
Dendritic Cell-Derived Artificial Microvesicles Inhibit RLS40 Lymphosarcoma Growth in Mice via Stimulation of Th1/Th17 Immune Response
 , , , , , , and
, , , , , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Liposomes
2.2. Cells
2.3. Mice
2.4. Preparation of Vaccines
2.4.1. Dendritic Cell-Based Vaccines
2.4.2. Liposome-Based Vaccines
2.4.3. Cytochalasin B-Induced Membrane Vesicle (CIMV)-Based Vaccines
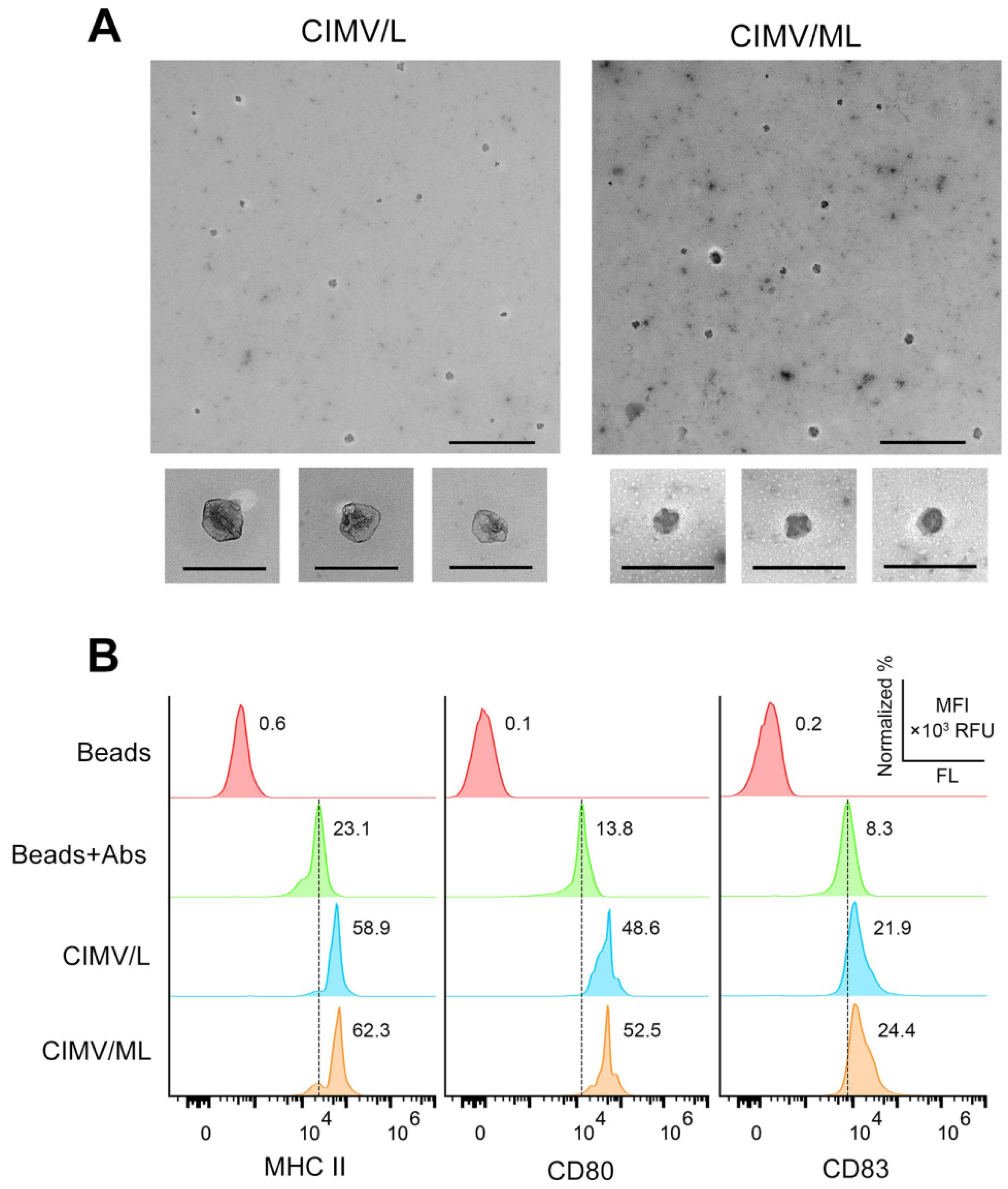
2.5. Transmission Electron Microscopy of CIMVs
2.6. Flow Cytometry Analysis of CIMVs
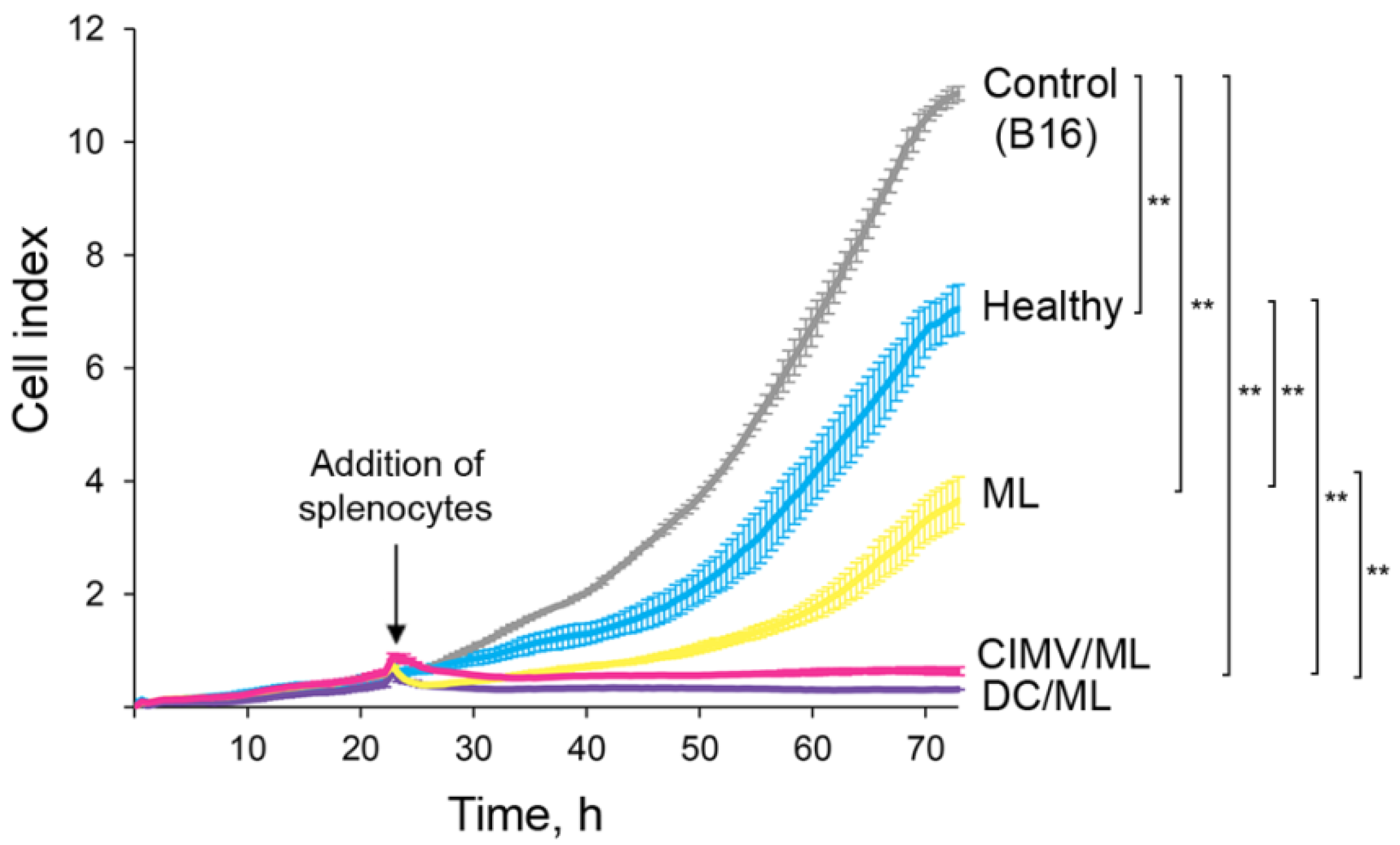
2.7. Evaluation of Antitumor CTL Response Activated by the Vaccines
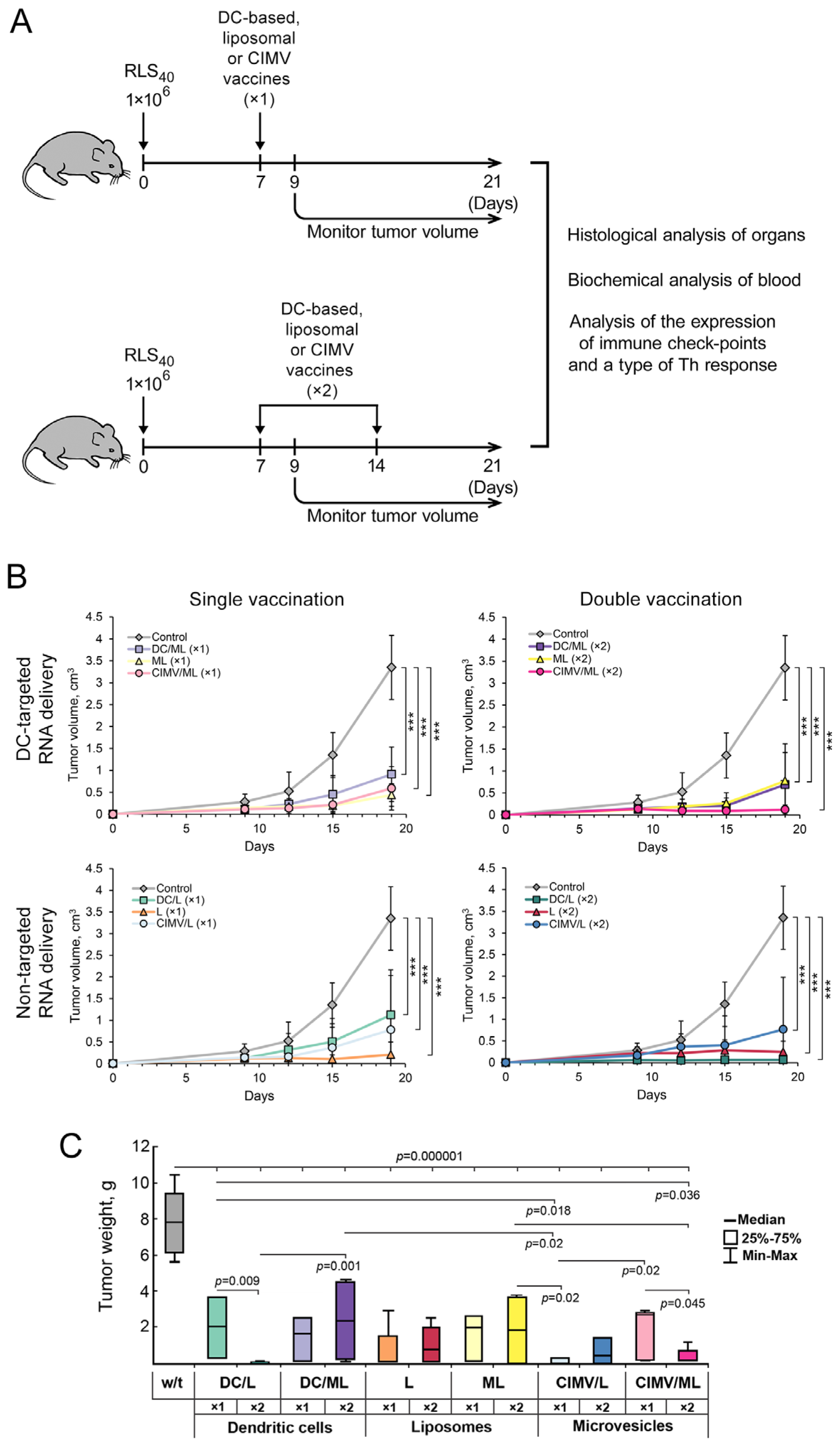
2.8. Tumor Transplantation and Design of Animal Experiments
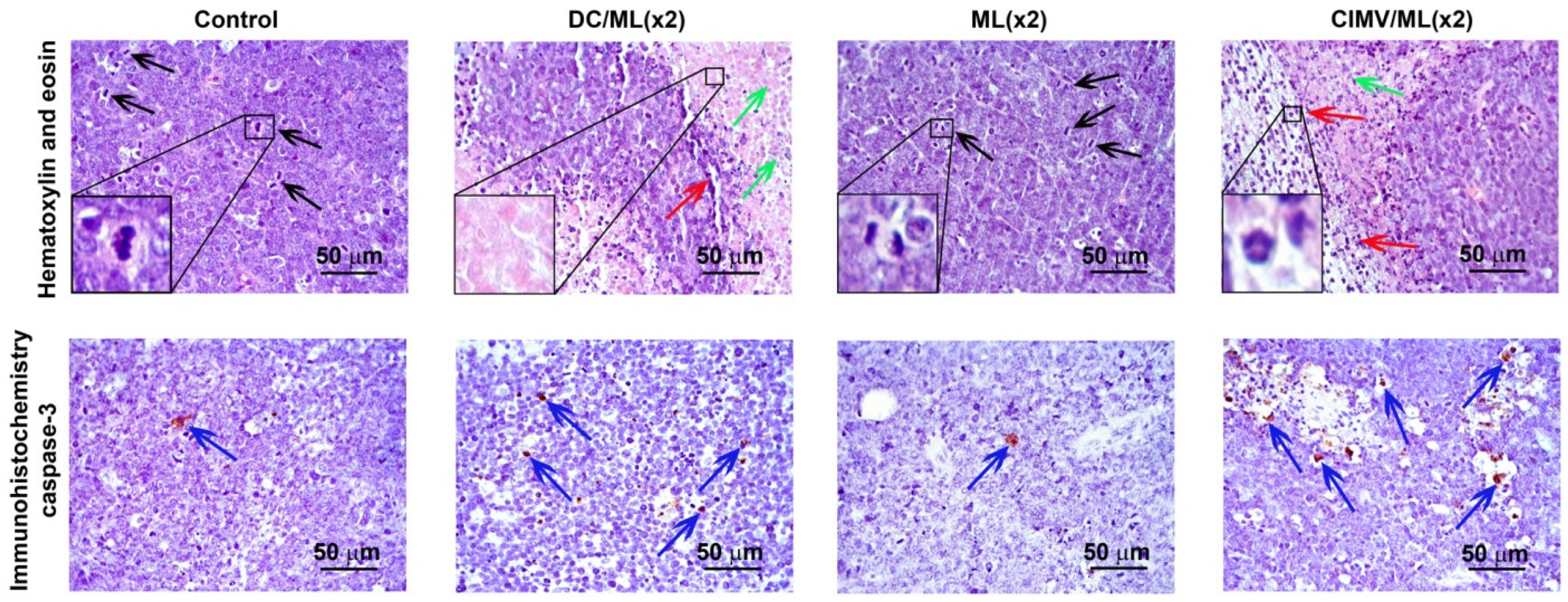
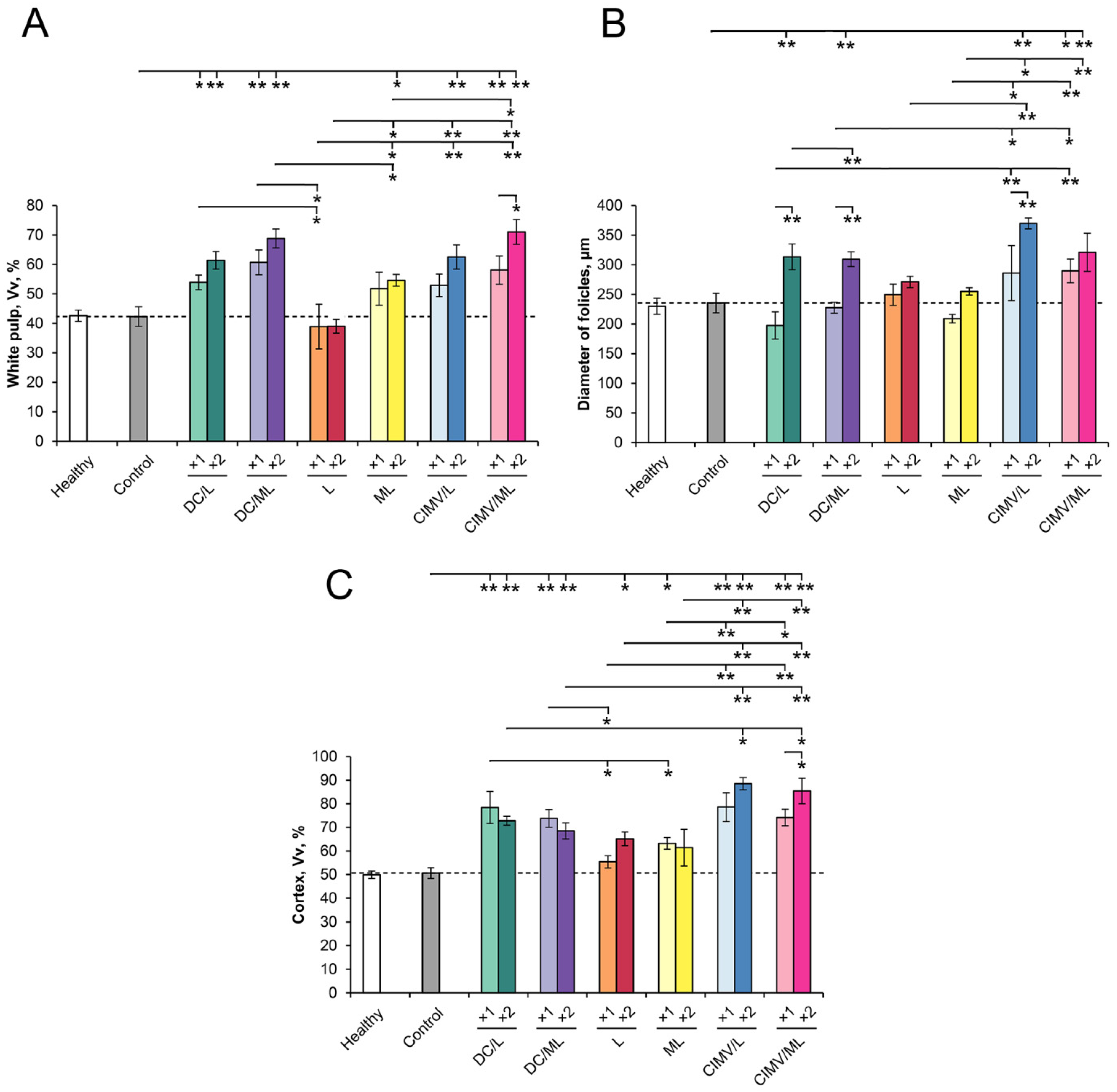
2.9. Histology and Immunohistochemistry
2.10. Blood Biochemistry
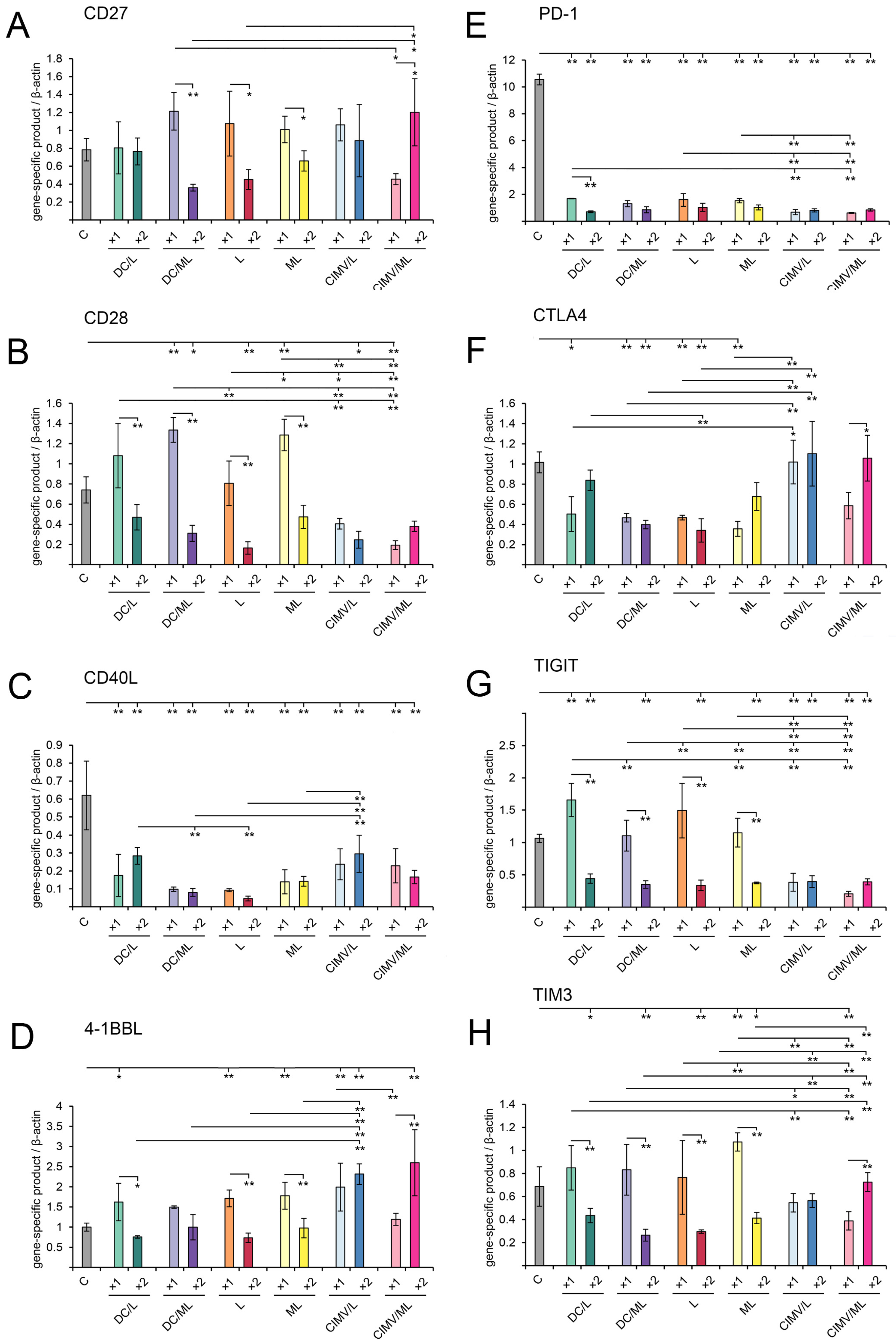
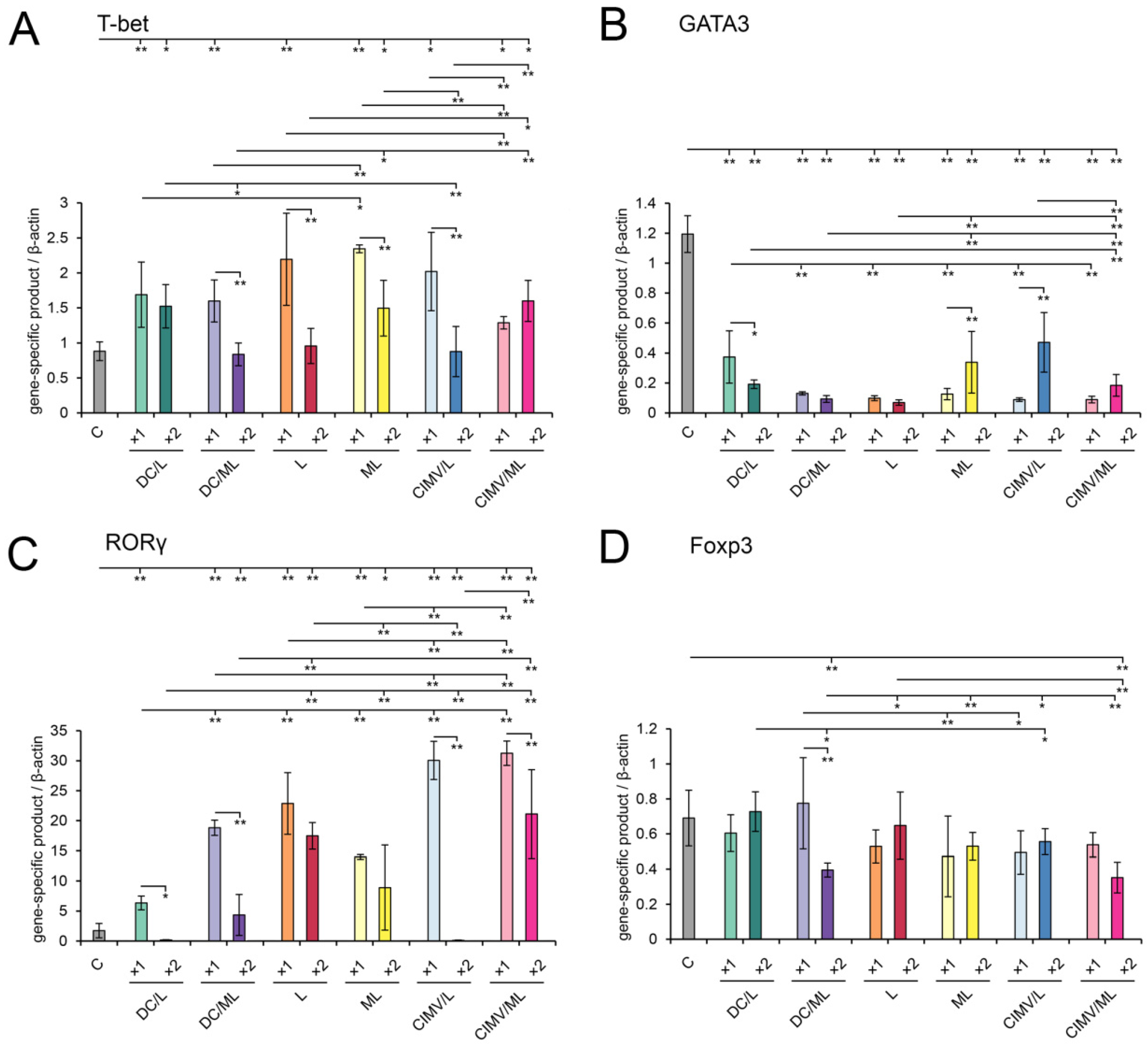
2.11. Immune Checkpoints and Master Regulators Analysis
2.12. Statistical Analysis
3. Results
3.1. Preparation and Characterization of Vaccines
3.2. DC-Derived CIMVs Efficiently Primed Antitumor Cytotoxic T-Cells In Vivo
3.3. Effect of Vaccination on RLS40 Lymphosarcoma Growth and Metastasis Development In Vivo
3.4. Liver Toxicity of Vaccines in RLS40-Bearing Mice
3.5. Immunomodulatory Effect of Different Types of Vaccines in RLS40-Bearing Mice
3.6. Immunologic Characteristics of Antitumor Immune Responses Caused by Vaccination
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stefanski, H.E.; Jonart, L.; Goren, E.; Mulé, J.J.; Blazar, B.R. A novel approach to improve immune effector responses post transplant by restoration of CCL21 expression. PLoS ONE 2018, 13, e0193461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Wan, J.; Hao, S.; Deng, X.; Chen, L.; Ma, L. TGF-β1-silenced leukemia cell-derived exosomes target dendritic cells to induce potent anti-leukemic immunity in a mouse model. Cancer Immunol. Immunother. 2017, 66, 1321–1331. [Google Scholar] [CrossRef] [PubMed]
- Nimanong, S.; Ostroumov, D.; Wingerath, J.; Knocke, S.; Woller, N.; Gürlevik, E.; Falk, C.S.; Manns, M.P.; Kühnel, F.; Wirth, T.C. CD40 signaling drives potent cellular immune responses in heterologous cancer vaccinations. Cancer Res. 2017, 77, 1918–1926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dammeijer, F.; Lievense, L.A.; Kaijen-Lambers, M.E.; Van Nimwegen, M.; Bezemer, K.; Hegmans, J.P.; Van Hall, T.; Hendriks, R.W.; Aerts, J.G. Depletion of tumor-associated macrophages with a CSF-1R kinase inhibitor enhances antitumor immunity and survival induced by DC immunotherapy. Cancer Immunol. Res. 2017, 5, 535–546. [Google Scholar] [CrossRef] [Green Version]
- Teng, C.F.; Wang, T.; Wu, T.H.; Lin, J.H.; Shih, F.Y.; Shyu, W.C.; Jeng, L. Bin Combination therapy with dendritic cell vaccine and programmed death ligand 1 immune checkpoint inhibitor for hepatocellular carcinoma in an orthotopic mouse model. Ther. Adv. Med. Oncol. 2020, 12, 1758835920922034. [Google Scholar] [CrossRef]
- Yao, Y.; Luo, F.; Tang, C.; Chen, D.; Qin, Z.; Hua, W.; Xu, M.; Zhong, P.; Yu, S.; Chen, D.; et al. Molecular subgroups and B7-H4 expression levels predict responses to dendritic cell vaccines in glioblastoma: An exploratory randomized phase II clinical trial. Cancer Immunol. Immunother. 2018, 67, 1777–1788. [Google Scholar] [CrossRef]
