
Rosinidin Flavonoid Ameliorates Hyperglycemia, Lipid Pathways and Proinflammatory Cytokines in Streptozotocin-Induced Diabetic Rats

 , , , , , , ,
, , , , , , ,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Methods
2.1. Chemicals and Drugs for Testing
2.2. Animals and STZ Paradigm
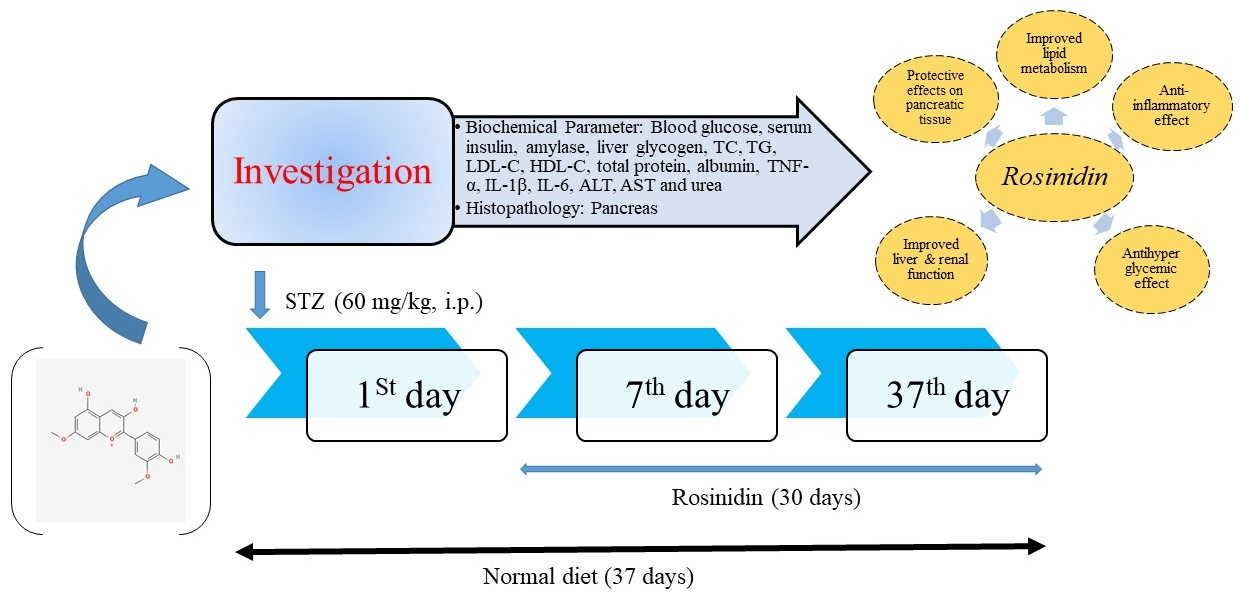
2.3. Research Protocol Demonstration
2.4. Parameters
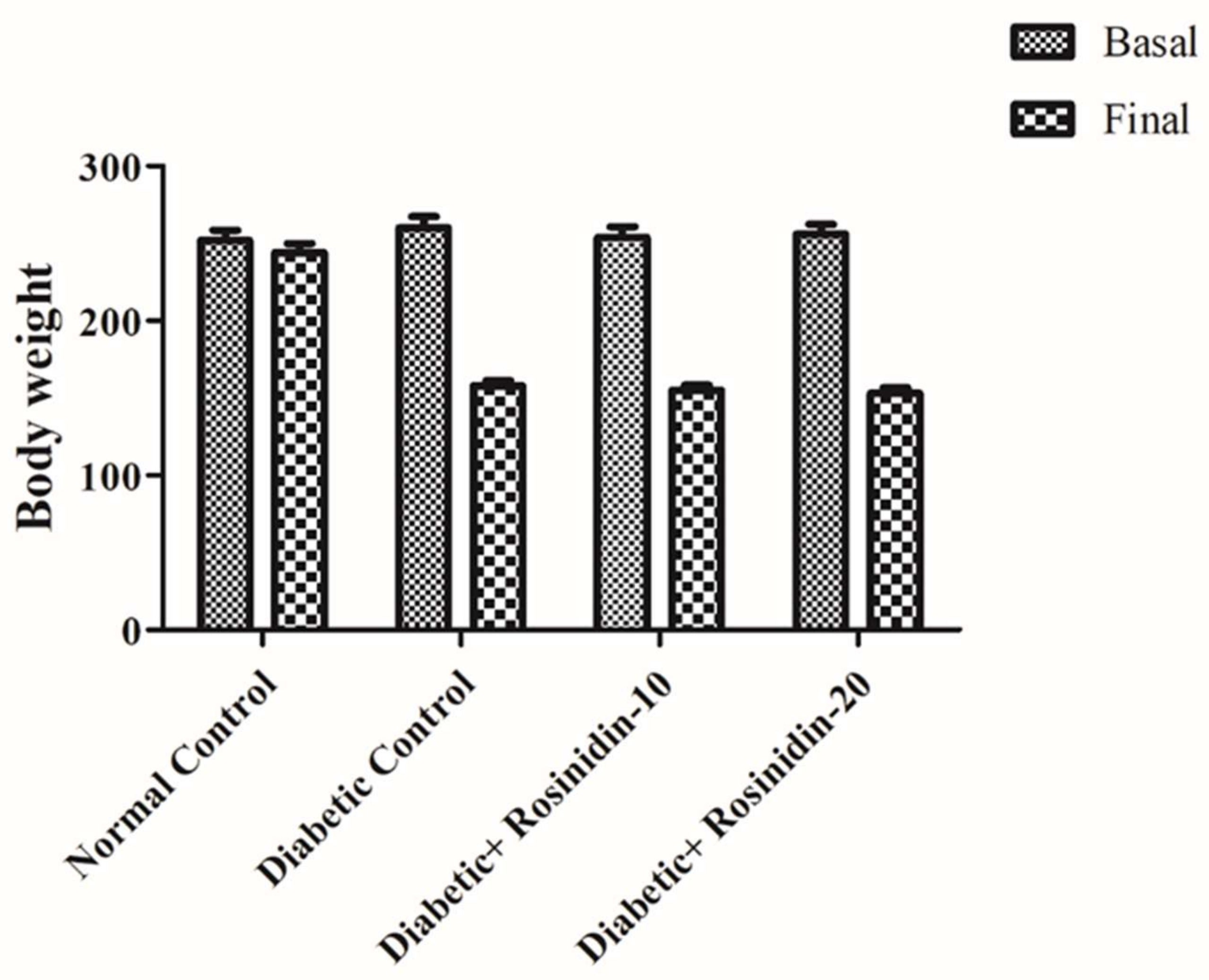
Changes in Body Weight
2.5. Fasting Blood Glucose
2.6. Estimation of Biochemical Parameters
2.7. Determination of Serum Lipid and Total Protein and Albumin
2.8. Assay of Serum Insulin
2.9. Determination of Liver Glycogen
2.10. Determination of α-Amylase
2.11. Determination of Proinflammatory Cytokines
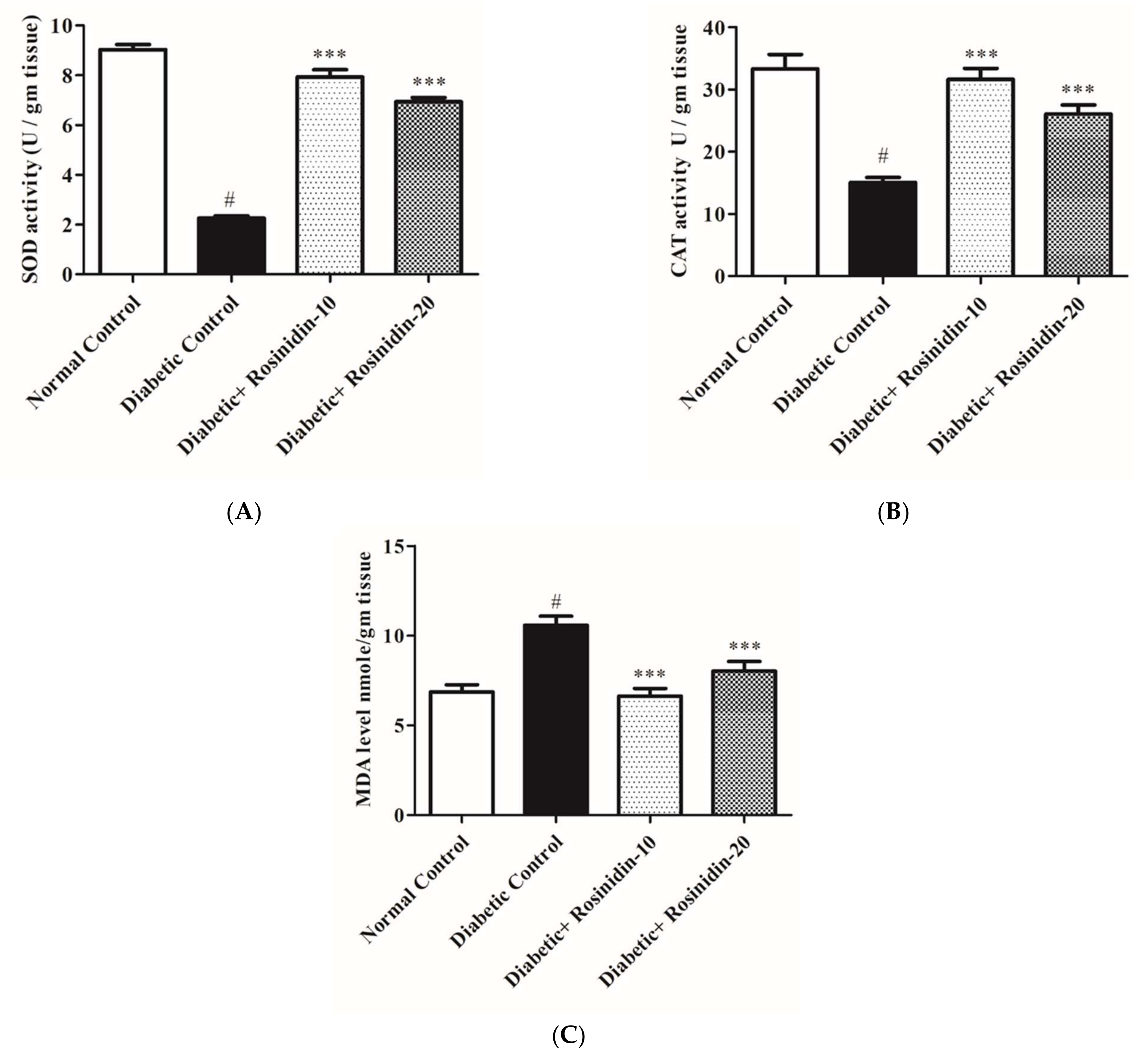
2.12. Determination of Lipid Peroxidation, Superoxide and Catalase
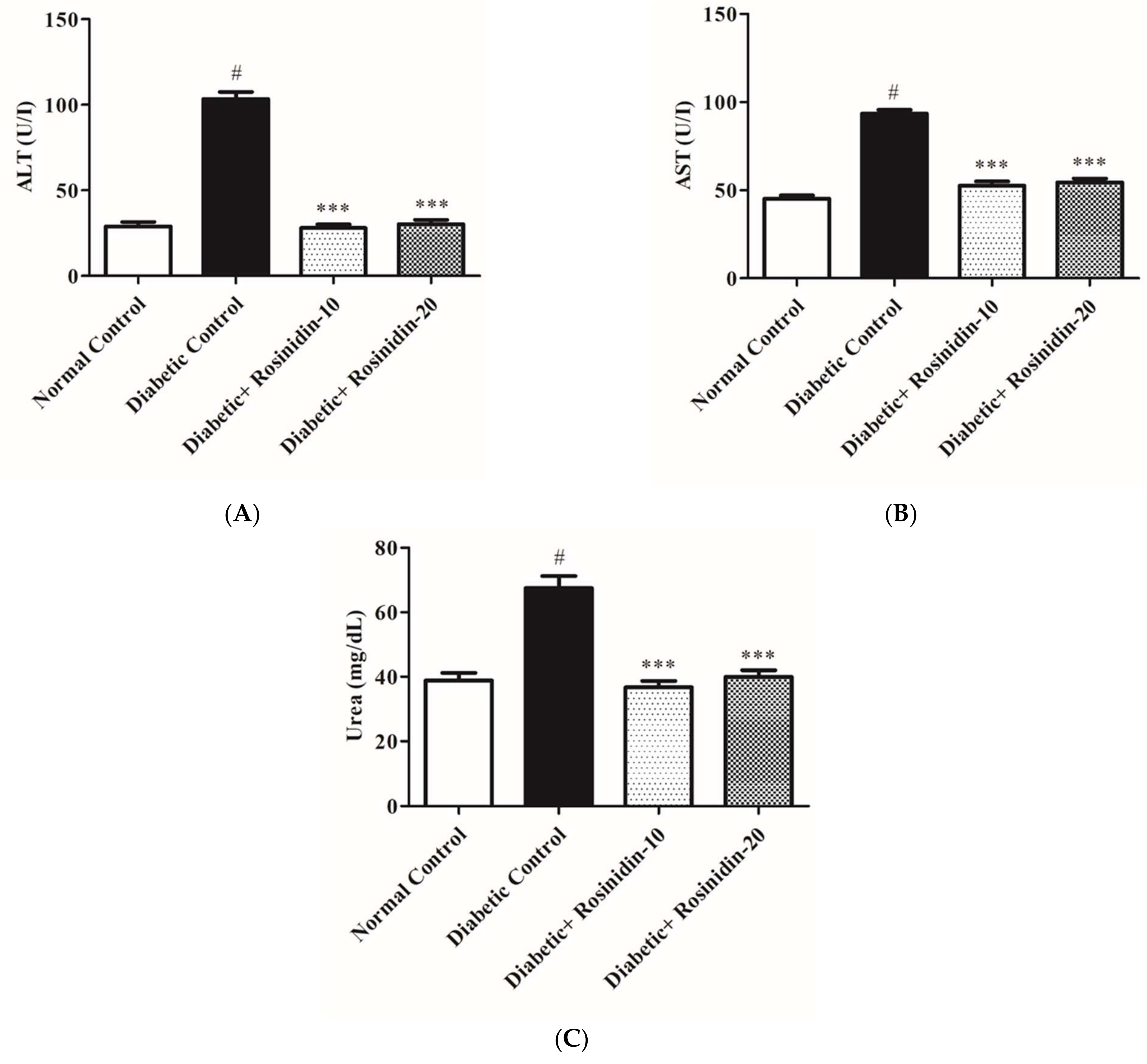
2.13. Determination of Aspartate Aminotransferase (AST), Alanine Aminotransferase (ALT), Urea
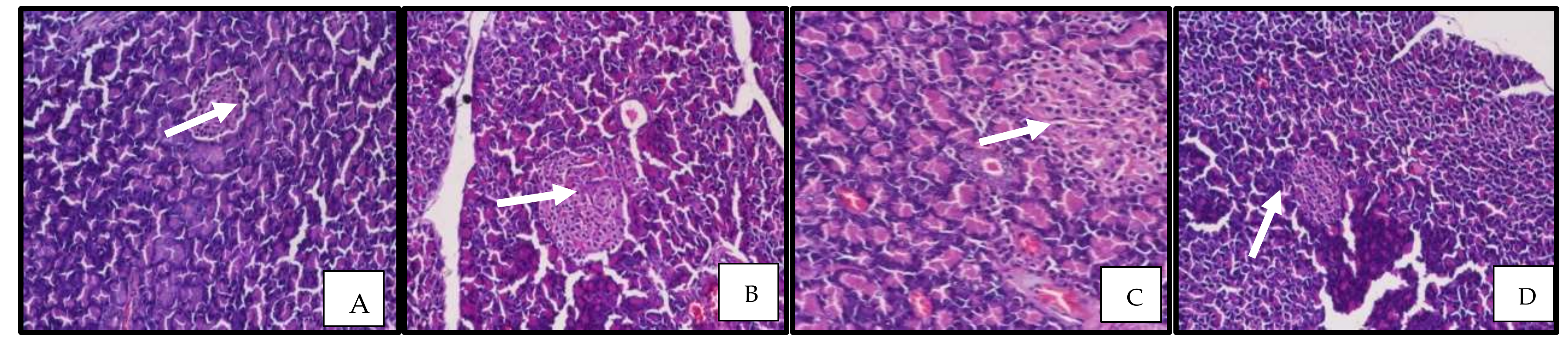
2.14. Histopathological Studies of the Pancreas
2.15. Statistical Analysis
3. Results
3.1. Assessment of Body Weight Changes in Diabetic Rats
3.2. Experimental Evidence for Proposed Place of Rosinidin as an Anti-diabetic Therapy
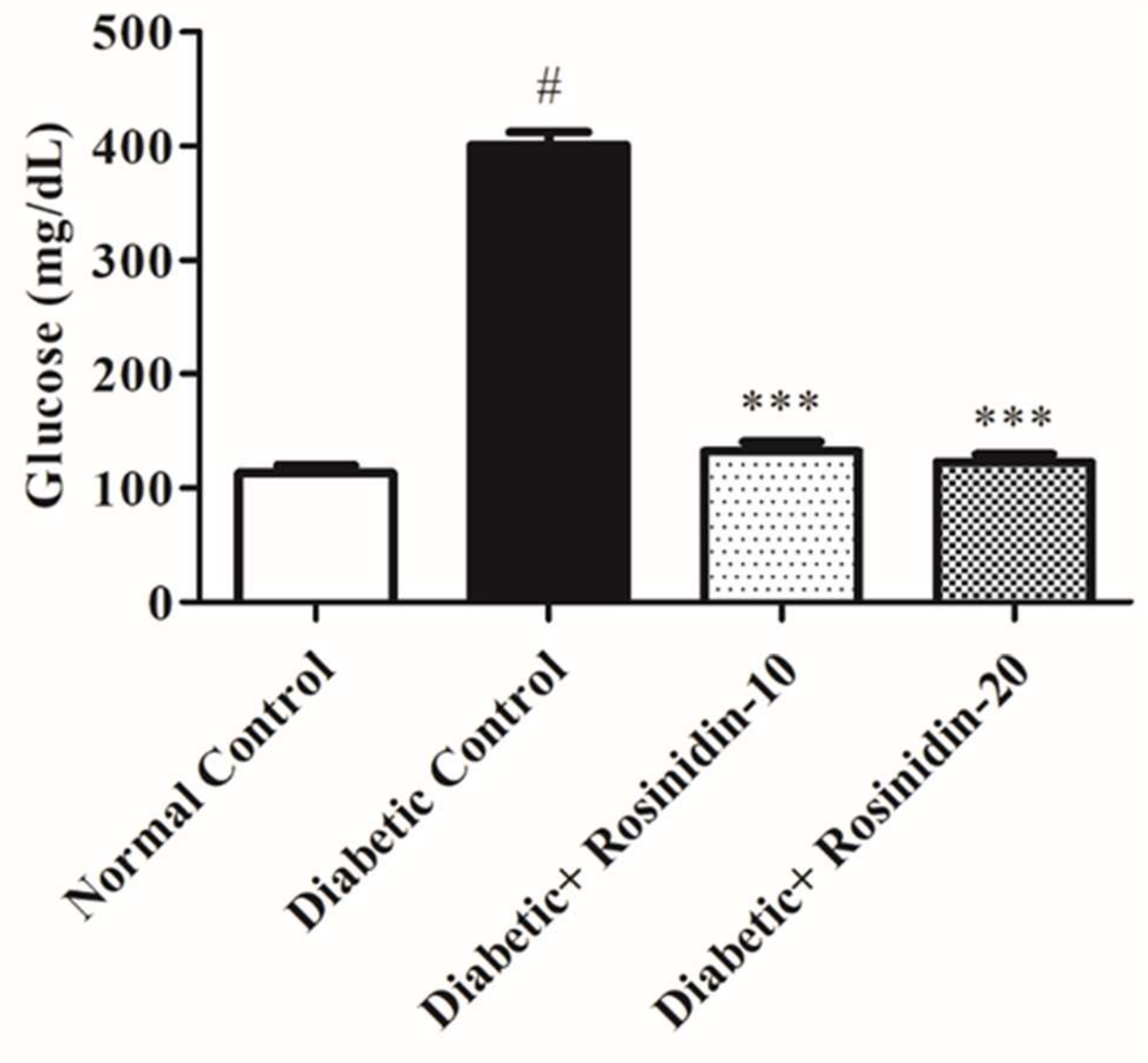
3.2.1. Rosinidin Effect on Glucose in Diabetic Rats
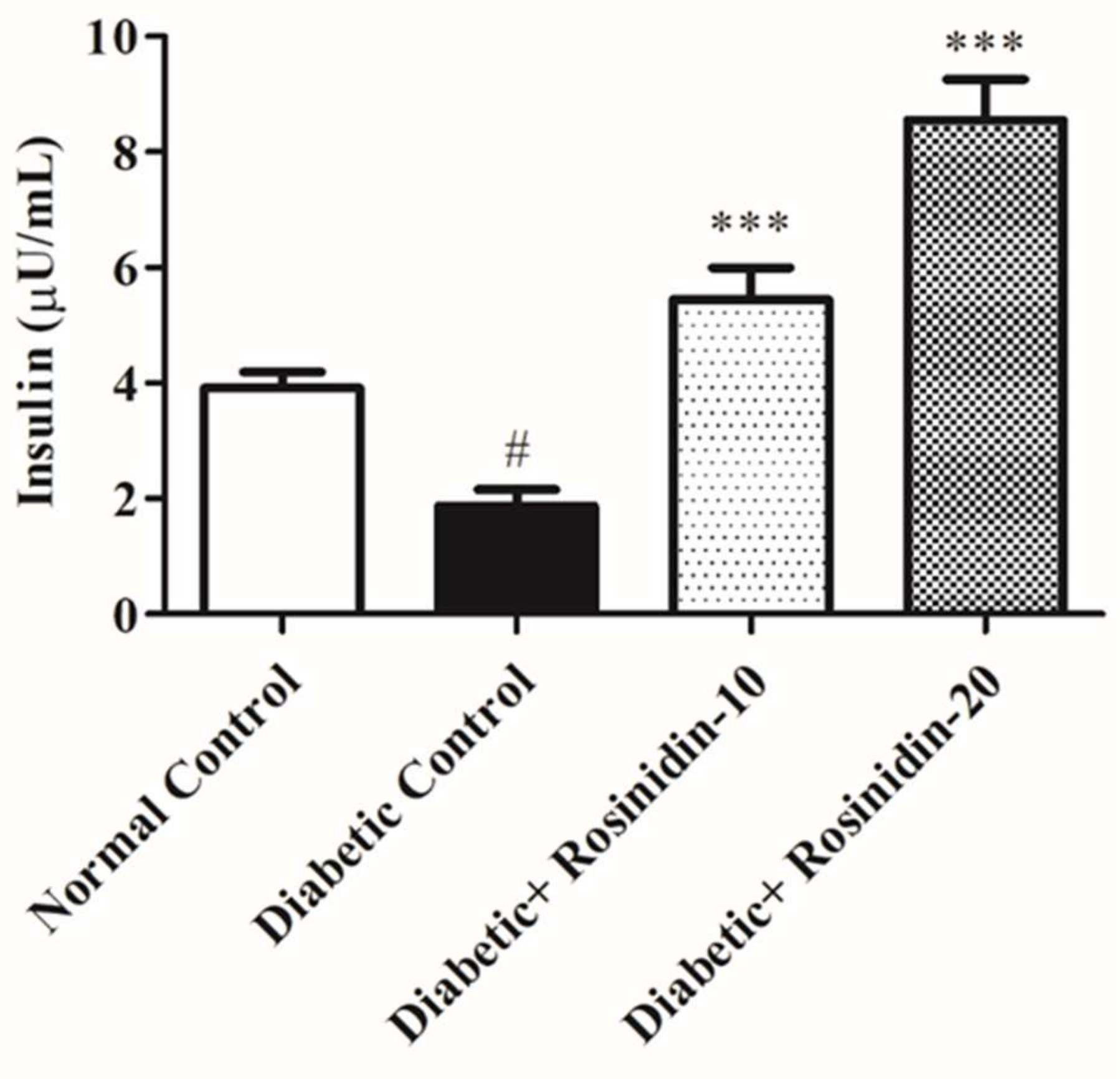
3.2.2. Rosinidin Effect on Insulin in Diabetic Rats
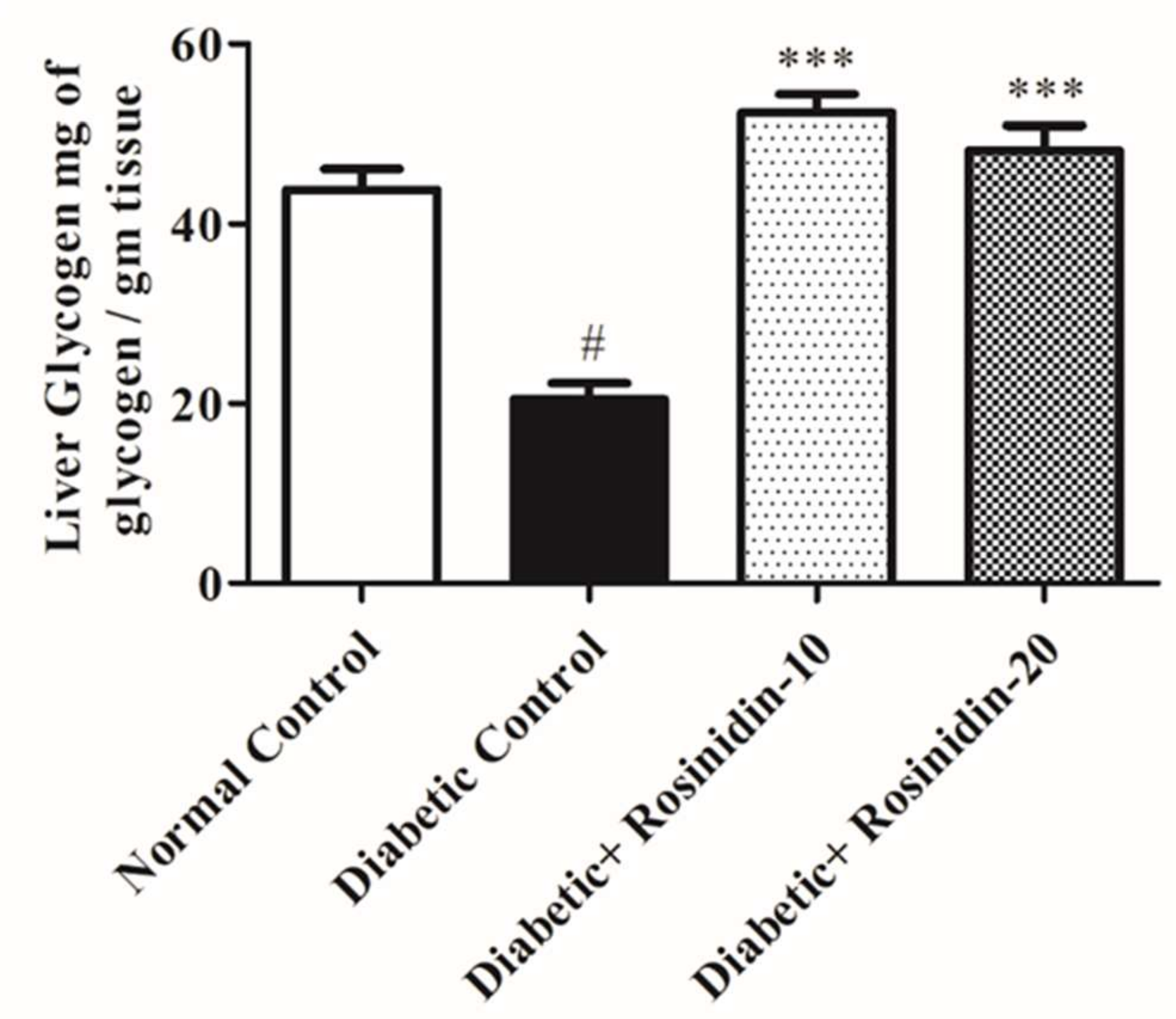
3.2.3. Rosinidin Effect on Liver Glycogen in Diabetic Rats
