
Ribozyme Assays to Quantify the Capping Efficiency of In Vitro-Transcribed mRNA

 , ,
, ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Templates for In Vitro Transcription
2.2. In Vitro Transcription and Capping of RNA
2.3. Design of Ribozymes and RNase H Probe
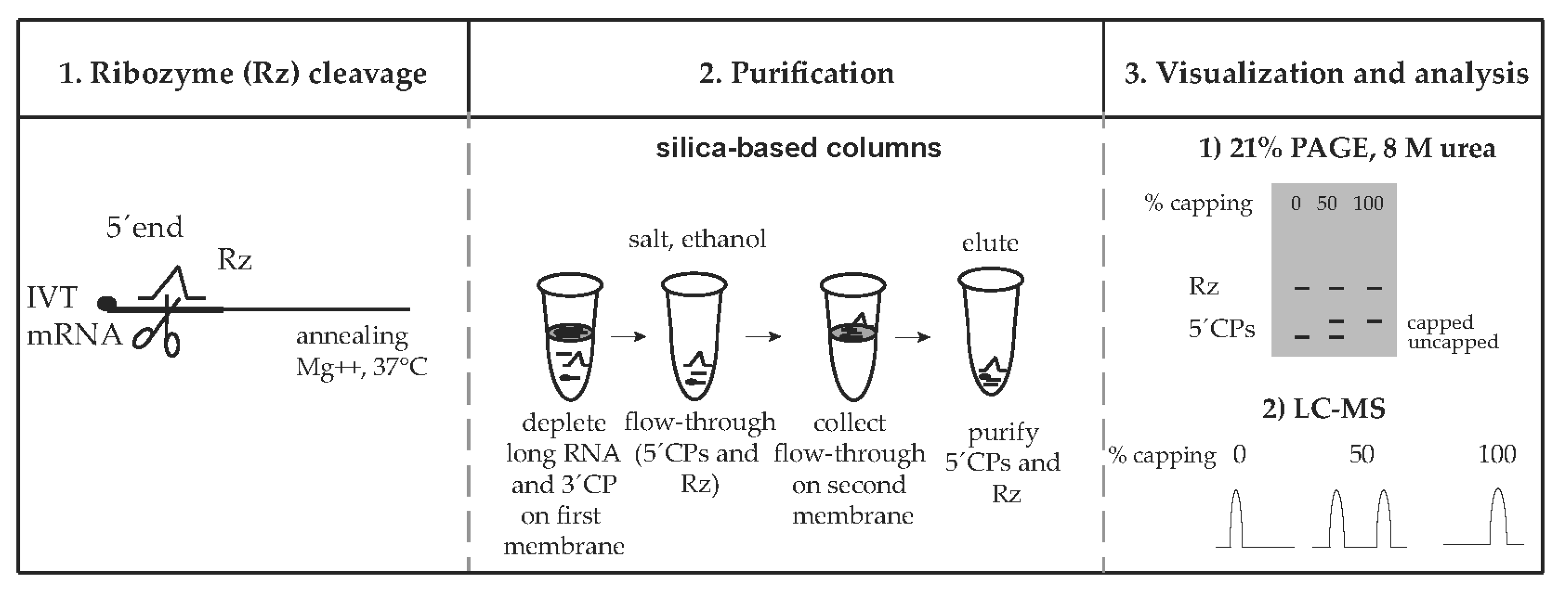
2.4. The mRNA Cleavage by Ribozyme
2.5. The mRNA Cleavage by RNase H
2.6. Purification of Cleaved, Short RNA Fragments Using Silica-Based Columns
2.7. Purification of Cleaved, Short Fragments by Elution from Polyacrylamide Gel
2.8. Visualization and Analysis of Cleaved, Short Fragments by PAGE
2.9. LC–MS Analysis of Ribozyme-Cleaved Products
3. Results
3.1. Ribozyme Assays to Quantify the Capping Efficiency of IVT mRNA
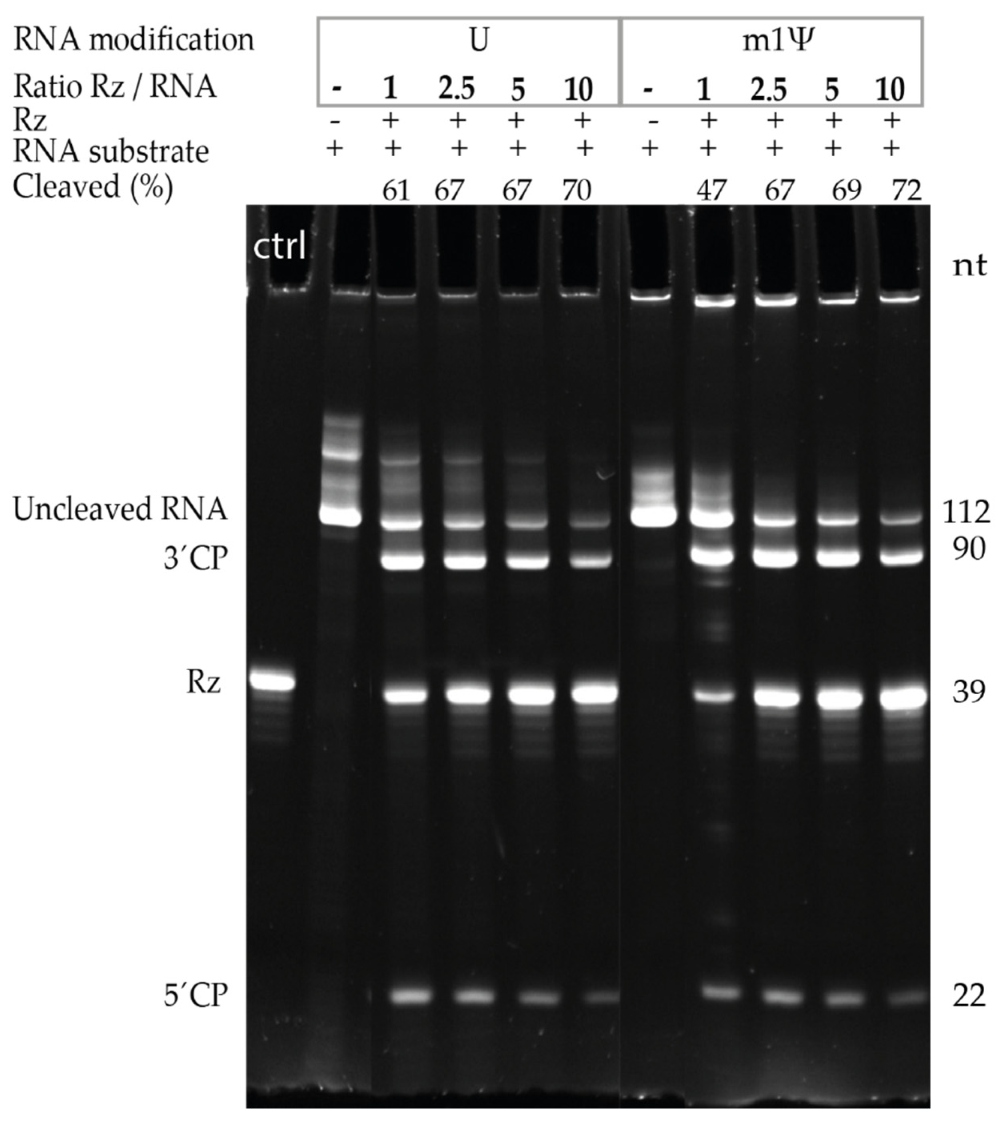
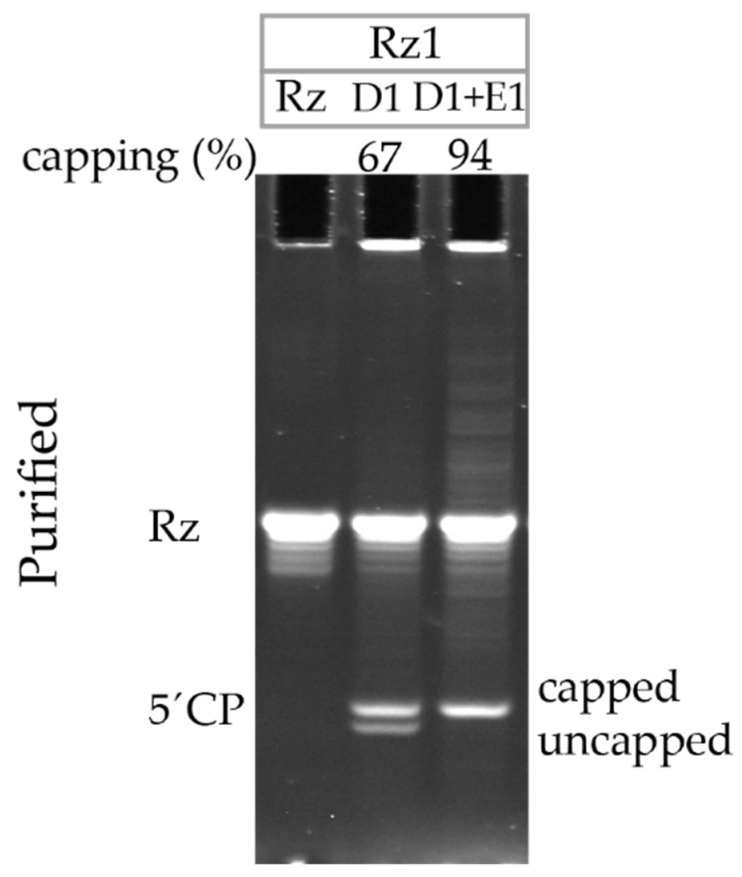
3.2. Ribozyme Cleavage Assay Optimization
3.3. Capping Efficiency Quantification after Visualizing 5′ Cleavage Products Using 21% PAGE, 8 M Urea
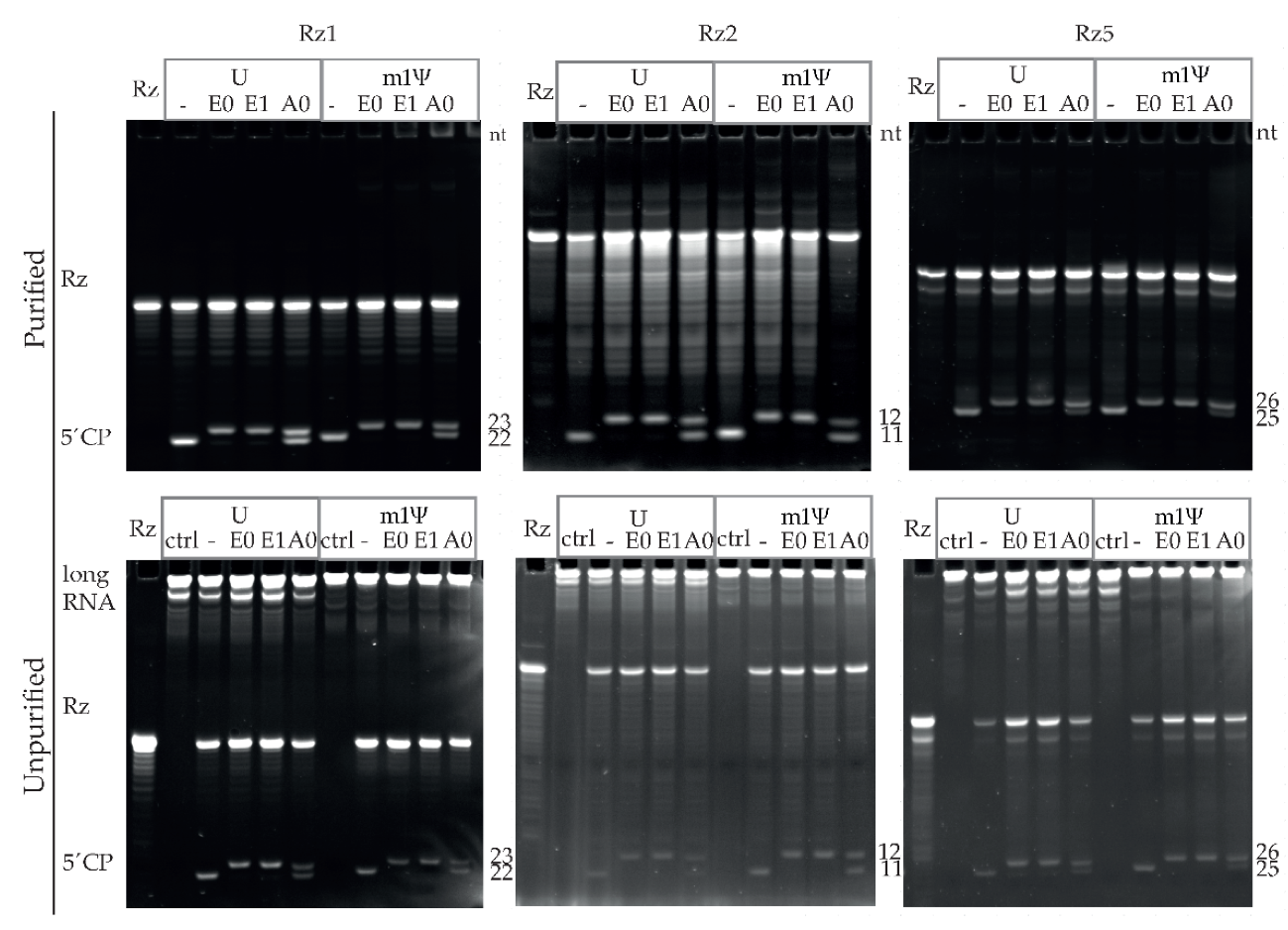
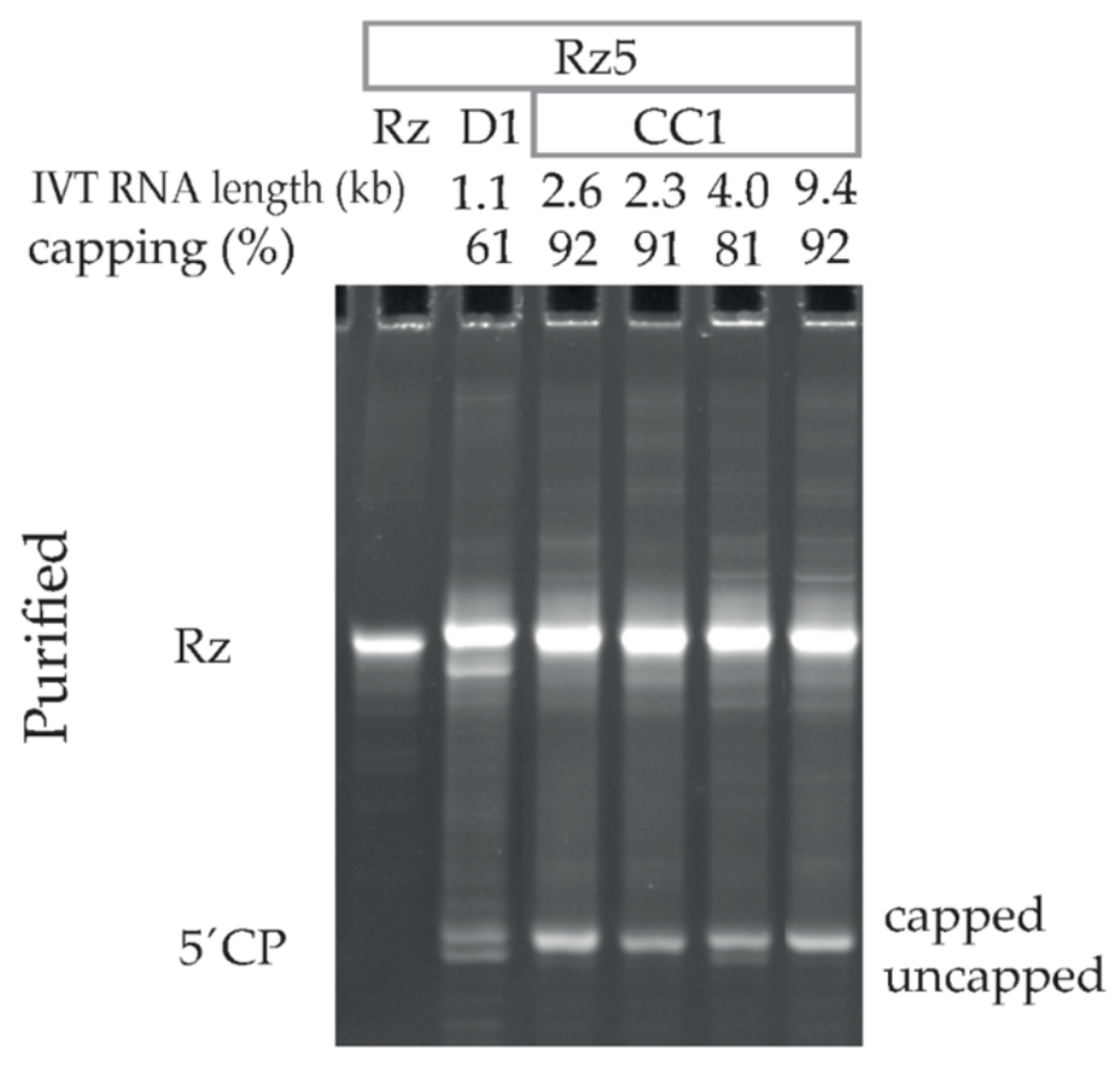
3.4. Capping Assay by Ribozyme-Mediated Cleavage Effectively Assesses Capping Efficiencies of Diversely Capped IVT mRNAs of Different Lengths
3.5. Ribozyme-Mediated Cleavage Assay Detects an Increase in Capping Efficiency after Additional Enzymatic Capping of Co-Transcriptionally Capped RNA
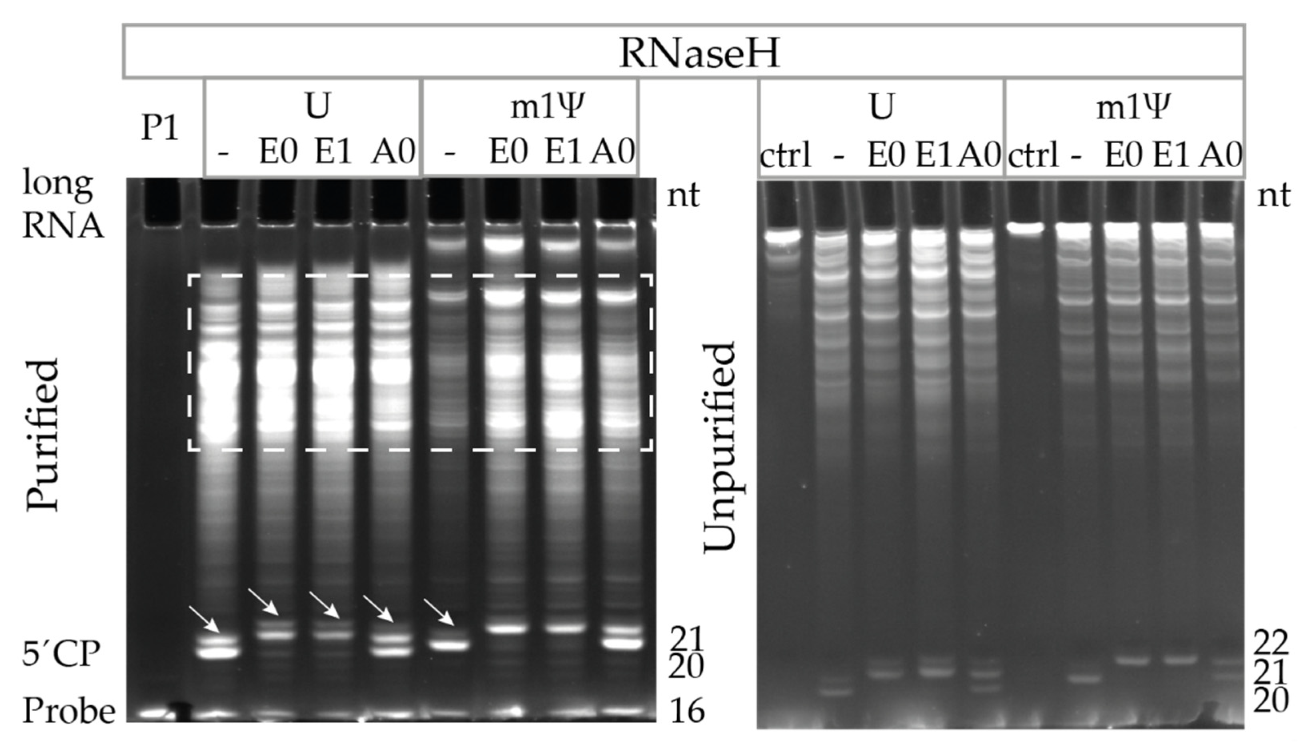
