
Isopropyl Gallate, a Gallic Acid Derivative: In Silico and In Vitro Investigation of Its Effects on Leishmania major

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
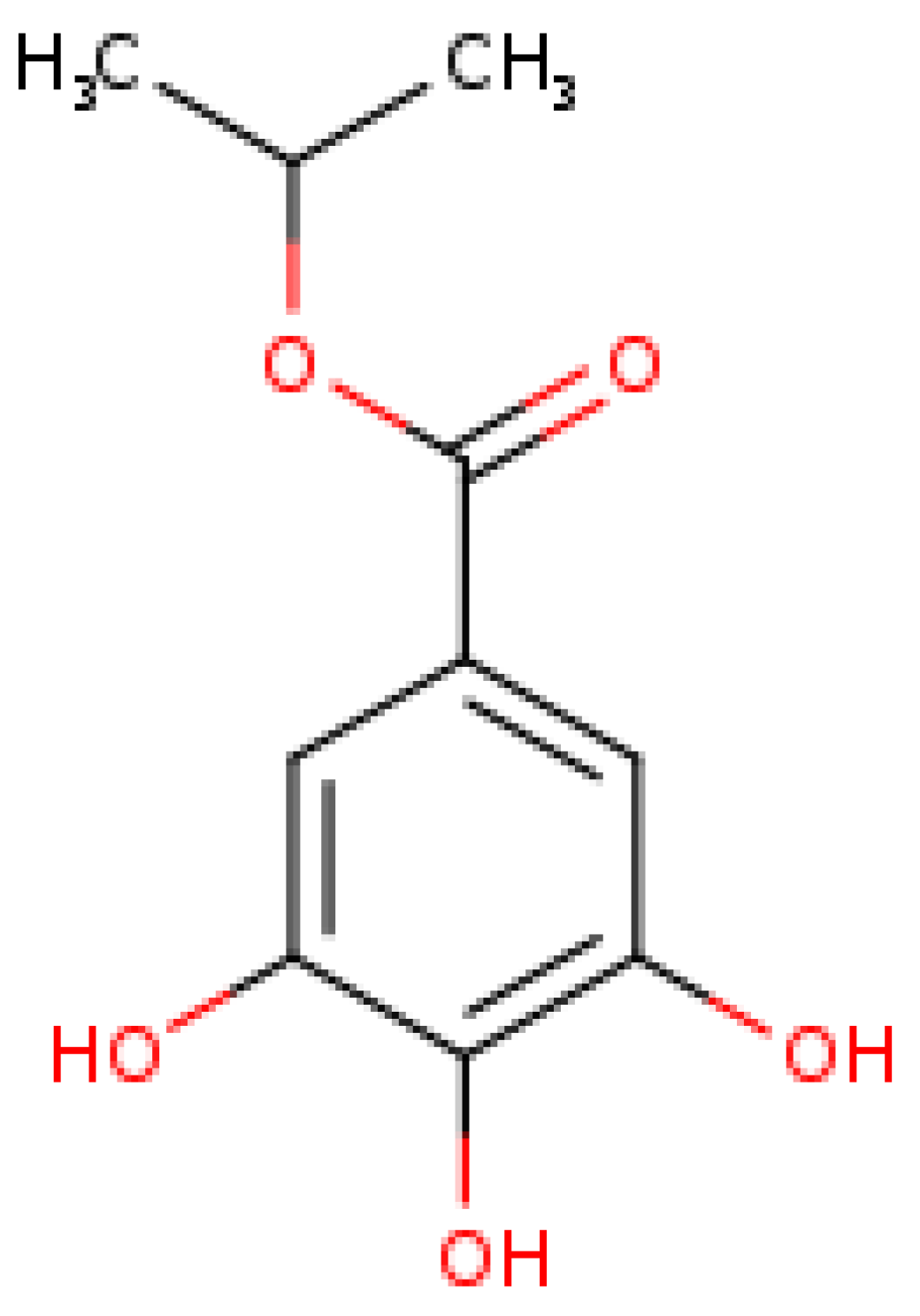
2.1. Isopropyl Gallate Compound
2.2. In Silico Analysis
2.2.1. Molecular Docking
2.2.2. Analysis of Physicochemical, Pharmacokinetic and Toxicological Properties
2.3. In Vitro Tests
2.3.1. Parasites and Cells
2.3.2. Investigation of the Antileishmanial Activity of IPG on Promastigote Forms
2.3.3. Cytotoxicity on Macrophages
2.3.4. Cell Viability in Red Cells
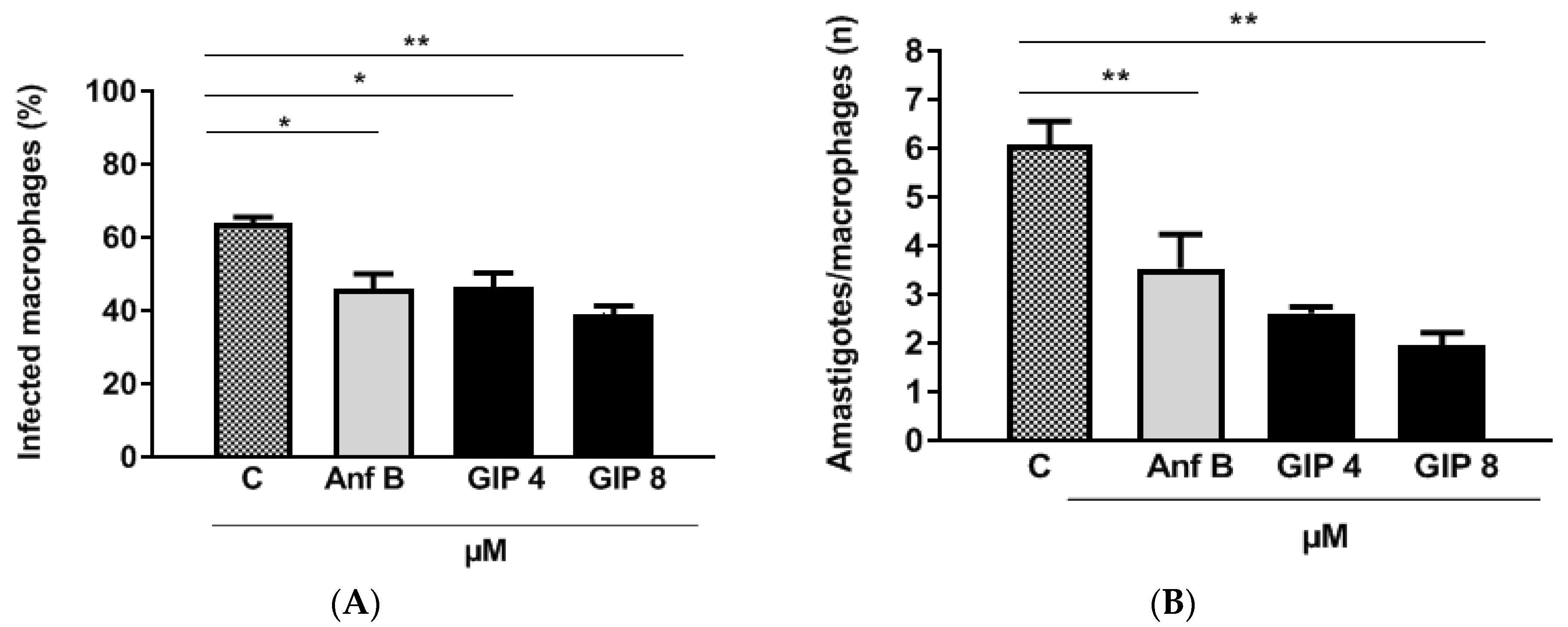
