
Formulated Phospholipids as Non-Canonical TLR4 Agonists
 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Reagents
2.2. Animal Ethics
2.3. Bone Marrow-Derived Dendritic Cell Prime and Stimulation In Vitro
2.4. Cytokine Enzyme-Linked Immunosorbent Assays (ELISAs)
2.5. Reporter Cell Assays
2.6. Human Whole Blood Immunogenicity Assay
2.7. Mice and Immunizations
2.8. Serum Endpoint Titer ELISA
2.9. Statistical Analysis
3. Results
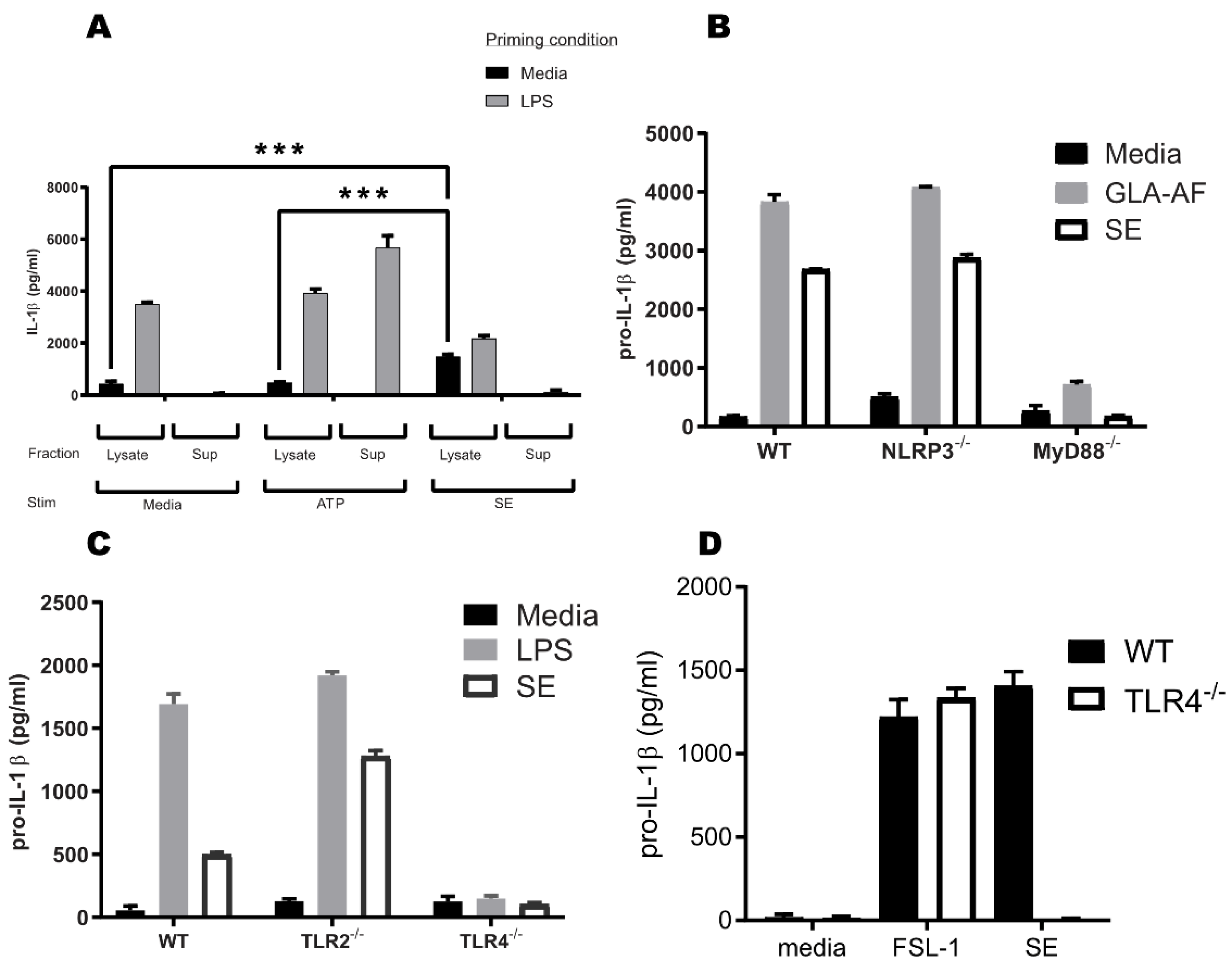
3.1. SE Does Not Induce Mature IL-1β Secretion through the Inflammasome
3.2. TLR4 and MyD88 Are Crucial for the Pro-IL-1β Induction Activity
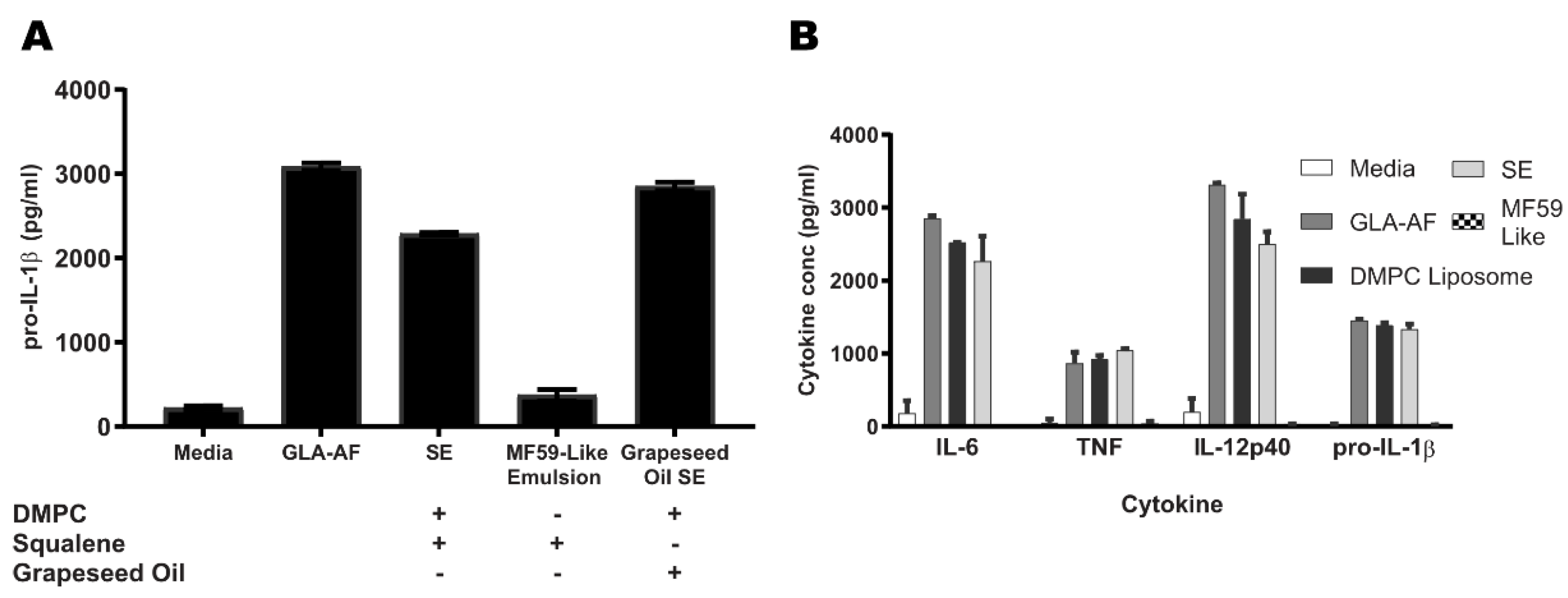
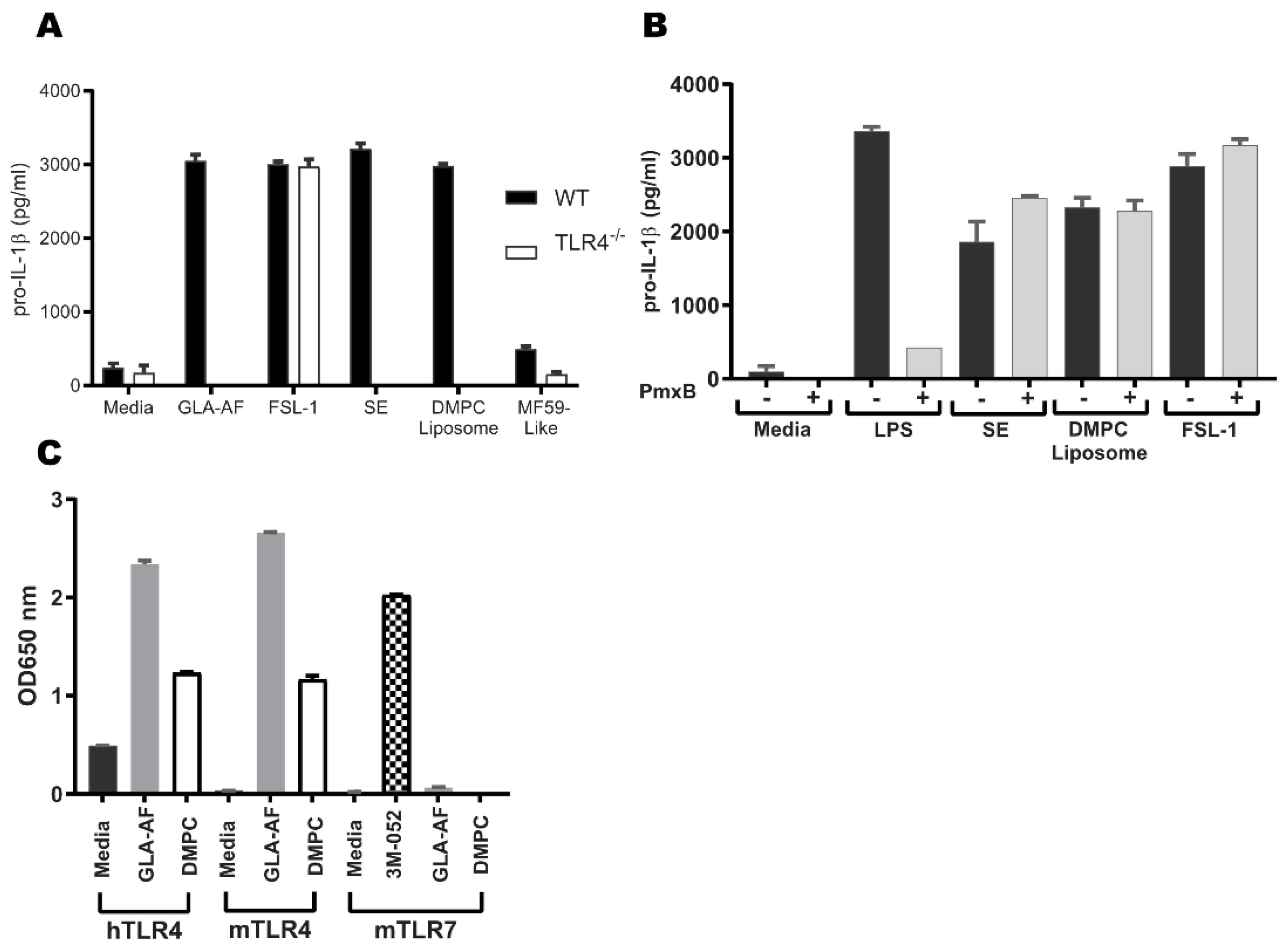
3.3. DMPC, a Component of SE, Is Essential for TLR4 Activity
3.4. DMPC Is an Agonist of the Murine and Human TLR4 Pathways
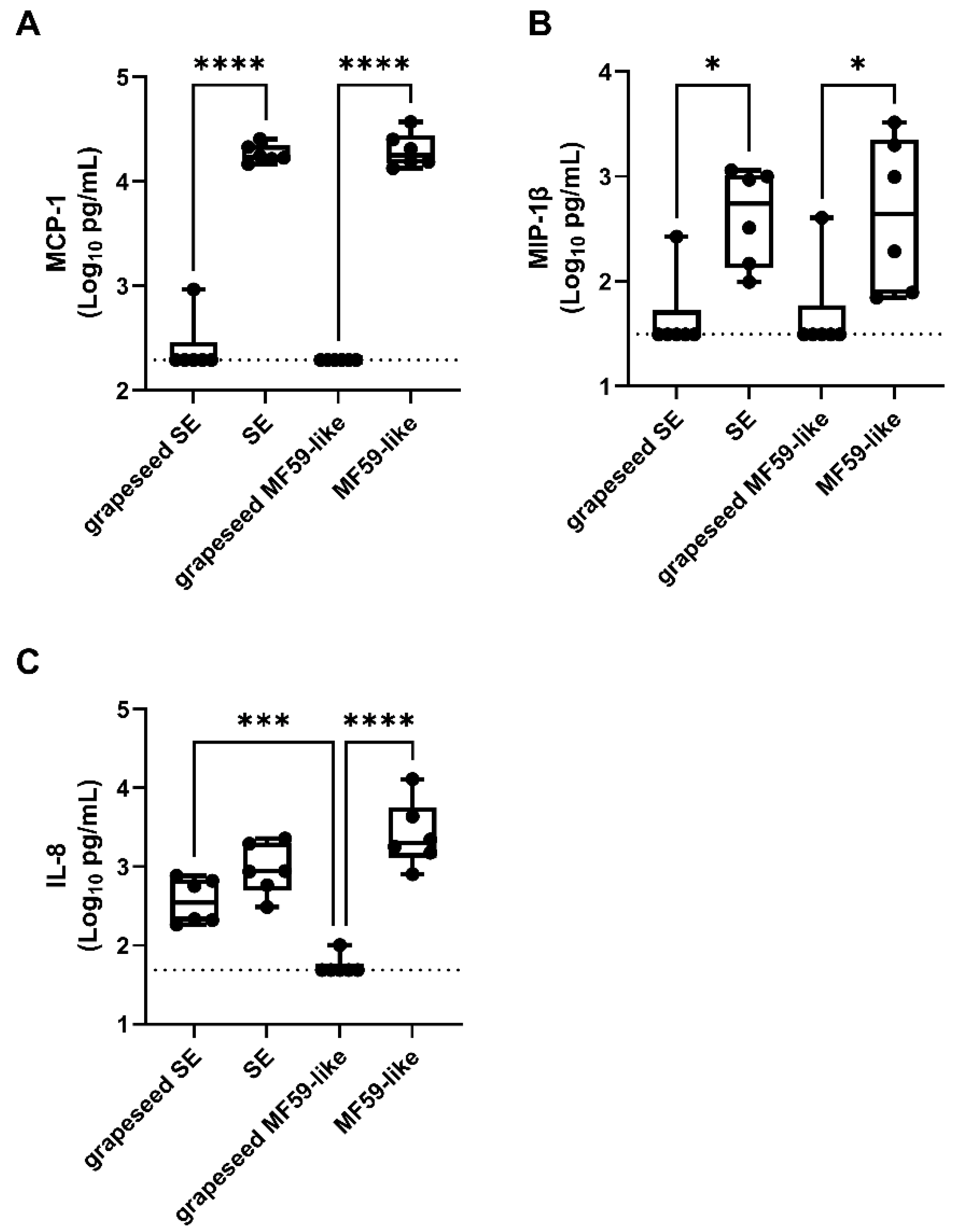
3.5. Emulsion Stimulation of Innate Immune Activity in Human Whole Blood Is Dominated by Oil-Phase Components
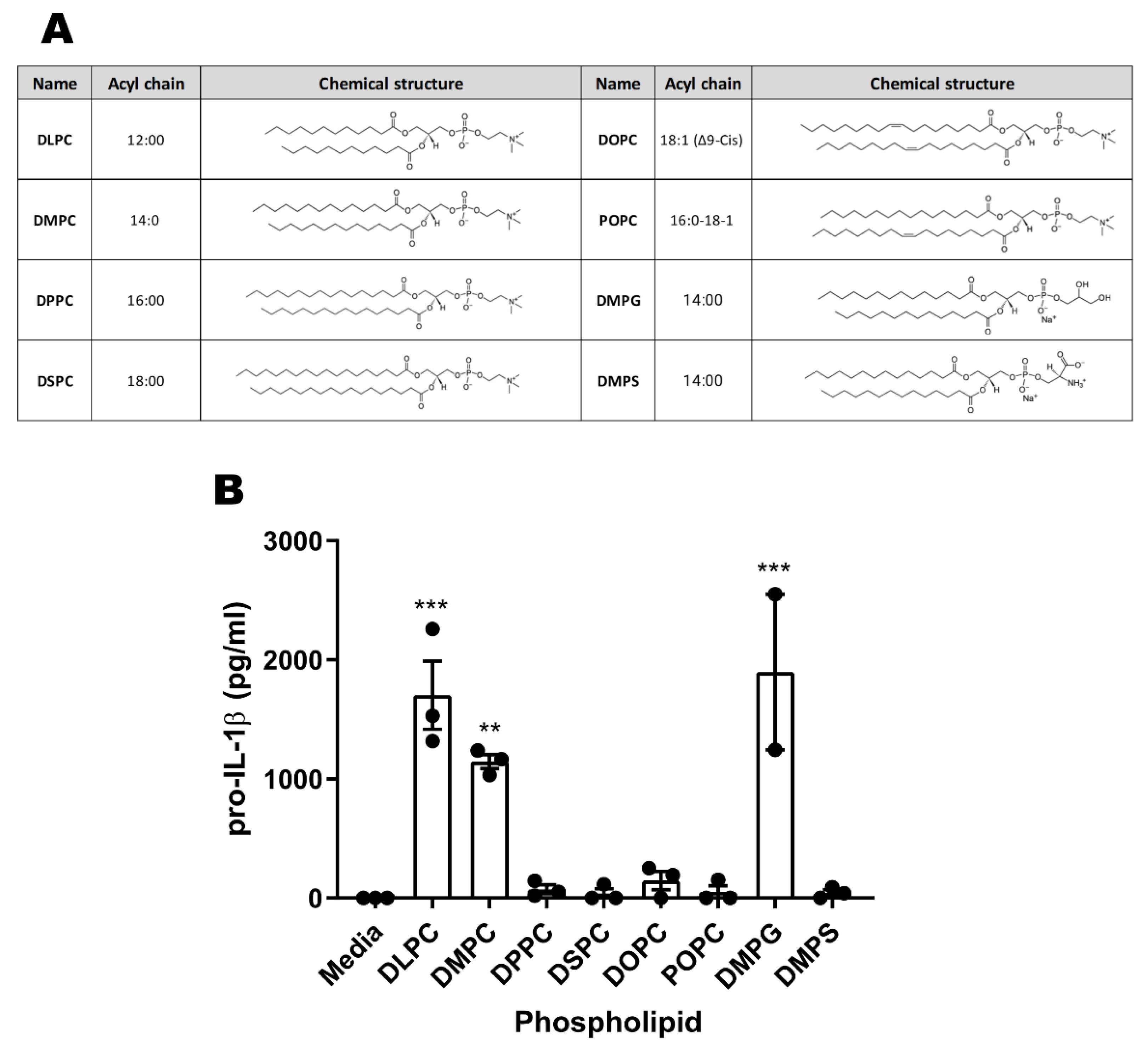
3.6. Phospholipid Acyl Chain Length, Saturation State, and Headgroup Structure Are Critical for TLR4 Activity
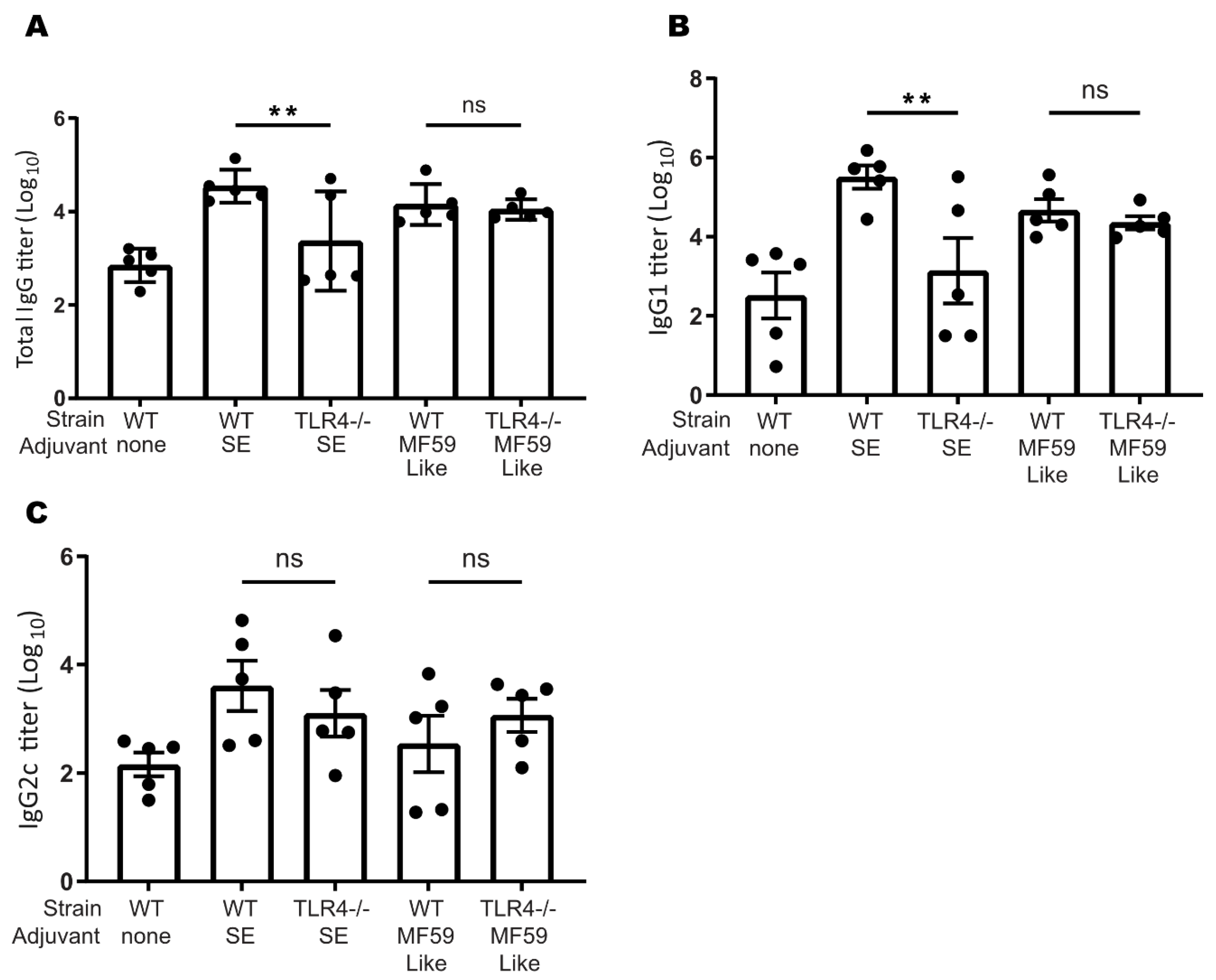
3.7. TLR4 Plays a Role in Murine SE-Stimulated Antibody Responses
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Champion, C.R. Heplisav-B: A Hepatitis B Vaccine with a Novel Adjuvant. Ann. Pharmacother. 2021, 55, 783–791. [Google Scholar] [CrossRef] [PubMed]
- Luchner, M.; Reinke, S.; Milicic, A. TLR Agonists as Vaccine Adjuvants Targeting Cancer and Infectious Diseases. Pharmaceutics 2021, 13, 142. [Google Scholar] [CrossRef] [PubMed]
- Keech, C.; Albert, G.; Cho, I.; Robertson, A.; Reed, P.; Neal, S.; Plested, J.S.; Zhu, M.; Cloney-Clark, S.; Zhou, H.; et al. Phase 1–2 Trial of a SARS-CoV-2 Recombinant Spike Protein Nanoparticle Vaccine. N. Engl. J. Med. 2020, 383, 2320–2332. [Google Scholar] [CrossRef] [PubMed]
- O’Hagan, D.T.; Lodaya, R.N.; Lofano, G. The Continued Advance of Vaccine Adjuvants—‘We Can Work It Out’. Semin. Immunol. 2020, 50, 101426. [Google Scholar] [CrossRef]
- Shi, S.; Zhu, H.; Xia, X.; Liang, Z.; Ma, X.; Sun, B. Vaccine Adjuvants: Understanding the Structure and Mechanism of Adjuvanticity. Vaccine 2019, 37, 3167–3178. [Google Scholar] [CrossRef]