3.6. Ribozyme-Mediated Cleavage Assay Performance Superior to RNase H Cleavage Assay
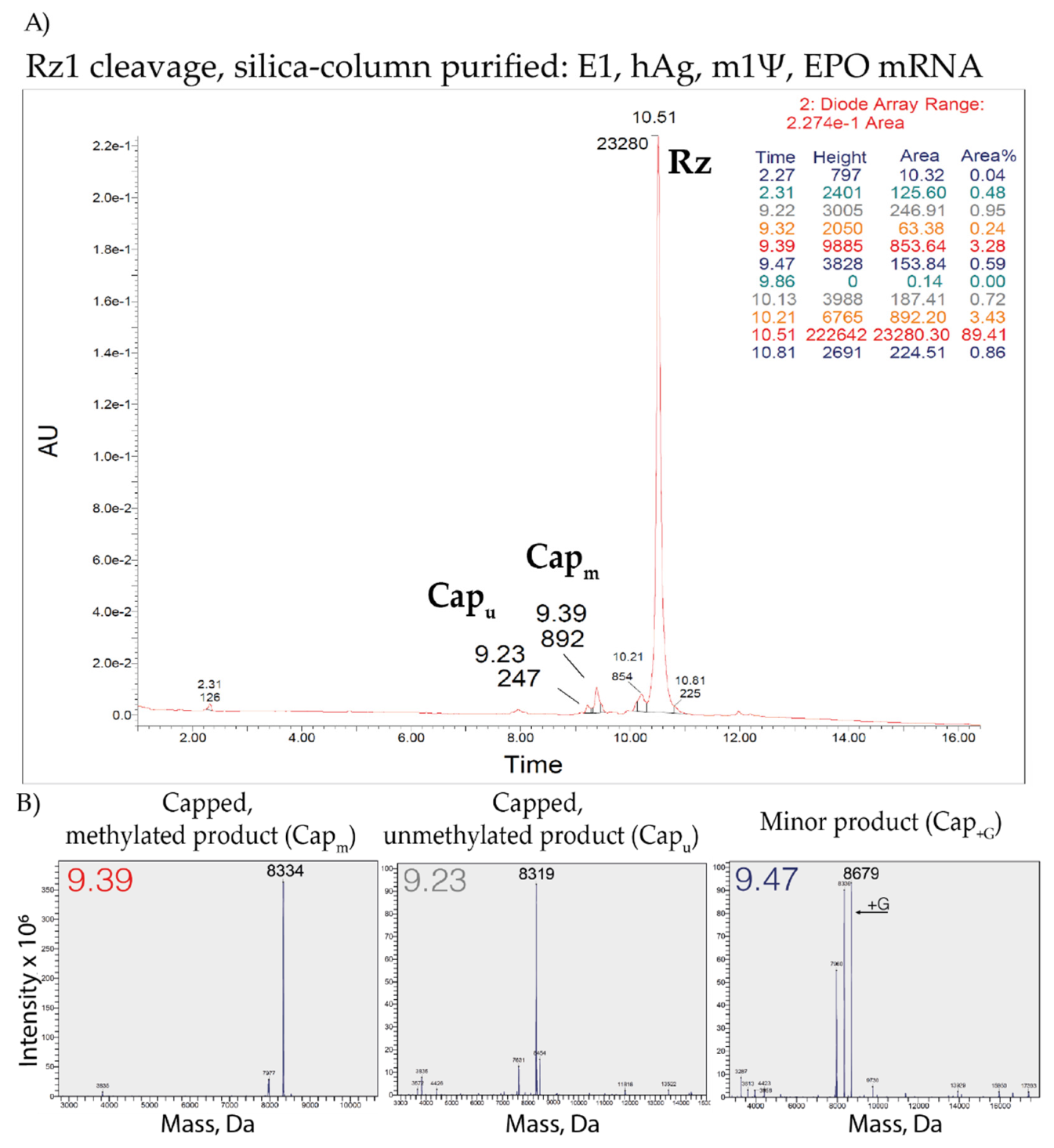
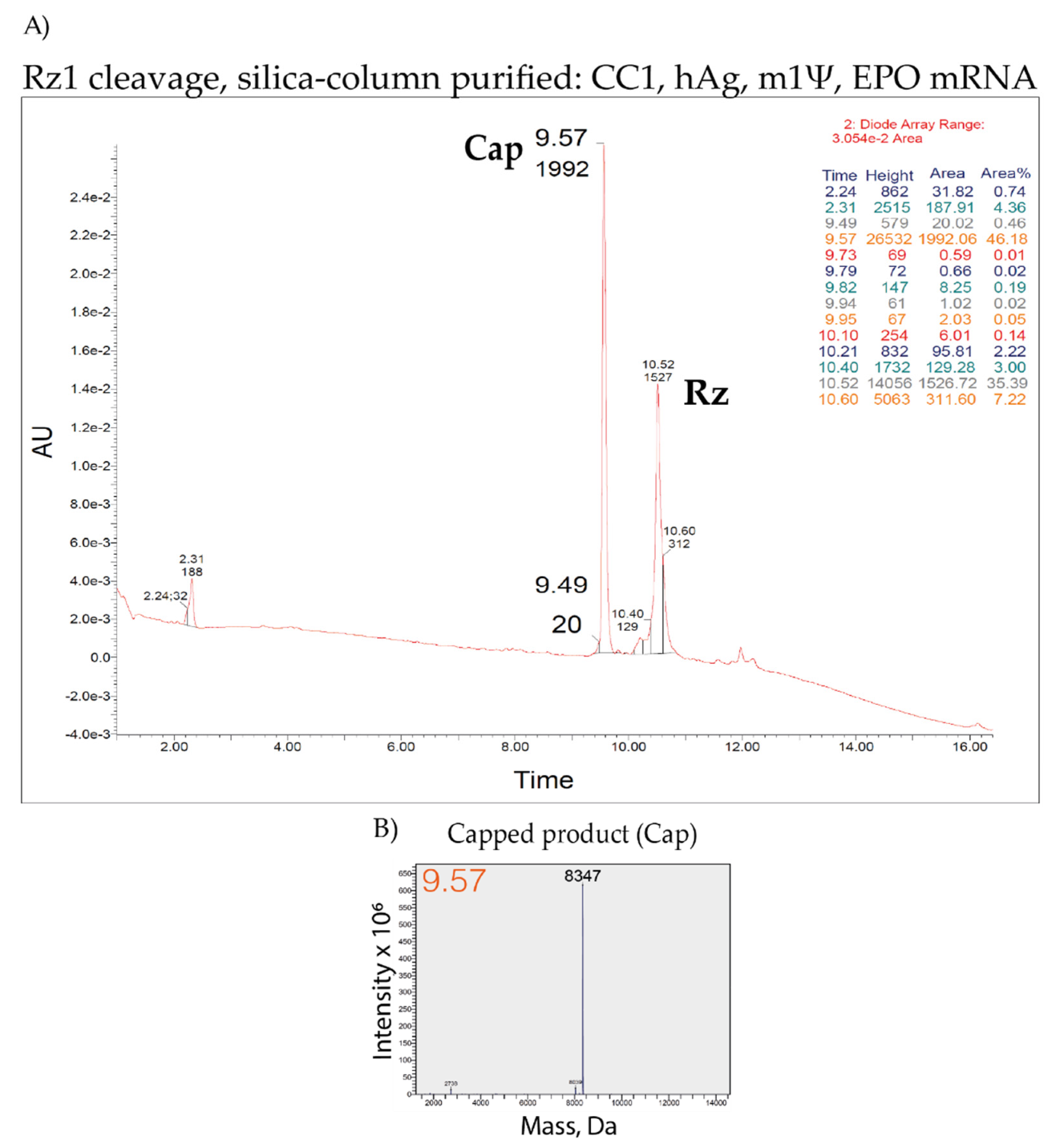
3.7. LC–MS Analysis of Capping Efficiency Using the Ribozyme-Mediated Cleavage Assay
- 68% of the expected major product(7MeGpppA(OMe)GGCGAACU*AGU*AU*U*CU*U*CU*GGU*C > p);
- 20% unmethylated product(7MeGpppAGGCGAACU*AGU*AU*U*CU*U*CU*GGU*C > p);
- and 1% additional product (+G).
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Sahin, U.; Karikó, K.; Türeci, Ö. mRNA-based therapeutics—Developing a new class of drugs. Nat. Rev. Drug Discov. 2014, 13, 759–780. [Google Scholar] [CrossRef]
- Sahin, U.; Derhovanessian, E.; Miller, M.; Kloke, B.-P.; Simon, P.; Löwer, M.; Bukur, V.; Tadmor, A.D.; Luxemburger, U.; Schrörs, B.; et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature 2017, 547, 222–226. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Porter, F.W.; Weissman, D. mRNA vaccines—A new era in vaccinology. Nat. Rev. Drug Discov. 2018, 17, 261–279. [Google Scholar] [CrossRef] [Green Version]
- Polack, F.P.; Thomas, S.J.; Kitchin, N.; Absalon, J.; Gurtman, A.; Lockhart, S.; Perez, J.L.; Pérez Marc, G.; Moreira, E.D.; Zerbini, C.; et al. Safety and efficacy of the BNT162b2 mRNA COVID-19 vaccine. N. Engl. J. Med. 2020, 383, 2603–2615. [Google Scholar] [CrossRef]