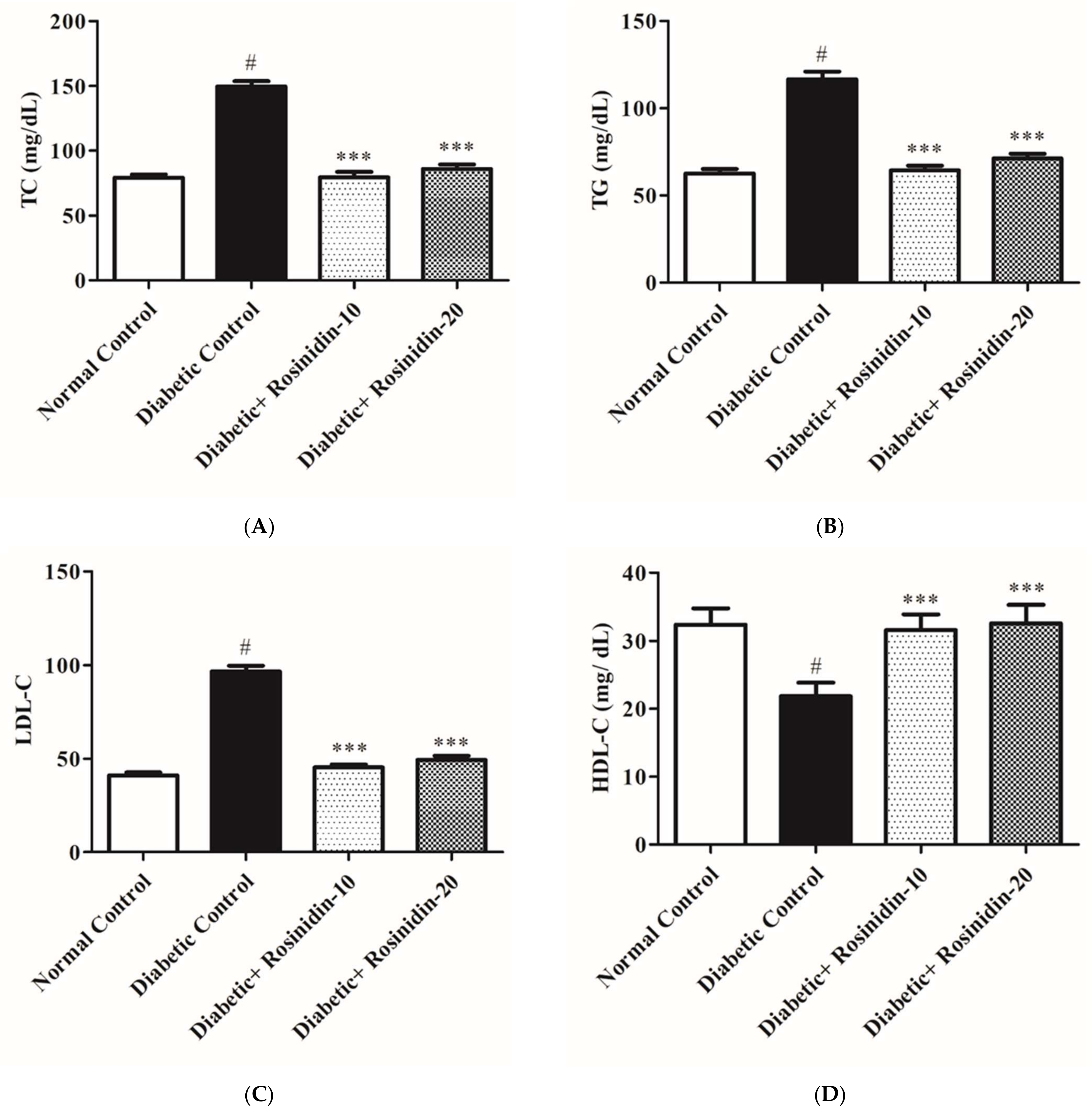
3.2.4. Rosinidin Effect on Lipid Profiles in Diabetic Rats
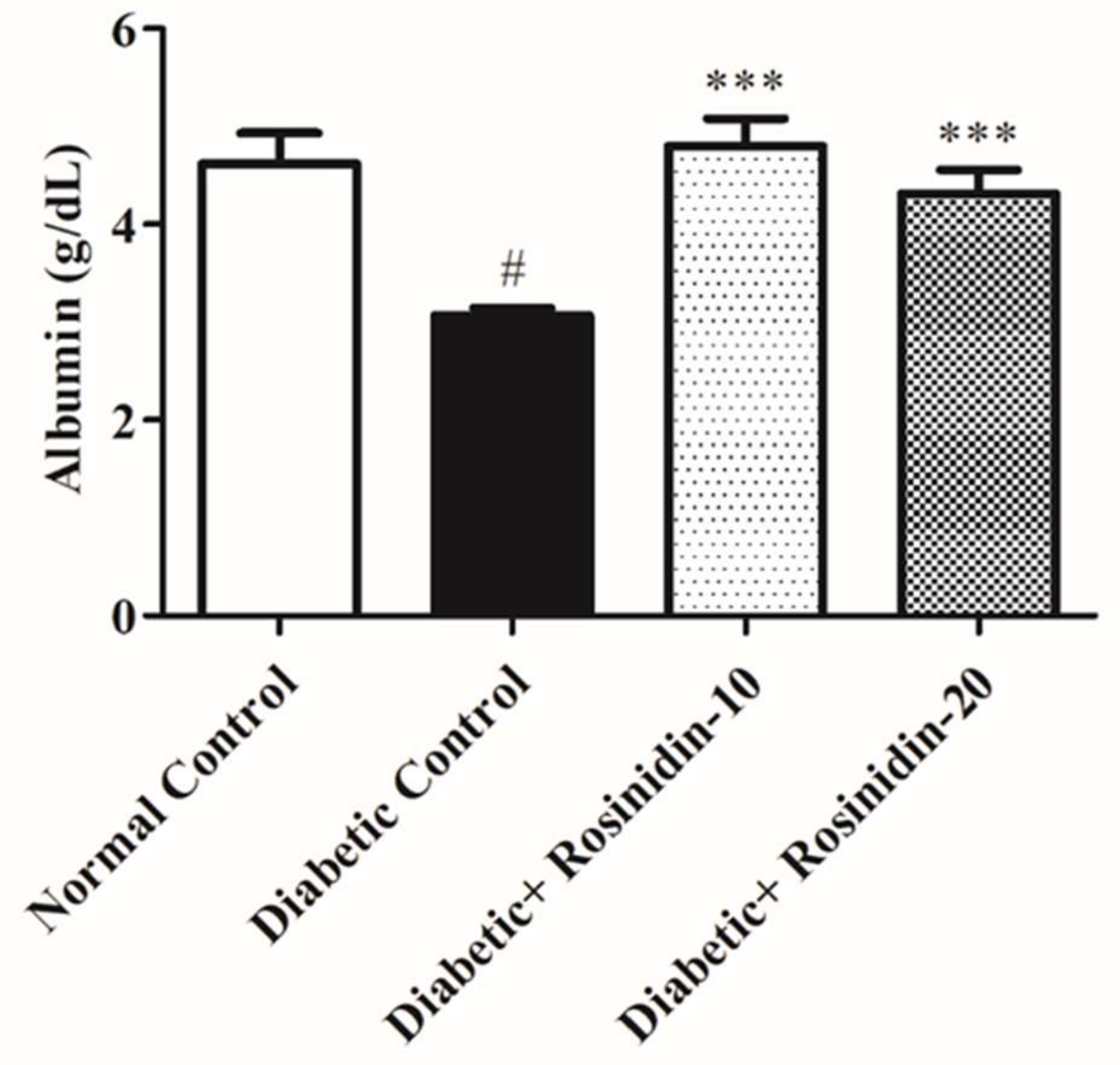
3.2.5. Rosinidin Effect on Total Protein and Albumin in Diabetic Rats
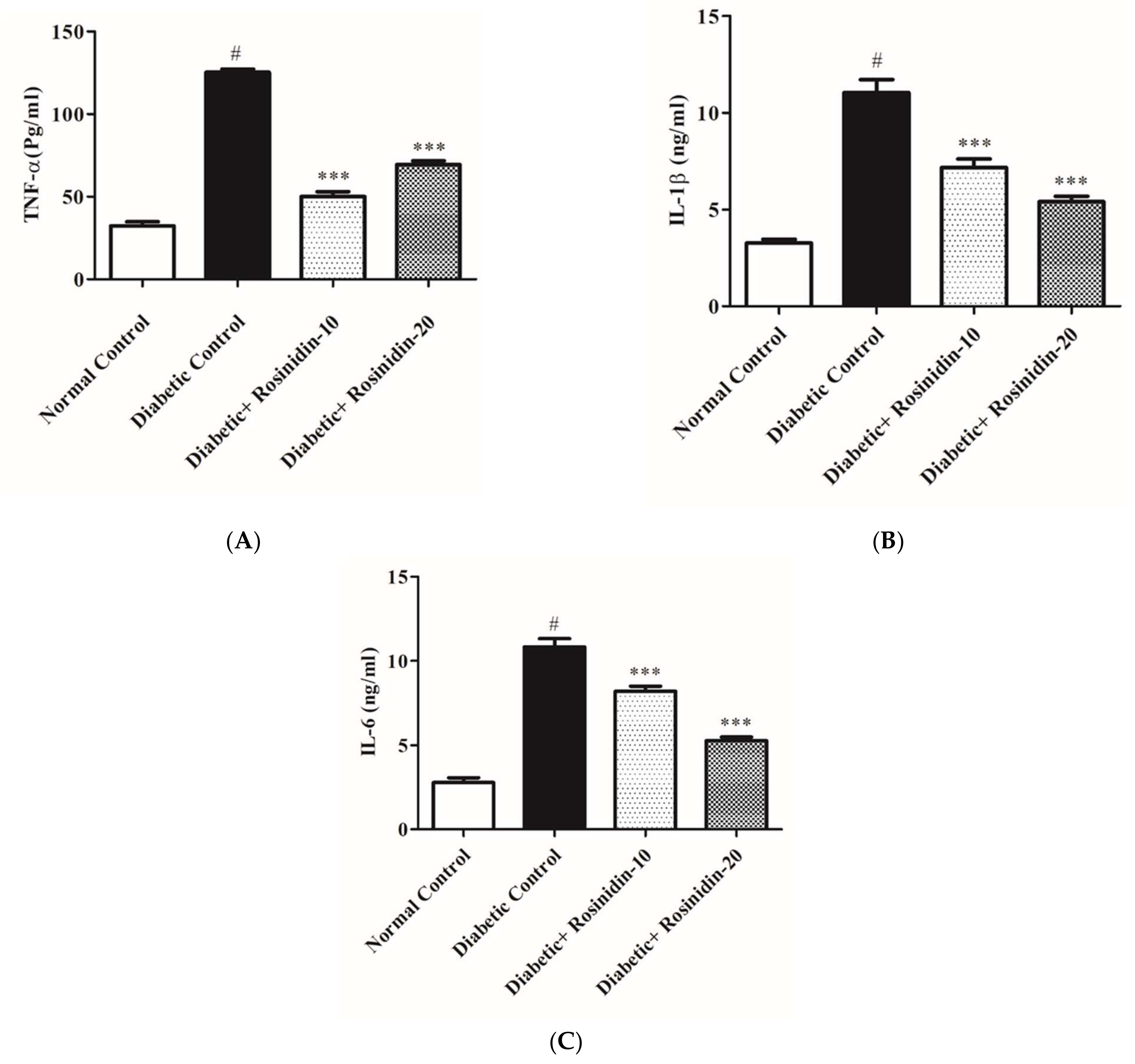
3.2.6. Rosinidin Effect on Proinflammatory Cytokines in Diabetic Rats
3.2.7. Rosinidin Effects on Antioxidant and Oxidative Stress Markers in Diabetic Rats
3.2.8. Rosinidin Effect on Hepatic and Renal Function in Diabetic Rats
3.2.9. Rosinidin Effect on α-Amylase Level in Diabetic Rats
3.2.10. Rosinidin Effect on Pancreatic Function and Photomicrographs of Pancreatic Tissue
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Kovács, N.; Nagy, A.; Dombrádi, V.; Bíró, K. Inequalities in the Global Burden of Chronic Kidney Disease Due to Type 2 Diabetes Mellitus: An Analysis of Trends from 1990 to 2019. Int. J. Environ. Res. Public Health 2021, 18, 4723. [Google Scholar] [CrossRef]
- Daryabor, G.; Atashzar, M.R.; Kabelitz, D.; Meri, S.; Kalantar, K. The Effects of Type 2Diabetes Mellitus on Organ Metabolism and the ImmuneSystem. Front. Immunol. 2020, 11, 1582. [Google Scholar] [CrossRef] [PubMed]
- Choudhury, H.; Pandey, M.; Hua, C.K.; Mun, C.S.; Jing, J.K.; Kong, L.; Ern, L.Y.; Ashraf, N.A.; Kit, S.W.; Yee, T.S.; et al. An update on natural compounds in the remedy of diabetes mellitus: A systematic review. J. Tradit. Complement. Med. 2017, 8, 361–376. [Google Scholar] [CrossRef]
