
A Transversal Approach Combining In Silico, In Vitro and In Vivo Models to Describe the Metabolism of the Receptor Interacting Protein 1 Kinase Inhibitor Sibiriline
 , , and
, , and
Abstract
:1. Introduction
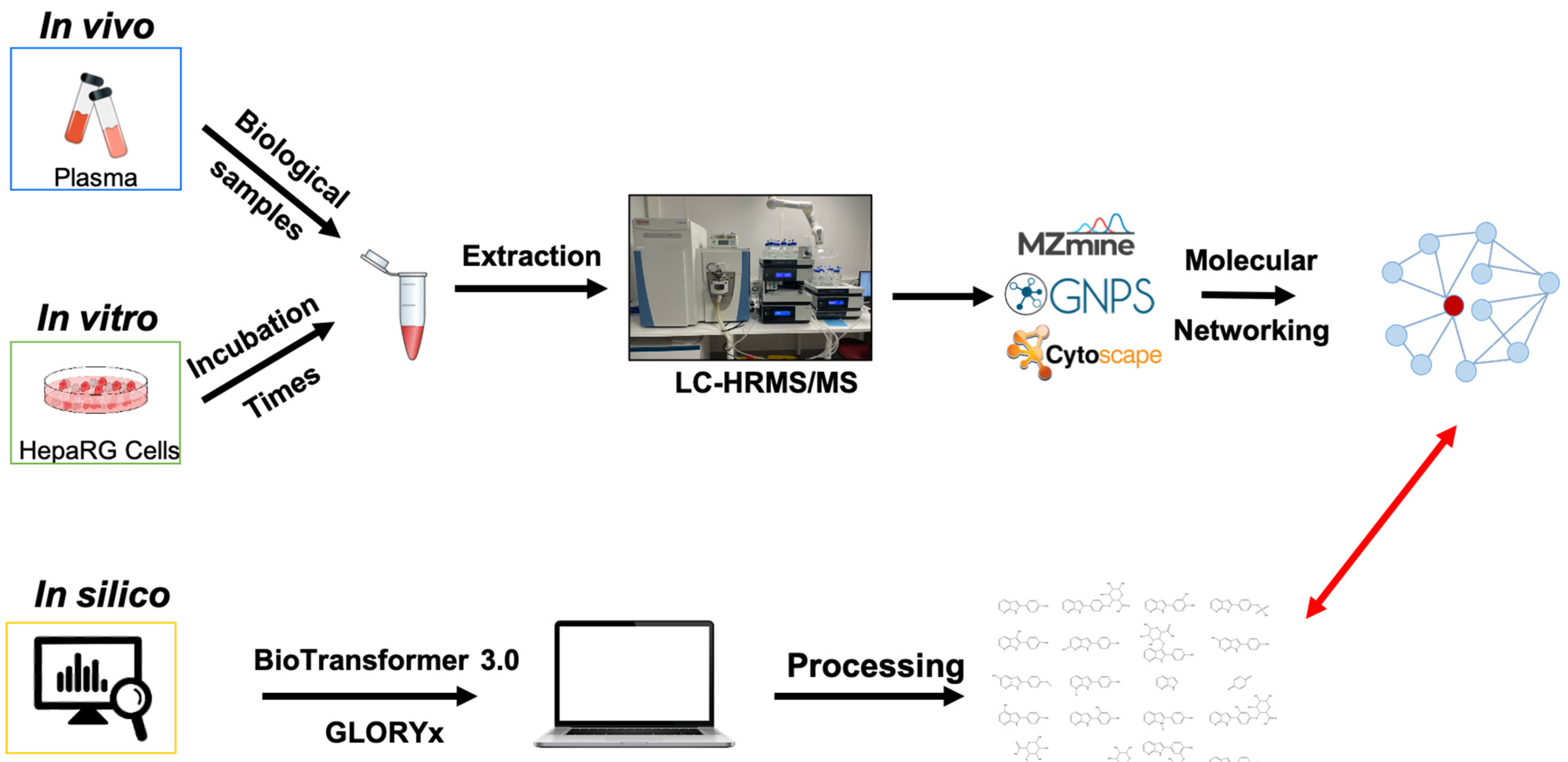
2. Materials and Methods
2.1. In Silico Metabolite Prediction
2.2. Pharmacokinetic Study in Mice
2.3. In Vitro and In Vivo Metabolism Studies
3. Results
3.1. In Silico Prediction of Sibiriline Metabolism
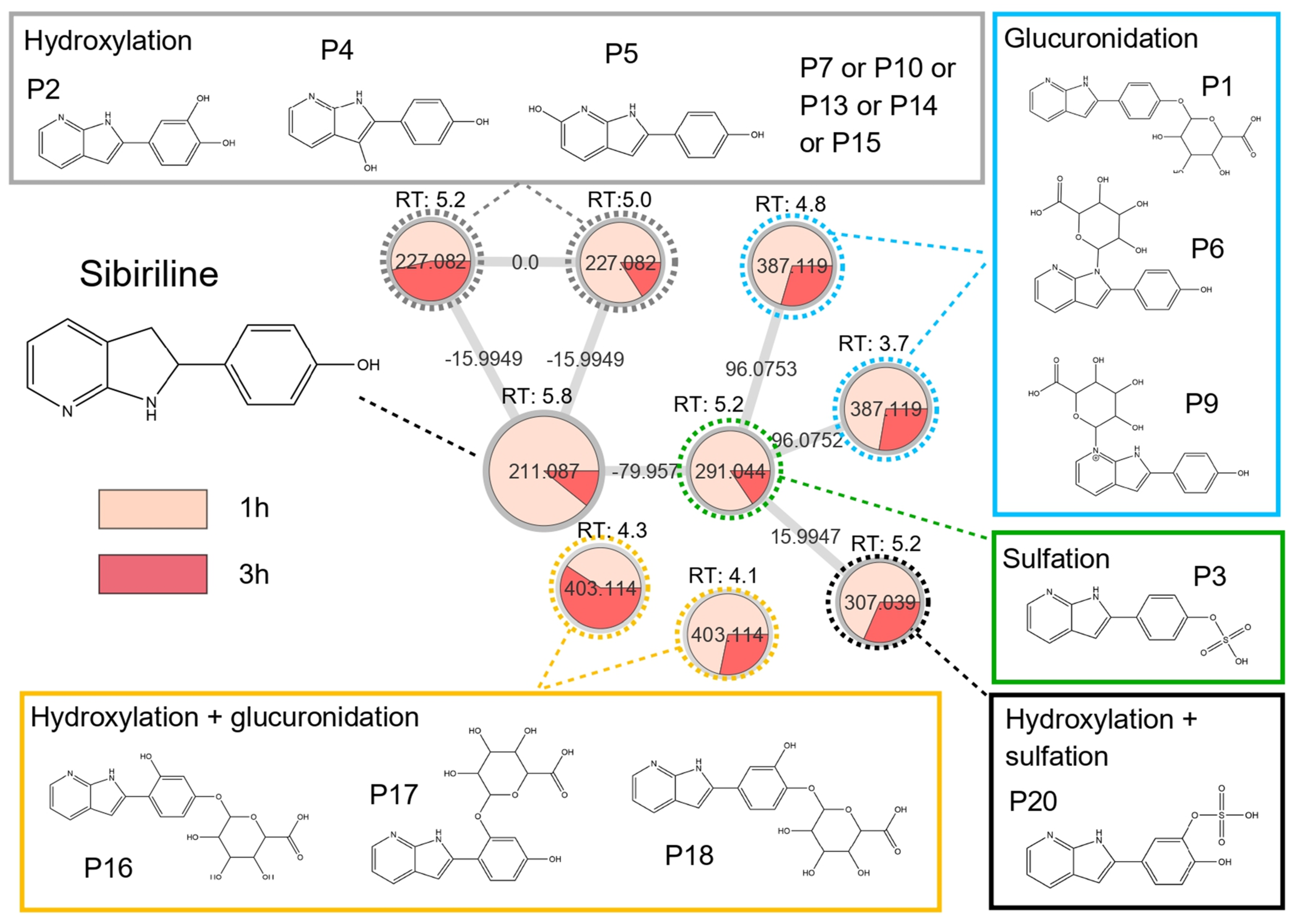
3.2. In Vitro Sibiriline Metabolism
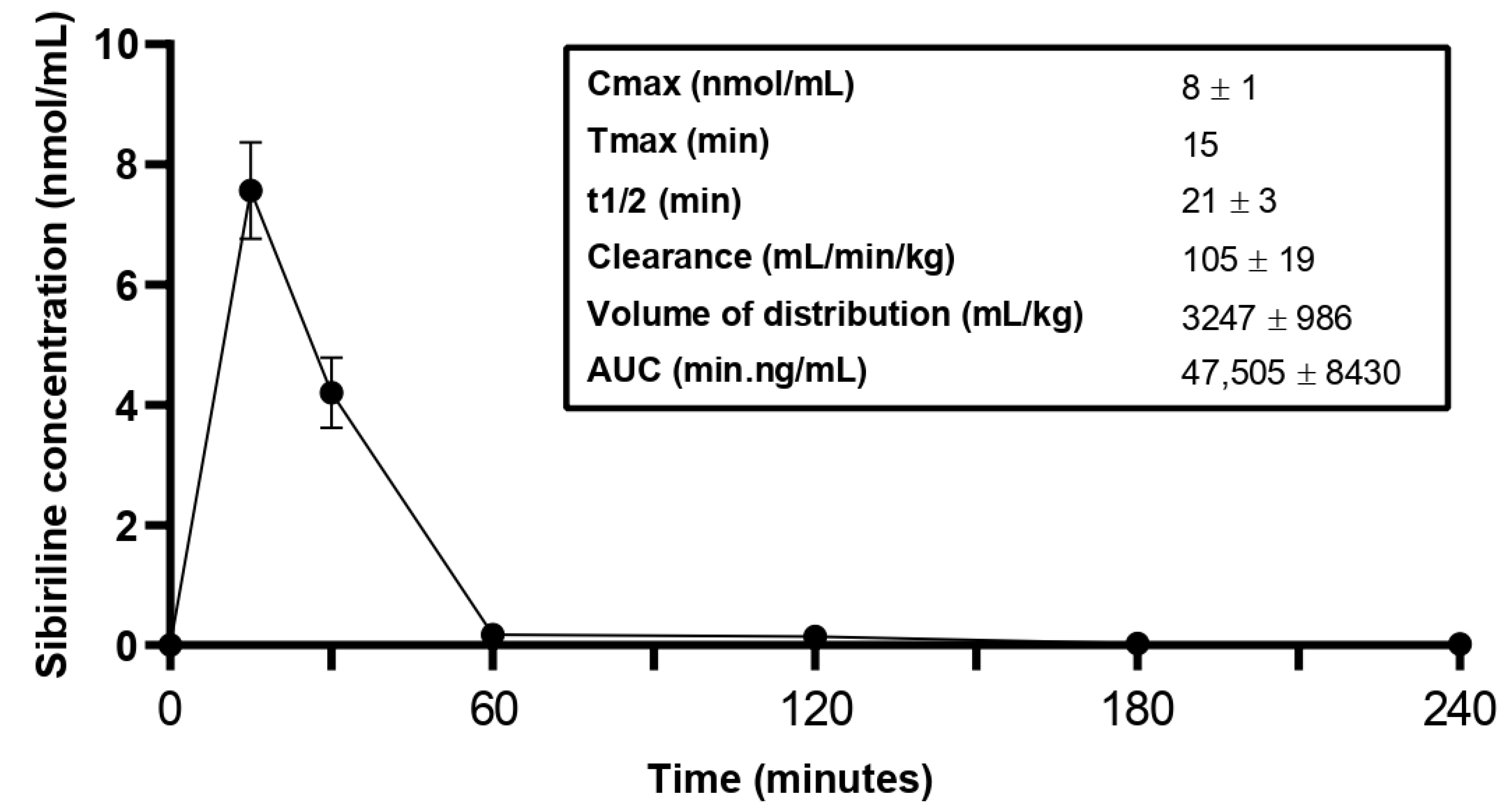
3.3. Sibiriline Pharmacokinetic Study in Mice
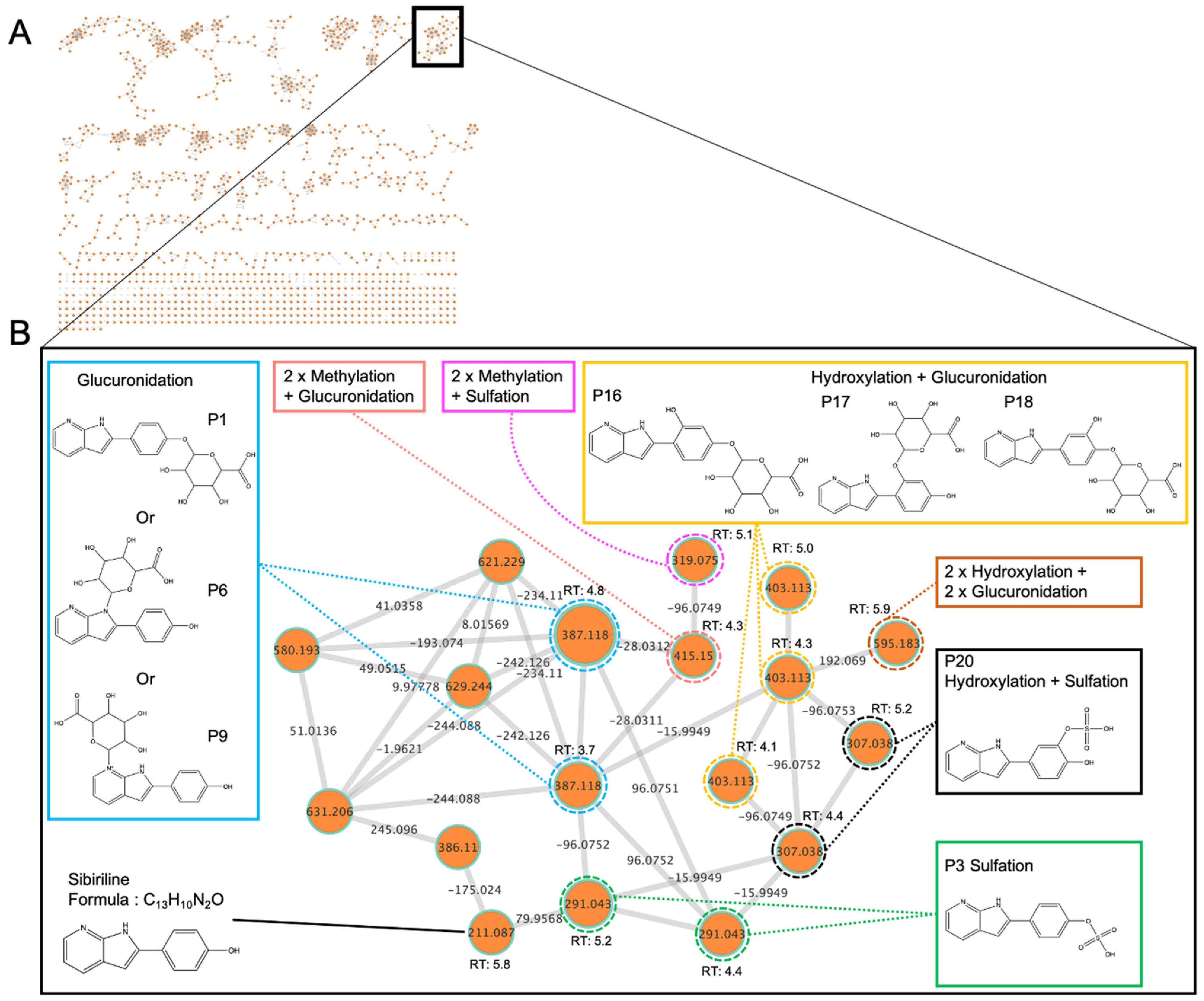
3.4. In Vivo Sibiriline Metabolism
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harris, P.A.; Berger, S.B.; Jeong, J.U.; Nagilla, R.; Bandyopadhyay, D.; Campobasso, N.; Capriotti, C.A.; Cox, J.A.; Dare, L.; Dong, X.; et al. Discovery of a First-in-Class Receptor Interacting Protein 1 (RIP1) Kinase Specific Clinical Candidate (GSK2982772) for the Treatment of Inflammatory Diseases. J. Med. Chem. 2017, 60, 1247–1261. [Google Scholar] [CrossRef]
- Weisel, K.; Scott, N.E.; Tompson, D.J.; Votta, B.J.; Madhavan, S.; Povey, K.; Wolstenholme, A.; Simeoni, M.; Rudo, T.; Richards-Peterson, L.; et al. Randomized Clinical Study of Safety, Pharmacokinetics, and Pharmacodynamics of RIPK1 Inhibitor GSK2982772 in Healthy Volunteers. Pharmacol. Res. Perspect. 2017, 5, e00365. [Google Scholar] [CrossRef]
- Sheridan, C. Death by Inflammation: Drug Makers Chase the Master Controller. Nat. Biotechnol. 2019, 37, 111–113. [Google Scholar] [CrossRef]
- Martens, S.; Hofmans, S.; Declercq, W.; Augustyns, K.; Vandenabeele, P. Inhibitors Targeting RIPK1/RIPK3: Old and New Drugs. Trends Pharmacol. Sci. 2020, 41, 209–224. [Google Scholar] [CrossRef]
