
Blockade of Cholecystokinin Type 2 Receptors Prevents the Onset of Vincristine-Induced Neuropathy in Mice
 , ,
, ,  , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Transcriptomic Study
2.2.1. Tissue Collection
2.2.2. mRNA Extraction
2.2.3. Transcriptomic Analysis
2.2.4. RT-qPCR
2.2.5. Localization of CCK2R in the Sensory Nervous System
2.3. Pharmacological Study
2.3.1. Chemicals
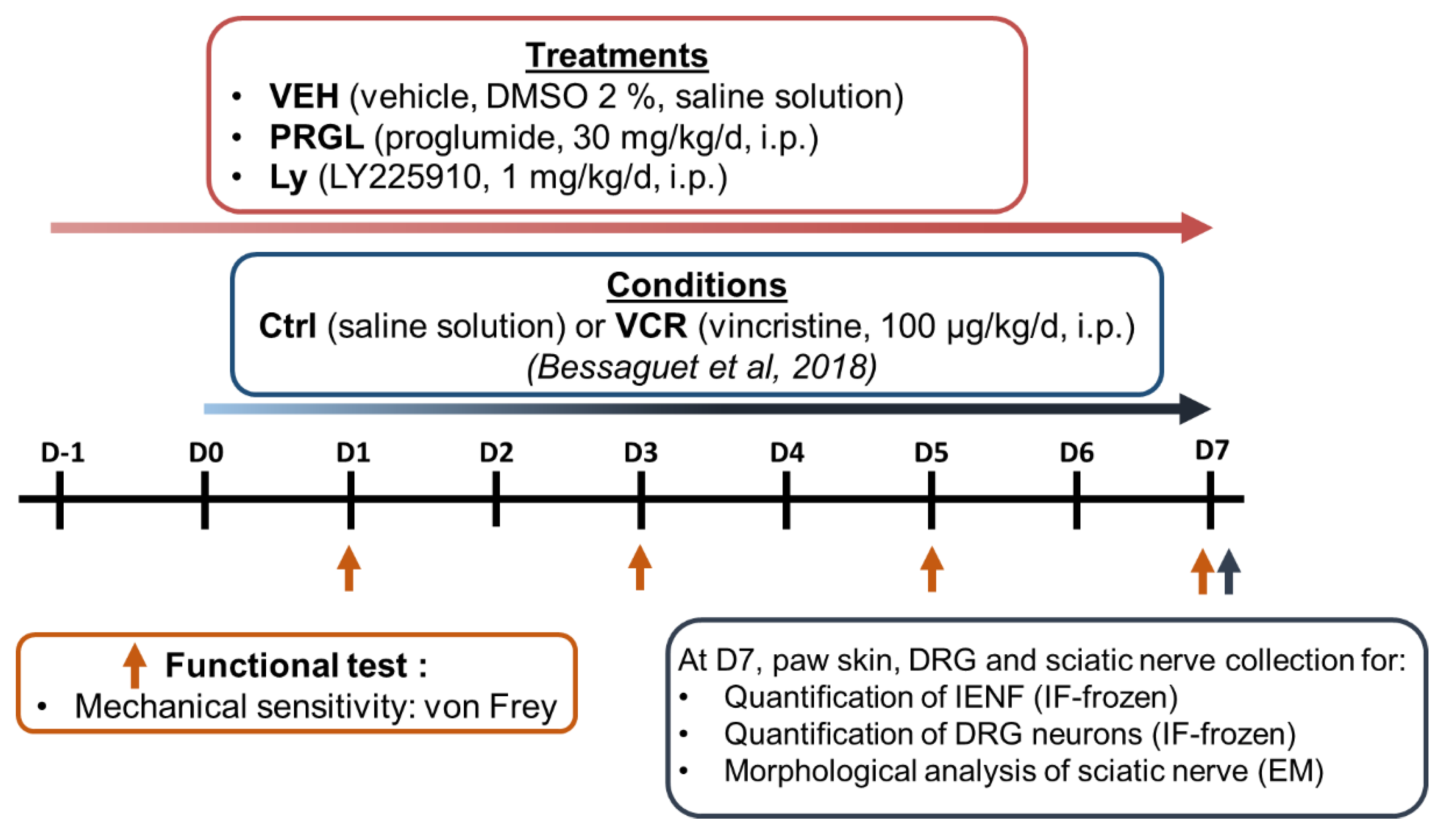
2.3.2. Schedule Treatments
- Ctrl-VEH
- Ctrl-PRGL
- Ctrl-Ly
- VCR-VEH
- VCR-PRGL
- VCR-Ly
2.3.3. Von Frey Filament Test
2.3.4. Quantification of Intraepidermal Nerve Fiber (IENF) and DRG Neuron Densities
2.3.5. Sciatic Nerve Ultrastructural Analysis
2.4. Evaluation of Cancer Cell Viability
2.5. Data Analysis
3. Results
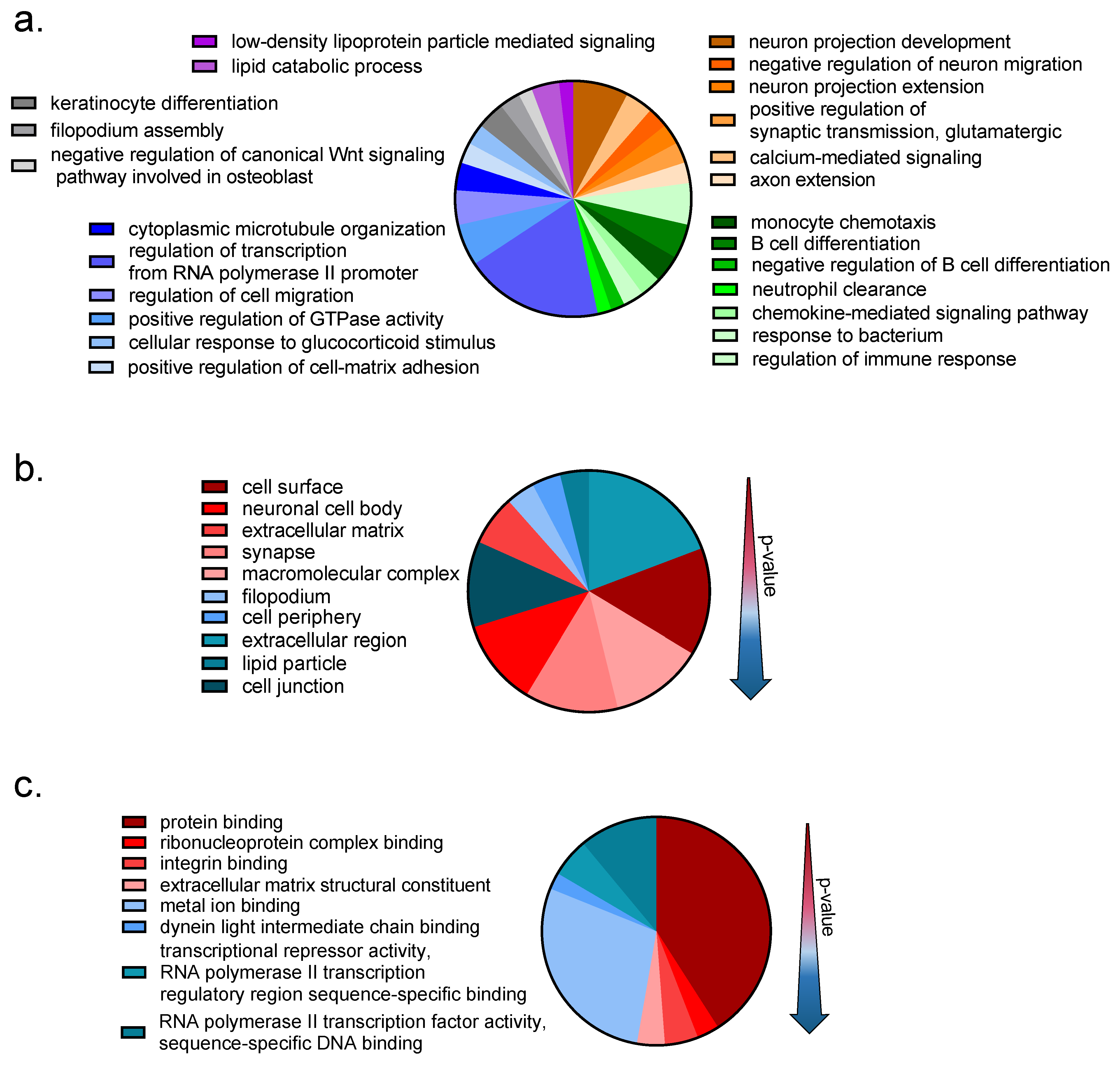
3.1. Transcriptomic Study
3.1.1. Transcriptomic Changes Induced by VCR Exposure in Mouse DRG
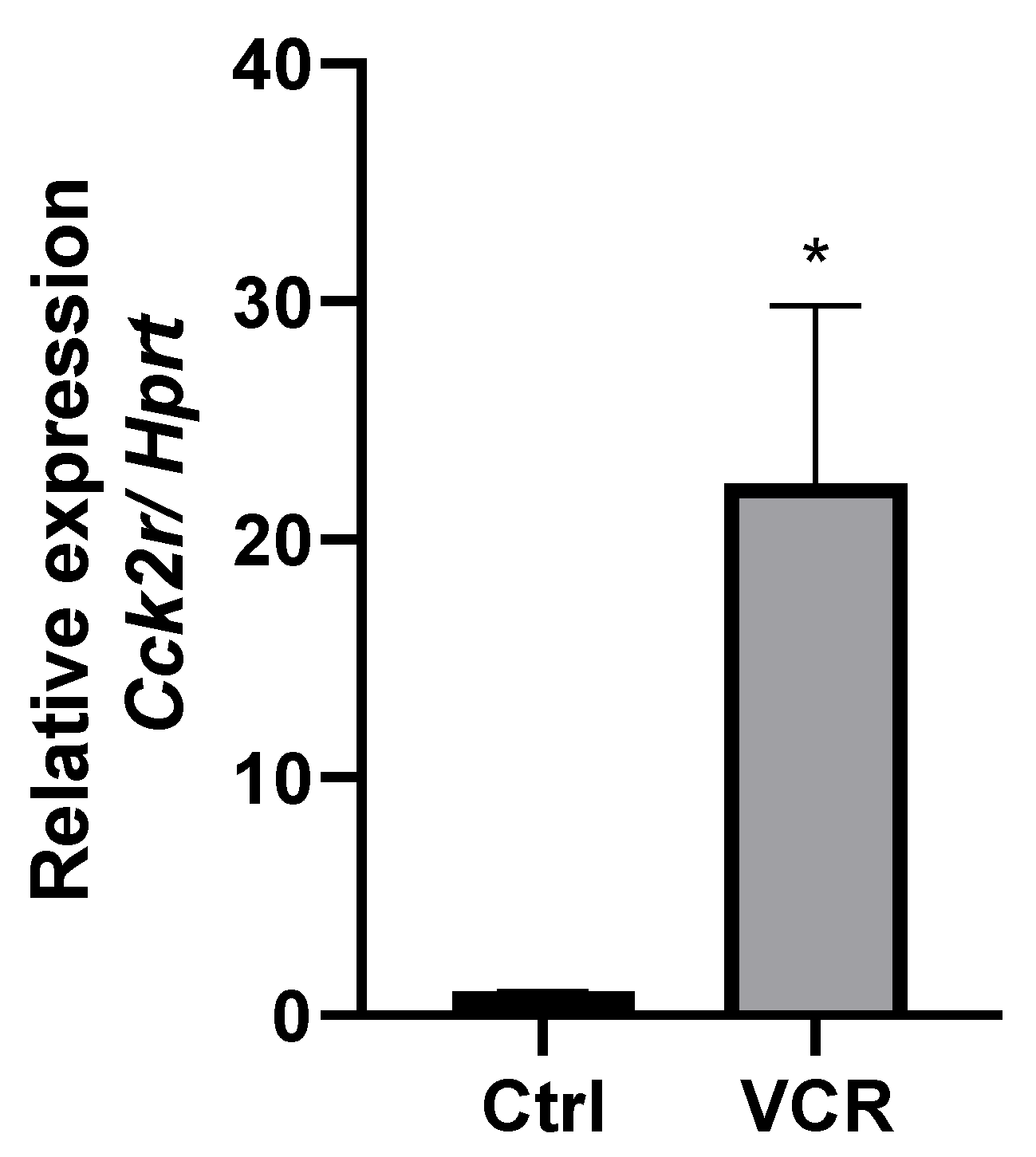
3.1.2. CCK2R mRNA Is Overexpressed in DRG of VCR-Administrated Mice
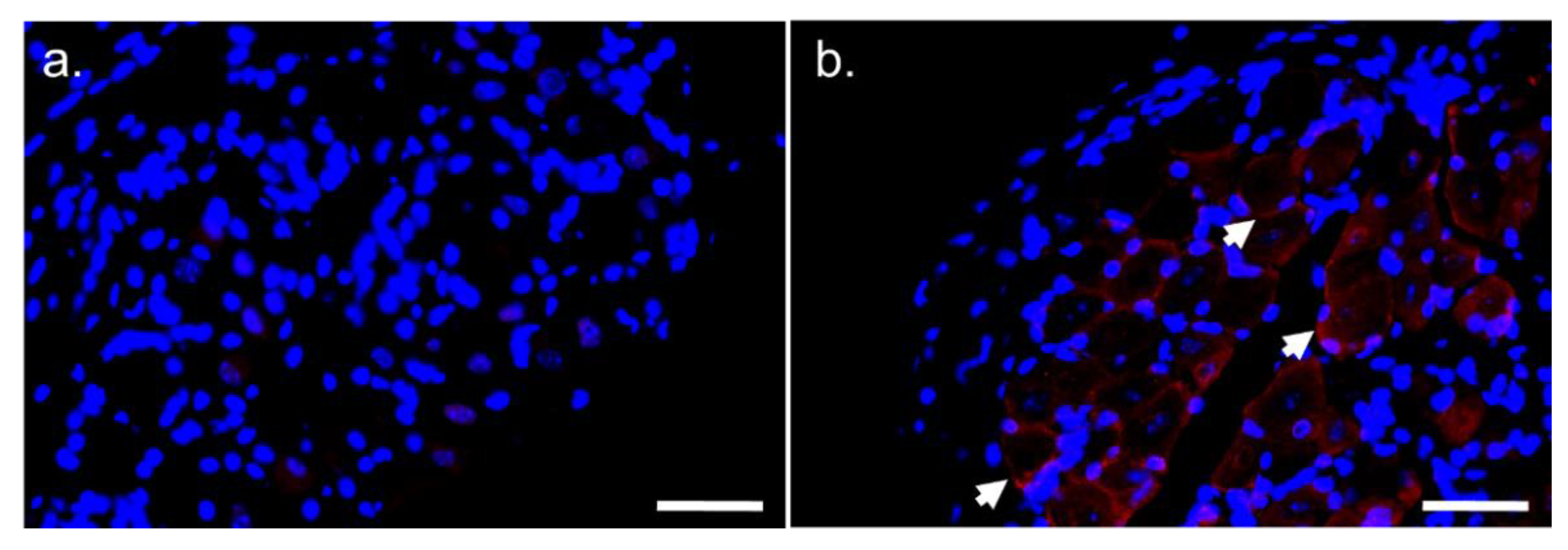
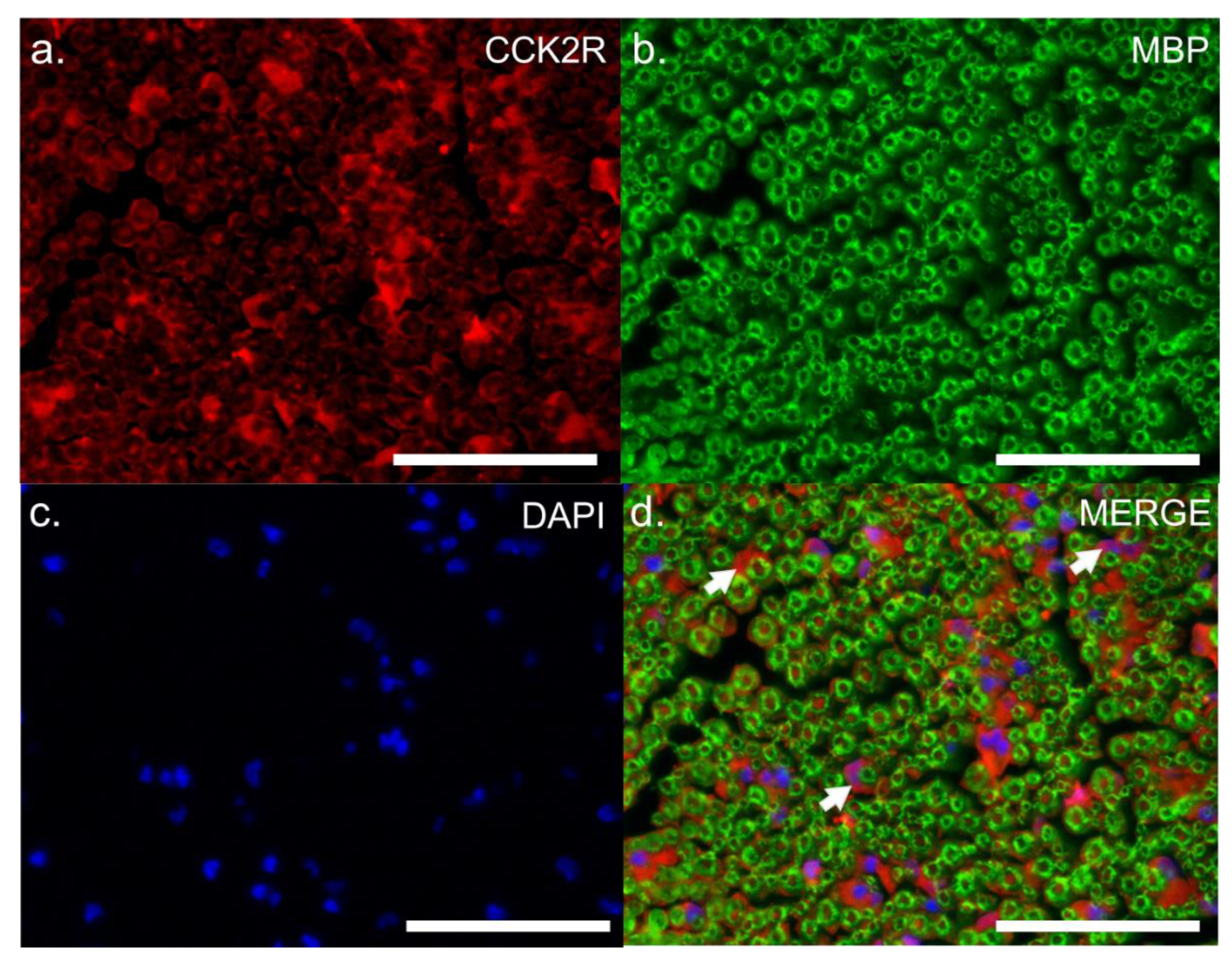
3.1.3. Location of CCK2R Protein in Mouse DRG and Sciatic Nerve
3.2. Pharmacological Study
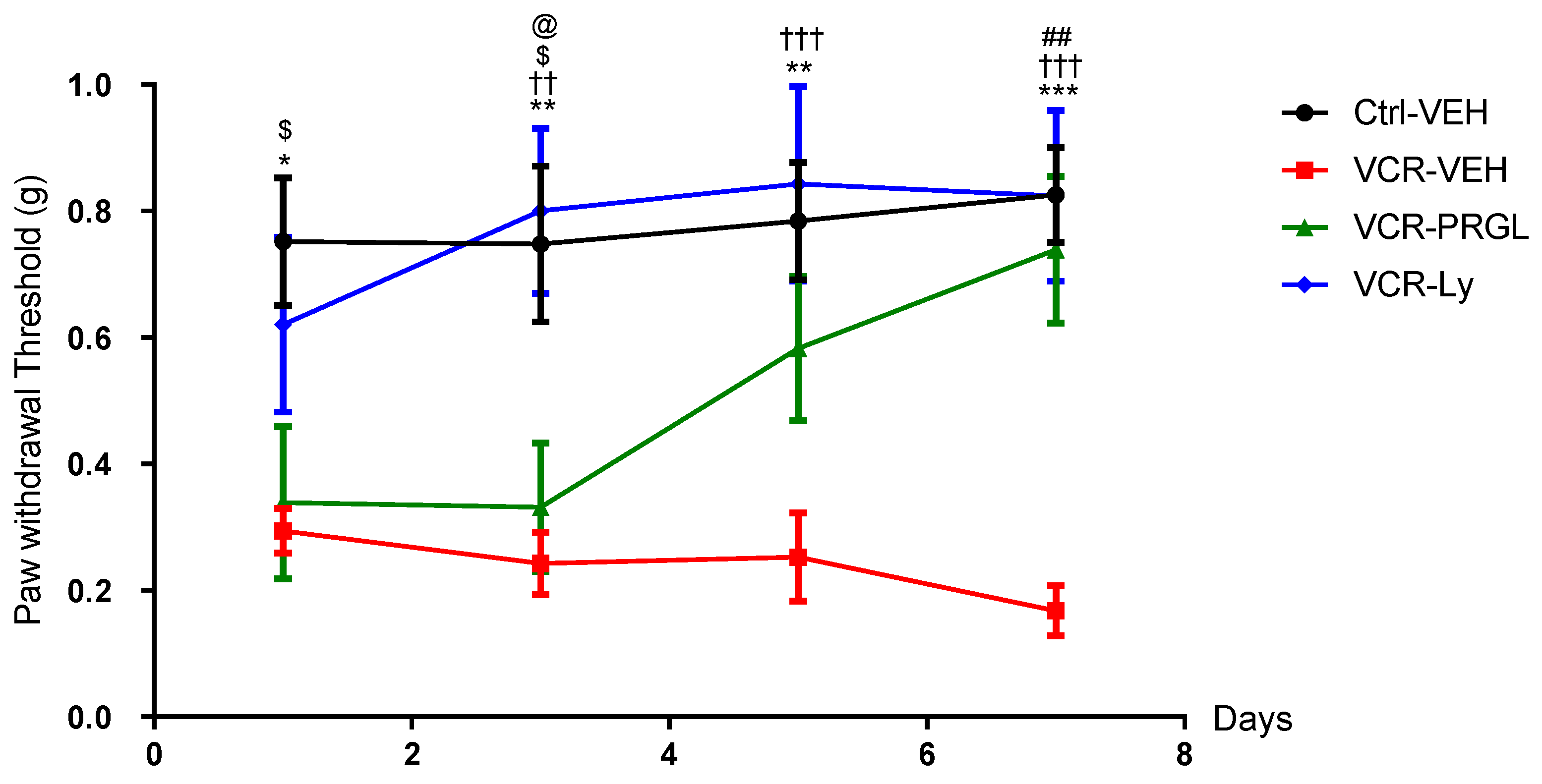
3.2.1. CCK2R Signaling Contributes to VCR-Induced Mechanical Allodynia
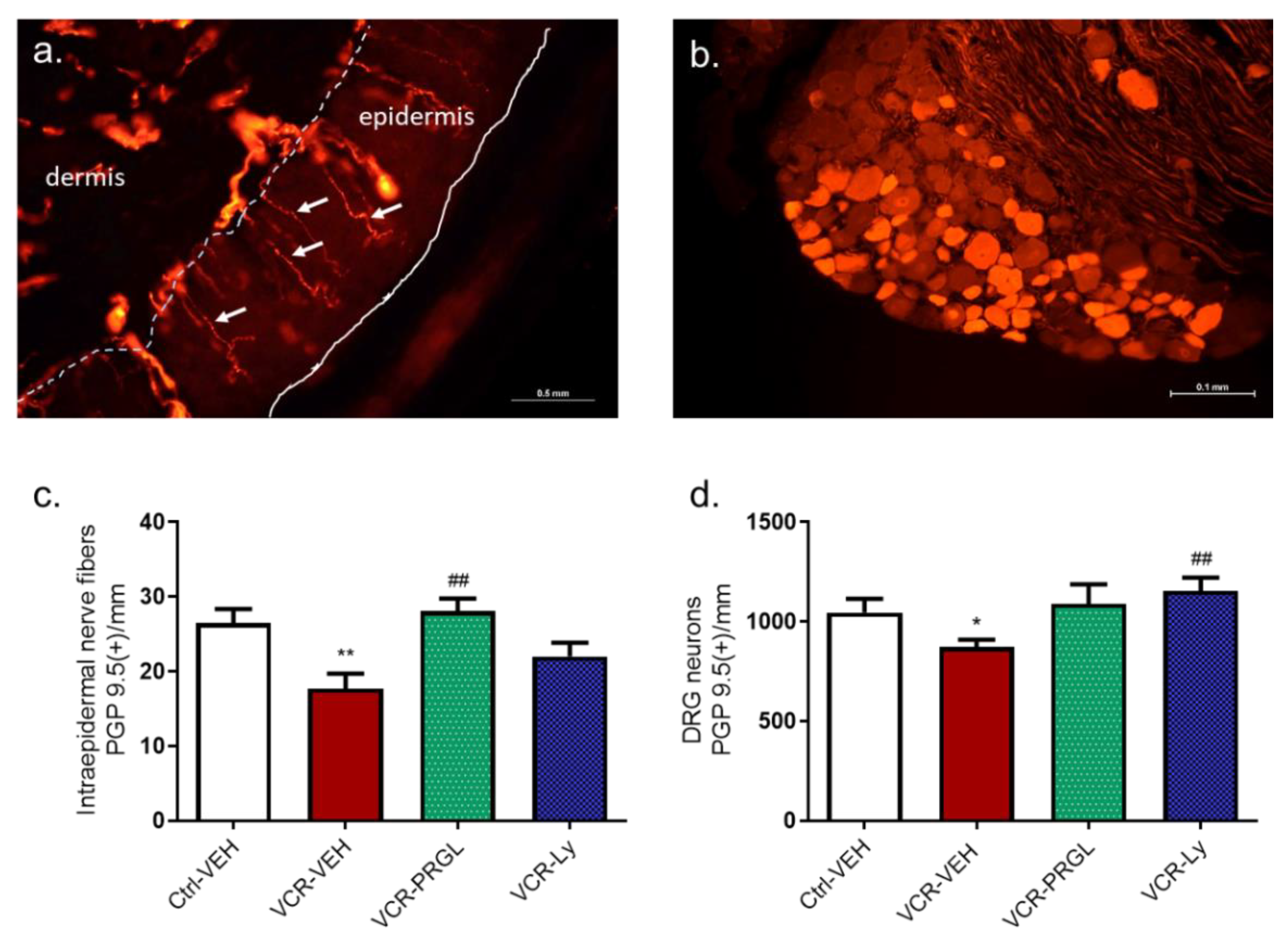
3.2.2. CCK2R Blockade Alleviates the Decrease in IENF and DRG Neuron Densities Induced by VCR
3.2.3. Effect of CCK2R Blockade on Alterations Induced by VCR on Myelinated Nerve Fiber Density and Morphology in Sciatic Nerves
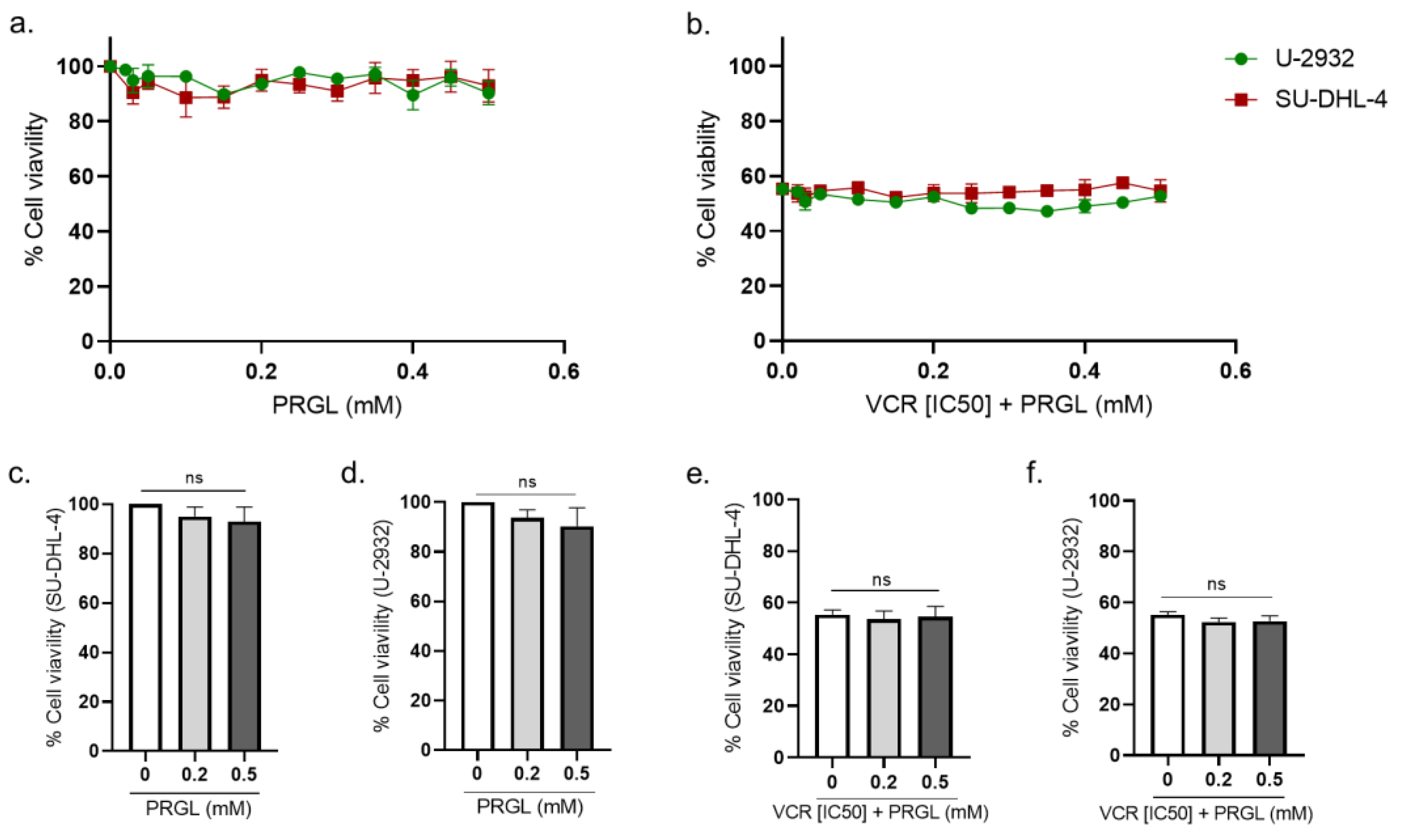
3.3. Pharmacological Modulation of CCK2R Did Not Alter the Anticancer Activity of VCR on Human Lymphoma Cell Lines
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Argyriou, A.A.; Bruna, J.; Marmiroli, P.; Cavaletti, G. Chemotherapy-Induced Peripheral Neurotoxicity (CIPN): An Update. Crit. Rev. Oncol. Hematol. 2012, 82, 51–77. [Google Scholar] [CrossRef]
- Jackson, D.V.; Sethi, V.S.; Spurr, C.L.; McWhorter, J.M. Pharmacokinetics of Vincristine in the Cerebrospinal Fluid of Humans. Cancer Res. 1981, 41, 1466–1468. [Google Scholar]
- Guo, L.; Hamre, J.; Eldridge, S.; Behrsing, H.P.; Cutuli, F.M.; Mussio, J.; Davis, M. Editor’s Highlight: Multiparametric Image Analysis of Rat Dorsal Root Ganglion Cultures to Evaluate Peripheral Neuropathy-Inducing Chemotherapeutics. Toxicol. Sci. 2017, 156, 275–288. [Google Scholar] [CrossRef] [Green Version]
- Silva, A.; Wang, Q.; Wang, M.; Ravula, S.K.; Glass, J.D. Evidence for Direct Axonal Toxicity in Vincristine Neuropathy. J. Peripher. Nerv. Syst. 2006, 11, 211–216. [Google Scholar] [CrossRef]
- Authier, N.; Balayssac, D.; Marchand, F.; Ling, B.; Zangarelli, A.; Descoeur, J.; Coudore, F.; Bourinet, E.; Eschalier, A. Animal Models of Chemotherapy-Evoked Painful Peripheral Neuropathies. Neurother. J. Am. Soc. Exp. Neurother. 2009, 6, 620–629. [Google Scholar] [CrossRef] [Green Version]
- Windebank, A.J.; Grisold, W. Chemotherapy-Induced Neuropathy. J. Peripher. Nerv. Syst. 2008, 13, 27–46. [Google Scholar] [CrossRef]
- Staff, N.P.; Grisold, A.; Grisold, W.; Windebank, A.J. Chemotherapy-Induced Peripheral Neuropathy: A Current Review: CIPN. Ann. Neurol. 2017, 81, 772–781. [Google Scholar] [CrossRef]