- Tritto, E.; Mosca, F.; De Gregorio, E. Mechanism of Action of Licensed Vaccine Adjuvants. Vaccine 2009, 27, 3331–3334. [Google Scholar] [CrossRef]
- Heitmann, J.S.; Bilich, T.; Tandler, C.; Nelde, A.; Maringer, Y.; Marconato, M.; Reusch, J.; Jäger, S.; Denk, M.; Richter, M.; et al. A COVID-19 Peptide Vaccine for the Induction of SARS-CoV-2 T Cell Immunity. Nature 2022, 601, 617–622. [Google Scholar] [CrossRef]
- Schroder, K.; Irvine, K.M.; Taylor, M.S.; Bokil, N.J.; Le Cao, K.-A.; Masterman, K.-A.; Labzin, L.I.; Semple, C.A.; Kapetanovic, R.; Fairbairn, L.; et al. Conservation and Divergence in Toll-like Receptor 4-Regulated Gene Expression in Primary Human versus Mouse Macrophages. Proc. Natl. Acad. Sci. USA 2012, 109, E944–E953. [Google Scholar] [CrossRef] [Green Version]
- Qu, G.; Liu, S.; Zhang, S.; Wang, L.; Wang, X.; Sun, B.; Yin, N.; Gao, X.; Xia, T.; Chen, J.J.; et al. Graphene Oxide Induces Toll-like Receptor 4 (TLR4)-Dependent Necrosis in Macrophages. ACS Nano 2013, 7, 5732–5745. [Google Scholar] [CrossRef]
- Uto, T.; Akagi, T.; Yoshinaga, K.; Toyama, M.; Akashi, M.; Baba, M. The Induction of Innate and Adaptive Immunity by Biodegradable Poly(γ-Glutamic Acid) Nanoparticles via a TLR4 and MyD88 Signaling Pathway. Biomaterials 2011, 32, 5206–5212. [Google Scholar] [CrossRef]
- Mano, S.S.; Kanehira, K.; Taniguchi, A. Comparison of Cellular Uptake and Inflammatory Response via Toll-like Receptor 4 to Lipopolysaccharide and Titanium Dioxide Nanoparticles. Int. J. Mol. Sci. 2013, 14, 13154–13170. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McKeown-Longo, P.J.; Higgins, P.J. Integration of Canonical and Noncanonical Pathways in TLR4 Signaling: Complex Regulation of the Wound Repair Program. Adv. Wound Care 2017, 6, 320–329. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaffaroni, L.; Peri, F. Recent Advances on Toll-like Receptor 4 Modulation: New Therapeutic Perspectives. Future Med. Chem. 2018, 10, 461–476. [Google Scholar] [CrossRef] [Green Version]
- O’Hagan, D.T.; Fox, C.B. New Generation Adjuvants—From Empiricism to Rational Design. Vaccine 2015, 33, B14–B20. [Google Scholar] [CrossRef]
