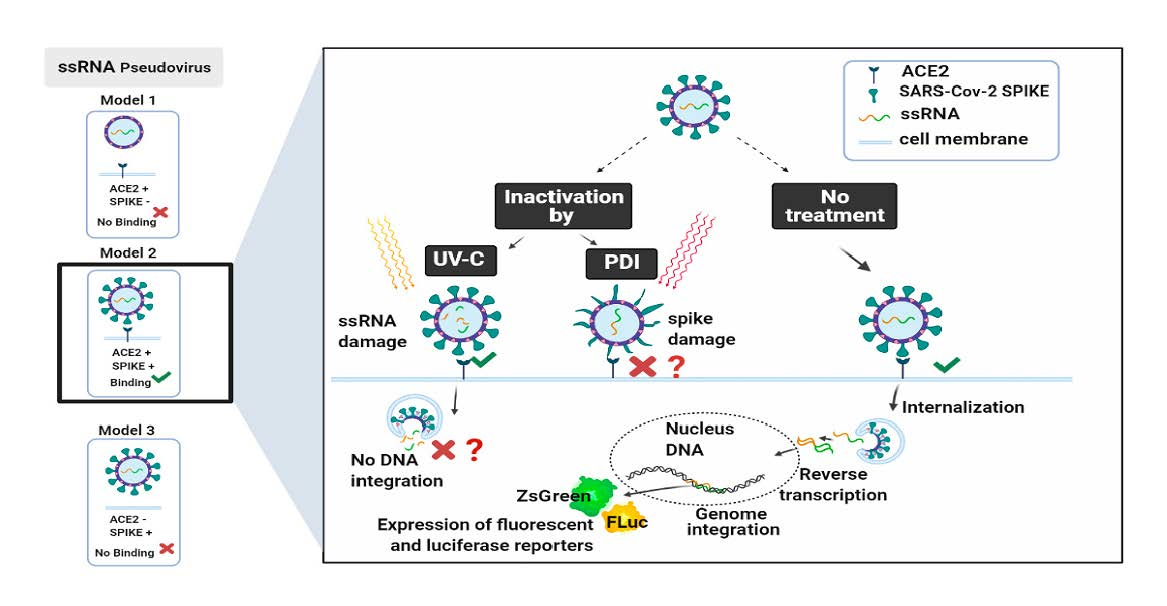
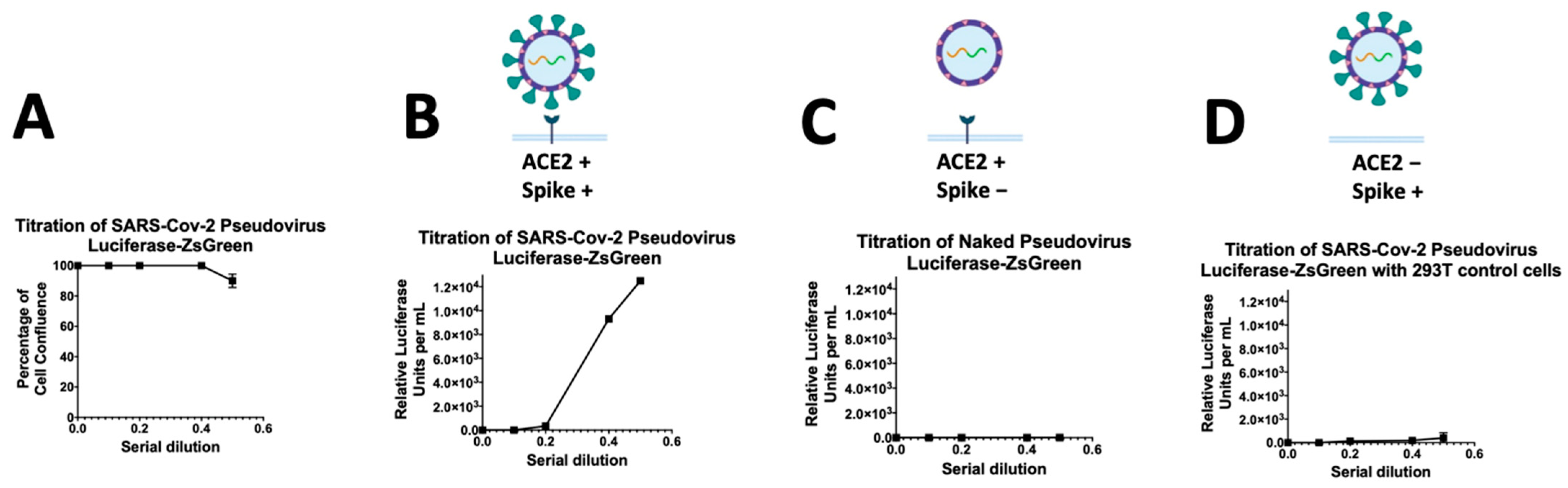
3.2. Titration of Pseudovirus
3.3. Viral Inactivation Using UV-C Irradiation or Photosensitizer-Based PDI
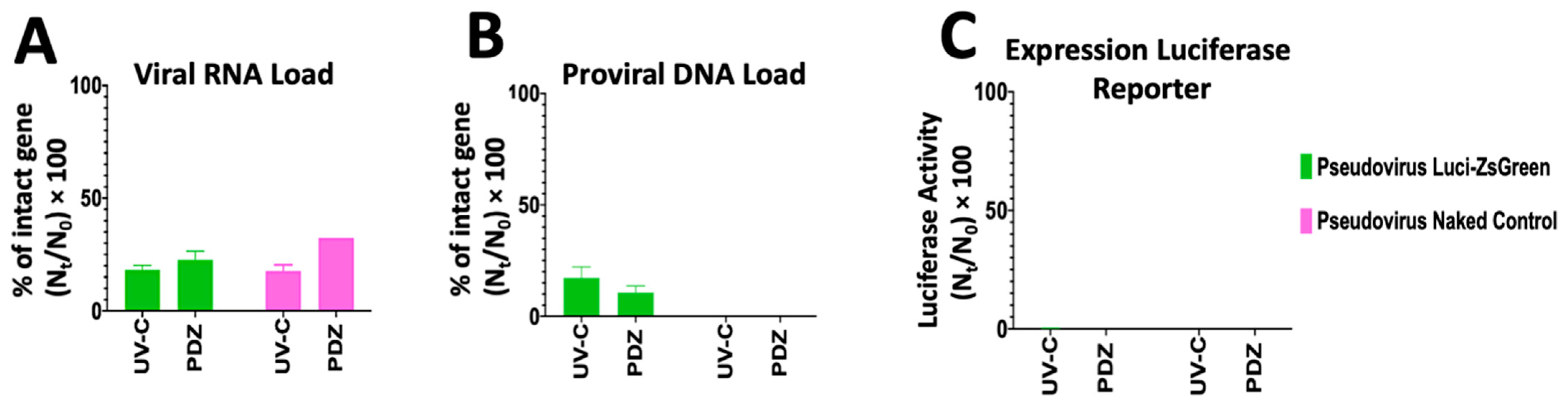
3.4. Study the Infectivity Mechanism of UV-C Irradiation and PDI Using qPCR
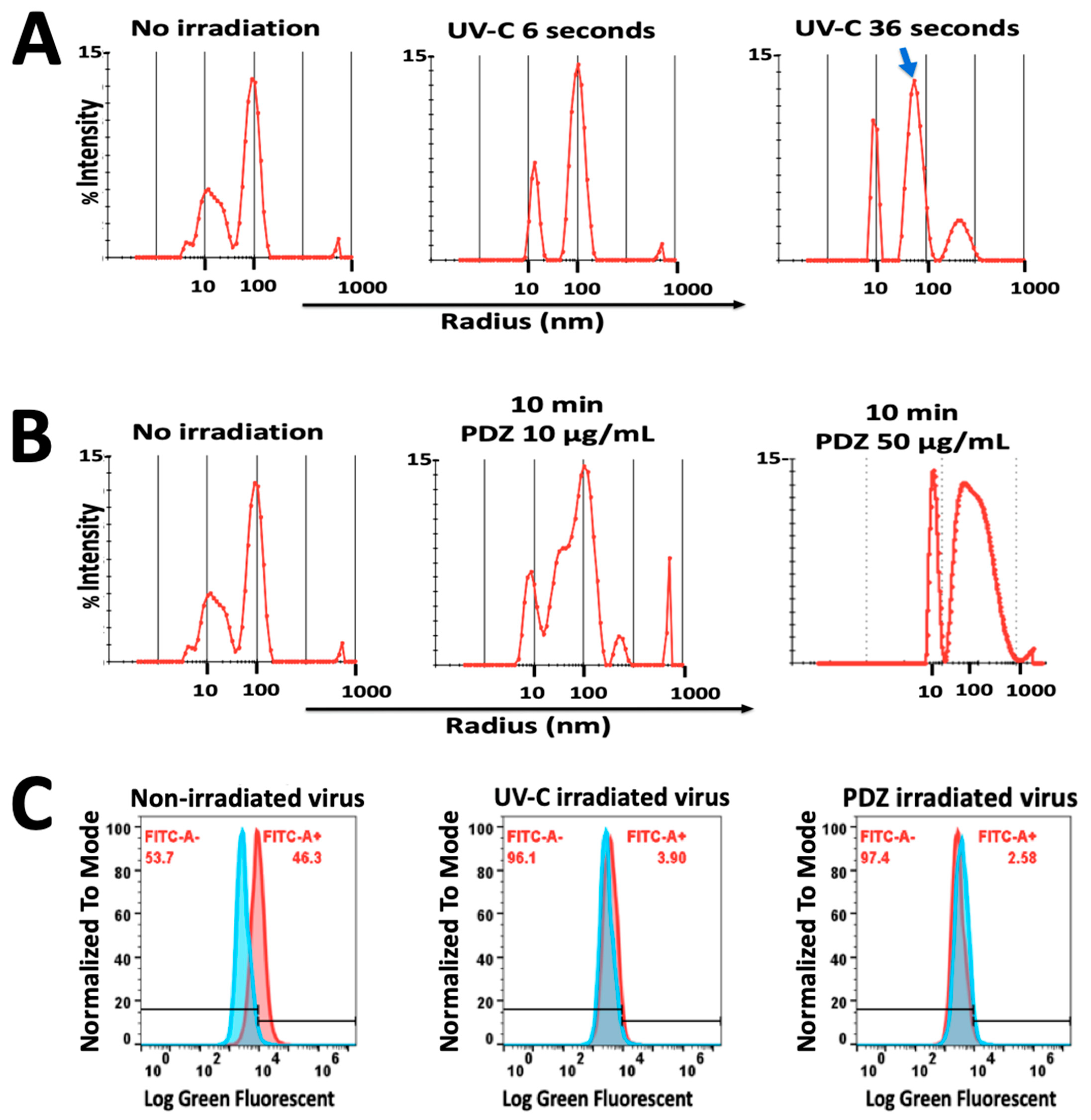
3.5. DLS Measurements before and after Irradiation
3.6. Green Fluorescent Measurement by Flow Cytometry
3.7. Observation of ZsGreen Expression by Confocal Microscopy
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| ACE-2 | Angiotensin Converting Enzyme 2 |
| DLS | Dynamic Light Scattering |
| PDI | Photodynamic Inactivation |
| PDT | Photodynamic Therapy |
| PDZ | Photoditazine |
| PS | Photosensitizer |
| RLU | Relative Luciferase Unit |
| SC2 | SARS-CoV-2 |
References
- World Health Organization. Coronavirus (COVID-19) Dashboard. Available online: https://covid19.who.int/ (accessed on 9 January 2022).
- Tran, H.D.M.; Boivin, S.; Kodamatani, H.; Ikehata, K.; Fujioka, T. Potential of UV-B and UV-C irradiation in disinfecting microorganisms and removing N-nitrosodimethylamine and 1,4-dioxane for potable water reuse: A review. Chemosphere 2021, 286, 131682. [Google Scholar] [CrossRef] [PubMed]
- Simonet, J.; Gantzer, C. Inactivation of poliovirus 1 and F-specific RNA phages and degradation of their genomes by UV irradiation at 254 nanometers. Appl. Environ. Microbiol. 2006, 72, 7671–7677. [Google Scholar] [CrossRef] [Green Version]
- Nogueira, M.S. Ultraviolet-based biophotonic technologies for control and prevention of COVID-19, SARS and related disorders. Photodiagn. Photodyn. Ther. 2020, 31, 101890. [Google Scholar] [CrossRef]
- Wallace, S.S.; Murphy, D.L.; Sweasy, J.B. Base excision repair and cancer. Cancer Lett. 2012, 327, 73–89. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hadi, J.; Dunowska, M.; Wu, S.; Brightwell, G. Control measures for SARS-CoV-2: A review on light-based inactivation of single-stranded rna viruses. Pathogens 2020, 9, 737. [Google Scholar] [CrossRef] [PubMed]