- Anderson, E.J.; Rouphael, N.G.; Widge, A.T.; Jackson, L.A.; Roberts, P.C.; Makhene, M.; Chappell, J.D.; Denison, M.R.; Stevens, L.J.; Pruijssers, A.J.; et al. Safety and Immunogenicity of SARS-CoV-2 mRNA-1273 Vaccine in Older Adults. N. Engl. J. Med. 2020, 383, 2427–2438. [Google Scholar] [CrossRef]
- Vlatkovic, I. Non-Immunotherapy Application of LNP-mRNA: Maximizing Efficacy and Safety. Biomedicines 2021, 9, 530. [Google Scholar] [CrossRef]
- Ramanathan, A.; Robb, G.B.; Chan, S.-H. mRNA capping: Biological functions and applications. Nucleic Acids Res. 2016, 44, 7511–7526. [Google Scholar] [CrossRef]
- Shimotohno, K.; Kodama, Y.; Hashimoto, J.; Miura, K.I. Importance of 5’-terminal blocking structure to stabilize mRNA in eukaryotic protein synthesis. Proc. Natl. Acad. Sci. USA 1977, 74, 2734–2738. [Google Scholar] [CrossRef] [Green Version]
- Paterson, B.M.; Rosenberg, M. Efficient translation of prokaryotic mRNAs in a eukaryotic cell-free system requires addition of a cap structure. Nature 1979, 279, 692–696. [Google Scholar] [CrossRef]
- Stepinski, J.; Waddell, C.; Stolarski, R.; Darzynkiewicz, E.; Rhoads, R.E. Synthesis and properties of mRNAs containing the novel "anti-reverse" cap analogs 7-methyl(3’-O-methyl)GpppG and 7-methyl (3’-deoxy)GpppG. RNA 2001, 7, 1486–1495. [Google Scholar]
- Grudzien, E.; Stepinski, J.; Jankowska-Anyszka, M.; Stolarski, R.; Darzynkiewicz, E.; Rhoads, R.E. Novel cap analogs for in vitro synthesis of mRNAs with high translational efficiency. RNA 2004, 10, 1479–1487. [Google Scholar] [CrossRef] [Green Version]