- Mutha, R.E.; Tatiya, A.U.; Surana, S.J. Flavonoids as natural phenolic compounds and their role in therapeutics: An overview. Futur. J. Pharm. Sci. 2021, 791, 25. [Google Scholar] [CrossRef]
- Lim, S.H.; Yu, J.S.; Lee, H.S.; Choi, C.-I.; Kim, K.H. Antidiabetic Flavonoids from Fruits of Morus alba Promoting Insulin-Stimulated Glucose Uptake via Akt and AMP-Activated Protein Kinase Activation in 3T3-L1 Adipocytes. Pharmaceutics 2021, 13, 526. [Google Scholar] [CrossRef]
- Martín, M.Á.; Ramos, S. Dietary Flavonoids and Insulin Signaling in Diabetes and Obesity. Cells 2021, 10, 1474. [Google Scholar] [CrossRef]
- Lang, X.; Li, N.; Li, L.; Zhang, S. Integrated Metabolome and Transcriptome Analysis Uncovers the Role of Anthocyanin Metabolism in Micheliamaudiae. Int. J. Genom. 2019, 3, 4393905. [Google Scholar]
- Khoo, H.E.; Azlan, A.; Tang, S.T.; Lim, S.M. Anthocyanidins and anthocyanins: Colored pigments as food, pharmaceutical ingredients, and the potential health benefits. Food Nutr. Res. 2017, 61, 1361779. [Google Scholar] [CrossRef] [Green Version]
- Monteiro, A.F.; Viana, J.D.; Nayarisseri, A.; Zondegoumba, E.N.; Mendonça Junior, F.J.; Scotti, M.T.; Scotti, L. ComputationalStudies Applied to Flavonoids against Alzheimer’s and Parkinson’s Diseases. Oxid. Med. Cell. Longev. 2018, 2018, 7912765. [Google Scholar] [CrossRef]
- Arokoyo, D.S.; Oyeyipo, I.P.; Du Plessis, S.S.; Chegouand, N.N.; Aboua, Y.G. Modulation of Inflammatory Cytokines and Islet Morphology as Therapeutic Mechanisms of Basella alba in Streptozotocin-Induced Diabetic Rats. Toxicol. Res. 2018, 34, 325–332. [Google Scholar] [CrossRef] [Green Version]
