
Hepatoprotective Effect of Carob Pulp Flour (Ceratonia siliqua L.) Extract Obtained by Optimized Microwave-Assisted Extraction

 , , ,
, , ,  , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Chemicals
2.3. Microwave-Assisted Extraction
2.4. Antioxidant Activity
2.4.1. DPPH Assay
2.4.2. FRAP Assay
2.4.3. ABTS Assay
2.5. Experimental Design and Statistical Analysis
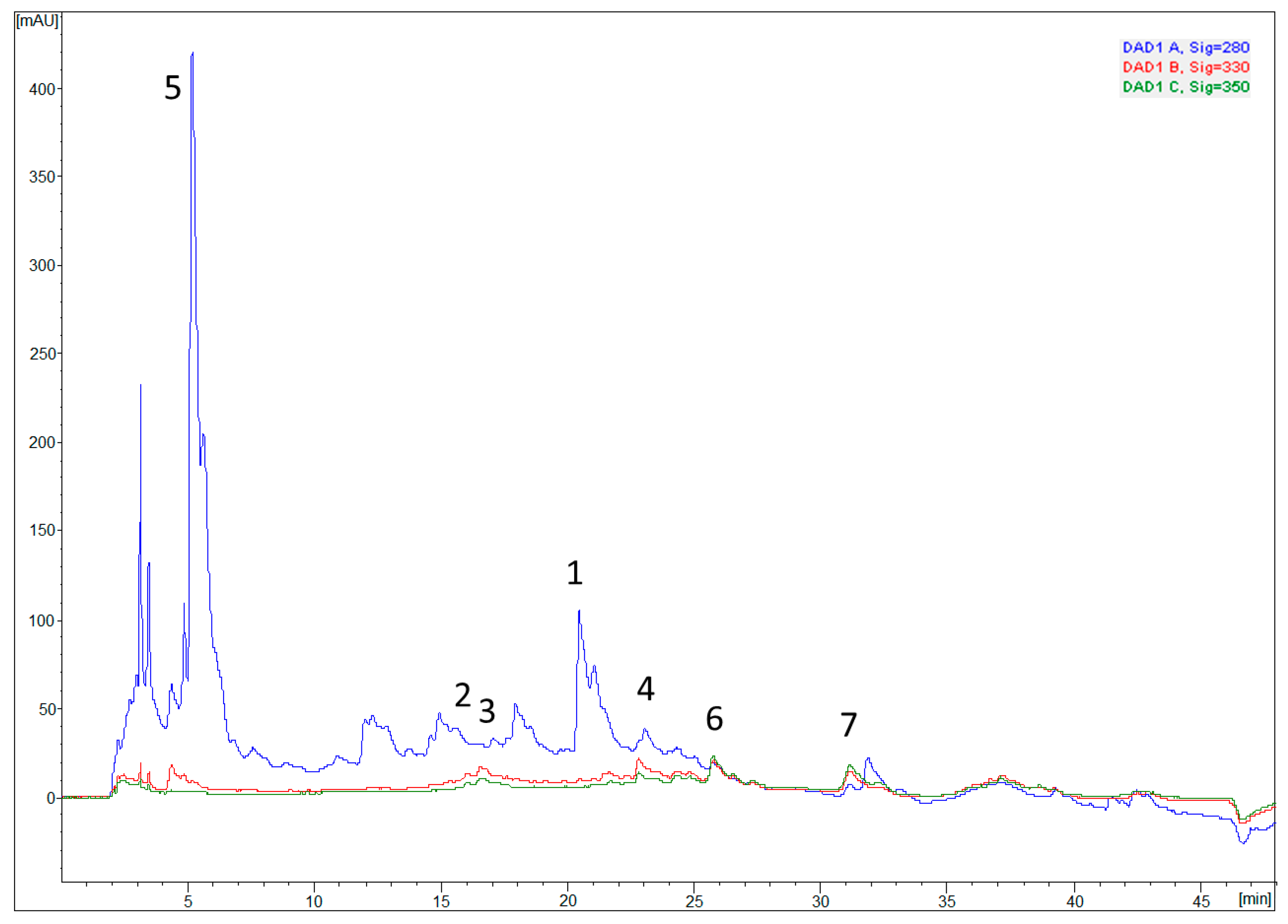
2.6. Chemical Characterization—Phenolic Profile
2.7. Experimental Animals and Ethical Statement
2.8. In Vivo Experimental Design
- ConS—saline 1 mL/kg, seven days p.o.;
- ConP—saline 1 mL/kg seven days p.o. +1 single dose of paracetamol 110 mg/kg p.o.;
- CE100—carob extract 100 mg/kg for seven days p.o.;
- CE100 + P—carob extract 100 mg/kg for seven days p.o. +1 single dose of paracetamol 110 mg/kg p.o.;
- CE200—carob extract 200 mg/kg for seven days p.o.;
- CE200 + P—carob extract 200 mg/kg for seven days p.o. +1 single dose of paracetamol 110 mg/kg p.o.
