
Combination of Nanovectorized siRNA Directed against Survivin with Doxorubicin for Efficient Anti-Cancer Activity in HER2+ Breast Cancer Cells
 , , ,
, , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Nanocarrier Synthesis and Formulation
2.3. Nanocarrier Characterization
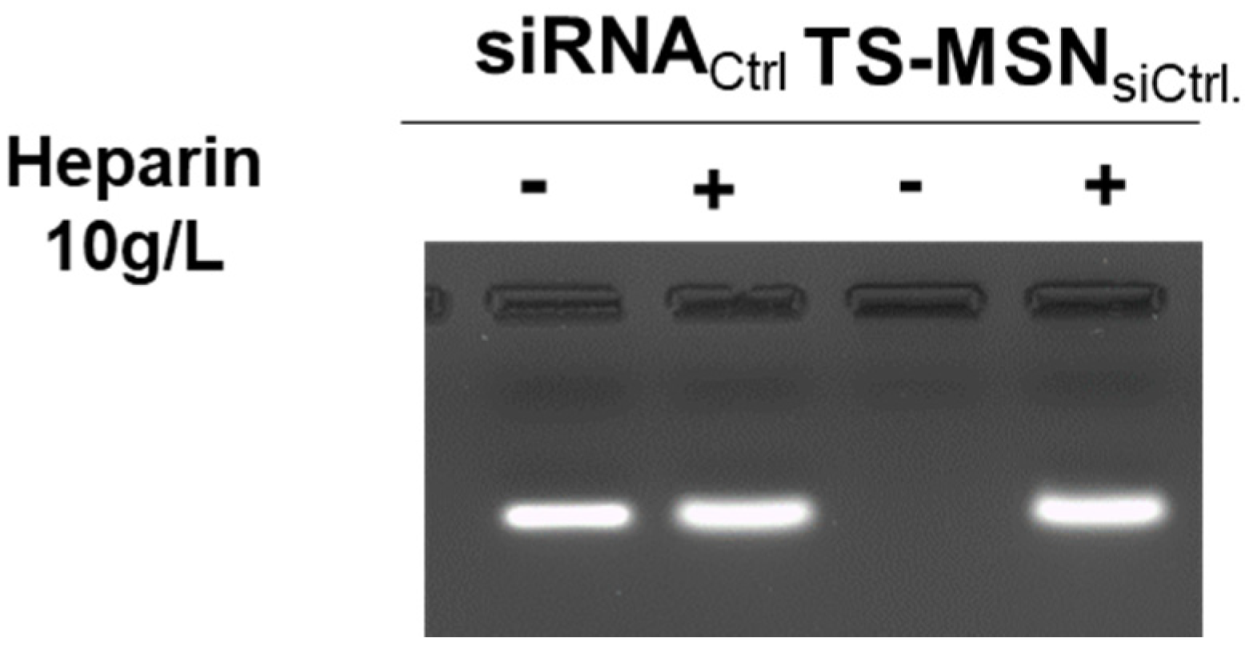
2.4. SiRNA Complexation Assay by Agarose Gel Electrophoresis
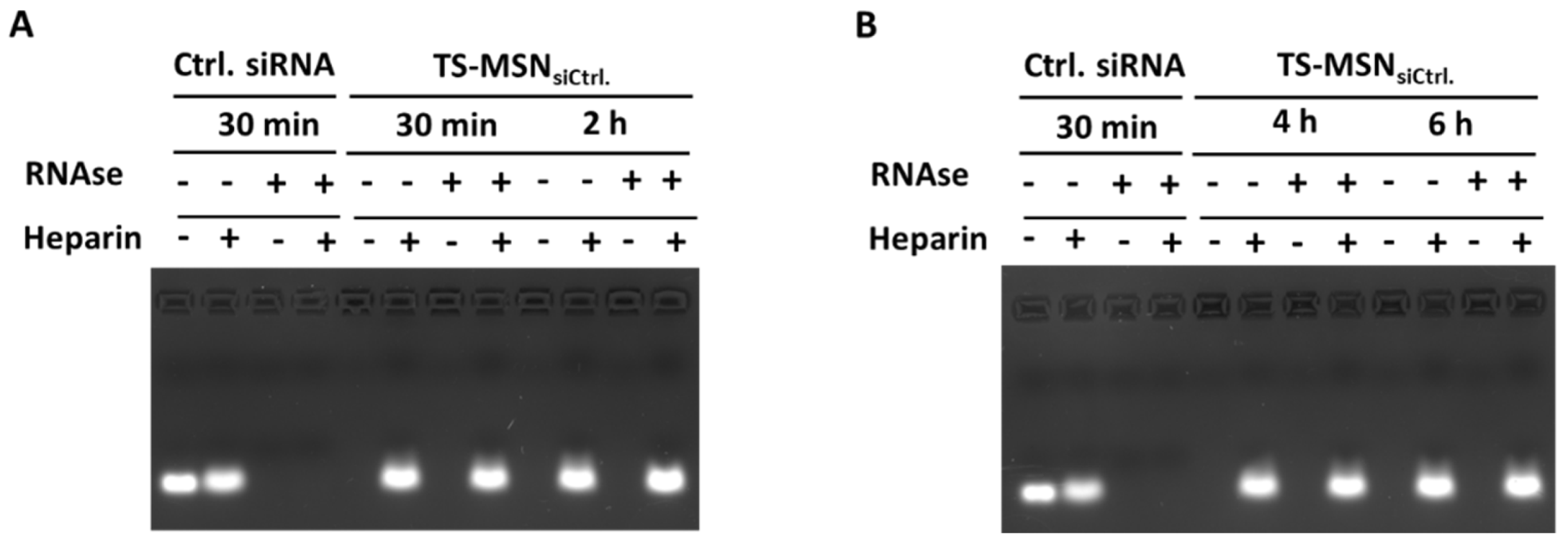
2.5. SiRNA Protection against Enzymatic Degradation
2.6. Cell Culture
2.7. SiRNA Transfection