2.3.5. IPG Activity in L. major-Infected Macrophages
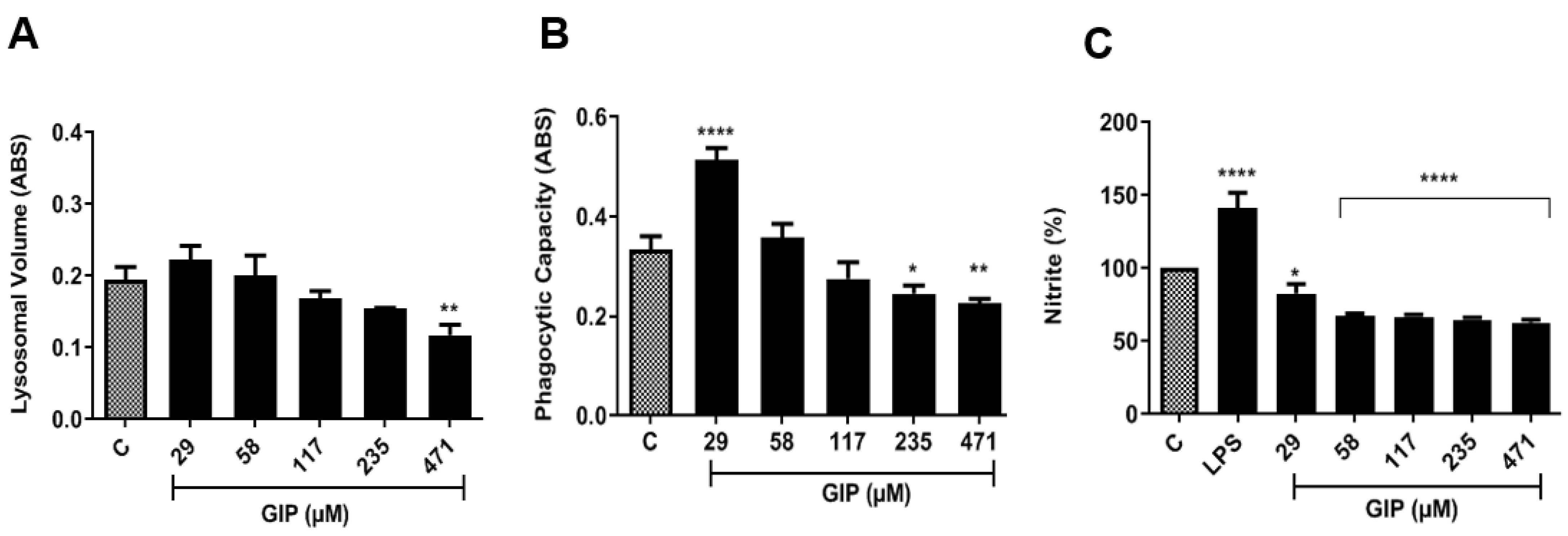
2.3.6. Evaluation of Macrophage Activation Parameters
Evaluation of Lysosomal Activity
Determination of Phagocytic Capacity
Determination of Nitrite
2.4. Statistical Analysis
3. Results and Discussion
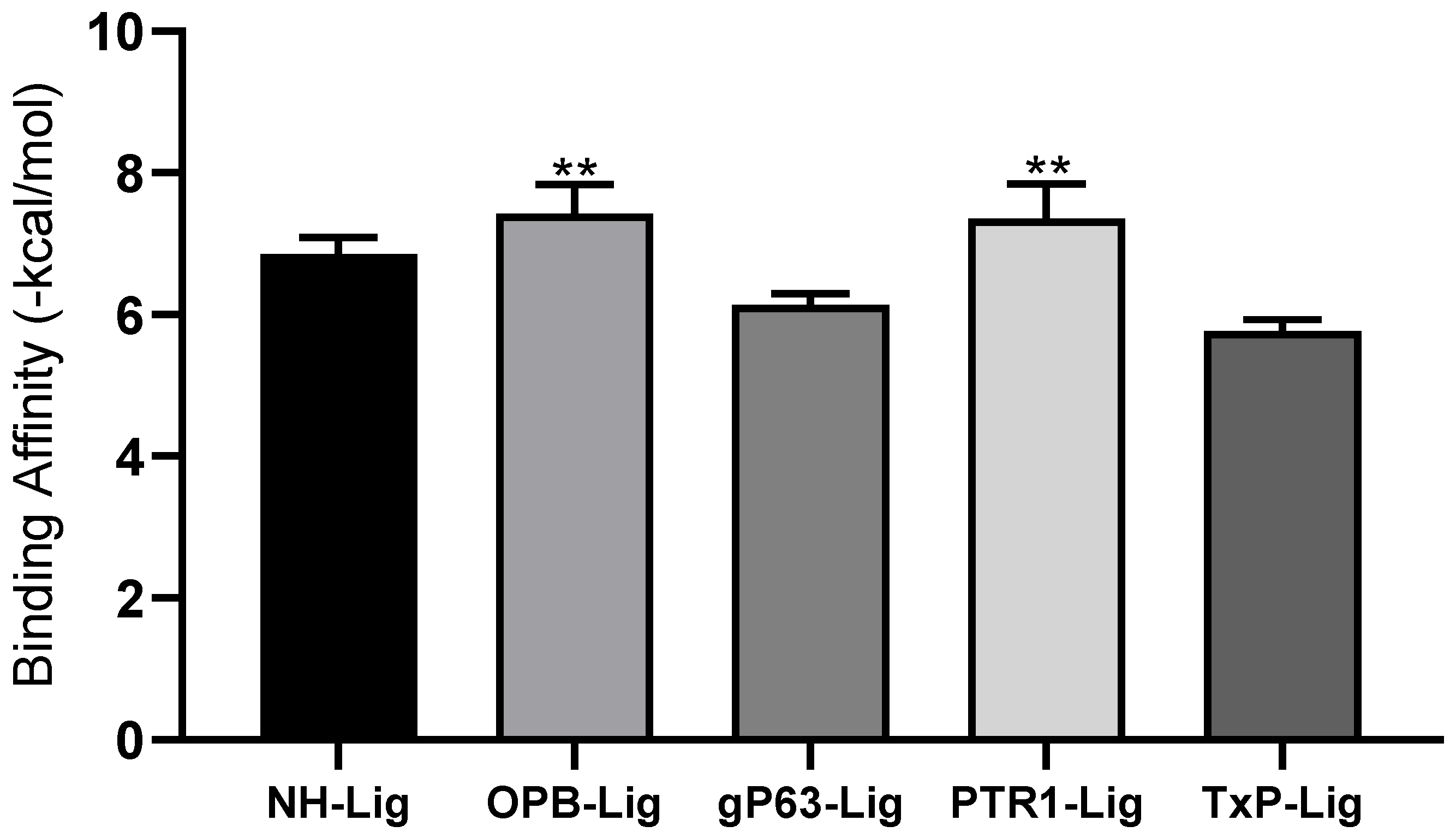
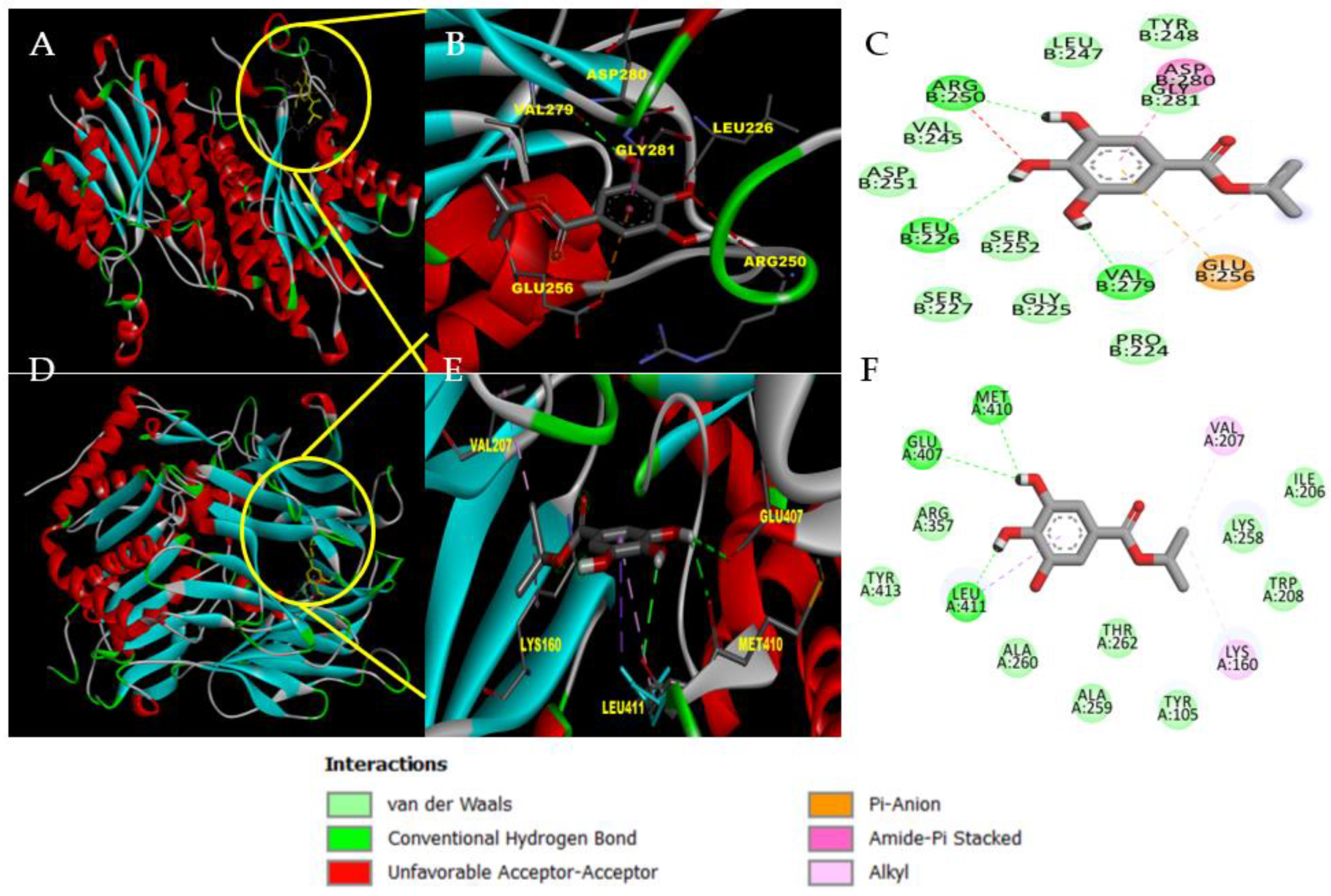
3.1. In Silico Analysis of IPG: Molecular Docking and ADMET Assessment
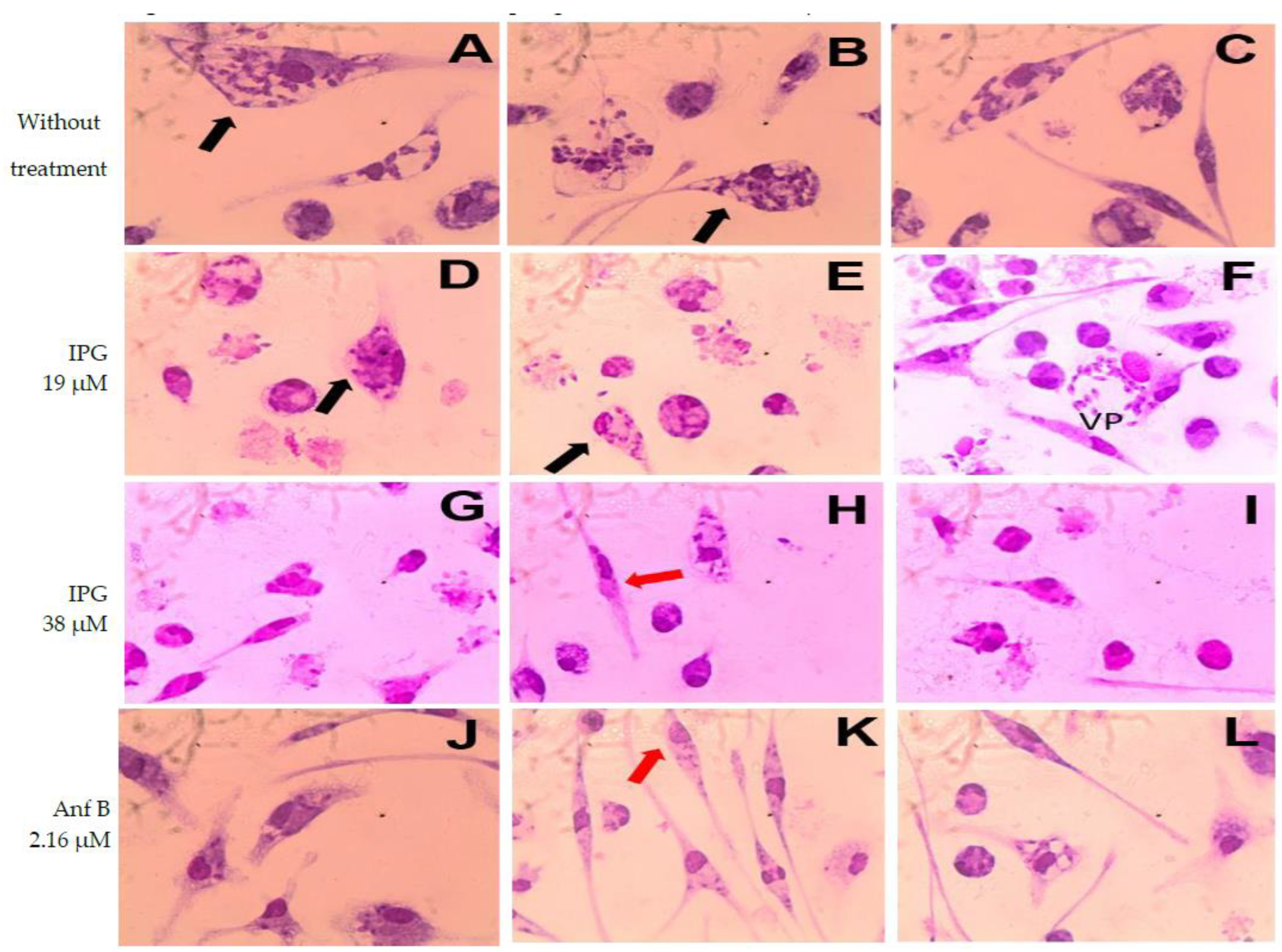
3.2. In Vitro Analysis: IPG-Induced Antileismanial Effects and Cell Viability