- Le Cann, F.; Delehouzé, C.; Leverrier-Penna, S.; Filliol, A.; Comte, A.; Delalande, O.; Desban, N.; Baratte, B.; Gallais, I.; Piquet-Pellorce, C.; et al. Sibiriline, a New Small Chemical Inhibitor of Receptor-Interacting Protein Kinase 1, Prevents Immune-Dependent Hepatitis. FEBS J. 2017, 284, 3050–3068. [Google Scholar] [CrossRef] [Green Version]
- Cerec, V.; Glaise, D.; Garnier, D.; Morosan, S.; Turlin, B.; Drenou, B.; Gripon, P.; Kremsdorf, D.; Guguen-Guillouzo, C.; Corlu, A. Transdifferentiation of Hepatocyte-like Cells from the Human Hepatoma HepaRG Cell Line through Bipotent Progenitor. Hepatology 2007, 45, 957–967. [Google Scholar] [CrossRef]
- Guguen-Guillouzo, C.; Guillouzo, A. Setup and Use of HepaRG Cells in Cholestasis Research. In Experimental Cholestasis Research; Vinken, M., Ed.; Methods in Molecular Biology; Springer: New York, NY, USA, 2019; Volume 1981, pp. 291–312. ISBN 978-1-4939-9419-9. [Google Scholar]
- Guillouzo, A.; Corlu, A.; Aninat, C.; Glaise, D.; Morel, F.; Guguen-Guillouzo, C. The Human Hepatoma HepaRG Cells: A Highly Differentiated Model for Studies of Liver Metabolism and Toxicity of Xenobiotics. Chem. Biol. Interact. 2007, 168, 66–73. [Google Scholar] [CrossRef]
- Le Daré, B.; Ferron, P.-J.; Allard, P.-M.; Clément, B.; Morel, I.; Gicquel, T. New Insights into Quetiapine Metabolism Using Molecular Networking. Sci. Rep. 2020, 10, 19921. [Google Scholar] [CrossRef]
- Aninat, C.; Piton, A.; Glaise, D.; Le Charpentier, T.; Langouët, S.; Morel, F.; Guguen-Guillouzo, C.; Guillouzo, A. Expression of Cytochromes P450, Conjugating Enzymes and Nuclear Receptors in Human Hepatoma HepaRG Cells. Drug Metab. Dispos. 2006, 34, 75–83. [Google Scholar] [CrossRef] [Green Version]
