
Engineering of a Fully Human Anti-MUC-16 Antibody and Evaluation as a PET Imaging Agent
 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Cloning of Domains SEA11-12 of MUC16
- HB103 (5′ to 3′)
- GCATTGCACTAAGTCTTGCACTTGTCACGAATTCGATAAATGGTTTCACCCAGCGG
- HB104 (5′ to 3′)
- GGCATGTGTGAGTTTTGTCAGATCTAACCATGGCCGATGATAAATTCTGGGGTGCATAGC
2.2. Expression and Purification of Recombinant SEA11-12-Fc Protein
2.3. Screening Naïve Antibody Libraries Using Phage Display
2.4. Expression of M16Ab Antibody in IgG1 Format
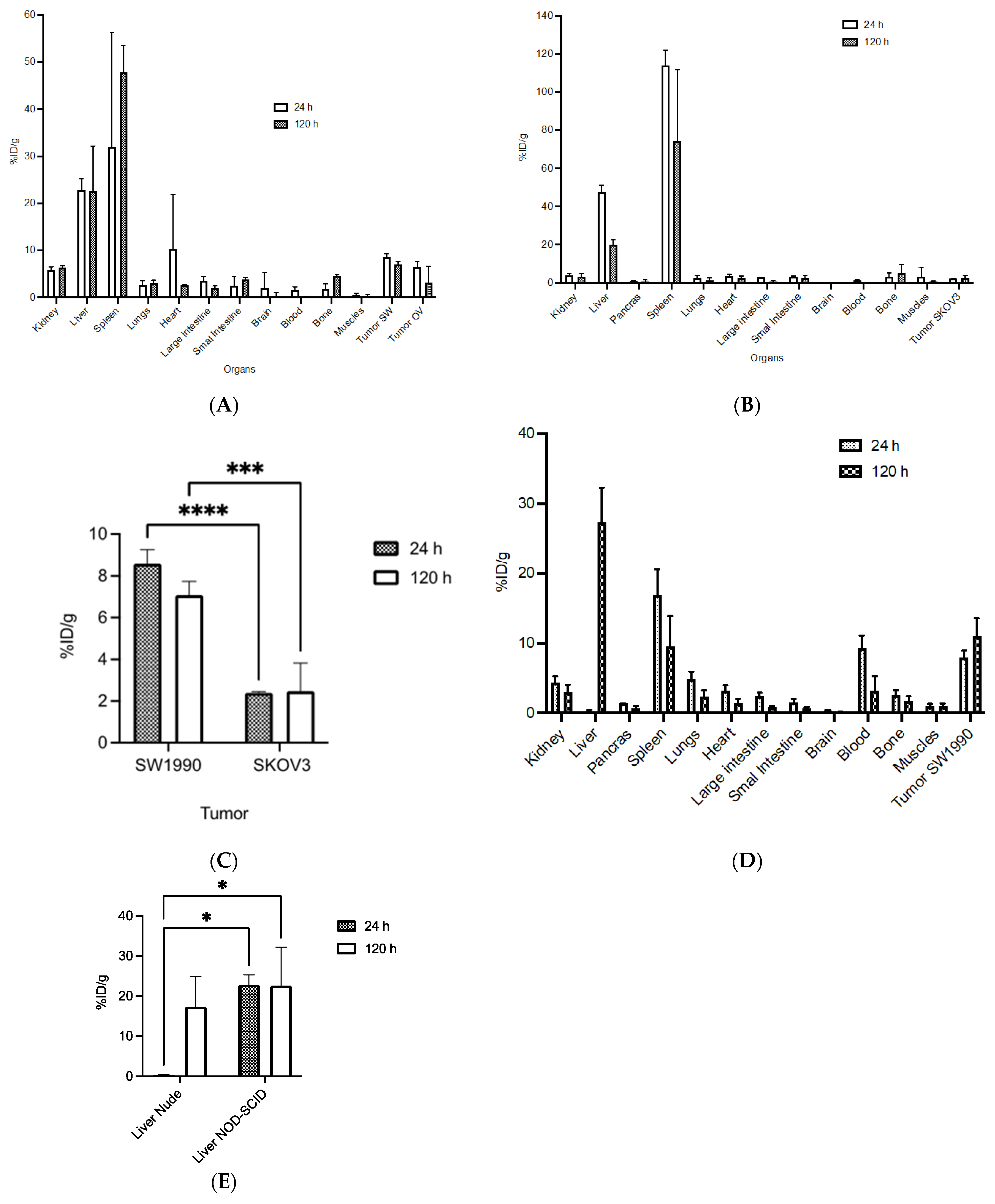
2.5. Cell Lines and Mouse Xenograft Models
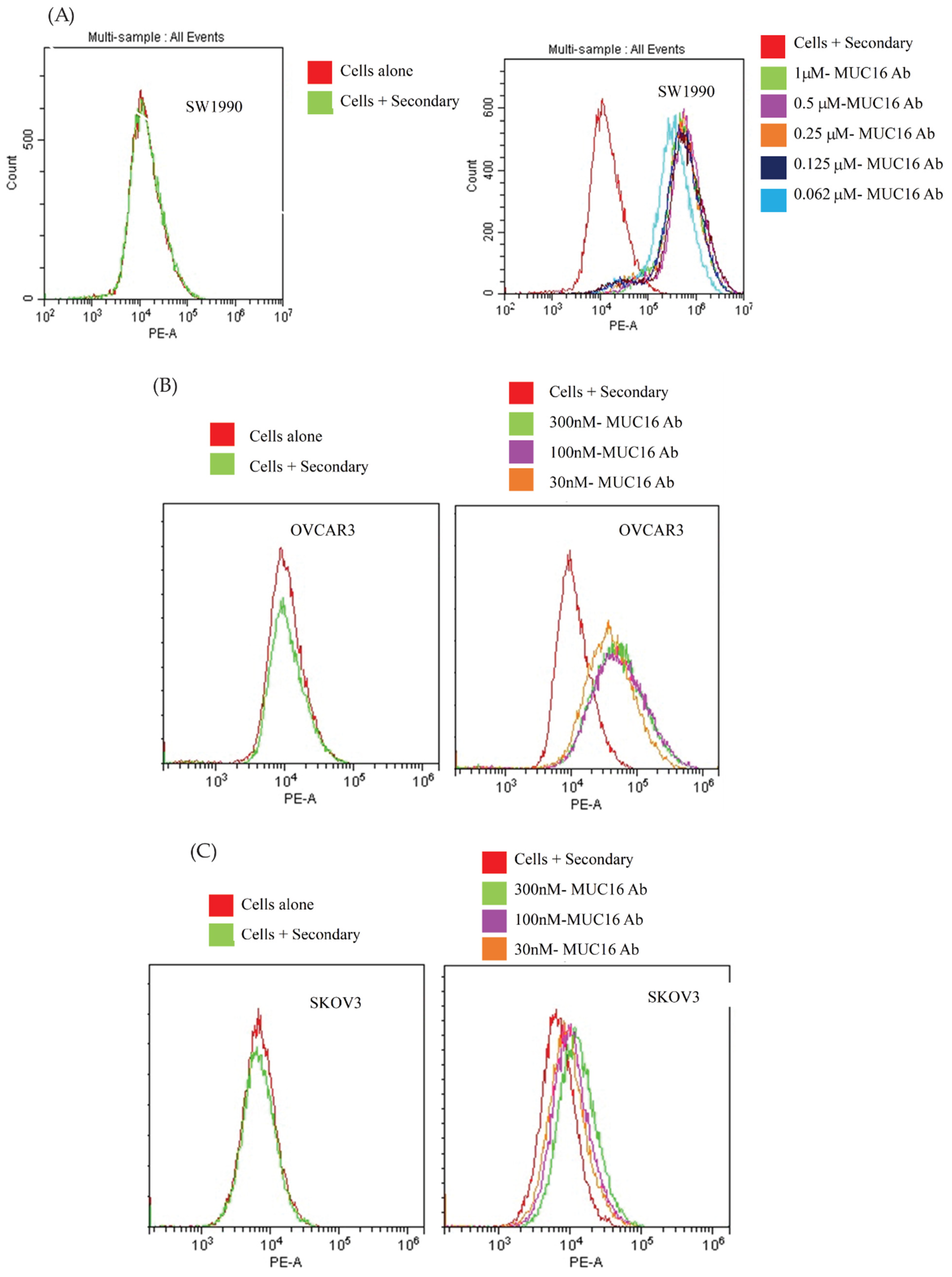
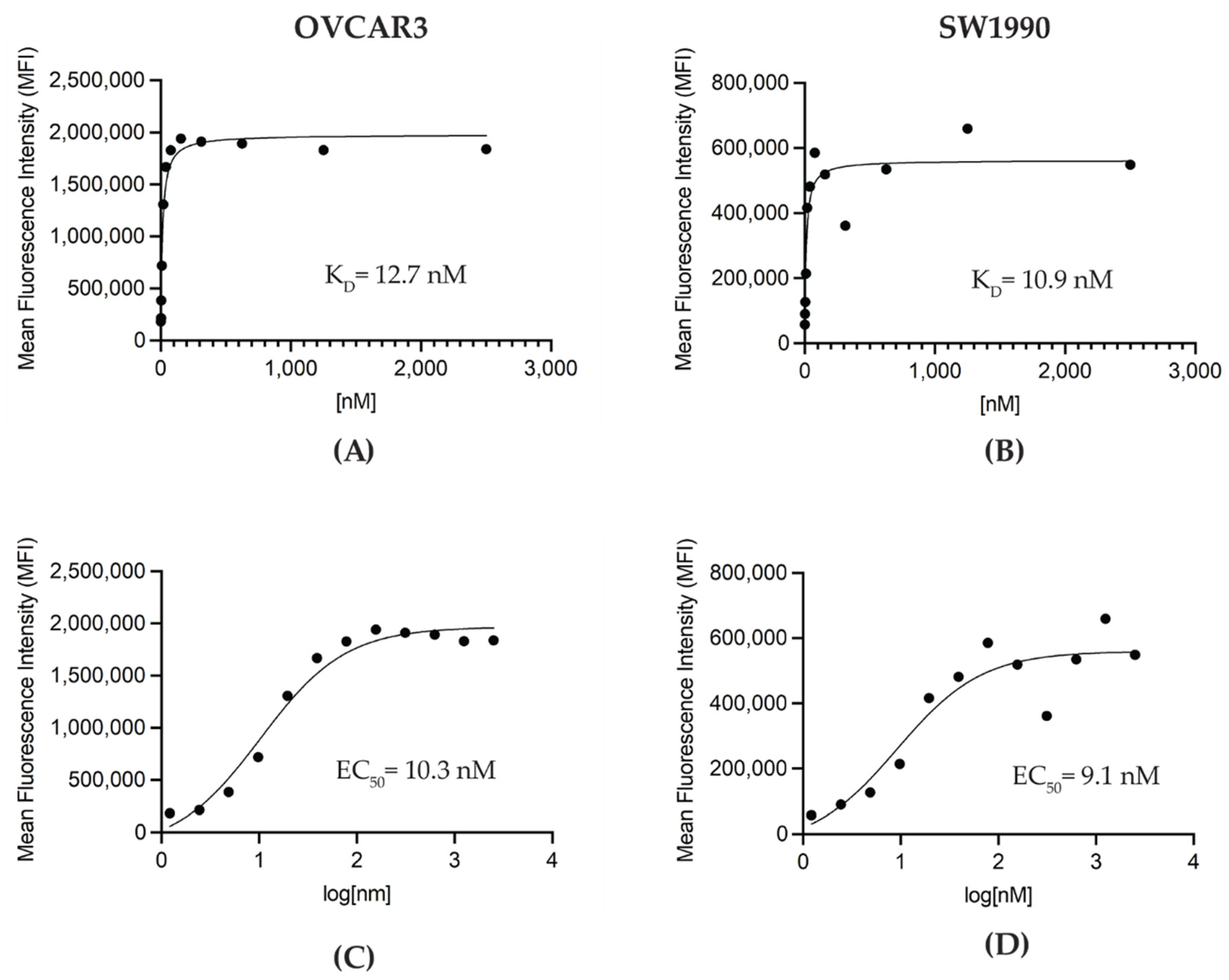
2.6. Flow Cytometry
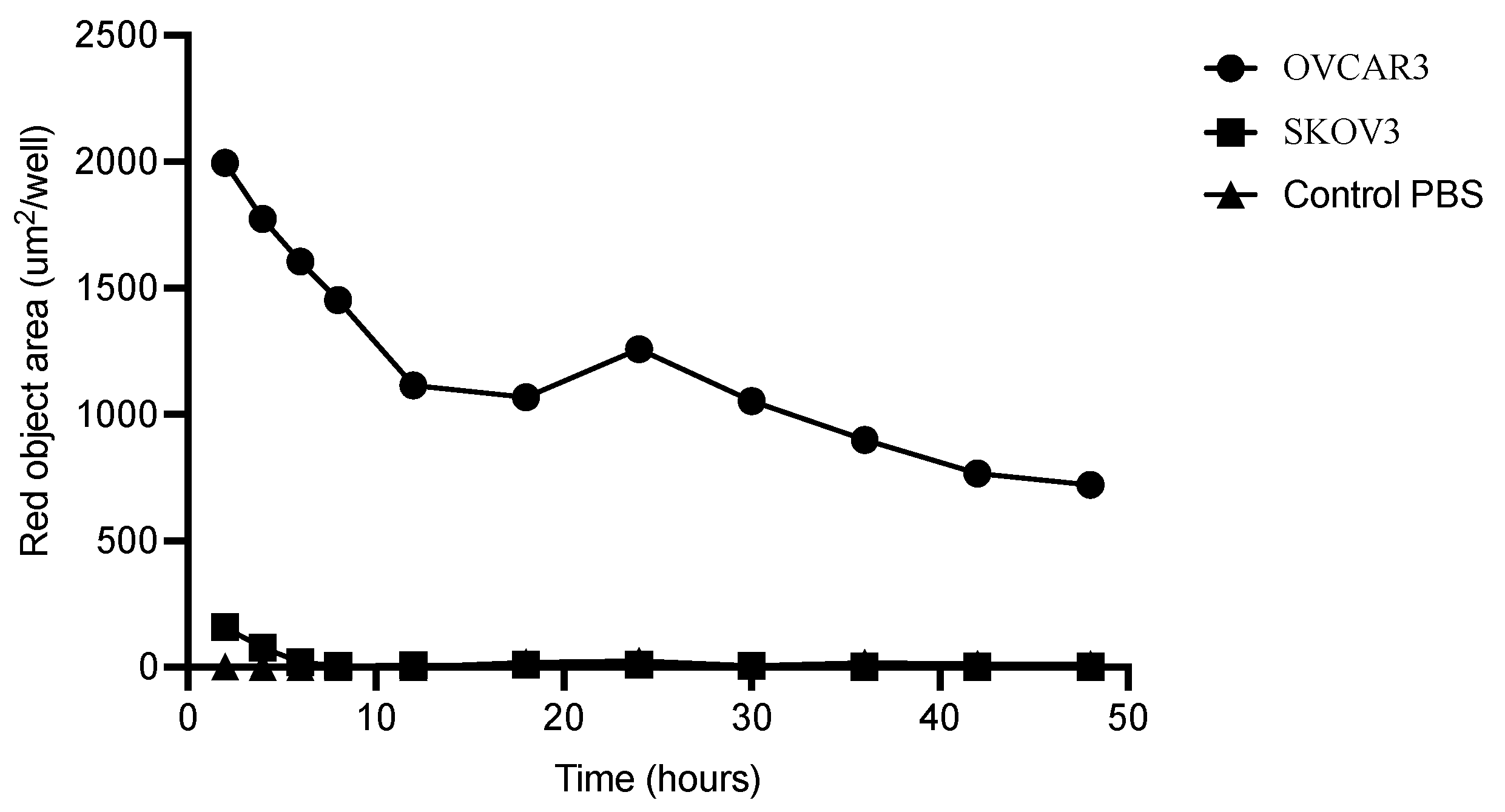
2.7. Antibody Internalization
2.8. Conjugation of Anti-MUC-16 Ab with p-SCN-Bz-Deferoxamine
2.9. Radiolabeling with 89Zr
2.10. Stability of 89Zr-MUC-16 Ab1
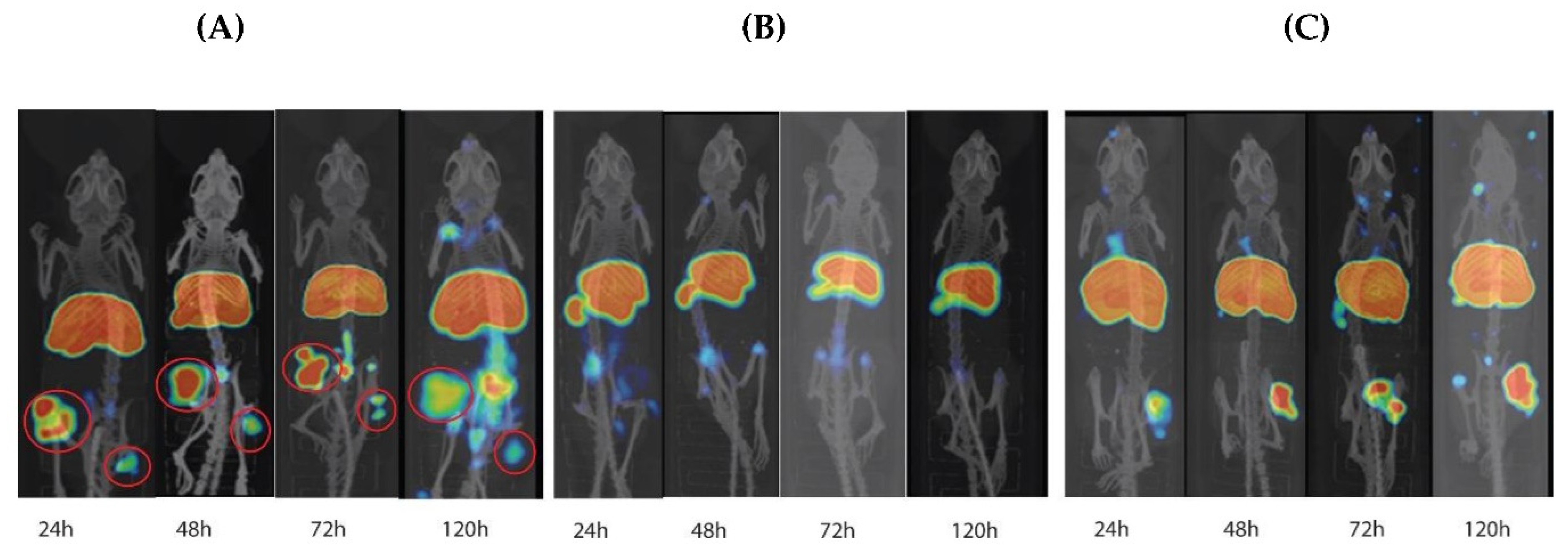
2.11. MicroPET/CT Imaging and Biodistribution
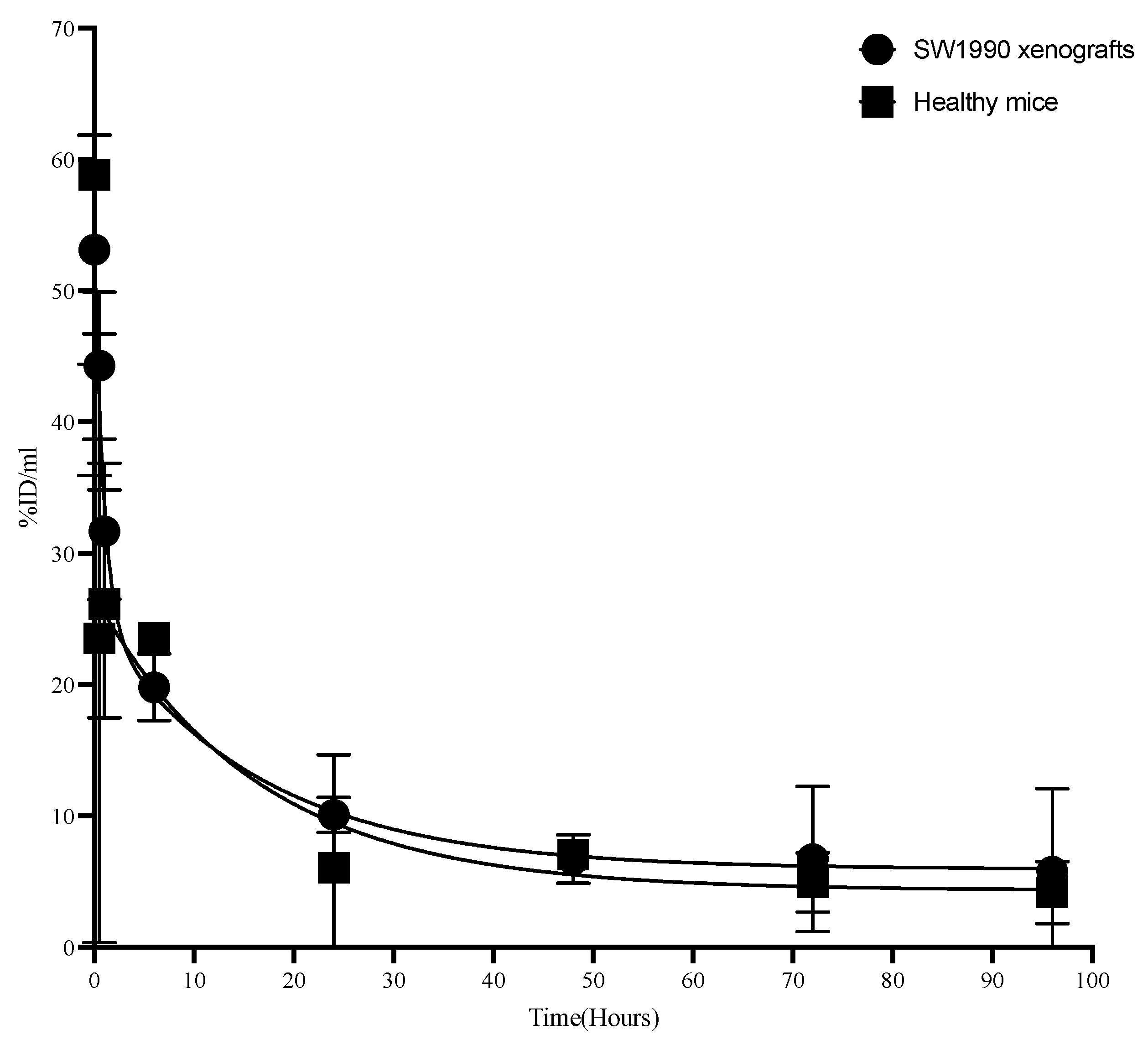
2.12. Pharmacokinetics
2.13. Statistical Analysis
3. Results
3.1. Generation and In Vitro Characterization of M16Ab Antibody
3.2. Conjugation of M16Ab and Quality Control of Immunoconjugate
3.3. Internalization of M16Ab
3.4. Radiolabeling and Characterization
3.5. Pharmacokinetics of M16Ab in CD-1 Mice
3.6. Biodistribution and MicroPET/CT Imaging in Tumor-Bearing Mice
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Lheureux, S.; Gourley, C.; Vergote, I.; Oza, A.M. Epithelial ovarian cancer. Lancet 2019, 393, 1240–1253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corrado, G.; Salutari, V.; Palluzzi, E.; Distefano, M.G.; Scambia, G.; Ferrandina, G. Optimizing treatment in recurrent epithelial ovarian cancer. Expert Rev. Anticancer Ther. 2017, 17, 1147–1158. [Google Scholar] [CrossRef] [PubMed]