- Anderson, B.R.; Muramatsu, H.; Nallagatla, S.R.; Bevilacqua, P.C.; Sansing, L.H.; Weissman, D.; Karikó, K. Incorporation of pseudouridine into mRNA enhances translation by diminishing PKR activation. Nucleic Acids Res. 2010, 38, 5884–5892. [Google Scholar] [CrossRef] [Green Version]
- Beverly, M.; Dell, A.; Parmar, P.; Houghton, L. Label-free analysis of mRNA capping efficiency using RNase H probes and LC-MS. Anal. Bioanal. Chem. 2016, 408, 5021–5030. [Google Scholar] [CrossRef]
- Moya-Ramírez, I.; Bouton, C.; Kontoravdi, C.; Polizzi, K. High resolution biosensor to test the capping level and integrity of mRNAs. Nucleic Acids Res. 2020, 48, e129. [Google Scholar] [CrossRef]
- von Niessen, A.G.O.; Poleganov, M.A.; Rechner, C.; Plaschke, A.; Kranz, L.; Fesser, S.; Diken, M.; Löwer, M.; Vallazza, B.; Beissert, T.; et al. Improving mRNA-Based Therapeutic Gene Delivery by Expression-Augmenting 3′ UTRs Identified by Cellular Library Screening. Mol. Ther. 2018, 27, 824–836. [Google Scholar] [CrossRef] [Green Version]
- Karikó, K.; Muramatsu, H.; A Welsh, F.; Ludwig, J.; Kato, H.; Akira, S.; Weissman, D. Incorporation of Pseudouridine Into mRNA Yields Superior Nonimmunogenic Vector With Increased Translational Capacity and Biological Stability. Mol. Ther. 2008, 16, 1833–1840. [Google Scholar] [CrossRef]