- Hugbart, C.; Verres, Y.; Le Daré, B.; Bucher, S.; Vène, E.; Bodin, A.; Lagente, V.; Fromenty, B.; Bouvet, R.; Morel, I.; et al. Non-Oxidative Ethanol Metabolism in Human Hepatic Cells in Vitro: Involvement of Uridine Diphospho-Glucuronosyltransferase 1A9 in Ethylglucuronide Production. Toxicol. Vitr. 2020, 66, 104842. [Google Scholar] [CrossRef]
- Quesnot, N.; Bucher, S.; Gade, C.; Vlach, M.; Vene, E.; Valença, S.; Gicquel, T.; Holst, H.; Robin, M.-A.; Loyer, P. Production of Chlorzoxazone Glucuronides via Cytochrome P4502E1 Dependent and Independent Pathways in Human Hepatocytes. Arch. Toxicol. 2018, 92, 3077–3091. [Google Scholar] [CrossRef]
- Richeval, C.; Gicquel, T.; Hugbart, C.; Le Dare, B.; Allorge, D.; Morel, I.; Gaulier, J. In Vitro Characterization of NPS Metabolites Produced by Human Liver Microsomes and the HepaRG Cell Line Using Liquid Chromatographyhigh Resolution Mass Spectrometry (LC-HRMS) Analysis: Application to Furanyl Fentanyl. Curr. Pharm. Biotechnol. 2018, 18, 806–814. [Google Scholar] [CrossRef]
- Katoh, M.; Sawada, T.; Soeno, Y.; Nakajima, M.; Tateno, C.; Yoshizato, K.; Yokoi, T. In Vivo Drug Metabolism Model for Human Cytochrome P450 Enzyme Using Chimeric Mice with Humanized Liver. J. Pharm. Sci. 2007, 96, 428–437. [Google Scholar] [CrossRef]
- Park, Y.M.; Meyer, M.R.; Müller, R.; Herrmann, J. Drug Administration Routes Impact the Metabolism of a Synthetic Cannabinoid in the Zebrafish Larvae Model. Molecules 2020, 25, 4474. [Google Scholar] [CrossRef]