2.9. Liver Function Tests
2.10. Determination of In Vivo Antioxidant Activity
2.11. Histopathology and Immunohistochemistry Assessment
3. Results and Discussion
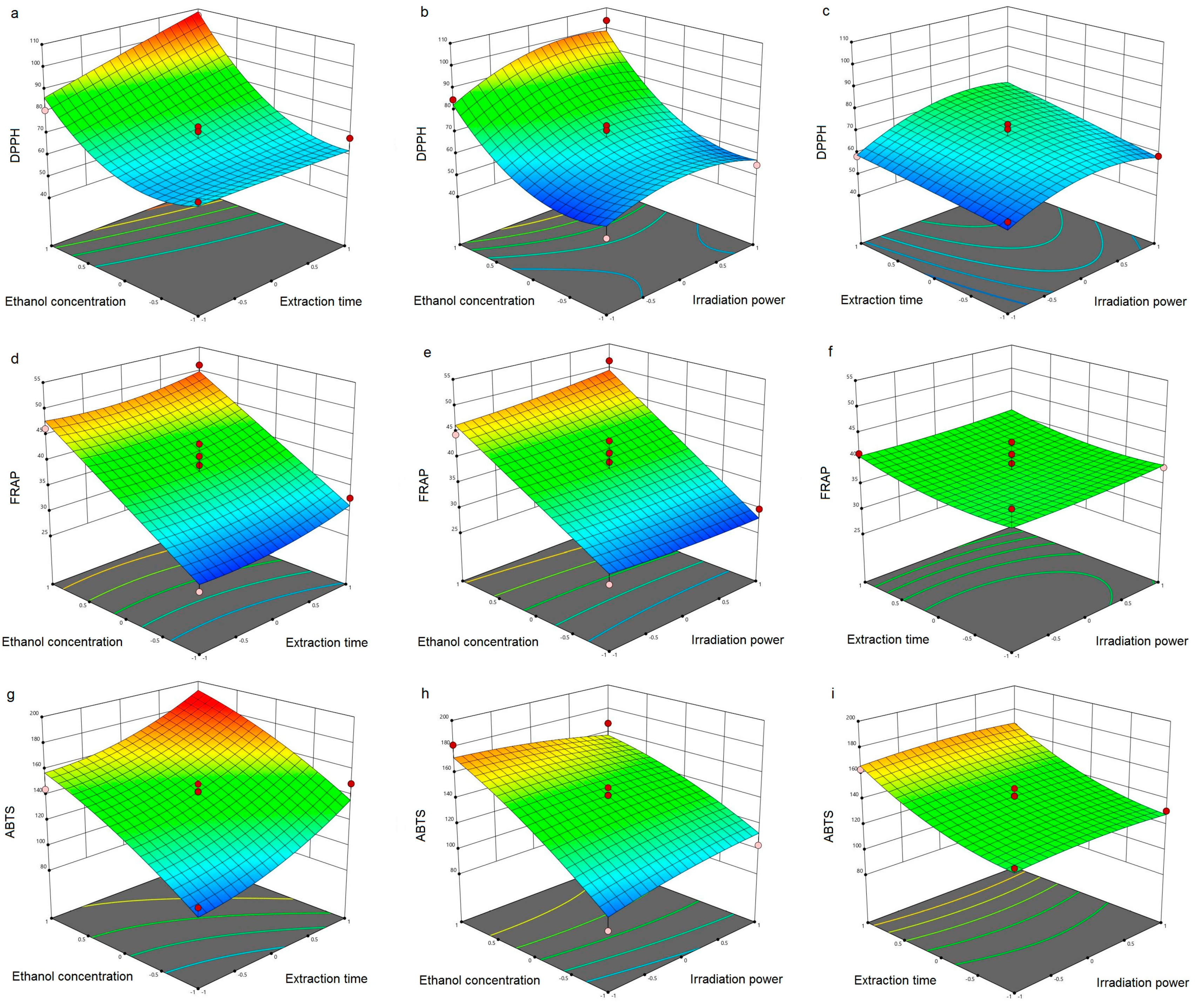
3.1. Model Adequacy
3.2. Antioxidant Activity of Carob Flour Extracts
3.3. Optimization of MAE Process
3.4. Chemical Characterization—Determination of Phenolic Profile
3.5. Effects of Carob Flour Extract on Paracetamol-Induced Serum Biochemical Parameters