- Seydoux, E.; Liang, H.; Dubois Cauwelaert, N.; Archer, M.; Rintala, N.D.; Kramer, R.; Carter, D.; Fox, C.B.; Orr, M.T. Effective Combination Adjuvants Engage Both TLR and Inflammasome Pathways to Promote Potent Adaptive Immune Responses. J. Immunol. 2018, 201, 98–112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gombault, A.; Baron, L.; Couillin, I. ATP Release and Purinergic Signaling in NLRP3 Inflammasome Activation. Front. Immunol. 2012, 3, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Fox, C.B.; Carter, D.; Kramer, R.M.; Beckmann, A.M.; Reed, S.G. Current Status of Toll-Like Receptor 4 Ligand Vaccine Adjuvants. In Immunopotentiators in Modern Vaccines; Elsevier: Amsterdam, The Netherlands, 2017; pp. 105–127. ISBN 9780128040195. [Google Scholar]
- Duthie, M.S.; Windish, H.P.; Fox, C.B.; Reed, S.G. Use of Defined TLR Ligands as Adjuvants within Human Vaccines. Immunol. Rev. 2011, 239, 178–196. [Google Scholar] [CrossRef]
- Cauwelaert, N.D.; Desbien, A.L.; Hudson, T.E.; Pine, S.O.; Reed, S.G.; Coler, R.N.; Orr, M.T. The TLR4 Agonist Vaccine Adjuvant, GLA-SE, Requires Canonical and Atypical Mechanisms of Action for TH1 Induction. PLoS ONE 2016, 11, e0146372. [Google Scholar] [CrossRef]
- Misquith, A.; Fung, H.W.M.; Dowling, Q.M.; Guderian, J.A.; Vedvick, T.S.; Fox, C.B. In Vitro Evaluation of TLR4 Agonist Activity: Formulation Effects. Colloids Surf. B Biointerfaces 2014, 113, 312–319. [Google Scholar] [CrossRef] [Green Version]
- Orr, M.T.; Duthie, M.S.; Windish, H.P.; Lucas, E.A.; Guderian, J.A.; Hudson, T.E.; Shaverdian, N.; O’Donnell, J.; Desbien, A.L.; Reed, S.G.; et al. MyD88 and TRIF Synergistic Interaction Is Required for TH1-Cell Polarization with a Synthetic TLR4 Agonist Adjuvant. Eur. J. Immunol. 2013, 43, 2398–2408. [Google Scholar] [CrossRef]
- Desbien, A.L.; Dubois Cauwelaert, N.; Reed, S.J.; Bailor, H.R.; Liang, H.; Carter, D.; Duthie, M.S.; Fox, C.B.; Reed, S.G.; Orr, M.T. IL-18 and Subcapsular Lymph Node Macrophages Are Essential for Enhanced B Cell Responses with TLR4 Agonist Adjuvants. J. Immunol. 2016, 197, 4351–4359. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chu, L.H.; Indramohan, M.; Ratsimandresy, R.A.; Gangopadhyay, A.; Morris, E.P.; Monack, D.M.; Dorfleutner, A.; Stehlik, C. The Oxidized Phospholipid OxPAPC Protects from Septic Shock by Targeting the Non-Canonical Inflammasome in Macrophages. Nat. Commun. 2018, 9, 996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bochkov, V.N.; Kadl, A.; Huber, J.; Gruber, F.; Binder, B.R.; Leitinger, N. Protective Role of Phospholipid Oxidation Products in Endotoxin-Induced Tissue Damage. Nature 2002, 419, 77–81. [Google Scholar] [CrossRef] [PubMed]