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Gutman, J.R.; Lucchi, N.W.; Cantey, P.T.; Steinhardt, L.C.; Samuels, A.M.; Kamb, M.L.; Kapella, B.K.; McElroy, P.D.; Udhayakumar, V.; Lindblade, K.A. Malaria and Parasitic Neglected Tropical Diseases: Potential Syndemics with COVID-19? Am. J. Trop. Med. Hyg. 2020, 103, 572–577. [Google Scholar] [CrossRef] [PubMed]
- Luna, E.J.A.; Campos, S.R.S.L.D.C. Vaccine development against neglected tropical diseases. Cad. Saude Publica 2020, 36 (Suppl. 2), e00215720. [Google Scholar] [CrossRef]
- Oliveira, R.G. Sentidos das Doenças Negligenciadas na agenda da Saúde Global: O lugar de populações e territórios [Meanings of Neglected Diseases in the Global Health agenda: The place of populations and territories]. Cien. Saude Colet 2018, 23, 2291–2302. (In Portuguese) [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hellmann, M.A.; Marchesan, E.D.; Velasquez, L.G. Leishmaniose e plantas medicinais: Uma revisão [leishmaniasis and medicinal plants: A review]. Arq. Cienc. Saúde UNIPAR 2018, 22, 217–231. (In Portuguese) [Google Scholar] [CrossRef]
- Pan American Health Organization. Leishmaniasis: Epidemiological Report of the Americas; No. 10; PAHO: Washington, DC, USA, 2021; Available online: https://iris.paho.org/handle/10665.2/51742 (accessed on 27 November 2022).