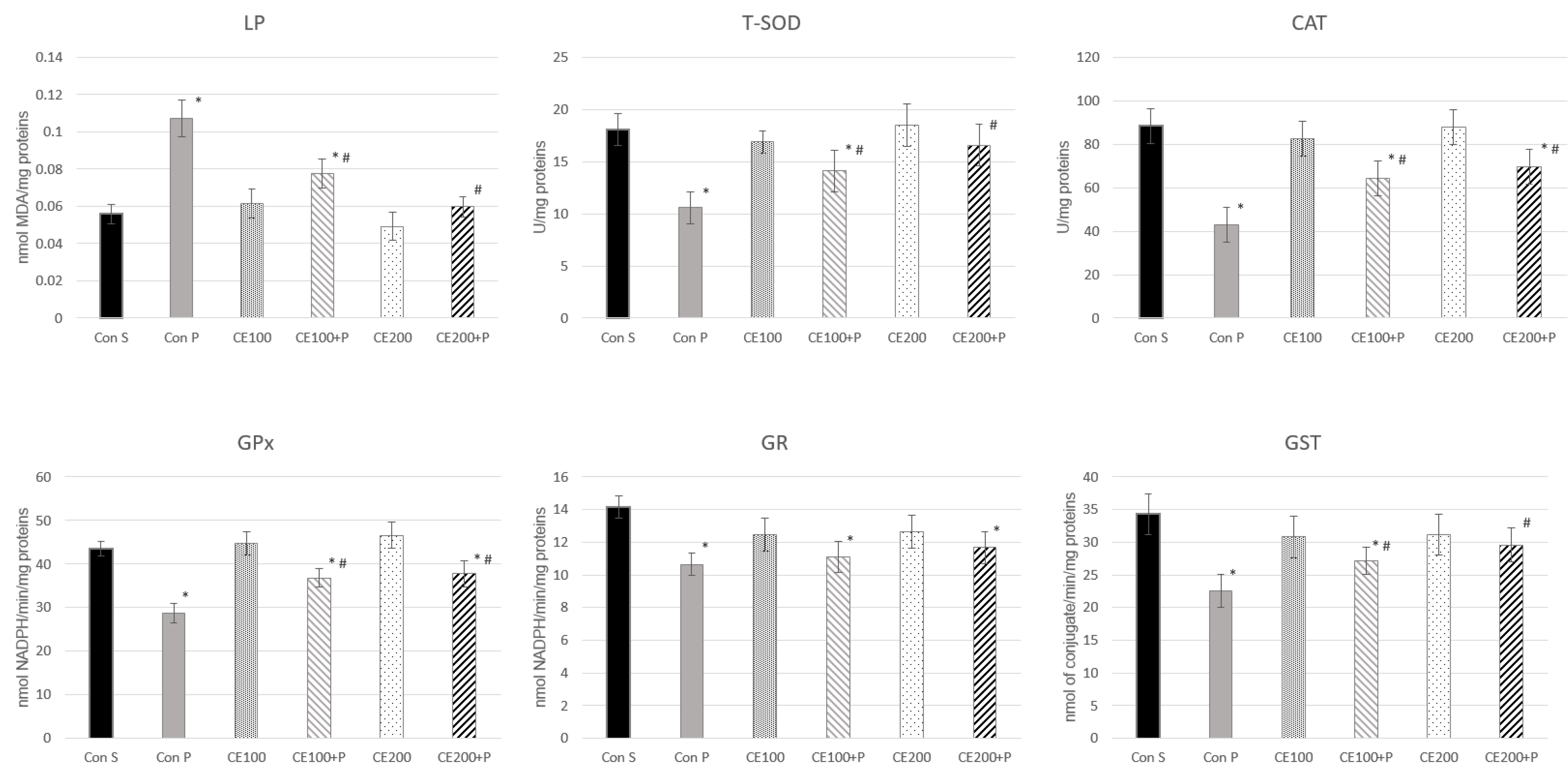
3.6. Effects of Carob Flour Extract on Paracetamol-Induced Oxidative Stress
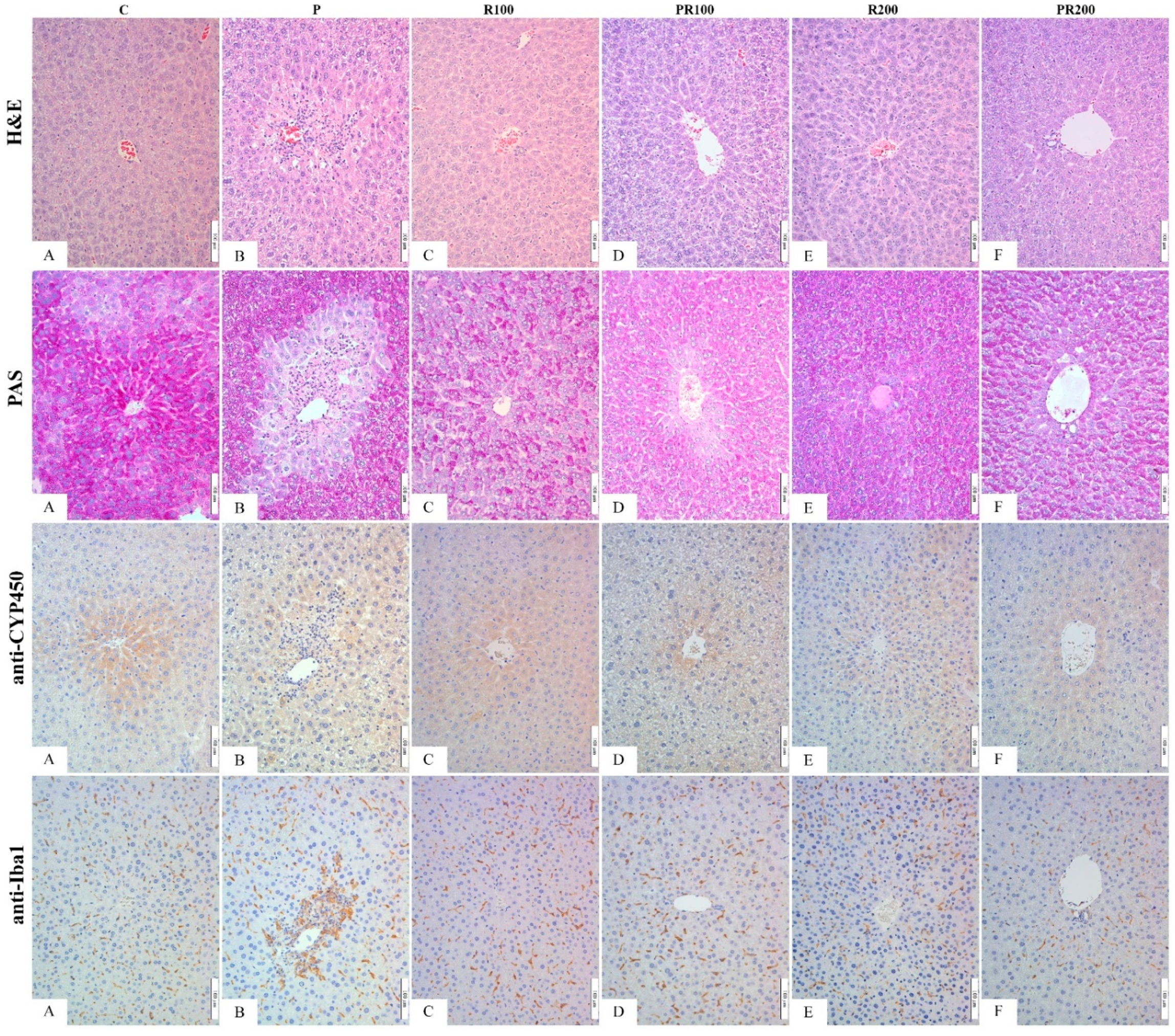
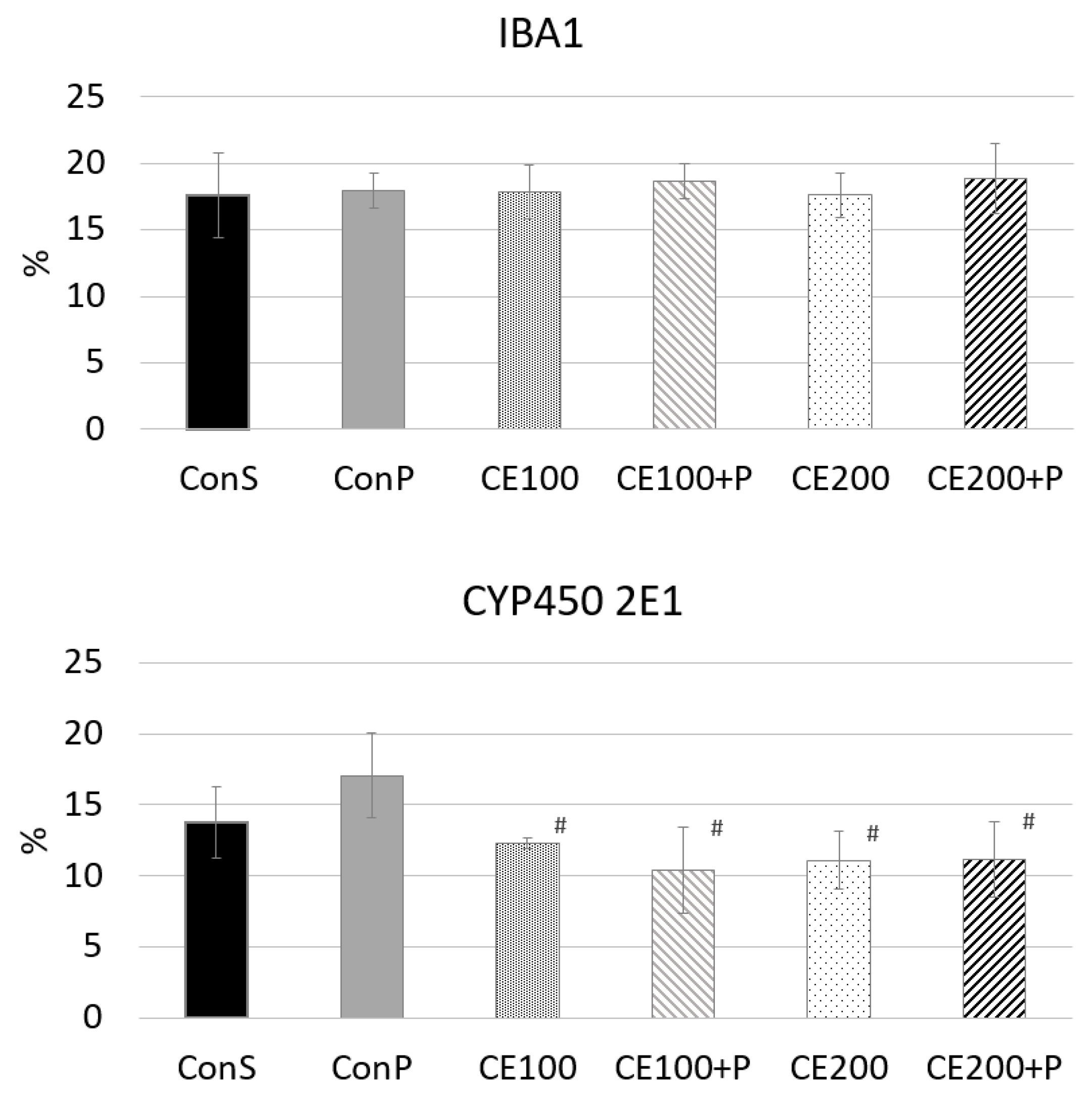
3.7. Histological, Immunohistochemical and Morphometric Analysis of Liver Tissue
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Brand, E. Carob. Nutr. Food Sci. 1984, 91, 22–24. [Google Scholar]
- Catarino, F. The carob tree. An exemplary plant. Naturopa 1993, 73, 14–15. [Google Scholar]
- Barak, S.; Mudgil, D. Locust bean gum: Processing, properties and food applications—A review. Int. J. Biol. Macromol. 2014, 66, 74–80. [Google Scholar] [CrossRef]
- Karabulut, A.; Canbolat, O.; Kamalak, A. Evaluation of carob, Ceratonia siliqua pods as a feed for sheep. Livest. Res. Rural Dev. 2006, 18, 104. Available online: https://www.lrrd.org/lrrd18/7/kara18104.htm. (accessed on 26 September 2021).