- Colvin, L.A. Chemotherapy-Induced Peripheral Neuropathy: Where Are We Now? Pain 2019, 160 (Suppl. S1), S1–S10. [Google Scholar] [CrossRef]
- Bernard, A.; Danigo, A.; Bourthoumieu, S.; Mroué, M.; Desmoulière, A.; Sturtz, F.; Rovini, A.; Demiot, C. The Cholecystokinin Type 2 Receptor, a Pharmacological Target for Pain Management. Pharmaceuticals 2021, 14, 1185. [Google Scholar] [CrossRef]
- LaVigne, J.E.; Alles, S.R.A. CCK2 Receptors in Chronic Pain. Neurobiol. Pain 2022, 11, 100092. [Google Scholar] [CrossRef]
- Greenough, A.; Cole, G.; Lewis, J.; Lockton, A.; Blundell, J. Untangling the Effects of Hunger, Anxiety, and Nausea on Energy Intake during Intravenous Cholecystokinin Octapeptide (CCK-8) Infusion. Physiol. Behav. 1998, 65, 303–310. [Google Scholar] [CrossRef]
- Yang, Y.; Li, Q.; He, Q.-H.; Han, J.-S.; Su, L.; Wan, Y. Heteromerization of μ-Opioid Receptor and Cholecystokinin B Receptor through the Third Transmembrane Domain of the μ-Opioid Receptor Contributes to the Anti-Opioid Effects of Cholecystokinin Octapeptide. Exp. Mol. Med. 2018, 50, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Jackson, D.L.; Graff, C.B.; Richardson, J.D.; Hargreaves, K.M. Glutamate Participates in the Peripheral Modulation of Thermal Hyperalgesia in Rats. Eur. J. Pharmacol. 1995, 284, 321–325. [Google Scholar] [CrossRef]
- Léna, I.; Dhôtel, H.; Garbay, C.; Daugé, V. Involvement of D 2 Dopamine Receptors in the Opposing Effects of Two CCK-B Agonists in a Spatial Recognition Memory Task: Role of the Anterior Nucleus Accumbens. Psychopharmacology 2001, 153, 170–179. [Google Scholar] [CrossRef]
- Yin, K.; Deuis, J.R.; Lewis, R.J.; Vetter, I. Transcriptomic and Behavioural Characterisation of a Mouse Model of Burn Pain Identify the Cholecystokinin 2 Receptor as an Analgesic Target. Mol. Pain 2016, 12, 1744806916665366. [Google Scholar] [CrossRef] [Green Version]
- Yamamoto, T.; Nozaki-Taguchi, N. Role of Cholecystokinin-B Receptor in the Maintenance of Thermal Hyperalgesia Induced by Unilateral Constriction Injury to the Sciatic Nerve in the Rat. Neurosci. Lett. 1995, 202, 89–92. [Google Scholar] [CrossRef]
- Zhang, X.; Dagerlind, A.; Elde, R.P.; Castel, M.N.; Broberger, C.; Wiesenfeld-Hallin, Z.; Hökfelt, T. Marked Increase in Cholecystokinin B Receptor Messenger RNA Levels in Rat Dorsal Root Ganglia after Peripheral Axotomy. Neuroscience 1993, 57, 227–233. [Google Scholar] [CrossRef]
- Percie du Sert, N.; Ahluwalia, A.; Alam, S.; Avey, M.T.; Baker, M.; Browne, W.J.; Clark, A.; Cuthill, I.C.; Dirnagl, U.; Emerson, M.; et al. Reporting Animal Research: Explanation and Elaboration for the ARRIVE Guidelines 2.0. PLoS Biol. 2020, 18, e3000411. [Google Scholar] [CrossRef]
- Bessaguet, F.; Danigo, A.; Bouchenaki, H.; Duchesne, M.; Magy, L.; Richard, L.; Sturtz, F.; Desmoulière, A.; Demiot, C. Neuroprotective Effect of Angiotensin II Type 2 Receptor Stimulation in Vincristine-Induced Mechanical Allodynia. Pain 2018, 159, 2538–2546. [Google Scholar] [CrossRef]
- Ballaz, S.J.; Akil, H.; Watson, S.J. The CCK-System Mediates Adaptation to Novelty-Induced Stress in the Rat: A Pharmacological Evidence. Neurosci. Lett. 2007, 428, 27–32. [Google Scholar] [CrossRef]
- Bouchenaki, H.; Danigo, A.; Bernard, A.; Bessaguet, F.; Richard, L.; Sturtz, F.; Balayssac, D.; Magy, L.; Demiot, C. Ramipril Alleviates Oxaliplatin-Induced Acute Pain Syndrome in Mice. Front. Pharmacol. 2021, 12, 712442. [Google Scholar] [CrossRef]