- Kowalska, J.; Lewdorowicz, M.; Zuberek, J.; Bojarska, E.; Wojcik, J.; Cohen, L.S.; Davis, R.E.; Stepinski, J.; Stolarski, R.; Darzynkiewicz, E.; et al. Synthesis and properties of mRNA cap analogs containing phosphorothioate moiety in 5’,5’-triphosphate chain. Nucleosides Nucleotides Nucleic Acids 2005, 24, 595–600. [Google Scholar] [CrossRef]
- Mašek, T.; Vopalensky, V.; Suchomelova, P.; Pospisek, M. Denaturing RNA electrophoresis in TAE agarose gels. Anal. Biochem. 2005, 336, 46–50. [Google Scholar] [CrossRef]
- Karikó, K.; Megyeri, K.; Xiao, Q.; Barnathan, E.S. Lipofectin-aided cell delivery of ribozyme targeted to human urokinase receptor mRNA. FEBS Lett. 1994, 352, 41–44. [Google Scholar] [CrossRef] [Green Version]
- Li, F.; Barnathan, E.S.; Karikó, K. Rapid method for screening and cloning cDNAs generated in differential mRNA display: Application of Northern blot for affinity capturing of cDNAs. Nucleic Acids Res. 1994, 22, 1764–1765. [Google Scholar] [CrossRef] [Green Version]
- Ludwig, J.; Blaschke, M.; Sproat, B.S. Extending the cleavage rules for the hammerhead ribozyme: Mutating adenosine15.1 to inosine15.1 changes the cleavage site specificity from N16.2U16.1H17 to N16.2C16.1H17. Nucleic Acids Res. 1998, 26, 2279–2285. [Google Scholar] [CrossRef] [Green Version]