- Nasar-Abbas, S.M.; e-Huma, Z.; Vu, T.-H.; Khan, M.K.; Esbenshade, H.; Jayasena, V. Carob kibble: A bioactive-rich food ingredient. Compr. Rev. Food Sci. Food Saf. 2016, 15, 63–72. [Google Scholar] [CrossRef] [Green Version]
- Goulas, V.; Stylos, E.; Chatziathanasiadou, M.V.; Mavromoustakos, T.; Tzakos, A.G. Functional components of carob fruit: Linking the chemical and biological space. Int. J. Mol. Sci. 2016, 17, 1875. [Google Scholar] [CrossRef]
- Galanakis, C.M. Phenols recovered from olive mill wastewater as additives in meat production. Trends Food Sci. Technol. 2018, 79, 98–105. [Google Scholar] [CrossRef]
- Sakakibara, H.; Honda, Y.; Nakagawa, S.; Ashida, H.; Kanazawa, K. Simultaneous determination of all polyphenols in vegetables, fruits, and teas. J. Agric. Food Chem. 2003, 51, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Youssef, M.K.E.; El-Manfaloty, M.M.; Ali, H.M. Assessment of proximate chemical composition, nutritional status, fatty acid composition and phenolic compounds of carob (Ceratonia siliqua L.). Food Public Health 2013, 3, 304–308. [Google Scholar] [CrossRef]
- Almanasrah, M.; Roseiro, L.B.; Bogel-Lukasik, R.; Carvalheiro, F.; Brazinha, C.; Crespo, J.; Kallioinen, M.; Mänttäri, M.; Duarte, L.C. Selective recovery of phenolic compounds and carbohydrates from carob kibbles using water-based extraction. Ind. Crops Prod. 2015, 70, 443–450. [Google Scholar] [CrossRef] [Green Version]
- Rtibi, K.; Selmi, S.; Grami, D.; Amri, M.; Eto, B.; El-benna, J.; Sebai, H.; Marzouki, L. Chemical constituents and pharmacological actions of carob pods and leaves (Ceratonia siliqua L.) on the gastrointestinal tract: A review. Biomed. Pharmacother. 2017, 93, 522–528. [Google Scholar] [CrossRef]
- Roseiro, L.B.; Tavares, C.S.; Roseiro, J.C.; Rauter, A.P. Antioxidants from aqueous decoction of carob pods biomass (Ceretonia siliqua L.): Optimisation using response surface methodology and phenolic profile by capillary electrophoresis. Ind. Crops Prod. 2013, 44, 119–126. [Google Scholar] [CrossRef]
- Stavrou, I.J.; Christou, A.; Kapnissi-Christodoulou, C.P. Polyphenols in carobs: A review on their composition, antioxidant capacity and cytotoxic effects, and health impact. Food Chem. 2018, 269, 355–374. [Google Scholar] [CrossRef]
- Chan, C.-H.; Yusoff, R.; Ngoh, G.-C.; Kung, F.W.-L. Microwave-assisted extractions of active ingredients from plants. J. Chromatogr. A 2011, 1218, 6213–6225. [Google Scholar] [CrossRef]
- Routray, W.; Orsat, V. Microwave-assisted extraction of flavonoids: A review. Food Bioproc. Technol. 2012, 5, 409–424. [Google Scholar] [CrossRef]
- Baş, D.; Boyaci, İ.H. Modeling and optimization I: Usability of response surface methodology. J. Food Eng. 2007, 78, 836–845. [Google Scholar] [CrossRef]
- Zeković, Z.; Vladić, J.; Vidović, S.E.; Adamović, D.; Pavlić, B. Optimization of microwave-assisted extraction (MAE) of coriander phenolic antioxidants—response surface methodology approach. J. Sci. Food Agric. 2016, 96, 4613–4622. [Google Scholar] [CrossRef]
- Huma, Z.E.; Jayasena, V.; Nasar-Abbas, S.M.; Imran, M.; Khan, M.K. Process optimization of polyphenol extraction from carob (Ceratonia siliqua) kibbles using microwave-assisted technique. J. Food Process. Preserv. 2018, 42, e13450. [Google Scholar] [CrossRef]
- Quiles-Carrillo, L.; Mellinas, C.; Garrigos, M.C.; Balart, R.; Torres-Giner, S. Optimization of microwave-assisted extraction of phenolic compounds with antioxidant activity from carob pods. Food Anal. Methods 2019, 12, 2480–2490. [Google Scholar] [CrossRef]
- Teslić, N.; Bojanić, N.; Rakić, D.; Takači, A.; Zeković, Z.; Fišteš, A.; Bodroža-Solarov, M.; Pavlić, B. Defatted wheat germ as source of polyphenols—Optimization of microwave-assisted extraction by RSM and ANN approach. Chem. Eng. Process. 2019, 143, 107634. [Google Scholar] [CrossRef]
- Al-Olayan, E.M.; El-Khadragy, M.F.; Alajmi, R.A.; Othman, M.S.; Bauomy, A.A.; Ibrahim, S.R.; Moneim, A.E. Ceratonia siliqua pod extract ameliorates Schistosoma mansoni-induced liver fibrosis and oxidative stress. BMC Complement. Altern. Med. 2016, 16, 434. [Google Scholar] [CrossRef] [Green Version]