- Anguille, S.; Van De Velde, A.L.; Smits, E.L.; Van Tendeloo, V.F.; Juliusson, G.; Cools, N.; Nijs, G.; Stein, B.; Lion, E.; Van Driessche, A.; et al. Dendritic cell vaccination as postremission treatment to prevent or delay relapse in acute myeloid leukemia. Blood 2017, 130, 1713–1721. [Google Scholar] [CrossRef] [Green Version]
- Markov, O.V.; Mironova, N.L.; Vlassov, V.V.; Zenkova, M.A. Antitumor vaccines based on dendritic cells: From experiments using animal tumor models to clinical trials. Acta Nat. 2017, 9, 27–38. [Google Scholar] [CrossRef]
- Cunningham, S.; Hackstein, H. Recent Advances in Good Manufacturing Practice-Grade Generation of Dendritic Cells. Transfus. Med. Hemotherapy 2020, 47, 454–462. [Google Scholar] [CrossRef]
- Li, R.; Johnson, R.; Yu, G.; McKenna, D.H.; Hubel, A. Preservation of cell-based immunotherapies for clinical trials. Cytotherapy 2019, 21, 943–957. [Google Scholar] [CrossRef]
- Kowal, J.; Tkach, M. Dendritic cell extracellular vesicles. Int. Rev. Cell Mol. Biol. 2019, 349, 213–249. [Google Scholar] [CrossRef] [PubMed]
- Pitt, J.M.; André, F.; Amigorena, S.; Soria, J.C.; Eggermont, A.; Kroemer, G.; Zitvogel, L. Dendritic cell-derived exosomes for cancer therapy. J. Clin. Investig. 2016, 126, 1224–1232. [Google Scholar] [CrossRef] [PubMed]
- Nikfarjam, S.; Rezaie, J.; Kashanchi, F.; Jafari, R. Dexosomes as a cell-free vaccine for cancer immunotherapy. J. Exp. Clin. Cancer Res. 2020, 39, 258. [Google Scholar] [CrossRef] [PubMed]
- Markov, O.; Oshchepkova, A.; Mironova, N. Immunotherapy based on dendritic cell-targeted/-derived extracellular vesicles—A novel strategy for enhancement of the anti-tumor immune response. Front. Pharmacol. 2019, 10, 1152. [Google Scholar] [CrossRef] [Green Version]
- Morse, M.A.; Garst, J.; Osada, T.; Khan, S.; Hobeika, A.; Clay, T.M.; Valente, N.; Shreeniwas, R.; Sutton, M.A.; Delcayre, A.; et al. A phase I study of dexosome immunotherapy in patients with advanced non-small cell lung cancer. J. Transl. Med. 2005, 3, 9. [Google Scholar] [CrossRef] [Green Version]
- Escudier, B.; Dorval, T.; Chaput, N.; André, F.; Caby, M.P.; Novault, S.; Flament, C.; Leboulaire, C.; Borg, C.; Amigorena, S.; et al. Vaccination of metastatic melanoma patients with autologous dendritic cell (DC) derived-exosomes: Results of the first phase 1 clinical trial. J. Transl. Med. 2005, 3, 10. [Google Scholar] [CrossRef] [Green Version]
- Besse, B.; Charrier, M.; Lapierre, V.; Dansin, E.; Lantz, O.; Planchard, D.; Le Chevalier, T.; Livartoski, A.; Barlesi, F.; Laplanche, A.; et al. Dendritic cell-derived exosomes as maintenance immunotherapy after first line chemotherapy in NSCLC. Oncoimmunology 2016, 5, e1071008. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Kaslan, M.; Lee, S.H.; Yao, J.; Gao, Z. Progress in exosome isolation techniques. Theranostics 2017, 7, 789–804. [Google Scholar] [CrossRef]
- Oshchepkova, A.; Neumestova, A.; Matveeva, V.; Artemyeva, L.; Morozova, K.; Kiseleva, E.; Zenkova, M.; Vlassov, V. Cytochalasin-B-inducible nanovesicle mimics of natural extracellular vesicles that are capable of nucleic acid transfer. Micromachines 2019, 10, 750. [Google Scholar] [CrossRef] [Green Version]
- Wahlund, C.J.E.; Güclüler, G.; Hiltbrunner, S.; Veerman, R.E.; Näslund, T.I.; Gabrielsson, S. Exosomes from antigen-pulsed dendritic cells induce stronger antigen-specific immune responses than microvesicles in vivo. Sci. Rep. 2017, 7, 17095. [Google Scholar] [CrossRef]