- Steverding, D. The history of leishmaniasis. Parasites Vectors 2017, 15, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yadagiri, G.; Singh, P.P. Chemotherapy and Experimental Models of Visceral Leishmaniasis. In Infectious Diseases and Your Health; Singh, P., Ed.; Springer: Singapore, 2018; pp. 63–97. [Google Scholar] [CrossRef]
- Lima, E.B.; Porto, C.; Motta, J.O.C.; Sampaio, R.N.R. Tratamento da Leishmaniose Tegumentar Americana [Treatment of american cutaneous leishmaniasis]. An. Bras. Dermatol. 2007, 82, 111–124. (In Portuguese) [Google Scholar] [CrossRef] [Green Version]
- Pimentel, M.I.; Baptista, C.; Rubin, E.F.; Vasconcellos, E.C.; Lyra, M.R.; Salgueiro, M.M.; Saheki, M.N.; Rosalino, C.M.; Madeira, M.F.; Silva, A.F.; et al. American cutaneous leishmaniasis caused by Leishmania (Viannia) braziliensis resistant to meglumine antimoniate, but with good response to pentamidine: A case report. Rev. Soc. Bras. Med. Trop. 2011, 44, 254–256. [Google Scholar] [CrossRef] [Green Version]
- Albuquerque, B.R.; Heleno, S.A.; Oliveira, M.B.P.P.; Barros, L.; Ferreira, I.C.F.R. Phenolic compounds: Current industrial applications, limitations and future challenges. Food Funct 2021, 12, 14–29. [Google Scholar] [CrossRef]
- Alves, M.M.M.; Brito, L.M.; Souza, A.C.; Queiroz, B.C.S.H.; Carvalho, T.P.; Batista, J.F.; Oliveira, J.S.S.M.; Mendonça, I.L.; Lira, S.R.S.; Chaves, M.H.; et al. Gallic and ellagic acids: Two natural immunomodulator compounds solve infection of macrophages by Leishmania major. Naunyn Schmiedebergs Arch. Pharmacol. 2017, 390, 893–903. [Google Scholar] [CrossRef]
- Xu, Y.; Tang, G.; Zhang, C.; Wang, N.; Feng, Y. Gallic Acid and Diabetes Mellitus: Its Association with Oxidative Stress. Molecules 2021, 26, 7115. [Google Scholar] [CrossRef]
- Steverding, D.; do Nascimento, L.G.; Perez-Castillo, Y.; de Sousa, D.P. Gallic Acid Alkyl Esters: Trypanocidal and Leishmanicidal Activity, and Target Identification via Modeling Studies. Molecules 2022, 27, 5876. [Google Scholar] [CrossRef] [PubMed]
- Almeida de Oliveira, L.S.; de Moura Bandeira, S.R.; Gomes Gonçalves, R.L.; Pereira de Sousa Neto, B.; Carvalho de Rezende, D.; dos Reis-Filho, A.C.; Sousa, I.J.O.; Pinheiro-Neto, F.R.; Timah Acha, B.; do Nascimento Caldas Trindade, G.; et al. The Isopropyl Gallate Counteracts Cyclophosphamide-Induced Hemorrhagic Cystitis in Mice. Biology 2022, 11, 728. [Google Scholar] [CrossRef]
- Figueiredo, K.A.; Figueiredo, J.F.S.; Costa, R.K.M.; Alves, M.M.M.; Magalhães, J.L.; Carvalho, A.L.M.; Lima, F.C.A. Prospecção de Alvos Bioquímicos Para Estudo in silico na Quimioterapia Antileishmania [Prospecting Biochemical Targets for in silico Study for Antileishmania Chemotherapy]. Rev. Virtual Quim. 2018, 10, 1485–1501. (In Portuguese) [Google Scholar] [CrossRef]
- Eberhardt, J.; Santos-Martins, D.; Tillack, A.F.; Forli, S. AutoDock Vina 1.2.0: New Docking Methods, Expanded Force Field, and Python Bindings. J. Chem. Inf. Model. 2021, 61, 3891–3898. [Google Scholar] [CrossRef] [PubMed]
- Trott, O.; Olson, A. AutoDock Vina: Improving the Speed and Accuracy of Docking with a New Scoring Function, Efficient Optimization, and Multithreading. J. Comput. Chem. 2010, 31, 455–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Daina, A.; Michielin, O.; Zoete, V. SwissADME: A free web tool to evaluate pharmacokinetics, drug-likeness and medicinal chemistry friendliness of small molecules. Sci. Rep. 2017, 7, 42717. [Google Scholar] [CrossRef] [Green Version]
- Lee, S.K.; Lee, I.H.; Kim, H.J.; Chang, G.S.; Chung, J.E.; No, K.T. The PreADME Approach: Web-based program for rapid prediction of physico-chemical, drug absorption and drug-like Properties. In EuroQSAR 2002 Designing Drugs and Crop Protectants: Processes, Problems and Solutions; Blackwell Publishing: Cambridge, MA, USA, 2003; pp. 418–420. [Google Scholar]
- Cheng, F.; Li, W.; Zhou, Y.; Shen, J.; Wu, Z.; Liu, G.; Lee, P.W.; Tang, Y. admetSAR: A comprehensive source and free tool for evaluating chemical ADMET properties. J. Chem. Inf. Model. 2012, 52, 3099–3105. [Google Scholar] [CrossRef]