- Bonin, R.P.; Bories, C.; De Koninck, Y. A Simplified Up-Down Method (SUDO) for Measuring Mechanical Nociception in Rodents Using von Frey Filaments. Mol. Pain 2014, 10, 26. [Google Scholar] [CrossRef]
- Lauria, G.; Cornblath, D.R.; Johansson, O.; McArthur, J.C.; Mellgren, S.I.; Nolano, M.; Rosenberg, N.; Sommer, C. EFNS Guidelines on the Use of Skin Biopsy in the Diagnosis of Peripheral Neuropathy. Eur. J. Neurol. Off. J. Eur. Fed. Neurol. Soc. 2005, 12, 747–758. [Google Scholar] [CrossRef]
- Bouchenaki, H.; Bernard, A.; Bessaguet, F.; Frachet, S.; Richard, L.; Sturtz, F.; Magy, L.; Bourthoumieu, S.; Demiot, C.; Danigo, A. Neuroprotective Effect of Ramipril Is Mediated by AT2 in a Mouse MODEL of Paclitaxel-Induced Peripheral Neuropathy. Pharmaceutics 2022, 14, 848. [Google Scholar] [CrossRef]
- Nga, N.T.H.; Ngoc, T.T.B.; Trinh, N.T.M.; Thuoc, T.L.; Thao, D.T.P. Optimization and Application of MTT Assay in Determining Density of Suspension Cells. Anal. Biochem. 2020, 610, 113937. [Google Scholar] [CrossRef]
- Abuirmeileh, A.; Lever, R.; Kingsbury, A.E.; Lees, A.J.; Locke, I.C.; Knight, R.A.; Chowdrey, H.S.; Biggs, C.S.; Whitton, P.S. The Corticotrophin-Releasing Factor-like Peptide Urocortin Reverses Key Deficits in Two Rodent Models of Parkinson’s Disease. Eur. J. Neurosci. 2007, 26, 417–423. [Google Scholar] [CrossRef]
- Bonilla, I.E.; Tanabe, K.; Strittmatter, S.M. Small Proline-Rich Repeat Protein 1A Is Expressed by Axotomized Neurons and Promotes Axonal Outgrowth. J. Neurosci. 2002, 22, 1303–1315. [Google Scholar] [CrossRef]
- Hafezparast, M.; Klocke, R.; Ruhrberg, C.; Marquardt, A.; Ahmad-Annuar, A.; Bowen, S.; Lalli, G.; Witherden, A.S.; Hummerich, H.; Nicholson, S.; et al. Mutations in Dynein Link Motor Neuron Degeneration to Defects in Retrograde Transport. Science 2003, 300, 808–812. [Google Scholar] [CrossRef]
- Seijffers, R.; Mills, C.D.; Woolf, C.J. Cellular/Molecular ATF3 Increases the Intrinsic Growth State of DRG Neurons to Enhance Peripheral Nerve Regeneration. J. Neurosci. 2007, 27, 7911–7920. [Google Scholar] [CrossRef] [Green Version]
- Kiryu-Seo, S.; Nagata, K.; Saido, T.C.; Kiyama, H. New Insights of a Neuronal Peptidase DINE/ECEL1: Nerve Development, Nerve Regeneration and Neurogenic Pathogenesis. Neurochem. Res. 2019, 44, 1279–1288. [Google Scholar] [CrossRef]
- Meyer zu Hörste, G.; Heidenreich, H.; Mausberg, A.K.; Lehmann, H.C.; ten Asbroek, A.L.M.A.; Saavedra, J.T.; Baas, F.; Hartung, H.-P.; Wiendl, H.; Kieseier, B.C. Mouse Schwann Cells Activate MHC Class I and II Restricted T-Cell Responses, but Require External Peptide Processing for MHC Class II Presentation. Neurobiol. Dis. 2010, 37, 483–490. [Google Scholar] [CrossRef]
- Mi, S.; Miller, R.H.; Lee, X.; Scott, M.L.; Shulag-Morskaya, S.; Shao, Z.; Chang, J.; Thill, G.; Levesque, M.; Zhang, M.; et al. LINGO-1 Negatively Regulates Myelination by Oligodendrocytes. Nat. Neurosci. 2005, 8, 745–751. [Google Scholar] [CrossRef]
- Duda, S.; Witte, T.; Stangel, M.; Adams, J.; Schmidt, R.E.; Baerlecken, N.T. Autoantibodies Binding to Stathmin-4: New Marker for Polyneuropathy in Primary Sjögren’s Syndrome. Immunol. Res. 2017, 65, 1099–1102. [Google Scholar] [CrossRef]