- Gehrmann, U.; Hiltbrunner, S.; Georgoudaki, A.M.; Karlsson, M.C.; Näslund, T.I.; Gabrielsson, S. Synergistic induction of adaptive antitumor immunity by codelivery of antigen with α-galactosylceramide on exosomes. Cancer Res. 2013, 73, 3865–3876. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damo, M.; Wilson, D.S.; Simeoni, E.; Hubbell, J.A. TLR-3 stimulation improves anti-tumor immunity elicited by dendritic cell exosome-based vaccines in a murine model of melanoma. Sci. Rep. 2015, 5, 17622. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Theodoropoulos, P.A.; Gravanis, A.; Tsapara, A.; Margioris, A.N.; Papadogiorgaki, E.; Galanopoulos, V.; Stournaras, C. Cytochalasin B may shorten actin filaments by a mechanism independent of barbed end capping. Biochem. Pharmacol. 1994, 47, 1875–1881. [Google Scholar] [CrossRef]
- Gomzikova, M.O.; Aimaletdinov, A.M.; Bondar, O.V.; Starostina, I.G.; Gorshkova, N.V.; Neustroeva, O.A.; Kletukhina, S.K.; Kurbangaleeva, S.V.; Vorobev, V.V.; Garanina, E.E.; et al. Immunosuppressive properties of cytochalasin B-induced membrane vesicles of mesenchymal stem cells: Comparing with extracellular vesicles derived from mesenchymal stem cells. Sci. Rep. 2020, 10, 10740. [Google Scholar] [CrossRef] [PubMed]
- Chulpanova, D.S.; Gilazieva, Z.E.; Kletukhina, S.K.; Aimaletdinov, A.M.; Garanina, E.E.; James, V.; Rizvanov, A.A.; Solovyeva, V.V. Cytochalasin B-Induced membrane vesicles from human mesenchymal stem cells overexpressing IL2 are able to stimulate CD8+ T-Killers to kill human triple negative breast cancer cells. Biology 2021, 10, 141. [Google Scholar] [CrossRef]
- Gomzikova, M.O.; Kletukhina, S.K.; Kurbangaleeva, S.V.; Neustroeva, O.A.; Vasileva, O.S.; Garanina, E.E.; Khaiboullina, S.F.; Rizvanov, A.A. Mesenchymal stem cell derived biocompatible membrane vesicles demonstrate immunomodulatory activity inhibiting activation and proliferation of human mononuclear cells. Pharmaceutics 2020, 12, 577. [Google Scholar] [CrossRef]
- Gomzikova, M.; Kletukhina, S.; Kurbangaleeva, S.; Rizvanov, A. Evaluation of Cytochalasin B-Induced Membrane Vesicles Fusion Specificity with Target Cells. Biomed Res. Int. 2018, 2018, 7053623. [Google Scholar] [CrossRef] [Green Version]
- Gomzikova, M.O.; Zhuravleva, M.N.; Miftakhova, R.R.; Arkhipova, S.S.; Evtugin, V.G.; Khaiboullina, S.F.; Kiyasov, A.P.; Persson, J.L.; Mongan, N.P.; Pestell, R.G.; et al. Cytochalasin B-induced membrane vesicles convey angiogenic activity of parental cells. Oncotarget 2017, 8, 70496–70507. [Google Scholar] [CrossRef] [Green Version]
- Zitvogel, L.; Regnault, A.; Lozier, A.; Wolfers, J.; Flament, C.; Tenza, D.; Ricciardi-Castagnoli, P.; Raposo, G.; Amigorena, S. Eradication of established murine tumors using a novel cell-free vaccine: Dendritic cell-derived exosomes. Nat. Med. 1998, 4, 594–600. [Google Scholar] [CrossRef]
- Admyre, C.; Johansson, S.M.; Paulie, S.; Gabrielsson, S. Direct exosome stimulation of peripheral human T cells detected by ELISPOT. Eur. J. Immunol. 2006, 36, 1772–1781. [Google Scholar] [CrossRef]
- Mironova, N.; Shklyaeva, O.; Andreeva, E.; Popova, N.; Kaledin, V.; Nikolin, V.; Vlassov, V.; Zenkova, M. Animal model of drug-resistant tumor progression. Ann. N. Y. Acad. Sci. 2006, 1091, 490–500. [Google Scholar] [CrossRef] [PubMed]