- Fathiazad, F.; Hamedeyazdan, S.; Khosropanah, M.K.; Khaki, A. Hypoglycemic activity of Fumaria parviflora in streptozotocin-induced diabetic rats. Adv. Pharm. Bull. 2013, 3, 207. [Google Scholar]
- Ali, A.A.; Essawy, E.A.; Hamed, H.S.; Abdel Moneim, A.E.; Attaby, F.A. The ameliorative role of Physalis pubescens L. against neurological impairment associated with streptozotocin induced diabetes in rats. Metab. Brain Dis. 2021, 36, 1191–1200. [Google Scholar] [CrossRef]
- Soni, L.K.; Dobhal, M.P.; Arya, D.; Bhagour, K.; Parasher, P.; Gupta, R.S. In vitro and in vivo antidiabetic activity of isolated fraction of Prosopis cineraria against streptozotocin-induced experimental diabetes: A mechanistic study. Biomed. Pharmacother. 2018, 108, 1015–1021. [Google Scholar] [CrossRef]
- Al-Ishaq, R.K.; Abotaleb, M.; Kubatka, P.; Kajo, K.; Büsselberg, D. Flavonoids and Their Anti-Diabetic Effects: Cellular Mechanisms and Effects to Improve Blood Sugar Levels. Biomolecules 2019, 9, 430. [Google Scholar] [CrossRef] [Green Version]
- Elshemy, M.M.; Asem, M.; Allemailem, K.S.; Uto, K.; Ebara, M.; Nabil, A. Antioxidative Capacity of Liver- and Adipose-Derived Mesenchymal Stem Cell-Conditioned Media and Their Applicability in Treatment of Type 2 Diabetic Rats. Oxid. Med. Cell. Longev. 2021, 2021, 8833467. [Google Scholar] [CrossRef]