- Biasin, M.; Bianco, A.; Pareschi, G.; Cavalleri, A.; Cavatorta, C.; Fenizia, C.; Galli, P.; Lessio, L.; Lualdi, M.; Tombetti, E.; et al. UV-C irradiation is highly effective in inactivating SARS-CoV-2 replication. Sci. Rep. 2021, 11, 6260. [Google Scholar] [CrossRef] [PubMed]
- Lo, C.-W.; Matsuura, R.; Iimura, K.; Wada, S.; Shinjo, A.; Benno, Y.; Nakagawa, M.; Takei, M.; Aida, Y. UVC disinfects SARS-CoV-2 by induction of viral genome damage without apparent effects on viral morphology and proteins. Sci. Rep. 2021, 11, 13804. [Google Scholar] [CrossRef] [PubMed]
- Bekmukhametova, A.; Ruprai, H.; Hook, J.M.; Mawad, D.; Houang, J.; Lauto, A. Photodynamic therapy with nanoparticles to combat microbial infection and resistance. Nanoscale 2020, 12, 21034–21059. [Google Scholar] [CrossRef]
- Dobre, M.; Boscencu, R.; Neagoe, I.V.; Surcel, M.; Milanesi, E.; Manda, G. Insight into the web of stress responses triggered at gene expression level by porphyrin-pdt in ht29 human colon carcinoma cells. Pharmaceutics 2021, 13, 1032. [Google Scholar] [CrossRef]
- Higuchi, N.; Hayashi, J.-I.; Fujita, M.; Iwamura, Y.; Sasaki, Y.; Goto, R.; Ohno, T.; Nishida, E.; Yamamoto, G.; Kikuchi, T.; et al. Photodynamic Inactivation of an Endodontic Bacteria Using Diode Laser and Indocyanine Green-Loaded Nanosphere. Int. J. Mol. Sci. 2021, 22, 8384. [Google Scholar] [CrossRef]
- Zühlke, M.; Meiling, T.T.; Order, P.; Riebe, D.; Beitz, T.; Bald, I.; Löhmannsröben, H.-G.; Janßen, T.; Erhard, M.; Repp, A. Photodynamic Inactivation of E. coli Bacteria via Carbon Nanodots. ACS Omega 2021, 6, 23742–23749. [Google Scholar] [CrossRef] [PubMed]
- Sadraeian, M.; da Cruz, E.F.; Boyle, R.W.; Bahou, C.; Chudasama, V.; Janini, L.M.R.; Diaz, R.S.; Guimarães, F.E.G. Photoinduced Photosensitizer–Antibody Conjugates Kill HIV Env-Expressing Cells, Also Inactivating HIV. ACS Omega 2021, 6, 16524–16534. [Google Scholar] [CrossRef] [PubMed]