- Di Gioia, M.; Spreafico, R.; Springstead, J.R.; Mendelson, M.M.; Joehanes, R.; Levy, D.; Zanoni, I. Endogenous Oxidized Phospholipids Reprogram Cellular Metabolism and Boost Hyperinflammation. Nat. Immunol. 2020, 21, 42–53. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.D.; Leitinger, N.; Navab, M.; Faull, K.F.; Hörkkö, S.; Witztum, J.L.; Palinski, W.; Schwenke, D.; Salomon, R.G.; Sha, W.; et al. Structural Identification by Mass Spectrometry of Oxidized Phospholipids in Minimally Oxidized Low Density Lipoprotein That Induce Monocyte/Endothelial Interactions and Evidence for Their Presence in Vivo. J. Biol. Chem. 1997, 272, 13597–13607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zanoni, I.; Tan, Y.; Di Gioia, M.; Springstead, J.R.; Kagan, J.C. By Capturing Inflammatory Lipids Released from Dying Cells, the Receptor CD14 Induces Inflammasome-Dependent Phagocyte Hyperactivation. Immunity 2017, 47, 697–709.e3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knapp, S.; Matt, U.; Leitinger, N.; van der Poll, T. Oxidized Phospholipids Inhibit Phagocytosis and Impair Outcome in Gram-Negative Sepsis In Vivo. J. Immunol. 2007, 178, 993–1001. [Google Scholar] [CrossRef] [Green Version]
- Hill, D.L.; Pierson, W.; Bolland, D.J.; Mkindi, C.; Carr, E.J.; Wang, J.; Houard, S.; Wingett, S.W.; Audran, R.; Wallin, E.F.; et al. The Adjuvant GLA-SE Promotes Human Tfh Cell Expansion and Emergence of Public TCRβ Clonotypes. J. Exp. Med. 2019, 216, 1857–1873. [Google Scholar] [CrossRef]
- Duthie, M.S.; Frevol, A.; Day, T.; Coler, R.N.; Vergara, J.; Rolf, T.; Sagawa, Z.K.; Marie Beckmann, A.; Casper, C.; Reed, S.G. A Phase 1 Antigen Dose Escalation Trial to Evaluate Safety, Tolerability and Immunogenicity of the Leprosy Vaccine Candidate LepVax (LEP-F1 + GLA–SE) in Healthy Adults. Vaccine 2020, 38, 1700–1707. [Google Scholar] [CrossRef]