- Rtibi, K.; Selmi, S.; Jabri, M.A.; El-Benna, J.; Amri, M.; Marzouki, L.; Sebai, H. Protective effect of Ceratonia siliqua L. against a dextran sulfate sodium-induced alterations in liver and kidney in rat. J. Med. Food 2016, 19, 882–889. [Google Scholar] [CrossRef]
- Espín, J.C.; Soler-Rivas, C.; Wichers, H.J. Characterization of the total free radical scavenger capacity of vegetable oils and oil fractions using 2,2-diphenyl-1-picrylhydrazyl radical. J. Agric. Food Chem. 2000, 48, 648–656. [Google Scholar] [CrossRef]
- Oyaizu, M. Studies on products of browning reaction. Antioxidative activities of products of browning reaction prepared from glucosamine. Jpn. J. Nutr. Diet. 1986, 44, 307–315. [Google Scholar] [CrossRef] [Green Version]
- Re, R.; Pellegrini, N.; Proteggenete, A.; Pannala, A.; Yang, M.; Rice-Evans, C. Antioxidant activity applying an improved ABTS radical cation decolorization assay. Free Radic. Biol. Med. 1999, 26, 1231–1237. [Google Scholar] [CrossRef]
- Derringer, G. Simultaneous optimization of several response variables. J. Qual. Technol. 1980, 12, 214–219. [Google Scholar] [CrossRef]
- Salaj, N.; Kladar, N.; Srđenović, Č.B.; Jeremić, K.; Barjaktarović, J.; Hitl, M.; Gavarić, N.; Božin, B. Stabilization of sunflower and olive oils with savory (Satureja kitaibelii, Lamiaceae). J. Food Nutr. Res. 2020, 59, 259–271. [Google Scholar]
- Rašković, A.; Ćućuz, V.; Torović, L.; Tomas, A.; Gojković-Bukarica, L.; Ćebović, T.; Milijašević, B.; Stilinović, N.; Hogervorst, J.C. Resveratrol supplementation improves metabolic control in rats with induced hyperlipidemia and type 2 diabetes. Saudi Pharm. J. 2019, 27, 1036–1043. [Google Scholar] [CrossRef]
- Buege, J.A.; Aust, S.D. Microsomal lipid peroxidation. Meth. Enzymol. 1978, 52, 302–310. [Google Scholar] [CrossRef]
- McCord, J.M.; Fridovich, I. Superoxide dismutase: An enzymic function for erythrocuprein (hemocuprein). J. Biol. Chem. 1969, 244, 6049–6055. [Google Scholar] [CrossRef]
- Liyana-Pathirana, C.; Shahidi, F. Optimization of extraction of phenolic compounds from wheat using response surface methodology. Food Chem. 2005, 93, 47–56. [Google Scholar] [CrossRef]
- Beers, R.F.; Sizer, I.W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Beutler, E. Red Cell Metabolism: A Manual of Biochemical Methods, 3rd ed.; Grune & Stratton Inc.: New York, NY, USA, 1984. [Google Scholar]
- Goldberg, D.M.; Spooner, R.J. Assay of Glutathione Reductase. In Methods of Enzymatic Analysis, 3rd ed.; Bergmeyen, H.V., Ed.; Verlog Chemie: Deerfiled Beach, FL, USA, 1983; Volume 3, pp. 258–265. [Google Scholar]
- Sharma, R. Polyphenols in health and disease: Practice and mechanisms of benefits. In Polyphenols in Human Health and Disease; Watson, R.R., Preedy, V.R., Zibadi, S., Eds.; Academic Press: Cambridge, MA, USA, 2014; Volume 1, pp. 757–778. [Google Scholar]
- Roseiro, L.B.; Duarte, L.C.; Oliviera, D.L.; Roque, R.; Bernardo-Gil, M.G.; Martins, A.I.; Sepúlveda, C.; Almeida, J.; Meireles, M.; Gírio, F.M.; et al. Supercritical, ultrasound and conventional extracts from carob (Ceratonia siliqua L.) biomass: Effect on the phenolic profile and antiproliferative activity. Ind. Crops Prod. 2013, 47, 132–138. [Google Scholar] [CrossRef]
- Bernardo-Gil, M.G.; Roque, R.; Roseiro, L.B.; Duarte, L.C.; Gírio, F.; Esteves, P. Supercritical extraction of carob kibbles (Ceratonia siliqua L.). J. Supercrit. Fluids 2011, 59, 36–42. [Google Scholar] [CrossRef]
- Goulas, V.; Georgiou, E. Utilization of carob fruit as sources of phenolic compounds with antioxidant potential: Extraction optimization and application in food models. Foods 2020, 9, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rašković, A.; Bukumirović, N.; Paut Kusturica, M.; Milić, N.; Čabarkapa, V.; Borišev, I.; Čapo, I.; Miljković, D.; Stilinović, N.; Mikov, M. Hepatoprotective and antioxidant potential of Pycnogenol® in acetaminophen-induced hepatotoxicity in rats. Phytother. Res. 2019, 33, 631–639. [Google Scholar] [CrossRef]