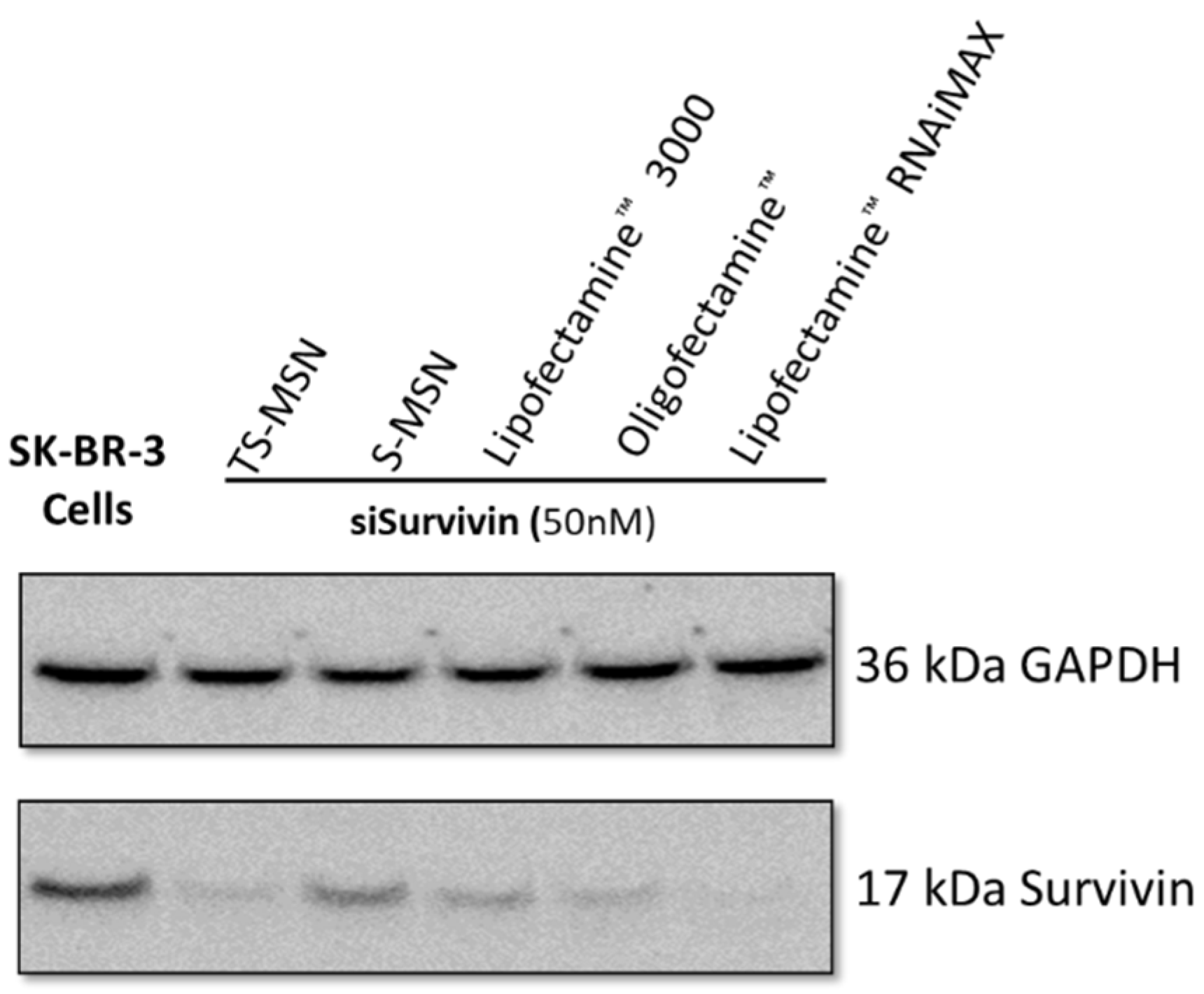
2.8. Protein Extraction and Western Blot
2.9. Design of Experiments
2.10. RNA Extraction and QRT-PCR Analysis
2.11. Cytotoxicity of TS-MSN Components
2.12. Cytotoxicity of DOX (IC50 Determination)