- Markov, O.O.; Mironova, N.L.; Maslov, M.A.; Petukhov, I.A.; Morozova, N.G.; Vlassov, V.V.; Zenkova, M.A. Novel cationic liposomes provide highly efficient delivery of DNA and RNA into dendritic cell progenitors and their immature offsets. J. Control. Release 2012, 160, 200–210. [Google Scholar] [CrossRef] [PubMed]
- Markov, O.V.; Mironova, N.L.; Shmendel, E.V.; Serikov, R.N.; Morozova, N.G.; Maslov, M.A.; Vlassov, V.V.; Zenkova, M.A. Multicomponent mannose-containing liposomes efficiently deliver RNA in murine immature dendritic cells and provide productive anti-tumour response in murine melanoma model. J. Control. Release 2015, 213, 45–56. [Google Scholar] [CrossRef] [PubMed]
- Petukhov, I.A.; Maslov, M.A.; Morozova, N.G.; Serebrennikova, G.A. Synthesis of polycationic lipids based on cholesterol and spermine. Russ. Chem. Bull. 2010, 59, 260–268. [Google Scholar] [CrossRef]
- Shmendel’, E.V.; Timakova, A.A.; Maslov, M.A.; Morozova, N.G.; Chupin, V.V. Synthesis of mannose-containing neoglycolipids as a component of targeted delivery system for transfer of nucleic acids into antigen-presenting cells. Russ. Chem. Bull. 2012, 61, 1497–1501. [Google Scholar] [CrossRef]
- Markov, O.V.; Mironova, N.L.; Shmendel, E.V.; Maslov, M.A.; Zenkova, M.A. Systemic delivery of complexes of melanoma RNA with mannosylated liposomes activates highly efficient murine melanoma-specific cytotoxic T cells in vivo. Mol. Biol. 2017, 51, 102–107. [Google Scholar] [CrossRef]
- Oshchepkova, A.; Markov, O.; Evtushenko, E.; Chernonosov, A.; Kiseleva, E.; Morozova, K.; Matveeva, V.; Artemyeva, L.; Vlassov, V.; Zenkova, M. Tropism of extracellular vesicles and cell-derived nanovesicles to normal and cancer cells: New perspectives in tumor-targeted nucleic acid delivery. Pharmaceutics 2021, 13, 1911. [Google Scholar] [CrossRef]
- Dias, M.V.S.; Martins, V.R.; Hajj, G.N.M. Stress-Inducible Protein 1 (STI1): Extracellular Vesicle Analysis and Quantification; Humana Press: New York, NY, USA, 2016; Volume 1459. [Google Scholar]
- Markov, O.V.; Mironova, N.L.; Sennikov, S.V.; Vlassov, V.V.; Zenkova, M.A. Prophylactic dendritic cell-based vaccines efficiently inhibit metastases in murine metastatic melanoma. PLoS ONE 2015, 10, e0136911. [Google Scholar] [CrossRef] [Green Version]
- Huck, S.P.; Tang, S.C.; Andrew, K.A.; Yang, J.; Harper, J.L.; Ronchese, F. Activation and route of administration both determine the ability of bone marrow-derived dendritic cells to accumulate in secondary lymphoid organs and prime CD8+ T cells against tumors. Cancer Immunol. Immunother. 2008, 57, 63–71. [Google Scholar] [CrossRef]
- Baek, S.; Lee, S.J.; Kim, M.J.; Lee, H. Dendritic Cell (DC) Vaccine in Mouse Lung Cancer Minimal Residual Model; Comparison of Monocyte-derived DC vs. Hematopoietic Stem Cell Derived-DC. Immune Netw. 2012, 12, 269. [Google Scholar] [CrossRef]
- Pardi, N.; Hogan, M.J.; Pelc, R.S.; Muramatsu, H.; Andersen, H.; DeMaso, C.R.; Dowd, K.A.; Sutherland, L.L.; Scearce, R.M.; Parks, R.; et al. Zika virus protection by a single low-dose nucleoside-modified mRNA vaccination. Nature 2017, 543, 248–251. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, H.; You, X.; Wang, X.; Cui, L.; Wang, Z.; Xu, F.; Li, M.; Yang, Z.; Liu, J.; Huang, P.; et al. Delivery of mRNA vaccine with a lipid-like material potentiates antitumor efficacy through toll-like receptor 4 signaling. Proc. Natl. Acad. Sci. USA 2021, 118, e2005191118. [Google Scholar] [CrossRef] [PubMed]