- Sadraeian, M.; Bahou, C.; Da Cruz, E.; Janini, L.; Diaz, R.S.; Boyle, R.; Chudasama, V.; Guimarães, F.E.G. Photoimmunotherapy Using Cationic and Anionic Photosensitizer-Antibody Conjugates against HIV Env-Expressing Cells. Int. J. Mol. Sci. 2020, 21, 9151. [Google Scholar] [CrossRef] [PubMed]
- Romano, R.A.; Pratavieira, S.; Da Silva, A.P.; Kurachi, C.; Guimarães, F.E.G. Light-driven photosensitizer uptake increases Candida albicans photodynamic inactivation. J. Biophotonics 2017, 10, 1538–1546. [Google Scholar] [CrossRef] [PubMed]
- Khorsandi, K.; Fekrazad, S.; Vahdatinia, F.; Farmany, A.; Fekrazad, R. Nano Antiviral Photodynamic Therapy: A Probable Biophysicochemical Management Modality in SARS-CoV-2. Expert Opin. Drug Deliv. 2021, 18, 265–272. [Google Scholar] [CrossRef]
- Almeida, A.; Faustino, M.A.F.; Neves, M.G.P.M.S. Antimicrobial Photodynamic Therapy in the Control of COVID-19. Antibiotics 2020, 9, 320. [Google Scholar] [CrossRef]
- Pires, L.; Bosco, S.D.M.G.; Baptista, M.S.; Kurachi, C. Photodynamic therapy in pythium insidiosum—An in vitro study of the correlation of sensitizer localization and cell death. PLoS ONE 2014, 9, e85431. [Google Scholar] [CrossRef]
- Sharshov, K.; Solomatina, M.; Kurskaya, O.; Kovalenko, I.; Kholina, E.; Fedorov, V.; Meerovich, G.; Rubin, A.; Strakhovskaya, M. The Photosensitizer Octakis(cholinyl)zinc Phthalocyanine with Ability to Bind to a Model Spike Protein Leads to a Loss of SARS-CoV-2 Infectivity In Vitro When Exposed to Far-Red LED. Viruses 2021, 13, 643. [Google Scholar] [CrossRef]
- Sabino, C.P.; Sellera, F.P.; Sales-Medina, D.F.; Machado, R.R.G.; Durigon, E.L.; Freitas-Junior, L.H.; Ribeiro, M.S. UV-C (254 nm) lethal doses for SARS-CoV-2. Photodiagn. Photodyn. Ther. 2020, 32, 101995. Available online: https://linkinghub.elsevier.com/retrieve/pii/S1572100020303495 (accessed on 14 March 2022). [CrossRef]