- Caspar, A.T.; Gaab, J.B.; Michely, J.A.; Brandt, S.D.; Meyer, M.R.; Maurer, H.H. Metabolism of the Tryptamine-Derived New Psychoactive Substances 5-MeO-2-Me-DALT, 5-MeO-2-Me-ALCHT, and 5-MeO-2-Me-DIPT and Their Detectability in Urine Studied by GC-MS, LC-MS n, and LC-HR-MS/MS: Metabolism of 5-MeO-2-Me-DALT, 5-MeO-2-Me-ALCHT, and 5-MeO-2-Me-DIPT. Drug Test. Anal. 2018, 10, 184–195. [Google Scholar] [CrossRef] [Green Version]
- Wink, C.S.D.; Michely, J.A.; Jacobsen-Bauer, A.; Zapp, J.; Maurer, H.H. Diphenidine, a New Psychoactive Substance: Metabolic Fate Elucidated with Rat Urine and Human Liver Preparations and Detectability in Urine Using GC-MS, LC-MS n, and LC-HR-MS n: Metabolism of Diphenidine. Drug Test. Anal. 2016, 8, 1005–1014. [Google Scholar] [CrossRef]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and Community Curation of Mass Spectrometry Data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Allard, S.; Allard, P.-M.; Morel, I.; Gicquel, T. Application of a Molecular Networking Approach for Clinical and Forensic Toxicology Exemplified in Three Cases Involving 3-MeO-PCP, Doxylamine, and Chlormequat. Drug Test. Anal. 2018, 11, 669–677. [Google Scholar] [CrossRef]
- Le Daré, B.; Allard, S.; Bouvet, R.; Baert, A.; Allard, P.-M.; Morel, I.; Gicquel, T. A Case of Fatal Acebutolol Poisoning: An Illustration of the Potential of Molecular Networking. Int. J. Leg. Med. 2019, 134, 251–256. [Google Scholar] [CrossRef]
- Le Daré, B.; Ferron, P.-J.; Couette, A.; Ribault, C.; Morel, I.; Gicquel, T. In Vivo and in Vitro α-Amanitin Metabolism Studies Using Molecular Networking. Toxicol. Lett. 2021, 346, 1–6. [Google Scholar] [CrossRef]
- Ferron, P.-J.; Le Daré, B.; Bronsard, J.; Steichen, C.; Babina, E.; Pelletier, R.; Hauet, T.; Morel, I.; Tarte, K.; Reizine, F.; et al. Molecular Networking for Drug Toxicities Studies: The Case of Hydroxychloroquine in COVID-19 Patients. IJMS 2021, 23, 82. [Google Scholar] [CrossRef]