- Orr, M.T.; Beebe, E.A.; Hudson, T.E.; Moon, J.J.; Fox, C.B.; Reed, S.G.; Coler, R.N. A Dual TLR Agonist Adjuvant Enhances the Immunogenicity and Protective Efficacy of the Tuberculosis Vaccine Antigen ID93. PLoS ONE 2014, 9, e83884. [Google Scholar] [CrossRef]
- Fox, C.B.; Barnes, V.L.; Evers, T.; Chesko, J.D.; Vedvick, T.S.; Coler, R.N.; Reed, S.G.; Baldwin, S.L. Adjuvanted Pandemic Influenza Vaccine: Variation of Emulsion Components Affects Stability, Antigen Structure, and Vaccine Efficacy. Influenza Other Respir. Viruses 2013, 7, 815–826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lutz, M.B.; Kukutsch, N.; Ogilvie, A.L.J.; Rößner, S.; Koch, F.; Romani, N.; Schuler, G. An Advanced Culture Method for Generating Large Quantities of Highly Pure Dendritic Cells from Mouse Bone Marrow. J. Immunol. Methods 1999, 223, 77–92. [Google Scholar] [CrossRef]
- Fox, C.B.; Van Hoeven, N.; Granger, B.; Lin, S.; Guderian, J.A.; Hartwig, A.; Marlenee, N.; Bowen, R.A.; Soultanov, V.; Carter, D. Vaccine Adjuvant Activity of Emulsified Oils from Species of the Pinaceae Family. Phytomedicine 2019, 64, 152927. [Google Scholar] [CrossRef] [PubMed]
- Orr, M.T.; Ireton, G.C.; Beebe, E.A.; Huang, P.-W.D.; Reese, V.A.; Argilla, D.; Coler, R.N.; Reed, S.G. Immune Subdominant Antigens as Vaccine Candidates against Mycobacterium Tuberculosis. J. Immunol. 2014, 193, 2911–2918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mariathasan, S.; Weiss, D.S.; Newton, K.; McBride, J.; O’Rourke, K.; Roose-Girma, M.; Lee, W.P.; Weinrauch, Y.; Monack, D.M.; Dixit, V.M. Cryopyrin Activates the Inflammasome in Response to Toxins and ATP. Nature 2006, 440, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saikh, K.U. MyD88 and beyond: A Perspective on MyD88-Targeted Therapeutic Approach for Modulation of Host Immunity. Immunol. Res. 2021, 69, 117–128. [Google Scholar] [CrossRef]
- Fox, C.B.; Baldwin, S.L.; Duthie, M.S.; Reed, S.G.; Vedvick, T.S. Immunomodulatory and Physical Effects of Oil Composition in Vaccine Adjuvant Emulsions. Vaccine 2011, 29, 9563–9572. [Google Scholar] [CrossRef] [Green Version]
- Danner, R.L.; Joiner, K.A.; Rubin, M.; Patterson, W.H.; Johnson, N.; Ayers, K.M.; Parrillo, J.E. Purification, Toxicity, and Antiendotoxin Activity of Polymyxin B Nonapeptide. Antimicrob. Agents Chemother. 1989, 33, 1428–1434. [Google Scholar] [CrossRef] [Green Version]