- Ou, X.; Liu, Y.; Lei, X.; Li, P.; Mi, D.; Ren, L.; Guo, L.; Guo, R.; Chen, T.; Hu, J.; et al. Characterization of spike glycoprotein of SARS-CoV-2 on virus entry and its immune cross-reactivity with SARS-CoV. Nat. Commun. 2020, 11, 1620. [Google Scholar] [CrossRef] [Green Version]
- Lam, B.; Kung, Y.J.; Lin, J.; Tseng, S.-H.; Tsai, Y.C.; He, L.; Castiglione, G.; Egbert, E.; Duh, E.J.; Bloch, E.M.; et al. In vivo characterization of emerging SARS-CoV-2 variant infectivity and human antibody escape potential. Cell Rep. 2021, 37, 109838. [Google Scholar] [CrossRef]
- Crawford, K.H.D.; Eguia, R.; Dingens, A.S.; Loes, A.N.; Malone, K.D.; Wolf, C.R.; Chu, H.Y.; Tortorici, M.A.; Veesler, D.; Murphy, M.; et al. Protocol and Reagents for Pseudotyping Lentiviral Particles with SARS-CoV-2 Spike Protein for Neutralization Assays. Viruses 2020, 12, 513. [Google Scholar] [CrossRef] [PubMed]
- Robinson, E.; Nunes, J.P.M.; Vassileva, V.; Maruani, A.; Nogueira, J.C.F.; Smith, M.E.B.; Pedley, R.B.; Caddick, S.; Baker, J.R.; Chudasama, V. Pyridazinediones deliver potent, stable, targeted and efficacious antibody–drug conjugates (ADCs) with a controlled loading of 4 drugs per antibody. RSC Adv. 2017, 7, 9073–9077. [Google Scholar] [CrossRef] [Green Version]
- Marinho, R.D.S.S.; Duro, R.L.S.; Santos, G.L.; Hunter, J.; Teles, M.D.A.R.; Brustulin, R.; Milagres, F.A.D.P.; Sabino, E.C.; Diaz, R.S.; Komninakis, S.V. Detection of coinfection with Chikungunya virus and Dengue virus serotype 2 in serum samples of patients in State of Tocantins, Brazil. J. Infect. Public Health 2020, 13, 724–729. [Google Scholar] [CrossRef]
- Sucupira, M.C.A.; Sanabani, S.; Cortes, R.M.; Giret, M.T.M.; Tomiyama, H.; Sauer, M.M.; Sabino, E.C.; Janini, L.M.; Kallas, E.G.; Diaz, R.S. Faster HIV-1 Disease Progression among Brazilian Individuals Recently Infected with CXCR4-Utilizing Strains. PLoS ONE 2012, 7, e30292. [Google Scholar]
- Kumar, A.M.; Fernandez, J.B.; Singer, E.J.; Commins, D.; Waldrop-Valverde, D.; Ownby, R.L.; Kumar, M. Human immunodeficiency virus type 1 in the central nervous system leads to decreased dopamine in different regions of postmortem human brains. J. Neuro Virol. 2009, 15, 257–274. [Google Scholar] [CrossRef] [PubMed]