- Gupta, N.; Yelamanchi, R. Pancreatic adenocarcinoma: A review of recent paradigms and advances in epidemiology, clinical diagnosis and management. World J. Gastroenterol. 2021, 27, 3158–3181. [Google Scholar] [CrossRef] [PubMed]
- Elsayed, M.; Abdelrahim, M. The Latest Advancement in Pancreatic Ductal Adenocarcinoma Therapy: A Review Article for the Latest Guidelines and Novel Therapies. Biomedicines 2021, 9, 389. [Google Scholar] [CrossRef]
- Corfield, A.P. Mucins: A biologically relevant glycan barrier in mucosal protection. Biochim. Biophys. Acta 2015, 1850, 236–252. [Google Scholar] [CrossRef]
- Kufe, D.W. Mucins in cancer: Function, prognosis and therapy. Nat. Rev. Cancer 2009, 9, 874–885. [Google Scholar] [CrossRef] [Green Version]
- van Putten, J.P.M.; Strijbis, K. Transmembrane Mucins: Signaling Receptors at the Intersection of Inflammation and Cancer. J. Innate Immun. 2017, 9, 281–299. [Google Scholar] [CrossRef]
- Reynolds, I.S.; Fichtner, M.; McNamara, D.A.; Kay, E.W.; Prehn, J.H.M.; Burke, J.P. Mucin glycoproteins block apoptosis; promote invasion, proliferation, and migration; and cause chemoresistance through diverse pathways in epithelial cancers. Cancer Metastasis Rev. 2019, 28, 237–257. [Google Scholar] [CrossRef]
- Bast, R.C., Jr.; Klug, T.L.; St John, E.; Jenison, E.; Niloff, J.M.; Lazarus, H.; Berkowitz, R.S.; Leavitt, T.; Griffiths, C.T.; Parker, L.; et al. A radioimmunoassay using a monoclonal antibody to monitor the course of epithelial ovarian cancer. N. Engl. J. Med. 1983, 309, 883–887. [Google Scholar] [CrossRef]