- Ahmed, M.M. Biochemical studies on nephroprotective effect of carob (Ceratonia siliqua L.) growing in Egypt. Nat. Sci. 2010, 8, 41–47. [Google Scholar]
- Suzek, H.; Celik, I.; Dogan, A. Nephroprotective hepatoprotective potential and antioxidant role of carob pods (Cerotonia siliqua L.) against carbon tetrachloride-induced toxicity in rats. Indian J. Pharm. Educ. Res. 2017, 51, 312–320. [Google Scholar] [CrossRef] [Green Version]
- Stilinović, N.; Čapo, I.; Vukmirović, S.; Rašković, A.; Tomas, A.; Popović, M.; Sabo, A. Chemical composition, nutritional profile and in vivo antioxidant properties of the cultivated mushroom Coprinus comatus. R. Soc. Open Sci. 2020, 7, 200900. [Google Scholar] [CrossRef]
- Hsouna, A.B.; Saoudi, M.; Trigui, M.; Jamoussi, K.; Boudawara, T.; Jaoua, S.; El Feki, A. Characterization of bioactive compounds and ameliorative effects of Ceratonia siliqua leaf extract against CCl4 induced hepatic oxidative damage and renal failure in rats. Food Chem. Toxicol. 2011, 49, 3183–3191. [Google Scholar] [CrossRef]
- Souli, A.; Sebai, H.; Chehimi, L.; Rtibi, K.; Tounsi, H.; Boubaker, S.; Sakly, M.; El-Benna, J.; Amri, M. Hepatoprotective effect of carob against acute ethanol-induced oxidative stress in rat. Toxicol. Ind. Health 2015, 31, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Rtibi, K.; Jabri, M.A.; Selmi, S.; Souli, A.; Sebai, H.; El-Benna, J.; Amri, M.; Marzouki, L. Gastroprotective effect of carob (Ceratonia siliqua L.) against ethanol-induced oxidative stress in rat. BMC Complement. Altern. Med. 2015, 15, 292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pandit, A.; Sachdeva, T.; Bafna, P. Drug-induced hepatotoxicity: A review. J. Appl. Pharm. Sci. 2012, 2, 233–243. [Google Scholar] [CrossRef] [Green Version]
- Akakpo, J.Y.; Ramachandran, A.; Curry, C.S.; Rumack, B.H.; Jaeschke, H. Comparing N-acetylcysteine and 4-methylpyrazole as antidotes for acetaminophen overdose. Arch. Toxicol. 2022, 96, 453–465. [Google Scholar] [CrossRef] [PubMed]
- Woolbright, B.L.; Jaeschke, H. Role of the inflammasome in acetaminophen-induced liver injury and acute liver failure. J. Hepatol. 2017, 66, 836–848. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mossanen, J.C.; Krenkel, O.; Ergen, C.; Govaere, O.; Liepelt, A.; Puengel, T.; Heymann, F.; Kalthoff, S.; Lefebvre, E.; Eulberg, D.; et al. Chemokine (C-C motif) receptor 2-positive monocytes aggravate the early phase of acetaminophen-induced acute liver injury. Hepatology 2016, 64, 1667–1682. [Google Scholar] [CrossRef] [PubMed]
- Rahman, N.; Pervin, M.; Kuramochi, M.; Karim, M.R.; Izawa, T.; Kuwamura, M.; Yamate, J. M1/M2-macrophage polarization-based hepatotoxicity in d-galactosamine-induced acute liver injury in rats. Toxicol. Pathol. 2018, 46, 764–776. [Google Scholar] [CrossRef] [Green Version]
- Frodermann, V.; Nahrendorf, M. Macrophages and cardiovascular health. Physiol. Rev. 2018, 98, 2523–2569. [Google Scholar] [CrossRef] [Green Version]
- Yamate, J.; Izawa, T.; Kuwamura, M. Histopathological analysis of rat hepatotoxicity based on macrophage functions: In particular, an analysis for thioacetamide-induced hepatic lesions. Food Saf. 2016, 4, 61–73. [Google Scholar] [CrossRef] [Green Version]
- Tosello-Trampont, A.C.; Landes, S.G.; Nguyen, V.; Novobrantseva, T.I.; Hahn, Y.S. Kuppfer cells trigger nonalcoholic steatohepatitis development in diet-induced mouse model through tumor necrosis factor-α production. J. Biol. Chem. 2012, 287, 40161–40172. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Run | Independent Variables | Investigated Responses | ||||
|---|---|---|---|---|---|---|
| X1-Ethanol Concentration (%) | X2-Extraction Time (min) | X3-Irradiation Power (W) | DPPH (μM TE/g DW) | FRAP (μM Fe2+/g DW) | ABTS (μM TE/g DW) | |
| 1 | 40 (1) | 15 (0) | 400 (−1) | 85.00 | 44.49 | 181.50 |
| 2 | 20 (0) | 15 (0) | 600 (0) | 71.05 | 43.30 | 142.82 |
| 3 | 20 (0) | 25 (1) | 800 (1) | 68.92 | 37.96 | 165.54 |
| 4 | 0 (−1) | 5 (−1) | 600 (0) | 65.52 | 26.70 | 100.00 |