- Komninakis, S.V.; Santos, D.E.M.; Santos, C.; Oliveros, M.P.R.; Sanabani, S.; Diaz, R.S. HIV-1 Proviral DNA Loads (as Determined by Quantitative PCR) in Patients Subjected to Structured Treatment Interruption after Antiretroviral Therapy Failure. J. Clin. Microbiol. 2012, 50, 2132–2133. [Google Scholar] [CrossRef] [Green Version]
- Hu, J.; Gao, Q.; He, C.; Huang, A.; Tang, N.; Wang, K. Development of cell-based pseudovirus entry assay to identify potential viral entry inhibitors and neutralizing antibodies against SARS-CoV-2. Genes Dis. 2020, 7, 551–557. [Google Scholar] [CrossRef]
- Svyatchenko, V.A.; Nikonov, S.D.; Mayorov, A.P.; Gelfond, M.L.; Loktev, V.B. Antiviral photodynamic therapy: Inactivation and inhibition of SARS-CoV-2 in vitro using methylene blue and Radachlorin. Photodiagn. Photodyn. Ther. 2021, 33, 102112. [Google Scholar] [CrossRef]
- Alves, F.; Ayala, E.T.P.; Pratavieira, S. Sonophotodynamic Inactivation: The power of light and ultrasound in the battle against microorganisms. J. Photochem. Photobiol. 2021, 7, 100039. [Google Scholar] [CrossRef]
- Sabino, C.P.; Wainwright, M.; dos Anjos, C.; Sellera, F.; Baptista, M.S.; Lincopan, N.; Ribeiro, M. Inactivation kinetics and lethal dose analysis of antimicrobial blue light and photodynamic therapy. Photodiagn. Photodyn. Ther. 2019, 28, 186–191. [Google Scholar] [CrossRef]
- Case, J.B.; Rothlauf, P.W.; Chen, R.E.; Liu, Z.; Zhao, H.; Kim, A.S.; Bloyet, L.-M.; Zeng, Q.; Tahan, S.; Droit, L.; et al. Neutralizing Antibody and Soluble ACE2 Inhibition of a Replication-Competent VSV-SARS-CoV-2 and a Clinical Isolate of SARS-CoV-2. Cell Host Microbe 2020, 28, 475–485.e5. [Google Scholar] [CrossRef] [PubMed]