- Wang, J.; Xu, W.; Kong, Y.; Huang, J.; Ding, Z.; Deng, M.; Guo, Q.; Zou, W. SNAP-25 Contributes to Neuropathic Pain by Regulation of VGLuT2 Expression in Rats. Neuroscience 2019, 423, 86–97. [Google Scholar] [CrossRef]
- Ferraro, G.B.; Morrison, C.J.; Overall, C.M.; Strittmatter, S.M.; Fournier, A.E. Membrane-Type Matrix Metalloproteinase-3 Regulates Neuronal Responsiveness to Myelin through Nogo-66 Receptor 1 Cleavage. J. Biol. Chem. 2011, 286, 31418–31424. [Google Scholar] [CrossRef] [Green Version]
- Cao, S.; Yuan, J.; Zhang, D.; Wen, S.; Wang, J.; Li, Y.; Deng, W. Transcriptome Changes in Dorsal Spinal Cord of Rats with Neuropathic Pain. J. Pain Res. 2019, 12, 3013–3023. [Google Scholar] [CrossRef] [Green Version]
- Sun, C.; Wu, G.; Zhang, Z.; Cao, R.; Cui, S. Protein Tyrosine Phosphatase Receptor Type D Regulates Neuropathic Pain After Nerve Injury via the STING-IFN-I Pathway. Front. Mol. Neurosci. 2022, 15, 859166. [Google Scholar] [CrossRef]
- Wu, Y.; Fu, Q.; Huang, X.; Luo, Y.; Wan, S.; Peng, M.; Su, S.; Xu, X.; Li, Y.; Li, X.; et al. NWD1 Facilitates Synaptic Transmission and Contributes to Neuropathic Pain. Neuropharmacology 2022, 205, 108919. [Google Scholar] [CrossRef]
- Zhang, D.-Y.; Yu, K.; Yang, Z.; Liu, X.-Z.; Ma, X.-F.; Li, Y.-X. Variation in Expression of Small Ubiquitin-like Modifiers in Injured Sciatic Nerve of Mice. Neural Regen. Res. 2019, 14, 1455–1461. [Google Scholar] [CrossRef]
- Cao, Y.; Lipka, J.; Stucchi, R.; Burute, M.; Pan, X.; Portegies, S.; Tas, R.; Willems, J.; Will, L.; MacGillavry, H.; et al. Microtubule Minus-End Binding Protein CAMSAP2 and Kinesin-14 Motor KIFC3 Control Dendritic Microtubule Organization. Curr. Biol. 2020, 30, 899–908.e6. [Google Scholar] [CrossRef] [Green Version]
- Rivat, C.; Sar, C.; Mechaly, I.; Leyris, J.-P.; Diouloufet, L.; Sonrier, C.; Philipson, Y.; Lucas, O.; Mallié, S.; Jouvenel, A.; et al. Inhibition of Neuronal FLT3 Receptor Tyrosine Kinase Alleviates Peripheral Neuropathic Pain in Mice. Nat. Commun. 2018, 9, 1042. [Google Scholar] [CrossRef] [PubMed]
- Leung, T.; Rajendran, R.; Singh, S.; Garva, R.; Krstic-Demonacos, M.; Demonacos, C. Cytochrome P450 2E1 (CYP2E1) Regulates the Response to Oxidative Stress and Migration of Breast Cancer Cells. Breast Cancer Res. 2013, 15, R107. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brosius Lutz, A.; Lucas, T.A.; Carson, G.A.; Caneda, C.; Zhou, L.; Barres, B.A.; Buckwalter, M.S.; Sloan, S.A. An RNA-Sequencing Transcriptome of the Rodent Schwann Cell Response to Peripheral Nerve Injury. J. Neuroinflamm. 2022, 19, 105. [Google Scholar] [CrossRef] [PubMed]
- Hilario, J.D.; Rodino-Klapac, L.R.; Wang, C.; Beattie, C.E. Semaphorin 5A Is a Bifunctional Axon Guidance Cue for Axial Motoneurons in Vivo. Dev. Biol. 2009, 326, 190–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ugarte, S.D.; Homanics, G.E.; Firestone, L.L.; Hammond, D.L. Sensory Thresholds and the Antinociceptive Effects of GABA Receptor Agonists in Mice Lacking the Β3 Subunit of the GABAA Receptor. Neuroscience 1999, 95, 795–806. [Google Scholar] [CrossRef] [PubMed]
- Mietto, B.S.; Jhelum, P.; Schulz, K.; David, S. Schwann Cells Provide Iron to Axonal Mitochondria and Its Role in Nerve Regeneration. J. Neurosci. Off. J. Soc. Neurosci. 2021, 41, 7300–7313. [Google Scholar] [CrossRef]