- Magny, R.; Auzeil, N.; Lefrère, B.; Mégarbane, B.; Houzé, P.; Labat, L. Molecular Network-Based Identification of Tramadol Metabolites in a Fatal Tramadol Poisoning. Metabolites 2022, 12, 665. [Google Scholar] [CrossRef]
- Allard, S.; Le Daré, B.; Allard, P.-M.; Morel, I.; Gicquel, T. Comparative Molecular Networking Analysis of a Rauwolfia Plant Powder and Biological Matrices in a Fatal Ingestion Case. Forensic Toxicol. 2020, 38, 447–454. [Google Scholar] [CrossRef]
- Kirchmair, J.; Göller, A.H.; Lang, D.; Kunze, J.; Testa, B.; Wilson, I.D.; Glen, R.C.; Schneider, G. Predicting Drug Metabolism: Experiment and/or Computation? Nat. Rev. Drug Discov. 2015, 14, 387–404. [Google Scholar] [CrossRef] [Green Version]
- Gertrudes, J.C.; Maltarollo, V.G.; Silva, R.A.; Oliveira, P.R.; Honorio, K.M.; da Silva, A.B.F. Machine Learning Techniques and Drug Design. CMC 2012, 19, 4289–4297. [Google Scholar] [CrossRef]
- Du, Q.-S.; Huang, R.-B.; Chou, K.-C. Recent Advances in QSAR and Their Applications in Predicting the Activities of Chemical Molecules, Peptides and Proteins for Drug Design. CPPS 2008, 9, 248–259. [Google Scholar] [CrossRef]
- Kazmi, S.R.; Jun, R.; Yu, M.-S.; Jung, C.; Na, D. In Silico Approaches and Tools for the Prediction of Drug Metabolism and Fate: A Review. Comput. Biol. Med. 2019, 106, 54–64. [Google Scholar] [CrossRef]
- Tyzack, J.D.; Kirchmair, J. Computational Methods and Tools to Predict Cytochrome P450 Metabolism for Drug Discovery. Chem. Biol. Drug Des. 2019, 93, 377–386. [Google Scholar] [CrossRef]