- Kumar, U.S.; Afjei, R.; Ferrara, K.; Massoud, T.F.; Paulmurugan, R. Gold-Nanostar-Chitosan-Mediated Delivery of SARS-CoV-2 DNA Vaccine for Respiratory Mucosal Immunization: Development and Proof-of-Principle. ACS Nano 2021, 15, 17582–17601. [Google Scholar] [CrossRef] [PubMed]
- Duguay, B.A.; Herod, A.; Pringle, E.S.; Monro, S.M.A.; Hetu, M.; Cameron, C.G.; McFarland, S.A.; McCormick, C. Photodynamic Inactivation of Human Coronaviruses. Viruses 2022, 14, 110. [Google Scholar] [CrossRef]
- Arentz, J.; von der Heide, H.-J. Evaluation of methylene blue based photodynamic inactivation (PDI) against intracellular B-CoV and SARS-CoV2 viruses under different light sources in vitro as a basis for new local treatment strategies in the early phase of a Covid19 infection. Photodiagn. Photodyn. Ther. 2022, 37, 102642. Available online: https://linkinghub.elsevier.com/retrieve/pii/S1572100021004609 (accessed on 14 March 2022). [CrossRef]
- Kipshidze, N.; Yeo, N.; Kipshidze, N. Photodynamic therapy for COVID-19. Nat. Photon. 2020, 14, 651–652. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sadraeian, M.; Junior, F.F.P.; Miranda, M.; Galinskas, J.; Fernandes, R.S.; da Cruz, E.F.; Fu, L.; Zhang, L.; Diaz, R.S.; Cabral-Miranda, G.; et al. Study of Viral Photoinactivation by UV-C Light and Photosensitizer Using a Pseudotyped Model. Pharmaceutics 2022, 14, 683. https://doi.org/10.3390/pharmaceutics14030683
Sadraeian M, Junior FFP, Miranda M, Galinskas J, Fernandes RS, da Cruz EF, Fu L, Zhang L, Diaz RS, Cabral-Miranda G, et al. Study of Viral Photoinactivation by UV-C Light and Photosensitizer Using a Pseudotyped Model. Pharmaceutics. 2022; 14(3):683. https://doi.org/10.3390/pharmaceutics14030683
Chicago/Turabian StyleSadraeian, Mohammad, Fabio Francisco Pinto Junior, Marcela Miranda, Juliana Galinskas, Rafaela Sachetto Fernandes, Edgar Ferreira da Cruz, Libing Fu, Le Zhang, Ricardo Sobhie Diaz, Gustavo Cabral-Miranda, and et al. 2022. "Study of Viral Photoinactivation by UV-C Light and Photosensitizer Using a Pseudotyped Model" Pharmaceutics 14, no. 3: 683. https://doi.org/10.3390/pharmaceutics14030683