| 5 | 20 (0) | 25 (1) | 400 (−1) | 58.37 | 41.04 | 163.39 |
| 6 | 40 (1) | 15 (0) | 800 (1) | 103.79 | 51.45 | 167.69 |
| 7 | 40 (1) | 5 (−1) | 600 (0) | 80.66 | 46.25 | 144.67 |
| 8 | 0 (−1) | 25 (1) | 600 (0) | 67.98 | 32.65 | 149.12 |
| 9 | 20 (0) | 5 (−1) | 800 (1) | 58.54 | 38.33 | 131.16 |
| 10 | 0 (−1) | 15 (0) | 800 (1) | 54.89 | 29.81 | 103.23 |
| 11 | 40 (1) | 25 (1) | 600 (0) | 106.85 | 51.22 | 185.64 |
| 12 | 20 (0) | 5 (−1) | 400 (−1) | 56.25 | 41.07 | 131.16 |
| 13 | 20 (0) | 15 (0) | 600 (0) | 65.09 | 40.94 | 142.79 |
| 14 | 20 (0) | 15 (0) | 600 (0) | 64.58 | 35.17 | 135.77 |
| 15 | 20 (0) | 15 (0) | 600 (0) | 65.77 | 31.19 | 143.02 |
| 16 | 20 (0) | 15 (0) | 600 (0) | 73.26 | 39.14 | 148.90 |
| 17 | 0 (−1) | 15 (0) | 400 (−1) | 49.87 | 27.44 | 85.73 |
| Source | DPPH | FRAP | ABTS |
|---|---|---|---|
| p-Value | p-Value | p-Value | |
| Model | 0.0013 * | 0.0329 * | 0.0040 * |
| X1-EtOH (%) | <0.0001 ** | 0.0005 ** | 0.0002 ** |
| X2-Time (min) | 0.0364 * | 0.4286 | 0.0020 ** |
| X3-Power (W) | 0.0550 | 0.7869 | 0.8634 |
| X12 | 0.0733 | 0.9149 | 0.7355 |
| X13 | 0.2611 | 0.6194 | 0.2180 |
| X23 | 0.4880 | 0.9706 | 0.9286 |
| X11 | 0.0025 * | 0.8676 | 0.3585 |
| X22 | 0.9215 | 0.5723 | 0.2124 |
| X33 | 0.0353 * | 0.6381 | 0.6603 |
| Lack of fit | 0.1314 | 0.6249 | 0.0155 * |
| Mean value | 70.38 | 38.71 | 142.48 |
| Std. Dev. | 5.64 | 4.43 | 11.57 |
| CV | 8.01 | 11.43 | 8.12 |
| R2 | 0.945 | 0.848 | 0.922 |
| Optimized Conditions | Ethanol Concentration (%) | Extraction Time (min) | Irradiation Power (W) |
|---|---|---|---|
| DPPH | 40 | 22.4 | 400 |
| FRAP | 40 | 25.0 | 800 |
| ABTS | 40 | 23.7 | 600 |
| Total antioxidant activity | 40 | 25.0 | 800 |
| Compound | Content (mg/g of Dry Extract) * |
|---|---|
| Caffeic acid | 0.05353 ± 0.00268 |
| p-coumaric acid | 0.02386 ± 0.00239 |
| Quercetin | 0.00318 ± 0.00022 |
| Trans-cinnamic acid | <LOD |
| Chlorogenic acid | 0.01667 ± 0.00083 |
| Rosmarinic acid | <LOD |
| Gallic acid | 0.46424 ± 0.06964 |
| Rutin | 0.01883 ± 0.00151 |
| Quercitrin | 0.03967 ± 0.00198 |
| Ferulic acid | <LOD |
| ConS | ConP | CE100 | CE100 + P | CE200 | CE200 + P | |
|---|---|---|---|---|---|---|
| ALT (U/I) | 68.00 ± 12.06 | 100.67 ± 8.39 a | 46.17 ± 12.98 b | 60.17 ± 18.02 b | 55.67 ± 10.35 b | 53.00 ± 7.90 b |
| AST (U/I) | 237.25 ± 72.06 | 348.00 ± 51.83 a | 187.80 ± 47.28 b | 204.67 ± 59.91 b | 193.33 ± 46.68 b | 190.83 ± 32.01 b |
| Direct bilirubin | 1.28 ± 0.31 | 1.45 ± 0.26 | 1.12 ± 0.08 | 1.17 ± 0.16 | 1.22 ± 0.10 | 1.14 ± 0.14 |
| Urea (mmol/L) | 7.12 ± 0.86 | 7.17 ± 1.94 | 7.18 ± 2.05 | 7.65 ± 1.22 | 7.28 ± 2.33 | 7.99 ± 0.42 |
| Creatinine (μmol/L) | 21.80 ± 2.28 | 24.00 ± 2.51 | 18.40 ± 3.65 b | 24.40 ± 2.61 | 19.33 ± 4.32 | 19.29 ± 2.43 b |
| Uric acid (μmol/L) | 181.00 ± 25.35 | 182.40 ± 27.40 | 174.00 ± 34.87 | 108.33 ± 24.81 a,b | 134.4 ± 30.21 | 63.57 ± 11.21 a,b |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Martić, N.; Zahorec, J.; Stilinović, N.; Andrejić-Višnjić, B.; Pavlić, B.; Kladar, N.; Šoronja-Simović, D.; Šereš, Z.; Vujčić, M.; Horvat, O.; et al. Hepatoprotective Effect of Carob Pulp Flour (Ceratonia siliqua L.) Extract Obtained by Optimized Microwave-Assisted Extraction. Pharmaceutics 2022, 14, 657. https://doi.org/10.3390/pharmaceutics14030657
Martić N, Zahorec J, Stilinović N, Andrejić-Višnjić B, Pavlić B, Kladar N, Šoronja-Simović D, Šereš Z, Vujčić M, Horvat O, et al. Hepatoprotective Effect of Carob Pulp Flour (Ceratonia siliqua L.) Extract Obtained by Optimized Microwave-Assisted Extraction. Pharmaceutics. 2022; 14(3):657. https://doi.org/10.3390/pharmaceutics14030657
Chicago/Turabian StyleMartić, Nikola, Jana Zahorec, Nebojša Stilinović, Bojana Andrejić-Višnjić, Branimir Pavlić, Nebojša Kladar, Dragana Šoronja-Simović, Zita Šereš, Miodrag Vujčić, Olga Horvat, and et al. 2022. "Hepatoprotective Effect of Carob Pulp Flour (Ceratonia siliqua L.) Extract Obtained by Optimized Microwave-Assisted Extraction" Pharmaceutics 14, no. 3: 657. https://doi.org/10.3390/pharmaceutics14030657