- Cohen, J.G.; White, M.; Cruz, A.; Farias-Eisner, R. In 2014, can we do better than CA125 in the early detection of ovarian cancer? World J. Biol. Chem. 2014, 5, 286–300. [Google Scholar] [CrossRef]
- Kim, N.; Hong, Y.; Kwon, D.; Yoon, S. Somatic mutaome profile in human cancer tissues. Genom. Inform. 2013, 11, 239–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gubbels, J.A.; Belisle, J.; Onda, M.; Rancourt, C.; Migneault, M.; Ho, M.; Bera, T.K.; Connor, J.; Sathyanarayana, B.K.; Lee, B.; et al. Mesothelin-MUC16 binding is a high affinity, N-glycan dependent interaction that facilitates peritoneal metastasis of ovarian tumors. Mol. Cancer 2006, 5, 50. [Google Scholar] [CrossRef] [Green Version]
- Theriault, C.; Pinard, M.; Comamala, M.; Migneault, M.; Beaudin, J.; Matte, I.; Boivin, M.; Piche, A.; Rancourt, C. MUC16 (CA125) regulates epithelial ovarian cancer cell growth, tumorigenesis and metastasis. Gynecol. Oncol. 2011, 121, 434–443. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.H.; Hung, W.C.; Wang, P.; Paul, C.; Konstantopoulos, K. Mesothelin binding to CA125/MUC16 promotes pancreatic cancer cell motility and invasion via MMP-7 activation. Sci. Rep. 2013, 3, 1870. [Google Scholar] [CrossRef] [Green Version]
- Higashi, M.; Yamada, N.; Yokoyama, S.; Kitamoto, S.; Tabata, K.; Koriyama, C.; Batra, S.K.; Yonezawa, S. Pathobiological implications of MUC16/CA125 expression in intrahepatic cholangiocarcinoma-mass forming type. Pathobiology 2012, 79, 101–106. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Qin, Y.; Zhang, B.; Ji, S.; Shi, S.; Xu, W.; Liu, J.; Xiang, J.; Liang, D.; Hu, Q.; et al. Oncogenic KRAS Targets MUC16/CA125 in Pancreatic Ductal Adenocarcinoma. Mol. Cancer Res. 2017, 15, 201–212. [Google Scholar] [CrossRef] [Green Version]
- Shimizu, A.; Hirono, S.; Tani, M.; Kawai, M.; Okada, K.; Miyazawa, M.; Kitahata, Y.; Nakamura, Y.; Noda, T.; Yokoyama, S.; et al. Coexpression of MUC16 and mesothelin is related to the invasion process in pancreatic ductal adenocarcinoma. Cancer Sci. 2012, 103, 739–746. [Google Scholar] [CrossRef]
- Haridas, D.; Chakraborty, S.; Ponnusamy, M.P.; Lakshmanan, I.; Rachagani, S.; Cruz, E.; Kumar, S.; Das, S.; Lele, S.M.; Anderson, J.M.; et al. Pathobiological implications of MUC16 expression in pancreatic cancer. PLoS ONE 2011, 6, e26839. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Clark, S.; Wong, T.; Chen, Y.; Chen, Y.; Dennis, M.S.; Luis, E.; Zhong, F.; Bheddah, S.; Koeppen, H.; et al. Armed antibodies targeting the mucin repeats of the ovarian cancer antigen, MUC16, are highly efficacious in animal tumor models. Cancer Res. 2007, 67, 4924–4932. [Google Scholar] [CrossRef] [Green Version]
- Junutula, J.R.; Raab, H.; Clark, S.; Bhakta, S.; Leipold, D.D.; Weir, S.; Chen, Y.; Simpson, M.; Tsai, S.P.; Dennis, M.S.; et al. Site-specific conjugation of a cytotoxic drug to an antibody improves the therapeutic index. Nat. Biotechnol. 2008, 26, 925–932. [Google Scholar] [CrossRef]