- Kuo, T.-Y.; Hong, C.-J.; Hsueh, Y.-P. Bcl11A/CTIP1 Regulates Expression of DCC and MAP1b in Control of Axon Branching and Dendrite Outgrowth. Mol. Cell. Neurosci. 2009, 42, 195–207. [Google Scholar] [CrossRef]
- Kreko-Pierce, T.; Pugh, J.R. Altered Synaptic Transmission and Excitability of Cerebellar Nuclear Neurons in a Mouse Model of Duchenne Muscular Dystrophy. Front. Cell. Neurosci. 2022, 16, 926518. [Google Scholar] [CrossRef]
- Meng, F.-W.; Jing, X.-N.; Song, G.-H.; Jie, L.-L.; Shen, F.-F. Prox1 Induces New Lymphatic Vessel Formation and Promotes Nerve Reconstruction in a Mouse Model of Sciatic Nerve Crush Injury. J. Anat. 2020, 237, 933–940. [Google Scholar] [CrossRef]
- Benedetti, F.; Amanzio, M.; Thoen, W. Disruption of Opioid-Induced Placebo Responses by Activation of Cholecystokinin Type-2 Receptors. Psychopharmacology 2011, 213, 791–797. [Google Scholar] [CrossRef]
- Bernstein, Z.P.; Yucht, S.; Battista, E.; Lema, M.; Spaulding, M.B. Proglumide as a Morphine Adjunct in Cancer Pain Management. J. Pain Symptom Manag. 1998, 15, 314–320. [Google Scholar] [CrossRef] [PubMed]
- Lavigne, G.J.; Hargreaves, K.M.; Schmidt, E.A.; Dionne, R.A. Proglumide Potentiates Morphine Analgesia for Acute Postsurgical Pain. Clin. Pharmacol. Ther. 1989, 45, 666–673. [Google Scholar] [CrossRef] [PubMed]
- Hill, D.; Horwell, D.C.; Hunter, J.C.; Kneen, C.O.; Pritchard, M.C.; Suman-Chauhan, N. Synthesis of a Potent and Selective Non-Peptide CCK-B/Gastrin Receptor Antagonist Tritiated Ligand. Bioorg. Med. Chem. Lett. 1993, 3, 885–888. [Google Scholar] [CrossRef]
- Starobova, H.; Mueller, A.; Deuis, J.R.; Carter, D.A.; Vetter, I. Inflammatory and Neuropathic Gene Expression Signatures of Chemotherapy-Induced Neuropathy Induced by Vincristine, Cisplatin, and Oxaliplatin in C57BL/6J Mice. J. Pain 2019, 21, 182–194. [Google Scholar] [CrossRef] [PubMed]
- Sun, W.; Yang, S.; Wu, S.; Ba, X.; Xiong, D.; Xiao, L.; Hao, Y. Transcriptome Analysis Reveals Dysregulation of Inflammatory and Neuronal Function in Dorsal Root Ganglion of Paclitaxel-Induced Peripheral Neuropathy Rats. Mol. Pain 2022, 17448069221106167. [Google Scholar] [CrossRef] [PubMed]
- Stone, L.S.; Molliver, D.C. In Search of Analgesia: Emerging Roles of GPCRs in Pain. Mol. Interv. 2009, 9, 234–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, J.; Kim, J.H.; Kim, Y.; Cho, H.; Hong, S.K.; Yoon, Y.W. Role of Spinal Cholecystokinin in Neuropathic Pain after Spinal Cord Hemisection in Rats. Neurosci. Lett. 2009, 462, 303–307. [Google Scholar] [CrossRef] [PubMed]
- Roca-Lapirot, O.; Fossat, P.; Ma, S.; Egron, K.; Trigilio, G.; López-González, M.-J.; Covita, J.; Bouali-Benazzouz, R.; Favereaux, A.; Gundlach, A.L.; et al. Acquisition of Analgesic Properties by the Cholecystokinin (CCK)/CCK2 Receptor System within the Amygdala in a Persistent Inflammatory Pain Condition. Pain 2019, 160, 345–357. [Google Scholar] [CrossRef]
- Kamei, J.; Zushida, K. The Role of Spinal Cholecystokinin B Receptors in Thermal Allodynia and Hyperalgesia in Diabetic Mice. Brain Res. 2001, 892, 370–375. [Google Scholar] [CrossRef]
- Juárez-Rojop, I.E.; Granados-Soto, V.; Díaz-Zagoya, J.C.; Flores-Murrieta, F.J.; Torres-López, J.E. Involvement of Cholecystokinin in Peripheral Nociceptive Sensitization during Diabetes in Rats as Revealed by the Formalin Response. Pain 2006, 122, 118–125. [Google Scholar] [CrossRef]