- Cruz, J.H.d.A.; Moreira, I.C.d.S.; Alves, M.d.F.V.; Oliveira, H.M.B.F.d.; Oliveira Filho, A.A.d.; Alves, M.A.S.G. Análise Da Atividade farmacológica E toxicológica Do Monoterpeno Relacionado à Odontologia: Estudo in Silico [Analysis of pharmacological and toxicological activity of neral monoterpenerelated to Dentistry: Study in silico]. Arch. Health Investig. 2019, 8, 674–680. (In Portuguese) [Google Scholar] [CrossRef]
- Carneiro, S.M.P.; Carvalho, F.A.A.; Santana, L.C.L.R.; Sousa, A.P.L.; Moita-Neto, J.M.; Chaves, M.H. The cytotoxic and antileishmanial activity of extracts and fractions of leaves and fruits of Azadirachta indica (A Juss.). Biol. Res. 2012, 45, 111–116. [Google Scholar] [CrossRef]
- Costa, L.M.; Alves, M.M.M.; Brito, L.M.; Abi-Chacra, E.A.; Barbosa-Filho, J.M.; Gutierrez, S.J.C.; Barreto, H.M.; Carvalho, F.A.A. In vitro antileishmanial and immunomodulatory activities of the synthetic analogue riparin E. Chem. Biol. Interact. 2021, 336, 109389. [Google Scholar] [CrossRef]
- Löfgren, S.E.; Miletti, L.C.; Steindel, M.; Bachère, E.; Barracco, M.A. Trypanocidal and leishmanicidal activities of different antimicrobial peptides (AMPs) isolated from aquatic animals. Exp. Parasitol. 2008, 118, 197–202. [Google Scholar] [CrossRef]
- Oliveira, L.G.C.; Brito, L.M.; Alves, M.M.M.; Amorim, L.V.; Sobrinho-Júnior, E.P.; Carvalho, C.E.S.; Rodrigues, K.A.F.; Arcanjo, D.D.R.; Citó, A.M.G.L.; Carvalho, F.A.A. In vitro effects of the Neolignan 2, 3-Dihydrobenzofuran against Leishmania amazonensis. Basic Clin. Pharmacol. Toxicol. 2017, 120, 52–58. [Google Scholar] [CrossRef]
- Grando, F.C.; Felício, C.A.; Twardowschy, A.; Paula, F.M.; Batista, V.G.; Fernandes, L.C.; Curi, R.; Nishiyama, A. Modulation of peritoneal macrophage activity by the saturation state of the fatty acid moiety of phosphatidylcholine. Braz. J. Med. Biol. Res. 2009, 42, 599–605. [Google Scholar] [CrossRef]
- Soares, D.C.; Pereira, C.G.; Meireles, M.A.A.; Saraiva, E.M. Leishmanicidal activity of a supercritical fluid fraction obtained from Tabernaemontana catharinensis. Parasitol. Int. 2007, 56, 135–139. [Google Scholar] [CrossRef]
- Mares-Sámano, S.; Garduño-Juarez, R. Computational Modeling of the Interactions of Drugs with Human Serum Albumin (HSA). Comp. Sist. 2021, 22, 1123–1135. [Google Scholar] [CrossRef] [Green Version]
- Munday, J.C.; McLuskey, K.; Brown, E.; Coombs, G.H.; Mottram, J.C. Oligopeptidase B deficient mutants of Leishmania major. Mol. Biochem. Parasitol. 2011, 175, 49–57. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lipinski, C.A. Lead- and drug-like compounds: The rule-of-five revolution. Drug Discov. Today Technol. 2004, 1, 337–341. [Google Scholar] [CrossRef]
- Sun, N.; Agyei, D.; Ji, D. Medicinal Chemistry Friendliness of Pigments from Monascus-Fermented Rice and the Molecular Docking Analysis of Their Anti-Hyperlipidemia Properties. Fermentation 2020, 6, 111. [Google Scholar] [CrossRef]
- Kim, I.; Kim, H.; Ro, J.; Jo, K.; Karki, S.; Khadka, P.; Yun, G.; Lee, J. Preclinical pharmacokinetic evaluation of β-lapachone: Characteristics of oral bioavailability and first-pass metabolism in rats. Biomol. Ther. 2015, 23, 296–300. [Google Scholar] [CrossRef]
- Pham-The, H.; Casañola-Martin, G.; Garrigues, T.; Bermejo, M.; González-Álvarez, I.; Nguyen-Hai, N.; Cabrera-Pérez, M.Á.; Le-Thi-Thu, H. Exploring different strategies for imbalanced ADME data problem: Case study on caco-2 permeability modeling. Mol. Divers 2016, 20, 93–109. [Google Scholar] [CrossRef]
- De Souza, J.; Freitas, Z.M.F.; Storpirtis, S. Modelos in vitro para determinação da absorção de fármacos e previsão da relação dissolução/absorção [In vitro models for the determination of drug absorption and a prediction of dissolution/absorption relationships]. Rev. Bras. Cienc. Farm 2007, 43, 515–527. (In Portuguese) [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Yang, H.; Li, J.; Wang, T.; Li, W.; Liu, G.; Tang, Y. In Silico Prediction of Compounds Binding to Human Plasma Proteins by QSAR Models. ChemMedChem 2018, 13, 572–581. [Google Scholar] [CrossRef]