- Van Lint, S.; Goyvaerts, C.; Maenhout, S.; Goethals, L.; Disy, A.; Benteyn, D.; Pen, J.; Bonehill, A.; Heirman, C.; Breckpot, K.; et al. Preclinical evaluation of TriMix and antigen mRNA-based antitumor therapy. Cancer Res. 2012, 72, 1661–1671. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Beuckelaer, A.; Pollard, C.; Van Lint, S.; Roose, K.; Van Hoecke, L.; Naessens, T.; Udhayakumar, V.K.; Smet, M.; Sanders, N.; Lienenklaus, S.; et al. Type I interferons interfere with the capacity of mRNA lipoplex vaccines to elicit Cytolytic T cell responses. Mol. Ther. 2016, 24, 2012–2020. [Google Scholar] [CrossRef] [Green Version]
- Phung, C.D.; Pham, T.T.; Nguyen, H.T.; Nguyen, T.T.; Ou, W.; Jeong, J.H.; Choi, H.G.; Ku, S.K.; Yong, C.S.; Kim, J.O. Anti-CTLA-4 antibody-functionalized dendritic cell-derived exosomes targeting tumor-draining lymph nodes for effective induction of antitumor T-cell responses. Acta Biomater. 2020, 115, 371–382. [Google Scholar] [CrossRef]
- Matsumoto, A.; Asuka, M.; Takahashi, Y.; Takakura, Y. Antitumor immunity by small extracellular vesicles collected from activated dendritic cells through effective induction of cellular and humoral immune responses. Biomaterials 2020, 252, 120112. [Google Scholar] [CrossRef]
- Kedmi, R.; Ben-Arie, N.; Peer, D. The systemic toxicity of positively charged lipid nanoparticles and the role of Toll-like receptor 4 in immune activation. Biomaterials 2010, 31, 6867–6875. [Google Scholar] [CrossRef]
- Lonez, C.; Vandenbranden, M.; Ruysschaert, J.M. Cationic lipids activate intracellular signaling pathways. Adv. Drug Deliv. Rev. 2012, 64, 1749–1758. [Google Scholar] [CrossRef]
- Sen’kova, A.V.; Mironova, N.L.; Patutina, O.A.; Ageeva, T.A.; Zenkova, M.A. The Toxic Effects of Polychemotherapy onto the Liver Are Accelerated by the Upregulated MDR of Lymphosarcoma. ISRN Oncol. 2012, 2012, 721612. [Google Scholar] [CrossRef] [Green Version]
- Mironova, N.L.; Petrushanko, I.Y.; Patutina, O.A.; Sen’kova, A.V.; Simonenko, O.V.; Mitkevich, V.A.; Markov, O.V.; Zenkova, M.A.; Makarov, A.A. Ribonuclease binase inhibits primary tumor growth and metastases via apoptosis induction in tumor cells. Cell Cycle 2013, 12, 2120–2131. [Google Scholar] [CrossRef]
- Clarke, S.R.M. The critical role of CD40/CD40L in the CD4-dependent generation of CD8+ T cell immunity. J. Leukoc. Biol. 2000, 67, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Jenkins, S.J.; Perona-Wright, G.; MacDonald, A.S. Full Development of Th2 Immunity Requires Both Innate and Adaptive Sources of CD154. J. Immunol. 2008, 180, 8083–8092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Díaz, Á.; González-Alayón, I.; Pérez-Torrado, V.; Suárez-Martins, M. CD40-CD154: A perspective from type 2 immunity. Semin. Immunol. 2021, 53, 101528. [Google Scholar] [CrossRef] [PubMed]
- Guevara, M.L.; Jilesen, Z.; Stojdl, D.; Persano, S. Codelivery of mRNA with α-galactosylceramide using a new lipopolyplex formulation induces a strong antitumor response upon intravenous administration. ACS Omega 2019, 4, 13015–13026. [Google Scholar] [CrossRef] [Green Version]
- Miao, L.; Li, L.; Huang, Y.; Delcassian, D.; Chahal, J.; Han, J.; Shi, Y.; Sadtler, K.; Gao, W.; Lin, J.; et al. Delivery of mRNA vaccines with heterocyclic lipids increases anti-tumor efficacy by STING-mediated immune cell activation. Nat. Biotechnol. 2019, 37, 1174–1185. [Google Scholar] [CrossRef]