- Ohto, U.; Fukase, K.; Miyake, K.; Shimizu, T. Structural Basis of Species-Specific Endotoxin Sensing by Innate Immune Receptor TLR4/MD-2. Proc. Natl. Acad. Sci. USA 2012, 109, 7421–7426. [Google Scholar] [CrossRef] [Green Version]
- Vijayan, V.; Pradhan, P.; Braud, L.; Fuchs, H.R.; Gueler, F.; Motterlini, R.; Foresti, R.; Immenschuh, S. Human and Murine Macrophages Exhibit Differential Metabolic Responses to Lipopolysaccharide—A Divergent Role for Glycolysis. Redox Biol. 2019, 22, 101147. [Google Scholar] [CrossRef]
- Nahori, M.-A.; Fournié-Amazouz, E.; Que-Gewirth, N.S.; Balloy, V.; Chignard, M.; Raetz, C.R.H.; Saint Girons, I.; Werts, C. Differential TLR Recognition of Leptospiral Lipid A and Lipopolysaccharide in Murine and Human Cells. J. Immunol. 2005, 175, 6022–6031. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stewart, E.; Triccas, J.A.; Petrovsky, N. Adjuvant Strategies for More Effective Tuberculosis Vaccine Immunity. Microorganisms 2019, 7, 255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reed, S.G.; Scott, P. T-Cell and Cytokine Responses in Leishmaniasis. Curr. Opin. Immunol. 1993, 5, 524–531. [Google Scholar] [CrossRef]
- Akashi, S.; Nagai, Y.; Ogata, H.; Oikawa, M.; Fukase, K.; Kusumoto, S.; Kawasaki, K.; Nishijima, M.; Hayashi, S.; Kimoto, M.; et al. Human MD-2 Confers on Mouse Toll-like Receptor 4 Species-Specific Lipopolysaccharide Recognition. Int. Immunol. 2001, 13, 1595–1599. [Google Scholar] [CrossRef] [Green Version]
- Abhyankar, M.M.; Orr, M.T.; Lin, S.; Suraju, M.O.; Simpson, A.; Blust, M.; Pham, T.; Guderian, J.A.; Tomai, M.A.; Elvecrog, J.; et al. Adjuvant Composition and Delivery Route Shape Immune Response Quality and Protective Efficacy of a Recombinant Vaccine for Entamoeba Histolytica. Npj Vaccines 2018, 3, 22. [Google Scholar] [CrossRef] [Green Version]
- Adeagbo, B.A.; Akinlalu, A.O.; Phan, T.; Guderian, J.; Boukes, G.; Willenburg, E.; Fenner, C.; Bolaji, O.O.; Fox, C.B. Controlled Covalent Conjugation of a Tuberculosis Subunit Antigen (ID93) to Liposome Improved In Vitro Th1-Type Cytokine Recall Responses in Human Whole Blood. ACS Omega 2020, 5, 31306–31313. [Google Scholar] [CrossRef]