- Rezaei, S.; Ashkar, F.; Koohpeyma, F.; Mahmoodi, M.; Gholamalizadeh, M.; Mazloom, Z.; Doaei, S. Hydroalcoholic extract of Achilleamillefolium improved blood glucose, liver enzymes and lipid profile compared to metformin in streptozotocin-induced diabetic rats. Lipids Health Dis. 2020, 19, 81. [Google Scholar] [CrossRef]
- Hussain, T.; Tan, B.; Murtaza, G.; Liu, G.; Rahu, N.; Saleem-Kalhoro, M.; Hussain-Kalhoro, D.; Adebowale, T.O.; Usman-Mazhar, M.; Rehman, Z.U.; et al. Flavonoids and type 2 diabetes: Evidence of efficacy in clinical and animal studies and delivery strategies to enhance their therapeutic efficacy. Pharmacol. Res. 2020, 152, 104629. [Google Scholar] [CrossRef]
- Bai, L.; Li, X.; He, L.; Zheng, Y.; Lu, H.; Li, J.; Zhong, L.; Tong, R.; Jiang, Z.; Shi, J.; et al. Antidiabetic Potential of Flavonoids from Traditional Chinese Medicine: A Review. Am. J. Chin. Med. 2019, 47, 933–957. [Google Scholar] [CrossRef]
- Othman, M.S.; Khaled, A.M.; Al-Bagawi, A.H.; Fareid, M.A.; Ghany, R.A.; Habotta, O.A.; Moneim AE, A. Hepatorenal protective efficacy of flavonoids from Ocimumbasilicum extract in diabetic albino rats: A focus on hypoglycemic, antioxidant, anti-inflammatory and anti-apoptotic activities. Biomed. Pharmacother. 2021, 144, 112287. [Google Scholar] [CrossRef]
- Li, C.; Gan, H.; Tan, X.; Hu, Z.; Deng, B.; Sullivan, M.A.; Gilbert, R.G. Effects of active ingredients from traditional Chinese medicines on glycogen molecular structure in diabetic mice. Eur. Polym. J. 2019, 112, 67–72. [Google Scholar] [CrossRef]