- Friedrich, A.E.; Gebhart, G.F. Effects of Spinal Cholecystokinin Receptor Antagonists on Morphine Antinociception in a Model of Visceral Pain in the Rat. J. Pharmacol. Exp. Ther. 2000, 292, 538–544. [Google Scholar] [PubMed]
- Kellstein, D.E.; Mayer, D.J. Chronic Administration of Cholecystokinin Antagonists Reverses the Enhancement of Spinal Morphine Analgesia Induced by Acute Pretreatment. Brain Res. 1990, 516, 263–270. [Google Scholar] [CrossRef] [PubMed]
- Lehmann, K.A.; Schlüsener, M.; Arabatsis, P. Failure of Proglumide, a Cholecystokinin Antagonist, to Potentiate Clinical Morphine Analgesia. A Randomized Double-Blind Postoperative Study Using Patient-Controlled Analgesia (PCA). Anesth. Analg. 1989, 68, 51–56. [Google Scholar] [PubMed]
- McCleane, G.J. The Cholecystokinin Antagonist Proglumide Enhances the Analgesic Efficacy of Morphine in Humans with Chronic Benign Pain. Anesth. Analg. 1998, 87, 1117–1120. [Google Scholar] [CrossRef] [PubMed]
- Tortorici, V.; Nogueira, L.; Salas, R.; Vanegas, H. Involvement of Local Cholecystokinin in the Tolerance Induced by Morphine Microinjections into the Periaqueductal Gray of Rats. Pain 2003, 102, 9–16. [Google Scholar] [CrossRef]
- Torres-López, J.E.; Juárez-Rojop, I.E.; Granados-Soto, V.; Diaz-Zagoya, J.C.; Flores-Murrieta, F.J.; Ortíz-López, J.U.S.; Cruz-Vera, J. Peripheral Participation of Cholecystokinin in the Morphine-Induced Peripheral Antinociceptive Effect in Non-Diabetic and Diabetic Rats. Neuropharmacology 2007, 52, 788–795. [Google Scholar] [CrossRef]
- Sjöstedt, E.; Zhong, W.; Fagerberg, L.; Karlsson, M.; Mitsios, N.; Adori, C.; Oksvold, P.; Edfors, F.; Limiszewska, A.; Hikmet, F.; et al. An Atlas of the Protein-Coding Genes in the Human, Pig, and Mouse Brain. Science 2020, 367, eaay5947. [Google Scholar] [CrossRef]
- Yu, S.; Zhang, Y.; Zhao, X.; Chang, Z.; Wei, Y.; Sun, Y.; Jiang, D.; Jiang, X.; Tao, J. Cholecystokinin Type B Receptor-Mediated Inhibition of a-Type K+ Channels Enhances Sensory Neuronal Excitability Through the Phosphatidylinositol 3-Kinase and C-Src-Dependent Jnk Pathway. Cell Commun. Signal. 2019, 17, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Vydyanathan, A.; Wu, Z.-Z.; Chen, S.-R.; Pan, H.-L. A-Type Voltage-Gated K+ Currents Influence Firing Properties of Isolectin B4-Positive but Not Isolectin B4-Negative Primary Sensory Neurons. J. Neurophysiol. 2005, 93, 3401–3409. [Google Scholar] [CrossRef] [Green Version]
- Cao, X.-H.; Byun, H.-S.; Chen, S.-R.; Cai, Y.-Q.; Pan, H.-L. Reduction in Voltage-Gated K+ Channel Activity in Primary Sensory Neurons in Painful Diabetic Neuropathy: Role of Brain-Derived Neurotrophic Factor. J. Neurochem. 2010, 114, 1460–1475. [Google Scholar] [CrossRef]
- Chi, X.X.; Nicol, G.D. Manipulation of the Potassium Channel Kv1.1 and Its Effect on Neuronal Excitability in Rat Sensory Neurons. J. Neurophysiol. 2007, 98, 2683–2692. [Google Scholar] [CrossRef] [PubMed]