- Liu, L.; Wang, Y.; Miao, L.; Liu, Q.; Musetti, S.; Li, J.; Huang, L. Combination Immunotherapy of MUC1 mRNA Nano-vaccine and CTLA-4 Blockade Effectively Inhibits Growth of Triple Negative Breast Cancer. Mol. Ther. 2018, 26, 45–55. [Google Scholar] [CrossRef] [Green Version]
- Mai, Y.; Guo, J.; Zhao, Y.; Ma, S.; Hou, Y.; Yang, J. Intranasal delivery of cationic liposome-protamine complex mRNA vaccine elicits effective anti-tumor immunity. Cell. Immunol. 2020, 354, 104143. [Google Scholar] [CrossRef]
- Van Der Jeught, K.; De Koker, S.; Bialkowski, L.; Heirman, C.; Tjok Joe, P.; Perche, F.; Maenhout, S.; Bevers, S.; Broos, K.; Deswarte, K.; et al. Dendritic Cell Targeting mRNA Lipopolyplexes Combine Strong Antitumor T-Cell Immunity with Improved Inflammatory Safety. ACS Nano 2018, 12, 9815–9829. [Google Scholar] [CrossRef]
- Zhang, R.; Tang, L.; Tian, Y.; Ji, X.; Hu, Q.; Zhou, B.; Ding, Z.; Xu, H.; Yang, L. DP7-C-modified liposomes enhance immune responses and the antitumor effect of a neoantigen-based mRNA vaccine. J. Control. Release 2020, 328, 210–221. [Google Scholar] [CrossRef]
- Verbeke, R.; Lentacker, I.; Breckpot, K.; Janssens, J.; Van Calenbergh, S.; De Smedt, S.C.; Dewitte, H. Broadening the Message: A Nanovaccine Co-loaded with Messenger RNA and α-GalCer Induces Antitumor Immunity through Conventional and Natural Killer T Cells. ACS Nano 2019, 13, 1655–1669. [Google Scholar] [CrossRef]
- Yang, J.; Arya, S.; Lung, P.; Lin, Q.; Huang, J.; Li, Q. Hybrid nanovaccine for the co-delivery of the mRNA antigen and adjuvant. Nanoscale 2019, 11, 21782–21789. [Google Scholar] [CrossRef] [PubMed]
- Pick, H.; Schmidt, E.L.; Tairi, A.P.; Ilegems, E.; Hovius, R.; Vogel, H. Investigating cellular signaling reactions in single attoliter vesicles. J. Am. Chem. Soc. 2005, 127, 2908–2912. [Google Scholar] [CrossRef] [PubMed]
- Gomzikova, M.O.; Zhuravleva, M.N.; Vorobev, V.V.; Salafutdinov, I.I.; Laikov, A.V.; Kletukhina, S.K.; Martynova, E.V.; Tazetdinova, L.G.; Ntekim, A.I.; Khaiboullina, S.F.; et al. Angiogenic activity of cytochalasin B-induced membrane vesicles of human mesenchymal stem cells. Cells 2020, 9, 95. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chulpanova, D.S.; Gilazieva, Z.E.; Akhmetzyanova, E.R.; Kletukhina, S.K.; Rizvanov, A.A.; Solovyeva, V.V. Cytochalasin B-induced membrane vesicles from human mesenchymal stem cells overexpressing TRAIL, PTEN and IFN-β1 can kill carcinoma cancer cells. Tissue Cell 2021, 73, 101664. [Google Scholar] [CrossRef] [PubMed]
- Kletukhina, S.K.; Neustroeva, O.A.; Kurbangaleeva, S.V.; Salafutdinov, I.I.; Rogov, A.M.; James, V.; Rizvanov, A.A.; Gomzikova, M.O. Storage stability and delivery potential of cytochalasin B induced membrane vesicles. Biotechnol. Rep. 2021, 30, e00616. [Google Scholar] [CrossRef] [PubMed]
- Solovyeva, V.V.; Kitaeva, K.V.; Chulpanova, D.S.; Arkhipova, S.S.; Filin, I.Y.; Rizvanov, A.A. Analysis of the Interaction of Human Neuroblastoma Cell-Derived Cytochalasin B Induced Membrane Vesicles with Mesenchymal Stem Cells Using Imaging Flow Cytometry. Bionanoscience 2022, 12, 293–301. [Google Scholar] [CrossRef]
- Ukrainskaya, V.; Rubtsov, Y.; Pershin, D.; Podoplelova, N.; Terekhov, S.; Yaroshevich, I.; Sokolova, A.; Bagrov, D.; Kulakovskaya, E.; Shipunova, V.; et al. Antigen-Specific Stimulation and Expansion of CAR-T Cells Using Membrane Vesicles as Target Cell Surrogates. Small 2021, 17, 2102643. [Google Scholar] [CrossRef]