- Di Trana, A.; Brunetti, P.; Giorgetti, R.; Marinelli, E.; Zaami, S.; Busardò, F.P.; Carlier, J. In Silico Prediction, LC-HRMS/MS Analysis, and Targeted/Untargeted Data-Mining Workflow for the Profiling of Phenylfentanyl in Vitro Metabolites. Talanta 2021, 235, 122740. [Google Scholar] [CrossRef]
- de Bruyn Kops, C.; Šícho, M.; Mazzolari, A.; Kirchmair, J. GLORYx: Prediction of the Metabolites Resulting from Phase 1 and Phase 2 Biotransformations of Xenobiotics. Chem. Res. Toxicol. 2021, 34, 286–299. [Google Scholar] [CrossRef]
- Djoumbou-Feunang, Y.; Fiamoncini, J.; Gil-de-la-Fuente, A.; Greiner, R.; Manach, C.; Wishart, D.S. BioTransformer: A Comprehensive Computational Tool for Small Molecule Metabolism Prediction and Metabolite Identification. J. Cheminform. 2019, 11, 2. [Google Scholar] [CrossRef] [Green Version]
- Gicquel, T.; Pelletier, R.; Richeval, C.; Gish, A.; Hakim, F.; Ferron, P.; Mesli, V.; Allorge, D.; Morel, I.; Gaulier, J. Metabolite Elucidation of 2-fluoro-deschloroketamine (2F-DCK) Using Molecular Networking across Three Complementary in Vitro and in Vivo Models. Drug Test. Anal. 2021, 14, 144–153. [Google Scholar] [CrossRef]
- Kessner, D.; Chambers, M.; Burke, R.; Agus, D.; Mallick, P. ProteoWizard: Open Source Software for Rapid Proteomics Tools Development. Bioinformatics 2008, 24, 2534–2536. [Google Scholar] [CrossRef] [Green Version]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular Framework for Processing, Visualizing, and Analyzing Mass Spectrometry-Based Molecular Profile Data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [Green Version]
- Shannon, P. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- HighChem LLC MzCloud-Advanced Mass Spectral Database. Available online: https://www.mzcloud.org/ (accessed on 12 November 2018).