- Nilsen, T.W. Gel Purification of RNA. Cold Spring Harb. Protoc. 2013, 2013, 180–183. [Google Scholar] [CrossRef] [Green Version]
- Karikó, K.; Kuo, A.; Barnathan, E.S. Overexpression of urokinase receptor in mammalian cells following administration of the in vitro transcribed encoding mRNA. Gene Ther. 1999, 6, 1092–1100. [Google Scholar] [CrossRef] [Green Version]
- Holtkamp, S.; Kreiter, S.; Selmi, A.; Simon, P.; Koslowski, M.; Huber, C.; Türeci, O.; Sahin, U. Modification of antigen-encoding RNA increases stability, translational efficacy, and T-cell stimulatory capacity of dendritic cells. Blood 2006, 108, 4009–4017. [Google Scholar] [CrossRef]
- Scott, W.G.; Horan, L.H.; Martick, M. The Hammerhead Ribozyme: Structure, catalysis, and gene regulation. Prog. Mol. Biol. Transl. Sci. 2013, 120, 1–23. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takagi, Y.; Taira, K. Temperature-dependent change in the rate-determining step in a reaction catalyzed by a hammerhead ribozyme. FEBS Lett. 1995, 361, 273–276. [Google Scholar] [CrossRef] [Green Version]
- Sawata, S.; Shimayama, T.; Komiyama, M.; Kumar, P.K.R.; Nishikawa, S.; Taira, K. Enhancement of the cleavage rates of DNA-armed hammerhead ribozymes by various divalent metal ions. Nucleic Acids Res. 1993, 21, 5656–5660. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| 5′UTR | Rz | Rz Sequence (5′-3′) |
|---|---|---|
| hAg | Rz1 | UGU GGG CUG AUG AGG CCG UGA GGC CGA AAC CAG AAG AAU |