- He, W.; Qu, T.; Yu, Q.; Wang, Z.; Lv, H.; Zhang, J.; Zhao, X.; Wang, P. LPS Induces IL-8 Expression through TLR4, MyD88, NF-KappaB and MAPK Pathways in Human Dental Pulp Stem Cells. Int. Endod. J. 2013, 46, 128–136. [Google Scholar] [CrossRef]
- Marr, N.; Turvey, S.E. Role of Human TLR4 in Respiratory Syncytial Virus-Induced NF-ΚB Activation, Viral Entry and Replication. Innate Immun. 2012, 18, 856–865. [Google Scholar] [CrossRef]
- Lonez, C.; Irvine, K.L.; Pizzuto, M.; Schmidt, B.I.; Gay, N.J.; Ruysschaert, J.-M.; Gangloff, M.; Bryant, C.E. Critical Residues Involved in Toll-like Receptor 4 Activation by Cationic Lipid Nanocarriers Are Not Located at the Lipopolysaccharide-Binding Interface. Cell. Mol. Life Sci. 2015, 72, 3971–3982. [Google Scholar] [CrossRef] [Green Version]
- Wong-Baeza, C.; Tescucano, A.; Astudillo, H.; Reséndiz, A.; Landa, C.; España, L.; Serafín-López, J.; Estrada-García, I.; Estrada-Parra, S.; Flores-Romo, L.; et al. Nonbilayer Phospholipid Arrangements Are Toll-Like Receptor-2/6 and TLR-4 Agonists and Trigger Inflammation in a Mouse Model Resembling Human Lupus. J. Immunol. Res. 2015, 2015, 369462. [Google Scholar] [CrossRef]
- Lancaster, G.I.; Langley, K.G.; Berglund, N.A.; Kammoun, H.L.; Reibe, S.; Estevez, E.; Weir, J.; Mellett, N.A.; Pernes, G.; Conway, J.R.W.; et al. Evidence That TLR4 Is Not a Receptor for Saturated Fatty Acids but Mediates Lipid-Induced Inflammation by Reprogramming Macrophage Metabolism. Cell Metab. 2018, 27, 1096–1110.e5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisenbarth, S.C.; Colegio, O.R.; O’Connor, W.; Sutterwala, F.S.; Flavell, R.A. Crucial Role for the Nalp3 Inflammasome in the Immunostimulatory Properties of Aluminium Adjuvants. Nature 2008, 453, 1122–1126. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mestas, J.; Hughes, C.C.W. Of Mice and Not Men: Differences between Mouse and Human Immunology. J. Immunol. 2004, 172, 2731–2738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fox, C.B.; Baldwin, S.L.; Duthie, M.S.; Reed, S.G.; Vedvick, T.S. Immunomodulatory and Physical Effects of Phospholipid Composition in Vaccine Adjuvant Emulsions. AAPS PharmSciTech 2012, 13, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Calabro, S.; Tritto, E.; Pezzotti, A.; Taccone, M.; Muzzi, A.; Bertholet, S.; De Gregorio, E.; O’Hagan, D.T.; Baudner, B.; Seubert, A. The Adjuvant Effect of MF59 Is Due to the Oil-in-Water Emulsion Formulation, None of the Individual Components Induce a Comparable Adjuvant Effect. Vaccine 2013, 31, 3363–3369. [Google Scholar] [CrossRef]