- Zhang, H.; Xu, H.; Ashby, C.R., Jr.; Assaraf, Y.G.; Chen, Z.S.; Liu, H.M. Chemical molecular-based approach to overcome multidrug resistance in cancer by targeting P-glycoprotein (P-gp). Med. Res. Rev. 2021, 41, 525–555. [Google Scholar] [CrossRef]
- Kitagawa, S.; Nabekura, T.; Kamiyama, S.; Takahashi, T.; Nakamura, Y.; Kashiwada, Y.; Ikeshiro, Y. Effects of alkyl gallates on P-glycoprotein function. Biochem. Pharmacol. 2005, 70, 1262–1266. [Google Scholar] [CrossRef]
- Ma, X.; Chen, C.; Yang, J. Predictive model of blood-brain barrier penetration of organic compounds. Acta Pharmacol. Sin. 2005, 26, 500–512. [Google Scholar] [CrossRef] [Green Version]
- Yang, J.T.; Lee, I.N.; Lu, F.J.; Chung, C.Y.; Lee, M.H.; Cheng, Y.C.; Chen, K.T.; Chen, C.H. Propyl Gallate Exerts an Antimigration Effect on Temozolomide-Treated Malignant Glioma Cells through Inhibition of ROS and the NF-κB Pathway. J. Immunol. Res. 2017, 2017, 9489383. [Google Scholar] [CrossRef] [Green Version]
- Petersen, C.A.; Greenlee, M.H.W. Neurologic Manifestations of Leishmania spp. Infection. J. Neuroparasitol. 2011, 2, N110401. [Google Scholar] [CrossRef]
- Mitragotri, S.; Anissimov, Y.G.; Bunge, A.L.; Frasch, H.F.; Guy, R.H.; Hadgraft, J.; Kasting, G.B.; Lane, M.E.; Roberts, M.S. Mathematical models of skin permeability: An overview. Int. J. Pharm. 2011, 418, 115–129. [Google Scholar] [CrossRef] [Green Version]
- Alves, M.M.M.; Arcanjo, D.D.R.; Figueiredo, K.A.; Oliveira, J.S.S.M.; Viana, F.J.C.; Coelho, E.S.; Lopes, G.L.N.; Gonçalves, J.C.R.; Carvalho, A.L.M.; Rizzo, M.S.; et al. Gallic and Ellagic Acids Are Promising Adjuvants to Conventional Amphotericin B for the Treatment of Cutaneous Leishmaniasis. Antimicrob. Agents Chemother. 2020, 64, e00807-20. [Google Scholar] [CrossRef]
- Kirchmair, J.; Göller, A.H.; Lang, D.; Kunze, J.; Testa, B.; Wilson, I.D.; Glen, R.C.; Schneider, G. Predicting drug metabolism: Experiment and/or computation? Nat. Rev. Drug Discov. 2015, 14, 387–404. [Google Scholar] [CrossRef] [Green Version]
- Zhou, P.Z.; Babcock, J.; Liu, L.Q.; Li, M.; Gao, Z.B. Activation of human ether-a-go-go related gene (hERG) potassium channels by small molecules. Acta Pharmacol. Sin. 2011, 32, 781–788. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Chen, L.; Cheng, F.; Wu, Z.; Bian, H.; Xu, C.; Li, W.; Liu, G.; Shen, X.; Tang, Y. In Silico Prediction of Chemical Acute Oral Toxicity Using Multi-Classification Methods. J. Chem. Inf. Model. 2014, 54, 1061–1069. [Google Scholar] [CrossRef]
- Déciga-Campos, M.; Navarrete-Vázquez, G.; López-Muñoz, F.J.; Librowski, T.; Sánchez-Recillas, A.; Yañez-Pérez, V.; Ortiz-Andrade, R. Complementary pharmacological and toxicological characterization data on the pharmacological profile of N-(2,6-dichlorophenyl)-2-(4-methyl-1-piperidinyl) acetamide. Data Brief 2016, 8, 1007–1012. [Google Scholar] [CrossRef]
- Tayama, S.; Nakagawa, Y. Cytogenetic effects of propyl gallate in CHO-K1 cells. Mutat. Res. 2001, 498, 117–127. [Google Scholar] [CrossRef] [PubMed]
- Koutsoni, O.S.; Karampetsou, K.; Kyriazis, I.D.; Stathopoulos, P.; Aligiannis, N.; Halabalaki, M.; Skaltsounis, L.A.; Dotsika, E. Evaluation of total phenolic fraction derived from extra virgin olive oil for its antileishmanial activity. Phytomedicine 2018, 47, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Kolodziej, H.; Kiderlen, A.F. Antileishmanial activity and immune modulatory effects of tannins and related compounds on Leishmania parasitised RAW 264.7 cells. Phytochemistry 2005, 66, 2056–2071. [Google Scholar] [CrossRef]
- Don, R.; Ioset, J.R. Screening strategies to identify new chemical diversity for drug development to treat kinetoplastid infections. Parasitology 2014, 141, 140–146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wei, W.C.; Su, Y.H.; Chen, S.S.; Sheu, J.H.; Yang, N.S. GM-CSF plays a key role in zymosan-stimulated human dendritic cells for activation of Th1 and Th17 cells. Cytokine 2011, 55, 79–89. [Google Scholar] [CrossRef]