| Rz2 | GGG GAC CAG AAG AAC UGA UGA GGC CGU GAG GCC GAA ACUm AmGmUm UmCmGm | |
| Rz3 | UGU GGG CUG AUG AGG CCG UGA GGC CGA AAC CAG AA | |
| Rz4 | AGU CUG UGG GCU GAU GAG GCC GUG AGG CCG AAA CCA GAA GAA | |
| TEV | Rz5 | GUA UAC UGA UGA GGC CGU GAG GCC GAA IUmUm GmUmGm UmUmGm AmGmAm CmUmAm GmUmUm UmAm |
| 5′UTR | Rz | Rz (nt) | Example of Uncapped 5′CP (Capped: +7MeG) | 5′CP (nt) | |
|---|---|---|---|---|---|
| −Cap | +Cap | ||||
| hAg | Rz1 | 39 | pppGCGAACUAGUAUUCUUCUGGUC > CCCACAGACU… | 22 | 23 |
| Rz2 | 45 | pppGCGAACUAGUA > UUCUUCUGGUCCCCACAGACU... | 11 | 12 | |
| Rz3 | 35 | pppGCGAACUAGUAUUCUUCUGGUC > CCCACAGACU… | 22 | 23 | |
| Rz4 | 42 | pppGCGAACUAGUAUUCUUCUGGUC > CCCACAGACU… | 22 | 23 | |
| TEV | Rz5 | 47 | pppGGAAUAAACUAGUCUCAACACAACA > UAUACAAA... | 25 | 26 |
| 5′UTR | hAg | TEV | |||||
|---|---|---|---|---|---|---|---|
| Purification | Unpurified | Purified | Unpurified | Purified | |||
| Assay | Rz1 | Rz2 | Rz1 | Rz2 | Rz5 | ||
| Cap | Modification | Capping Efficiency (%) | |||||
| E0 | U | 100 | 86 | 90 | 95 | 93 | 89 |
| m1Ψ | 88 | 100 | 95 | 96 | 100 | 99 | |
| E1 | U | 100 | 84 | 91 | 95 | 95 | 90 |
| m1Ψ | 92 | 100 | 95 | 96 | 98 | 95 | |
| A0 | U | 52 | 51 | 52 | 52 | 75 | 67 |
| m1Ψ | 40 | 48 | 53 | 34 | 77 | 69 | |
| 5′UTR | Probe Name | Probe Sequence (5′-3′) | Probe Size (nt) | (1) 5′CP Sequence w/o Cap | 5′CP Size (nt) | |