- Durán-Aniotz, C.; Segal, G.; Salazar, L.; Pereda, C.; Falcón, C.; Tempio, F.; Aguilera, R.; González, R.; Pérez, C.; Tittarelli, A.; et al. The immunological response and post-treatment survival of DC-vaccinated melanoma patients are associated with increased Th1/Th17 and reduced Th3 cytokine responses. Cancer Immunol. Immunother. 2013, 62, 761–772. [Google Scholar] [CrossRef]
- Yamamoto, M.; Kamigaki, T.; Yamashita, K.; Hori, Y.; Hasegawa, H.; Kuroda, D.; Moriyama, H.; Nagata, M.; Ku, Y.; Kuroda, Y. Enhancement of anti-tumor immunity by high levels of Th1 and Th17 with a combination of dendritic cell fusion hybrids and regulatory T cell depletion in pancreatic cancer. Oncol. Rep. 2009, 22, 337–343. [Google Scholar] [CrossRef] [Green Version]
- Miyabayashi, T.; Kagamu, H.; Koshio, J.; Ichikawa, K.; Baba, J.; Watanabe, S.; Tanaka, H.; Tanaka, J.; Yoshizawa, H.; Nakata, K.; et al. Vaccination with CD133 + melanoma induces specific Th17 and Th1 cell-mediated antitumor reactivity against parental tumor. Cancer Immunol. Immunother. 2011, 60, 1597–1608. [Google Scholar] [CrossRef]
- Nistor, G.I.; Dillman, R.O. Cytokine network analysis of immune responses before and after autologous dendritic cell and tumor cell vaccine immunotherapies in a randomized trial. J. Transl. Med. 2020, 18, 176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Montico, B.; Lapenta, C.; Ravo, M.; Martorelli, D.; Muraro, E.; Zeng, B.; Comaro, E.; Spada, M.; Donati, S.; Santini, S.M.; et al. Exploiting a new strategy to induce immunogenic cell death to improve dendritic cell-based vaccines for lymphoma immunotherapy. Oncoimmunology 2017, 6, e1356964. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cannon, M.J.; Goyne, H.; Stone, P.J.B.; Chiriva-Internati, M. Dendritic cell vaccination against ovarian cancer-tipping the Treg/T H17 balance to therapeutic advantage? Expert Opin. Biol. Ther. 2011, 11, 441–445. [Google Scholar] [CrossRef] [Green Version]
- Terhune, J.; Berk, E.; Czerniecki, B. Dendritic Cell-Induced Th1 and Th17 Cell Differentiation for Cancer Therapy. Vaccines 2013, 1, 527–549. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martin-Orozco, N.; Muranski, P.; Chung, Y.; Yang, X.O.; Yamazaki, T.; Lu, S.; Hwu, P.; Restifo, N.P.; Overwijk, W.W.; Dong, C. T Helper 17 Cells Promote Cytotoxic T Cell Activation in Tumor Immunity. Immunity 2009, 31, 787–798. [Google Scholar] [CrossRef] [Green Version]
- Gnerlich, J.L.; Mitchem, J.B.; Weir, J.S.; Sankpal, N.V.; Kashiwagi, H.; Belt, B.A.; Porembka, M.R.; Herndon, J.M.; Eberlein, T.J.; Goedegebuure, P.; et al. Induction of Th17 Cells in the Tumor Microenvironment Improves Survival in a Murine Model of Pancreatic Cancer. J. Immunol. 2010, 185, 4063–4071. [Google Scholar] [CrossRef]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Sequences of Primers and Probes, 5′ → 3′ | Amplicon Size, bp 1 |
|---|---|---|
| CD27 | F 2: TTGTGACCTTCTCCAGCATG R: GTAAGGACAAGGCTCTTCAGG Probe: ((5,6)-FAM)-AAGAACAAGATTGCACCCAGGACGA-BHQ1 | 122 |
| CD28 | F: GTTGCTGGAGTCCTGTTTTG R: CTGGTAAGGCTTTCGAGTGAG Probe: ((5,6)-FAM)-ATGGCTTGCTAGTGACAGTGGCT–BHQ1 | 147 |
| CD40L | F: GTCAGCATGATAGAAACATACAGC R: TGGGTGATAAGGAAAACAGTAAGT Probe: ((5,6)-FAM)-CCCCCAGATCCGTGGCAACT–BHQ1 | 110 |
| 4-1BBL | F: TGCCCCAACACTACACAAC R: TCTTCGTACCTCAGACCTTGA Probe: ((5,6)-FAM)-CTCTCCTGTGTTCGCCAAGCTACT–BHQ1 | 146 |
| PD1 | F: GGTACCCTGGTCATTCACTTG R: ATTTGCTCCCTCTGACACTG Probe: ((5,6)-FAM)-ACCTCTAGAAGCCACCCTGATTGC–BHQ1 | 133 |