- Marcos-Silva, L.; Ricardo, S.; Chen, K.; Blixt, O.; Arigi, E.; Pereira, D.; Hogdall, E.; Mandel, U.; Bennett, E.P.; Vakhrushev, S.Y.; et al. A novel monoclonal antibody to a defined peptide epitope in MUC16. Glycobiology 2015, 25, 1172–1182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nemieboka, B.; Sharma, S.K.; Rao, T.D.; Edwards, K.J.; Yan, S.; Wang, P.; Ragupathi, A.; Piersigilli, A.; Spriggs, D.R.; Lewis, J.S. Radiopharmacologic screening of antibodies to the unshed ectodomain of MUC16 in ovarian cancer identifies a lead candidate for clinical translation. Nucl. Med. Biol. 2020, 86-87, 9–19. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Mack, K.N.; Piersigilli, A.; Pourat, J.; Edwards, K.J.; Keinanen, O.; Jiao, M.S.; Zhao, H.; White, B.; Brooks, C.L.; et al. ImmunoPET of Ovarian and Pancreatic Cancer with AR9.6, a Novel MUC16-Targeted Therapeutic Antibody. Clin. Cancer Res. 2022, 28, 948–959. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.K.; Sevak, K.K.; Monette, S.; Carlin, S.D.; Knight, J.C.; Wuest, F.R.; Sala, E.; Zeglis, B.M.; Lewis, J.S. Preclinical 89Zr Immuno-PET of High-Grade Serous Ovarian Cancer and Lymph Node Metastasis. J. Nucl. Med. 2016, 57, 771–776. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Olson, M.T.; Wojtynek, N.E.; Talmon, G.A.; Caffrey, T.C.; Radhakrishnan, P.; Ly, Q.P.; Hollingsworth, M.A.; Mohs, A.M. Development of a MUC16-Targeted Near-Infrared Fluorescent Antibody Conjugate for Intraoperative Imaging of Pancreatic Cancer. Mol. Cancer Ther. 2020, 19, 1670–1681. [Google Scholar] [CrossRef] [PubMed]
- Broqueza, J.; Prabaharan, C.B.; Andrahennadi, S.; Allen, K.J.H.; Dickinson, R.; MacDonald-Dickinson, V.; Dadachova, E.; Uppalapati, M. Novel Human Antibodies to Insulin Growth Factor 2 Receptor (IGF2R) for Radioimmunoimaging and Therapy of Canine and Human Osteosarcoma. Cancers 2021, 13, 2208. [Google Scholar] [CrossRef] [PubMed]