- Chien, L.-Y.; Cheng, J.-K.; Chu, D.; Cheng, C.-F.; Tsaur, M.-L. Reduced Expression of A-Type Potassium Channels in Primary Sensory Neurons Induces Mechanical Hypersensitivity. J. Neurosci. 2007, 27, 9855–9865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takeda, M.; Tsuboi, Y.; Kitagawa, J.; Nakagawa, K.; Iwata, K.; Matsumoto, S. Potassium Channels as a Potential Therapeutic Target for Trigeminal Neuropathic and Inflammatory Pain. Mol. Pain 2011, 7, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zemel, B.M.; Ritter, D.M.; Covarrubias, M.; Muqeem, T. A-Type KV Channels in Dorsal Root Ganglion Neurons: Diversity, Function, and Dysfunction. Front. Mol. Neurosci. 2018, 11, 253. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ji, X.-T.; Qian, N.-S.; Zhang, T.; Li, J.-M.; Li, X.-K.; Wang, P.; Zhao, D.-S.; Huang, G.; Zhang, L.; Fei, Z.; et al. Spinal Astrocytic Activation Contributes to Mechanical Allodynia in a Rat Chemotherapy-Induced Neuropathic Pain Model. PLoS ONE 2013, 8, e60733. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kiguchi, N.; Maeda, T.; Kobayashi, Y.; Saika, F.; Kishioka, S. Involvement of Inflammatory Mediators in Neuropathic Pain Caused by Vincristine. Int. Rev. Neurobiol. 2009, 85, 179–190. [Google Scholar] [CrossRef]
- Nadella, S.; Ciofoaia, V.; Cao, H.; Kallakury, B.; Tucker, R.D.; Smith, J.P. Cholecystokinin Receptor Antagonist Therapy Decreases Inflammation and Fibrosis in Chronic Pancreatitis. Dig. Dis. Sci. 2020, 65, 1376–1384. [Google Scholar] [CrossRef]
- Dong, X.; Wang, Y.; Qin, Z. Molecular Mechanisms of Excitotoxicity and Their Relevance to Pathogenesis of Neurodegenerative Diseases. Acta Pharmacol. Sin. 2009, 30, 379–387. [Google Scholar] [CrossRef] [Green Version]
- Boyce, M.; Dowen, S.; Turnbull, G.; Van den Berg, F.; Zhao, C.-M.; Chen, D.; Black, J. Effect of Netazepide, a Gastrin/CCK2 Receptor Antagonist, on Gastric Acid Secretion and Rabeprazole-Induced Hypergastrinaemia in Healthy Subjects. Br. J. Clin. Pharmacol. 2015, 79, 744–755. [Google Scholar] [CrossRef] [Green Version]
- Lloyd, K.A.; Parsons, B.N.; Burkitt, M.D.; Moore, A.R.; Papoutsopoulou, S.; Boyce, M.; Duckworth, C.A.; Exarchou, K.; Howes, N.; Rainbow, L.; et al. Netazepide Inhibits Expression of Pappalysin 2 in Type 1 Gastric Neuroendocrine Tumors. Cell. Mol. Gastroenterol. Hepatol. 2020, 10, 113–132. [Google Scholar] [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes (Size) | Primers (5′-3′) | Accession No. |
|---|---|---|
| Mouse cck2r (122 bp) | Designed by Qiagen (QuantiTect Primer assay) | NM_007627 |
| Mouse hprt (185 bp) | F: TGATCAGTCAACGGGGGACAT R: AGGTCCTTTTCACCAGCAAGC | NM_013556.2 |
| Gene Symbol | Description | p-Value | Up FC | Ref. |
|---|---|---|---|---|
| ucn | Urocortin (Ucn), transcript variant 1 | 4.42 × 10−4 | 33.59 | Oxidative stress, neuroprotective in Parkinson’s disease [26] repair |
| sprr1A | Small proline-rich protein 1A | 7.75 × 10−4 | 23.35 | Promotes axonal outgrowth [27] |
| dync1h1 | Dynein cytoplasmic 1 heavy chain 1 | 0.0018 | 15.76 | The main motor protein responsible for retrograde axonal transport in neurons [28] |
| atf3 | Activating transcription factor 3 | 0.0020 | 6.06 | Survival and the regeneration of axons following axotomy [29] |
| ecel1 | Endothelin converting enzyme-like 1 | 0.0039 | 6.05 | Nerve development, nerve regeneration [30] |
| mill2 | MHC I like leukocyte 2, transcript variant 1 | 0.0029 | 4.45 | Expressed by Schwann cells, involved in antigen presentation [31] |