| CTLA4 | F: TCTGCAAGGTGGAACTCATG R: AGCTAACTGCGACAAGGATC Probe: ((5,6)-FAM)-ATAAATCTGCGTCCCGTTGCCCA–BHQ1 | 132 |
| TIGIT | F: CTGTAGGCCTCTGGTTAGAAG R: TGACAGAGCCACCTTCCT Probe: ((5,6)-FAM)-CAGCCTGTATCAGCCCCTGGAC–BHQ1 | 149 |
| TIM3 | F: ACCCTGGCACTTATCATTGG R: TTTTCCTCAGAGCGAATCCTG Probe: ((5,6)-FAM)-TCCTGCATTTGCCAACCCTCCT–BHQ1 | 149 |
| T-bet | F: TTCAACCAGCACCAGACAG R: AGACCACATCCACAAACATCC Probe: ((5,6)-FAM)-TCACTAAGCAAGGACGGCGAATGT–BHQ1 | 124 |
| GATA3 | F: ACCTCACCACCCTTCCA R: TTCATGATACTGCTCCTGCG Probe: ((5,6)-FAM)-CTCCGACCCCTTCTACTTGCGTTTT–BHQ1 | 141 |
| RORγ | F: TTTCTGAGGATGAGATTGCCC R: TTGTCGATGAGTCTTGCAGAG Probe: ((5,6)-FAM)-CCAGGACGGTTGGCATTGATGAGA–BHQ1 | 146 |
| Foxp3 | F: AAGTACCACAATATGCGACCC R: TCTGAAGTAGGCGAACATGC Probe: ((5,6)-FAM)-TCACCTATGCCACCCTTATCCGATG–BHQ1 | 132 |
| β-actin | F: TATTGGCAACGAGCGGTTCC R: TGGCATAGAGGTCTTTACGG Probe: ((5,6)-ROX)-CCAGCCTTCCTTCTTGGGTATGGAATCC–BHQ2 | 140 |
| Type | Name 1 | Color Code 2 | Vaccine Composition | Vaccination Regimen 3 | Dose, per Mouse |
|---|---|---|---|---|---|
| Cellular vaccines | DC/L(×1) |  | DCs transfected with RNA-RLS40/L | d7 | 105 cells |
| DC/L(×2) |  | d7, d14 | |||
| DC/ML(×1) |  | DCs transfected with RNA-RLS40/ML | d7 | ||
| DC/ML(×2) |  | d7, d14 | |||
| Liposomal vaccines | L(×1) |  | RNA-RLS40/L lipoplexes | d7 | 5 µg RNA |
| L(×2) |  | d7, d14 | |||
| ML(×1) |  | RNA-RLS40/ML lipoplexes | d7 | ||
| ML(×2) |  | d7, d14 | |||
| Vesicular vaccines | MV/L(×1) |  | CIMVs isolated from DCs transfected with RNA-RLS40/L | d7 | 15 µg (total protein count) |
| MV/L(×2) |  | d7, d14 | |||
| MV/ML(×1) |  | CIMVs isolated from DCs transfected with RNA-RLS40/ML | d7 | ||
| MV/ML(×2) |  | d7, d14 |
| Groups | Color Code | Mitotic Cells, Nv | Caspase-3 Positive Cells, Nv |
|---|---|---|---|
| Control |  | 5.6 ± 0.5 | 1.4 ± 0.1 |
| DC/L(×1) |  | 4 ± 0.7 * | 1.6 ± 0.1 |
| DC/L(×2) |  | 2 ± 0.2 * | 6.6 ± 0.9 * |
| DC/ML(×1) |  | 2.6 ± 0.8 * | 1.9 ± 0.5 |
| DC/ML(×2) |  | 1.7 ± 0.4 * | 4.8 ± 0.4 * |
| L(×1) |  | 4 ± 1.5 | 2.2 ± 0.4 |
| L(×2) |  | 5.4 ± 1.5 | 1.3 ± 0.1 |
| ML(×1) |  | 4.1 ± 0.5 * | 0.9 ± 0.4 |
| ML(×2) |  | 5.2 ± 1 | 1.4 ± 0.1 |
| CIMV/L(×1) |  | 4.2 ± 0.2 * | 1.5 ± 0.5 |
| CIMV/L(×2) |  | 2.5 ± 0.6 * | 6.3 ± 0.4 * |
| CIMV/ML(×1) |  | 5 ± 0.7 | 2.6 ± 0.8 * |
| CIMV/ML(×2) |  | 2.1 ± 0.7* | 5.8 ± 1.4 * |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Markov, O.V.; Sen’kova, A.V.; Mohamed, I.S.; Shmendel, E.V.; Maslov, M.A.; Oshchepkova, A.L.; Brenner, E.V.; Mironova, N.L.; Zenkova, M.A. Dendritic Cell-Derived Artificial Microvesicles Inhibit RLS40 Lymphosarcoma Growth in Mice via Stimulation of Th1/Th17 Immune Response. Pharmaceutics 2022, 14, 2542. https://doi.org/10.3390/pharmaceutics14112542
Markov OV, Sen’kova AV, Mohamed IS, Shmendel EV, Maslov MA, Oshchepkova AL, Brenner EV, Mironova NL, Zenkova MA. Dendritic Cell-Derived Artificial Microvesicles Inhibit RLS40 Lymphosarcoma Growth in Mice via Stimulation of Th1/Th17 Immune Response. Pharmaceutics. 2022; 14(11):2542. https://doi.org/10.3390/pharmaceutics14112542
Chicago/Turabian StyleMarkov, Oleg V., Aleksandra V. Sen’kova, Islam S. Mohamed, Elena V. Shmendel, Mikhail A. Maslov, Anastasiya L. Oshchepkova, Evgeniy V. Brenner, Nadezhda L. Mironova, and Marina A. Zenkova. 2022. "Dendritic Cell-Derived Artificial Microvesicles Inhibit RLS40 Lymphosarcoma Growth in Mice via Stimulation of Th1/Th17 Immune Response" Pharmaceutics 14, no. 11: 2542. https://doi.org/10.3390/pharmaceutics14112542