- Copeland, L.A.; Swendsen, C.S.; Sears, D.M.; MacCarthy, A.A.; McNeal, C.J. Association between triglyceride levels and cardiovascular disease in patients with acute pancreatitis. PLoS ONE 2018, 13, e0179998. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El-Ouady, F.; Bachir, F.; Eddouks, M. Flavonoids Extracted from Asteris cusgraveolens improve glucose metabolism and lipid profile in diabetic Rats. Endocr. Metab. Immune Disord. Drug Targets 2021, 21, 895–904. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, Z.; Zhao, H.; Wang, X.; Pang, J.; Li, Q.; Yang, Y.; Ling, W. Anthocyanin supplementation improves anti-oxidative and anti-inflammatory capacity in a dose-response manner in subjects with dyslipidemia. Redox Biol. 2020, 32, 101474. [Google Scholar] [CrossRef] [PubMed]
- Kassab, R.B.; Lokman, M.S.; Daabo, H.M.; Gaber, D.A.; Habotta, O.A.; Hafez, M.M.; Zhery, A.S.; Moneim, A.E.A.; Fouda, M.S. Ferulic acid influences Nrf2 activation to restore testicular tissue from cadmium-induced oxidative challenge, inflammation, and apoptosis in rats. J. Food Biochem. 2020, 44, e13505. [Google Scholar] [CrossRef]
- Zou, J.; Sui, D.; Fu, W.; Li, Y.; Yu, P.; Yu, X.; Xu, H. Total flavonoids extracted from the leaves of Murraya paniculata (L.) Jack alleviate oxidative stress, inflammation and apoptosis in a rat model of diabetic cardiomyopathy. J. Funct. Foods 2021, 76, 104319. [Google Scholar] [CrossRef]