- Bogaards, J.J.P.; Bertrand, M.; Jackson, P.; Oudshoorn, M.J.; Weaver, R.J.; Van Bladeren, P.J.; Walther, B. Determining the Best Animal Model for Human Cytochrome P450 Activities: A Comparison of Mouse, Rat, Rabbit, Dog, Micropig, Monkey and Man. Xenobiotica 2000, 30, 1131–1152. [Google Scholar] [CrossRef]
- Guengerich, F.P. Comparisons of Catalytic Selectivity of Cytochrome P450 Subfamily Enzymes from Different Species. Chem. Biol. Interact. 1997, 106, 161–182. [Google Scholar] [CrossRef]
- Lin, J.H. Applications and Limitations of Interspecies Scaling and in Vitro Extrapolation in Pharmacokinetics. Drug Metab. Dispos. 1998, 26, 1202–1212. [Google Scholar]
- Lin, J.H. Species Similarities and Differences in Pharmacokinetics. Drug Metab. Dispos. 1995, 23, 1008–1021. [Google Scholar]
- Jeon, J.-H.; Lee, J.; Choi, M.-K.; Song, I.-S. Pharmacokinetics of Ginsenosides Following Repeated Oral Administration of Red Ginseng Extract Significantly Differ between Species of Experimental Animals. Arch. Pharm. Res. 2020, 43, 1335–1346. [Google Scholar] [CrossRef]
- Natsch, A.; Nordone, A.; Adamson, G.M.; Laue, H. A Species Specific Metabolism Leading to Male Rat Reprotoxicity of Cyclamen Aldehyde: In Vivo and in Vitro Evaluation. Food Chem. Toxicol. 2021, 153, 112243. [Google Scholar] [CrossRef] [PubMed]
- Arensdorff, L.; Boillat-Blanco, N.; Decosterd, L.; Buclin, T.; de Vallière, S. Adequate Plasma Drug Concentrations Suggest That Amoxicillin Can Be Administered by Continuous Infusion Using Elastomeric Pumps. J. Antimicrob. Chemother. 2017, 72, 2613–2615. [Google Scholar] [CrossRef] [PubMed]
- Powell, J.S. Liposomal Approach towards the Development of a Longer-Acting Factor VIII. Haemophilia 2007, 13, 23–28. [Google Scholar] [CrossRef]
- Pacchiarotti, I.; Tiihonen, J.; Kotzalidis, G.D.; Verdolini, N.; Murru, A.; Goikolea, J.M.; Valentí, M.; Aedo, A.; Vieta, E. Long-Acting Injectable Antipsychotics (LAIs) for Maintenance Treatment of Bipolar and Schizoaffective Disorders: A Systematic Review. Eur. Neuropsychopharmacol. 2019, 29, 457–470. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, S.; Isaacs, D.; Clements, J.N. Pharmacokinetics and Clinical Implications of Semaglutide: A New Glucagon-Like Peptide (GLP)-1 Receptor Agonist. Clin. Pharmacokinet. 2018, 57, 1529–1538. [Google Scholar] [CrossRef] [PubMed]
- Bialer, M.; Friedman, M.; Dubrovsky, J. Relation between Absorption Half-Life Values of Four Novel Sustained-Release Dosage Forms of Valproic Acid in Dogs and Human. Biopharm. Drug Dispos. 1986, 7, 495–500. [Google Scholar] [CrossRef]
- Gunaydin, H.; Altman, M.D.; Ellis, J.M.; Fuller, P.; Johnson, S.A.; Lahue, B.; Lapointe, B. Strategy for Extending Half-Life in Drug Design and Its Significance. ACS Med. Chem. Lett. 2018, 9, 528–533. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Predicted Metabolite (m/z) Elemental Composition Metabolite Reaction | Putative Biotransformation Reaction Exact Mass Shift | Molecular Structure | Prediction Software | Score and/or Enzyme Involved |
|---|---|---|---|---|
| Sibiriline (m/z 211.0871) C13H10N2O None | Not applicable |  | Not applicable | Not applicable |
| P1 (m/z 387.1192) C19H18N2O7 O-glucuronidation | +C6H8O6 +176.032 |  | GLORYx BT3 | 0.948 Predicted (UDP-glucuronosyltransferase) |
| P2 (m/z 227.0820) C13H10N2O2 Aromatic hydroxylation | +O +15.994 |  | GLORYx BT3 | 0.656 Predicted (CYP1A2) |
| P3 (m/z 291.0439) C13H10N2O4S Sulfation | +SO3 +79.956 |  | GLORYx BT3 | 0.628 Predicted (Aryl sulfotransferase) |
| P4 (m/z 227.0820) C13H10N2O2 Aromatic hydroxylation | +O +15.994 |  | GLORYx BT3 | 0.348 Not predicted |
| P5 (m/z 227.0820) C13H10N2O2 Aromatic hydroxylation (para to carbon) | +O +15.994 |  | GLORYx BT3 | 0.348 Not predicted |
| P6 (m/z 387.1192) C19H18N2O7 N-glucuronidation | +C6H8O6 +176.032 |  | GLORYx BT3 | 0.308 Not predicted |
| P7 (m/z 227.0820) C13H10N2O2 Aromatic hydroxylation (para to nitrogen) | +O +15.994 |  | GLORYx BT3 | 0.295 Predicted (CYP2C9) |
| P8 (m/z 225.1027) C14H12N2O Methylation (aromatic OH) | +CH2 +14.015 |  | GLORYx BT3 | 0.28 Not predicted |