- Fellouse, F.A.; Sidhu, S.S. Making Antibodies in Bacteria. In Making and Using Antibodies; Howard, G.C., Kaser, M.R., Eds.; CRC Press: Boca Raton, FL, USA, 2006. [Google Scholar]
- Vazquez-Lombardi, R.; Nevoltris, D.; Luthra, A.; Schofield, P.; Zimmermann, C.; Christ, D. Transient expression of human antibodies in mammalian cells. Nat. Protoc. 2018, 13, 99–117. [Google Scholar] [CrossRef]
- Alizadeh, E.; Behlol Ayaz Ahmed, K.; Raja Solomon, V.; Gaja, V.; Bernhard, W.; Makhlouf, A.; Gonzalez, C.; Barreto, K.; Casaco, A.; Geyer, C.R.; et al. (89)Zr-Labeled Domain II-Specific scFv-Fc ImmunoPET Probe for Imaging Epidermal Growth Factor Receptor In Vivo. Cancers 2021, 13, 560. [Google Scholar] [CrossRef]
- Gaja, V.; Cawthray, J.; Geyer, C.R.; Fonge, H. Production and Semi-Automated Processing of (89)Zr Using a Commercially Available TRASIS MiniAiO Module. Molecules 2020, 25, 2626. [Google Scholar] [CrossRef]
- Yang, Z.J.; Zhao, B.B.; Li, L. The significance of the change pattern of serum CA125 level for judging prognosis and diagnosing recurrences of epithelial ovarian cancer. J. Ovarian Res. 2016, 9, 57. [Google Scholar] [CrossRef]
- Giampaolino, P.; Foreste, V.; Della Corte, L.; Di Filippo, C.; Iorio, G.; Bifulco, G. Role of biomarkers for early detection of ovarian cancer recurrence. Gland Surg. 2020, 9, 1102–1111. [Google Scholar] [CrossRef] [PubMed]
- Buamah, P. Benign conditions associated with raised serum CA-125 concentration. J. Surg. Oncol. 2000, 75, 264–265. [Google Scholar] [CrossRef] [PubMed]