- Ajayi, A.M.; Umukoro, S.; Ben-Azu, B.; Adzu, B.; Ademowo, O.G. Toxicity and protective effect of phenolic-enriched ethylacetate fraction of Ocimum gratissimum (Linn.) leaf against acute inflammation and oxidative stress in rats. Drug Dev. Res. 2017, 78, 135–145. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.L.; Liu, W.P.; Wang, L.L.; Feng, L. Effects of flavonoids from Pyrrosiae folium on pathological changesand inflammatory response of diabetic nephropathy. China J. Chin. Mater. Med. 2018, 43, 2352–2357. [Google Scholar]
- Nazir, N.; Zahoor, M.; Ullah, R.; Ezzeldin, E.; Mostafa, G.A.E. Curative Effect of Catechin Isolated from Elaeagnus Umbellata Thunb. Berries for Diabetes and Related complications in Streptozotocin-Induced Diabetic Rats Model. Molecules 2020, 26, 137. [Google Scholar] [CrossRef]
- Zaky, A.S.; Kandeil, M.; Abdel-Gabbar, M.; Fahmy, E.M.; Almehmadi, M.M.; Ali, T.M.; Ahmed, O.M. The Antidiabetic Effects and Modes of Action of the Balanites aegyptiaca Fruit and Seed Aqueous Extracts in NA/STZ-Induced Diabetic Rats. Pharmaceutics 2022, 14, 263. [Google Scholar] [CrossRef]
- Arigela, C.S.; Nelli, G.; Gan, S.H.; Sirajudeen, K.N.S.; Krishnan, K.; Abdul Rahman, N.; Pasupuleti, V.R. Bitter Gourd Honey Ameliorates Hepatic and Renal Diabetic Complications on Type 2 Diabetes Rat Models by Antioxidant, Anti-Inflammatory, and Anti-Apoptotic Mechanisms. Foods 2021, 10, 2872. [Google Scholar] [CrossRef]
- Ogunlana, O.O.; Adetuyi, B.O.; Esalomi, E.F.; Rotimi, M.I.; Popoola, J.O.; Ogunlana, O.E.; Adetuyi, O.A. Antidiabetic and Antioxidant Activities of the Twigs of Andrograhis paniculata on Streptozotocin-Induced Diabetic Male Rats. BioChem 2021, 1, 238–249. [Google Scholar] [CrossRef]