| lingo1 | Leucin rich repeated and ig domain containing 1 | 0.0033 | 2.08 | Negatively regulates myelination by oligodendrocytes [32] |
| stmn4 | Stathmin-like 4 | 0.0093 | 1.95 | Marker for polyneuropathy in primary Sjögren’s syndrome [33] |
| snap25 | Synaptosomal-associated protein 25 | 0.0086 | 1.70 | Cell exocytosis during synaptic transmission. Cleaved by Botulinum toxin, implicated in neuropathic pain [34] |
| MMP16 | Matrix metallopeptidase 16 | 0.0014 | 1.68 | Regulates neuronal responsiveness to myelin [35] |
| ptprd | Protein tyrosine phosphatase, receptor type, D | 0.0098 | 1.64 | Increased in DRG, neuropathic pain in CCI rat and mice [36,37] |
| nwd1 | NACHT and WD repeat domain containing 1 | 0.0043 | 1.62 | Facilitates synaptogenesis in spinal cord and then neuropathic pain [38] |
| sumo2 | Small ubiquitin-like modifier 2 | 0.0017 | 1.60 | Overexpressed after nerve injury, role in regeneration [39] |
| camsap2 | Calmodulin regulated spectrin-associated protein family, member 2 | 0.0065 | 1.58 | Control of dendritic microtubule organization [40] |
| flt3 | FMS-like tyrosine kinase 3 | 0.0032 | 1.51 | Alleviates pain if downregulated [41] |
| Gene Symbol | Description | p-Value | Down FC | Ref. |
|---|---|---|---|---|
| ms4a1 | Membrane-spanning 4-domains, subfamily A, member 1 | 0.0013 | 24.72 | B-lymphocyte antigen (CD20) |
| cyp2e1 | Cytochrome P450, family 2, subfamily e, polypeptide 1 | 0.0079 | 4.28 | Regulates response to oxidative stress [42] |
| sema5a | Semaphorin 5A | 0.0097 | 3.75 | Schwann cell injury response [43], axon guidance molecule in the nervous system [44] |
| gabrg3 | Gamma-aminobutyric acid (GABA) A receptor, subunit gamma 3 | 0.0066 | 2.55 | Synaptogenesis, nociception [45] |
| cp | Ceruloplasmin | 0.0098 | 1.86 | Provide iron to axonal mitochondria [46] |
| bcl11a | B-Cell Lymphoma/Leukemia 11A | 6.63 × 10−4 | 1.66 | Negative regulation of dendrite extension [47] |
| dmd | dystrophin, muscular dystrophy | 0.0053 | 1.61 | Synaptic transmission [48] |
| prox1 | Prospero Homeobox 1 | 0.0087 | 4.63 | Induces new lymphatic vessel formation and promotes nerve reconstruction [49] |
| Gene | VCR | |
|---|---|---|
| FC | p-Value | |
| cck2r | 8.75 | 9.21 × 10−4 **** |
| cck1r | 1.2 | 0.15 |
| cck | 10.6 | 0.78 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bernard, A.; Danigo, A.; Mroué, M.; Rovini, A.; Richard, L.; Nizou, A.; Desmoulière, A.; Sturtz, F.; Demiot, C.; Bourthoumieu, S. Blockade of Cholecystokinin Type 2 Receptors Prevents the Onset of Vincristine-Induced Neuropathy in Mice. Pharmaceutics 2022, 14, 2823. https://doi.org/10.3390/pharmaceutics14122823
Bernard A, Danigo A, Mroué M, Rovini A, Richard L, Nizou A, Desmoulière A, Sturtz F, Demiot C, Bourthoumieu S. Blockade of Cholecystokinin Type 2 Receptors Prevents the Onset of Vincristine-Induced Neuropathy in Mice. Pharmaceutics. 2022; 14(12):2823. https://doi.org/10.3390/pharmaceutics14122823
Chicago/Turabian StyleBernard, Amandine, Aurore Danigo, Mohamad Mroué, Amandine Rovini, Laurence Richard, Angélique Nizou, Alexis Desmoulière, Franck Sturtz, Claire Demiot, and Sylvie Bourthoumieu. 2022. "Blockade of Cholecystokinin Type 2 Receptors Prevents the Onset of Vincristine-Induced Neuropathy in Mice" Pharmaceutics 14, no. 12: 2823. https://doi.org/10.3390/pharmaceutics14122823