| P9 (m/z 387.1192) C19H19N2O7+ N-glucuronidation | +C6H8O6 +176.032 |  | GLORYx BT3 | 0.224 Not predicted |
| P10 (m/z 227.0820) C13H10N2O2 N-oxidation | +O +15.994 |  | GLORYx BT3 | 0.212 Predicted (CYP2C8) |
| P11 (m/z 119.0609) C7H6N2 | Unknown |  | GLORYx BT3 | 0.1312 Not predicted |
| P12 (m/z 109.0289) C6H4O2 Oxidation of 4-substituted phenol to quinone | Unknown |  | GLORYx BT3 | 0.1312 Not predicted |
| P13 (m/z 227.0820) C13H10N2O2 Aromatic hydroxylation | +O +15.994 |  | GLORYx BT3 | 0.072 Predicted (CYP2C9) |
| P14 (m/z 227.0820) C13H10N2O2 Aromatic hydroxylation | +O +15.994 |  | GLORYx BT3 | 0.028 Predicted (CYP1A2) |
| P15 (m/z 227.0820) C13H10N2O2 Oxidation | +O +15.994 |  | GLORYx BT3 | None Predicted (CYP2E1) |
| P16 (m/z 403.1141) C19H18N2O8 Hydroxylation + O-glucuronidation | C6H8O7 +192.027 |  | GLORYx BT3 | None Predicted (UDP-glucuronosyltransferase) |
| P17 (m/z 403.1141) C19H18N2O8 Hydroxylation + O-glucuronidation | +C6H8O7 +192.027 |  | GLORYx BT3 | None Predicted (UDP-glucuronosyltransferase) |
| P18 (m/z 403.1141) C19H18N2O8 Hydroxylation + O-glucuronidation | +C6H8O7 +192.027 |  | GLORYx BT3 | None Predicted (UDP-glucuronosyltransferase) |
| P19 (m/z 403.1141) C19H18N2O8 Hydroxylation + O-glucuronidation | +C6H8O7 +192.027 |  | GLORYx BT3 | None Predicted (UDP-glucuronosyltransferase) |
| P20 (m/z 307.0388) C13H10N2O5S Hydroxylation + O-sulfation | +SO4 +95.951 |  | GLORYx BT3 | None Predicted (Aryl sulfotransferase) |
| P21 (m/z 241.0977) C14H12N2O2 O-methylation | +CH2 +14.015 |  | GLORYx BT3 | None Predicted (Catechol O-methyltransferase) |
| Metabolite Proposal | Formula Proposal | Biotransformation | m/z | Mass Shift | RT (min) | In Vivo | In Vitro | In Silico | |
|---|---|---|---|---|---|---|---|---|---|
| GLORYx | BT3 | ||||||||
| - | C6H4O2 | Unknown | 109.029 | −102.058 | - | - | - | X | - |
| - | C7H6N2 | Unknown | 119.061 | −92.026 | - | - | - | X | - |
| - | C13H10N2O | Parent molecule (sibiriline) | 211.087 | 0 | 5.8 | X | X | X | X |
| M1a | C13H10N2O2 | Hydroxylation | 227.082 | 15.995 | 5.0 | X | - | X | X |
| M1b | 5.2 | X | - | ||||||
| M2a | C13H10N2O4S | Sulfation | 291.044 | 79.957 | 4.4 | - | X | X | X |
| M2b | 5.2 | X | X | ||||||
| M3a | C13H10N2O5S | Hydroxylation + sulfation | 307.038 | 95.952 | 4.4 | - | X | - | X |
| M3b | 5.2 | X | X | ||||||
| M4 | C15H14N2O4S | 2 × methylation + sulfation | 319.075 | 107.988 | 5.1 | - | X | - | - |
| M5a | C19H18N2O7 | Glucuronidation | 387.119 | 176.033 | 3.7 | X | X | X | X |
| M5b | 4.8 | X | X | ||||||
| M6a | C19H18N2O8 | Hydroxylation + glucuronidation | 403.113 | 192.027 | 4.1 | X | X | - | X |
| M6b | 4.3 | X | X | ||||||
| M6c | 5.0 | X | |||||||
| M7 | C21H22N2O7 | 2 × methylation + glucuronidation | 415.150 | 204.063 | 4.3 | - | X | - | - |
| M8 | C25H26N2O15 | 2 × hydroxylation + 2 × glucuronidation | 595.183 | 384.096 | 5.9 | - | X | - | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pelletier, R.; Gicquel, T.; Simoes Eugenio, M.; Ferron, P.-J.; Morel, I.; Delehouzé, C.; Dimanche-Boitrel, M.-T.; Rousselot, M.; Le Daré, B. A Transversal Approach Combining In Silico, In Vitro and In Vivo Models to Describe the Metabolism of the Receptor Interacting Protein 1 Kinase Inhibitor Sibiriline. Pharmaceutics 2022, 14, 2665. https://doi.org/10.3390/pharmaceutics14122665
Pelletier R, Gicquel T, Simoes Eugenio M, Ferron P-J, Morel I, Delehouzé C, Dimanche-Boitrel M-T, Rousselot M, Le Daré B. A Transversal Approach Combining In Silico, In Vitro and In Vivo Models to Describe the Metabolism of the Receptor Interacting Protein 1 Kinase Inhibitor Sibiriline. Pharmaceutics. 2022; 14(12):2665. https://doi.org/10.3390/pharmaceutics14122665
Chicago/Turabian StylePelletier, Romain, Thomas Gicquel, Mélanie Simoes Eugenio, Pierre-Jean Ferron, Isabelle Morel, Claire Delehouzé, Marie-Thérèse Dimanche-Boitrel, Morgane Rousselot, and Brendan Le Daré. 2022. "A Transversal Approach Combining In Silico, In Vitro and In Vivo Models to Describe the Metabolism of the Receptor Interacting Protein 1 Kinase Inhibitor Sibiriline" Pharmaceutics 14, no. 12: 2665. https://doi.org/10.3390/pharmaceutics14122665