- Lovqvist, A.; Humm, J.L.; Sheikh, A.; Finn, R.D.; Koziorowski, J.; Ruan, S.; Pentlow, K.S.; Jungbluth, A.; Welt, S.; Lee, F.T.; et al. PET imaging of (86)Y-labeled anti-Lewis Y monoclonal antibodies in a nude mouse model: Comparison between (86)Y and (111)In radiolabels. J. Nucl. Med. 2001, 42, 1281–1287. [Google Scholar] [PubMed]
- Wright, B.D.; Lapi, S.E. Designing the magic bullet? The advancement of immuno-PET into clinical use. J. Nucl. Med. 2013, 54, 1171–1174. [Google Scholar] [CrossRef] [Green Version]
- Harding, F.A.; Stickler, M.M.; Razo, J.; DuBridge, R.B. The immunogenicity of humanized and fully human antibodies: Residual immunogenicity resides in the CDR regions. MAbs 2010, 2, 256–265. [Google Scholar] [CrossRef] [Green Version]
- Deri, M.A.; Zeglis, B.M.; Francesconi, L.C.; Lewis, J.S. PET imaging with (8)(9)Zr: From radiochemistry to the clinic. Nucl. Med. Biol. 2013, 40, 3–14. [Google Scholar] [CrossRef] [Green Version]
- Sharma, S.K.; Chow, A.; Monette, S.; Vivier, D.; Pourat, J.; Edwards, K.J.; Dilling, T.R.; Abdel-Atti, D.; Zeglis, B.M.; Poirier, J.T.; et al. Fc-Mediated Anomalous Biodistribution of Therapeutic Antibodies in Immunodeficient Mouse Models. Cancer Res. 2018, 78, 1820–1832. [Google Scholar] [CrossRef] [Green Version]
- Michel, R.B.; Ochakovskaya, R.; Mattes, M.J. Rapid blood clearance of injected mouse IgG2a in SCID mice. Cancer Immunol. Immunother. 2002, 51, 547–556. [Google Scholar] [CrossRef]
- Aithal, A.; Rauth, S.; Kshirsagar, P.; Shah, A.; Lakshmanan, I.; Junker, W.M.; Jain, M.; Ponnusamy, M.P.; Batra, S.K. MUC16 as a novel target for cancer therapy. Expert Opin. Ther. Targets 2018, 22, 675–686. [Google Scholar] [CrossRef]