- Chen, B.-M.; Cheng, T.-L.; Roffler, S.R. Polyethylene Glycol Immunogenicity: Theoretical, Clinical, and Practical Aspects of Anti-Polyethylene Glycol Antibodies. ACS Nano 2021, 15, 14022–14048. [Google Scholar] [CrossRef]
- Ahmad Ruzaidi, D.A.; Mahat, M.M.; Shafiee, S.A.; Mohamed Sofian, Z.; Mohmad Sabere, A.S.; Ramli, R.; Osman, H.; Hamzah, H.H.; Zainal Ariffin, Z.; Sadasivuni, K.K. Advocating Electrically Conductive Scaffolds with Low Immunogenicity for Biomedical Applications: A Review. Polymers 2021, 13, 3395. [Google Scholar] [CrossRef]
- Coler, R.N.; Bertholet, S.; Moutaftsi, M.; Guderian, J.A.; Windish, H.P.; Baldwin, S.L.; Laughlin, E.M.; Duthie, M.S.; Fox, C.B.; Carter, D.; et al. Development and Characterization of Synthetic Glucopyranosyl Lipid Adjuvant System as a Vaccine Adjuvant. PLoS ONE 2011, 6, e16333. [Google Scholar] [CrossRef] [Green Version]
- Seubert, A.; Calabro, S.; Santini, L.; Galli, B.; Genovese, A.; Valentini, S.; Aprea, S.; Colaprico, A.; D’Oro, U.; Giuliani, M.M.; et al. Adjuvanticity of the Oil-in-Water Emulsion MF59 Is Independent of Nlrp3 Inflammasome but Requires the Adaptor Protein MyD88. Proc. Natl. Acad. Sci. USA 2011, 108, 11169–11174. [Google Scholar] [CrossRef]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liang, H.; Lykins, W.R.; Seydoux, E.; Guderian, J.A.; Phan, T.; Fox, C.B.; Orr, M.T. Formulated Phospholipids as Non-Canonical TLR4 Agonists. Pharmaceutics 2022, 14, 2557. https://doi.org/10.3390/pharmaceutics14122557
Liang H, Lykins WR, Seydoux E, Guderian JA, Phan T, Fox CB, Orr MT. Formulated Phospholipids as Non-Canonical TLR4 Agonists. Pharmaceutics. 2022; 14(12):2557. https://doi.org/10.3390/pharmaceutics14122557
Chicago/Turabian StyleLiang, Hong, William R. Lykins, Emilie Seydoux, Jeffrey A. Guderian, Tony Phan, Christopher B. Fox, and Mark T. Orr. 2022. "Formulated Phospholipids as Non-Canonical TLR4 Agonists" Pharmaceutics 14, no. 12: 2557. https://doi.org/10.3390/pharmaceutics14122557