|---|---|---|---|---|---|---|
| (2) Minor Cleavage Product | −Cap | +Cap | ||||
| hAg | Probe 1(P1) | GACCAGAmAmGmAmAmUmAmCmUmAm | 16 |
| 20 | 21 |
| 21 | 22 | ||||
| 5’UTR | hAg | ||
|---|---|---|---|
| Purification | Unpurified | Purified | |
| Cap | Modification | Capping Efficiency (%) | |
| E0 | U | 75 | 82 |
| m1Ψ | 89 | 85 | |
| E1 | U | 82 | 78 |
| m1Ψ | 82 | 83 | |
| A0 | U | 56 | 47 |
| m1Ψ | 40 | 37 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Vlatkovic, I.; Ludwig, J.; Boros, G.; Szabó, G.T.; Reichert, J.; Buff, M.; Baiersdörfer, M.; Reinholz, J.; Mahiny, A.J.; Şahin, U.; et al. Ribozyme Assays to Quantify the Capping Efficiency of In Vitro-Transcribed mRNA. Pharmaceutics 2022, 14, 328. https://doi.org/10.3390/pharmaceutics14020328
Vlatkovic I, Ludwig J, Boros G, Szabó GT, Reichert J, Buff M, Baiersdörfer M, Reinholz J, Mahiny AJ, Şahin U, et al. Ribozyme Assays to Quantify the Capping Efficiency of In Vitro-Transcribed mRNA. Pharmaceutics. 2022; 14(2):328. https://doi.org/10.3390/pharmaceutics14020328
Chicago/Turabian StyleVlatkovic, Irena, János Ludwig, Gábor Boros, Gábor Tamás Szabó, Julia Reichert, Maximilian Buff, Markus Baiersdörfer, Jonas Reinholz, Azita Josefine Mahiny, Uğur Şahin, and et al. 2022. "Ribozyme Assays to Quantify the Capping Efficiency of In Vitro-Transcribed mRNA" Pharmaceutics 14, no. 2: 328. https://doi.org/10.3390/pharmaceutics14020328