- Hipolito, V.E.B.; Diaz, J.A.; Tandoc, K.V.; Oertlin, C.; Ristau, J.; Chauhan, N.; Saric, A.; Mclaughlan, S.; Larsson, O.; Topisirovic, I.; et al. Enhanced translation expands the endo-lysosome size and promotes antigen presentation during phagocyte activation. PLoS Biol. 2019, 17, e3000535. [Google Scholar] [CrossRef]
- Bezerra, É.A.; Alves, M.M.M.; Lima, S.K.R.; Pinheiro, E.E.A.; Amorim, L.V.; Lima Neto, J.S.; Carvalho, F.A.A.; Citó, A.M.G.L.; Arcanjo, D.D.R. Biflavones from Platonia insignis Mart. Flowers Promote In Vitro Antileishmanial and Immunomodulatory Effects Against Internalized Amastigote Forms of Leishmania amazonensis. Pathogens 2021, 10, 1166. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targets | PDB * | Metabolic Pathway/Function |
|---|---|---|
| Nucleoside Hydrolase (NH) | 1EZR | Nucleic acid metabolism |
| Oligopeptidase B (OPB) | 2XE4 | Protease/virulence factor |
| Leishmanolysin Proteinase (Gp63) | 1LML | Virulence factor |
| Pteridine Reductase (PTR1) | 1E7W | Nucleic acid metabolism |
| Triparedoxin peroxidase (TxP) | 3TUE | Redox metabolism (parasite survival) |
| GIP Molecule | Targets | ||||
|---|---|---|---|---|---|
| Interaction energy (kcal/mol) | NH | OPB | Gp63 | PTR1 | TxP |
| −7.3 | −8.0 | −6.4 | −8.2 | −6.0 | |
| Properties | Parameter | Prediction |
|---|---|---|
| Molecular Weight (g/mol) | 212.2 | |
| Log of lipophilicity (Log P) | 1.37 | |
| Physical Chemistry | Aqueous solubility (Log S) | −2.22 |
| H-bond donors (HBD) | 3 | |
| H-Bond Acceptors (HBA) | 5 | |
| Human oral bioavailability (%) | 0.5429 | |
| Human intestinal absorption (%) | 0.9963 | |
| Caco-2 permeability coefficient (nm/s) | 0.5405 | |
| Plasma protein binding (PPB) (%) | 76.0 | |
| Inhibition of P-glycoprotein (gp-P) | Yes | |
| Pharmacokinetics | Penetration of the blood–brain barrier | 0.8301 |
| Skin permeability | −3.3007 | |
| CYP2C19 inhibitor | No | |
| CYP2C9 inhibitor | No | |
| CYP1A2 inhibitor | No | |
| CYP2D6 inhibitor | No | |
| CYP3A4 inhibitor | No | |
| Organ Toxicity | ||
| Drug-induced liver-injury assay Human ether-a-go-go gene (hERG) K+ channels | - | |
| Toxicological | Acute oral toxicity * | III |
| Eye injuries | + | |
| Eye corrosion | - | |
| Genomic toxicity | ||
| Ames mutagenesis | - | |
| Carcinogenesis | - | |
| Micronucleus assay | + |
| Substance | Macrophage CC50 (μM) | Erythrocyte CH50 (μM) | Promastigote IC50 (μM) | Amastigote IC50 (μM) | ISm | ISe |
|---|---|---|---|---|---|---|
| IPG | 1260 ± 0.48 | >3770 | 90.81 ± 0.51 | 13.45 ± 0.35 | 93.8 | >280.3 |
| Anf B | 8.97 ± 0.02 | ND * | 1.40 ± 0.09 | 0.90 ± 0.36 | 9.97 | ND * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Melo, D.S.d.; Nery Neto, J.A.d.O.; Santos, M.d.S.d.; Pimentel, V.D.; Carvalho, R.d.C.V.; Sousa, V.C.d.; Sousa, R.G.C.; Nascimento, L.G.d.; Alves, M.M.d.M.; Arcanjo, D.D.R.; et al. Isopropyl Gallate, a Gallic Acid Derivative: In Silico and In Vitro Investigation of Its Effects on Leishmania major. Pharmaceutics 2022, 14, 2701. https://doi.org/10.3390/pharmaceutics14122701
Melo DSd, Nery Neto JAdO, Santos MdSd, Pimentel VD, Carvalho RdCV, Sousa VCd, Sousa RGC, Nascimento LGd, Alves MMdM, Arcanjo DDR, et al. Isopropyl Gallate, a Gallic Acid Derivative: In Silico and In Vitro Investigation of Its Effects on Leishmania major. Pharmaceutics. 2022; 14(12):2701. https://doi.org/10.3390/pharmaceutics14122701
Chicago/Turabian StyleMelo, Danielly Silva de, José Arimatéa de Oliveira Nery Neto, Maisa de Sousa dos Santos, Vinícius Duarte Pimentel, Rita de Cássia Viana Carvalho, Valéria Carlos de Sousa, Ruy Gabriel Costa Sousa, Lázaro Gomes do Nascimento, Michel Muálem de Moraes Alves, Daniel Dias Rufino Arcanjo, and et al. 2022. "Isopropyl Gallate, a Gallic Acid Derivative: In Silico and In Vitro Investigation of Its Effects on Leishmania major" Pharmaceutics 14, no. 12: 2701. https://doi.org/10.3390/pharmaceutics14122701