- Xiang, X.; Feng, M.; Felder, M.; Connor, J.P.; Man, Y.G.; Patankar, M.S.; Ho, M. HN125: A Novel Immunoadhesin Targeting MUC16 with Potential for Cancer Therapy. J. Cancer 2011, 2, 280–291. [Google Scholar] [CrossRef]
- Larson, S.M.; Carrasquillo, J.A.; Cheung, N.K.; Press, O.W. Radioimmunotherapy of human tumours. Nat. Rev. Cancer 2015, 15, 347–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| AUC (%IA.h/mL) | t1/2α (h) | t1/2ß (h) | CL (mL) | Vss (mL/h × 10−2) | |
|---|---|---|---|---|---|
| Tumor-bearing mice | 979.0 ± 197.6 | 5.8 ± 2.6 | 92.4 ± 15.2 | 0.11 ± 0.01 | 9.5 ± 2.4 |
| Normal mice | 850.0 ± 245 | 2.64 ± 1.3 | 63.58 ± 19.5 | 0.14 ± 0.04 | 11.1 ± 6.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Babeker, H.; Ketchemen, J.P.; Annan Sudarsan, A.; Andrahennadi, S.; Tikum, A.F.; Nambisan, A.K.; Fonge, H.; Uppalapati, M. Engineering of a Fully Human Anti-MUC-16 Antibody and Evaluation as a PET Imaging Agent. Pharmaceutics 2022, 14, 2824. https://doi.org/10.3390/pharmaceutics14122824
Babeker H, Ketchemen JP, Annan Sudarsan A, Andrahennadi S, Tikum AF, Nambisan AK, Fonge H, Uppalapati M. Engineering of a Fully Human Anti-MUC-16 Antibody and Evaluation as a PET Imaging Agent. Pharmaceutics. 2022; 14(12):2824. https://doi.org/10.3390/pharmaceutics14122824
Chicago/Turabian StyleBabeker, Hanan, Jessica Pougoue Ketchemen, Arunkumar Annan Sudarsan, Samitha Andrahennadi, Anjong Florence Tikum, Anand Krishnan Nambisan, Humphrey Fonge, and Maruti Uppalapati. 2022. "Engineering of a Fully Human Anti-MUC-16 Antibody and Evaluation as a PET Imaging Agent" Pharmaceutics 14, no. 12: 2824. https://doi.org/10.3390/pharmaceutics14122824