- Esquivel-Gutiérrez, E.R.; Manzo-Avalos, S.; Peña-Montes, D.J.; Saavedra-Molina, A.; Morreeuw, Z.P.; Reyes, A.G. Hypolipidemic and Antioxidant Effects of Guishe Extract from Agave lechuguilla, a Mexican Plant with Biotechnological Potential, on Streptozotocin-Induced Diabetic Male Rats. Plants 2021, 10, 2492. [Google Scholar] [CrossRef]












Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gilani, S.J.; Bin-Jumah, M.N.; Al-Abbasi, F.A.; Nadeem, M.S.; Imam, S.S.; Alshehri, S.; Ghoneim, M.M.; Afzal, M.; Alzarea, S.I.; Sayyed, N.; et al. Rosinidin Flavonoid Ameliorates Hyperglycemia, Lipid Pathways and Proinflammatory Cytokines in Streptozotocin-Induced Diabetic Rats. Pharmaceutics 2022, 14, 547. https://doi.org/10.3390/pharmaceutics14030547
Gilani SJ, Bin-Jumah MN, Al-Abbasi FA, Nadeem MS, Imam SS, Alshehri S, Ghoneim MM, Afzal M, Alzarea SI, Sayyed N, et al. Rosinidin Flavonoid Ameliorates Hyperglycemia, Lipid Pathways and Proinflammatory Cytokines in Streptozotocin-Induced Diabetic Rats. Pharmaceutics. 2022; 14(3):547. https://doi.org/10.3390/pharmaceutics14030547
Chicago/Turabian StyleGilani, Sadaf Jamal, May Nasser Bin-Jumah, Fahad A. Al-Abbasi, Muhammad Shahid Nadeem, Syed Sarim Imam, Sultan Alshehri, Mohammed M. Ghoneim, Muhammad Afzal, Sami I. Alzarea, Nadeem Sayyed, and et al. 2022. "Rosinidin Flavonoid Ameliorates Hyperglycemia, Lipid Pathways and Proinflammatory Cytokines in Streptozotocin-Induced Diabetic Rats" Pharmaceutics 14, no. 3: 547. https://doi.org/10.3390/pharmaceutics14030547