2.13. Cytotoxicity of The Combination of TS-MSN Formulated with SiSurvivin and DOX
2.14. Statistical Analysis
3. Results and Discussion
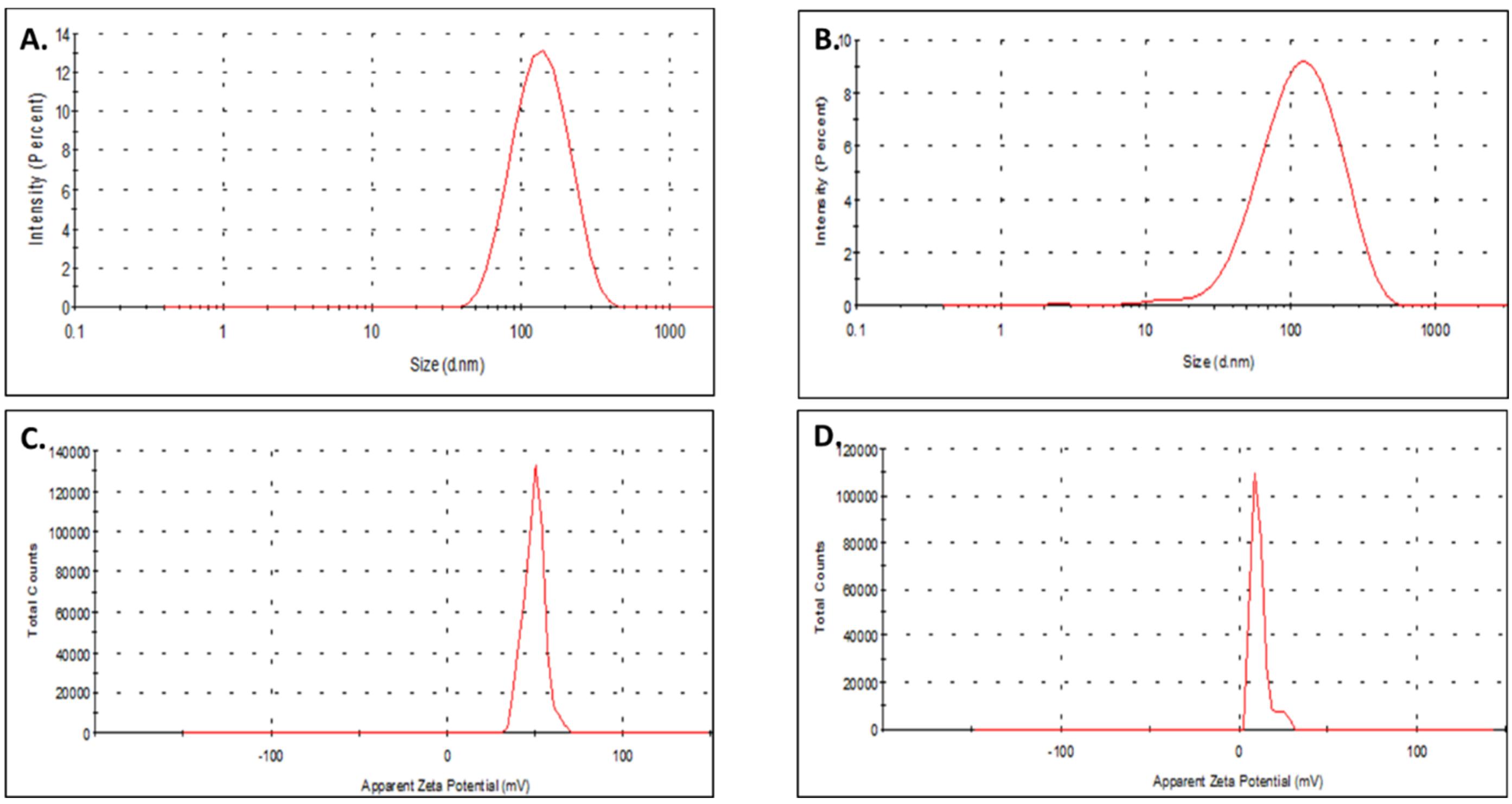
3.1. Formulation and Physicochemical Characterization of TS-MSN
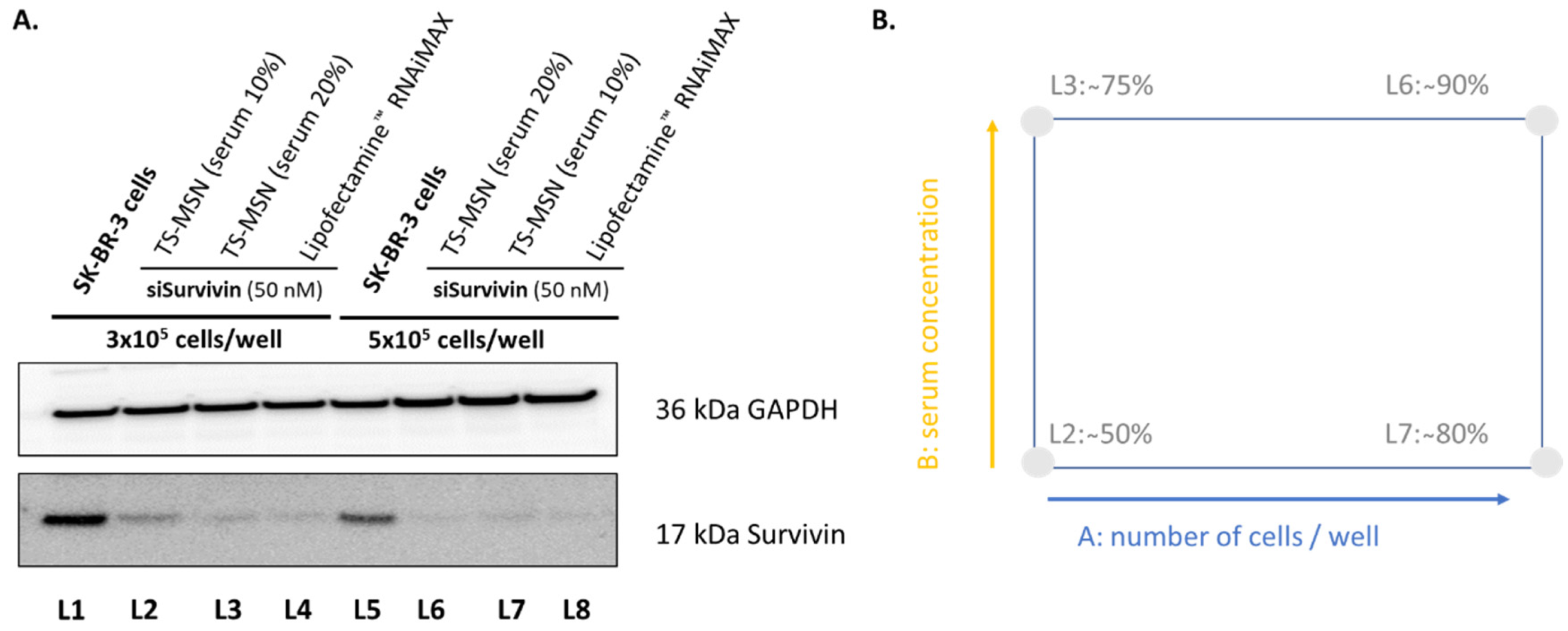
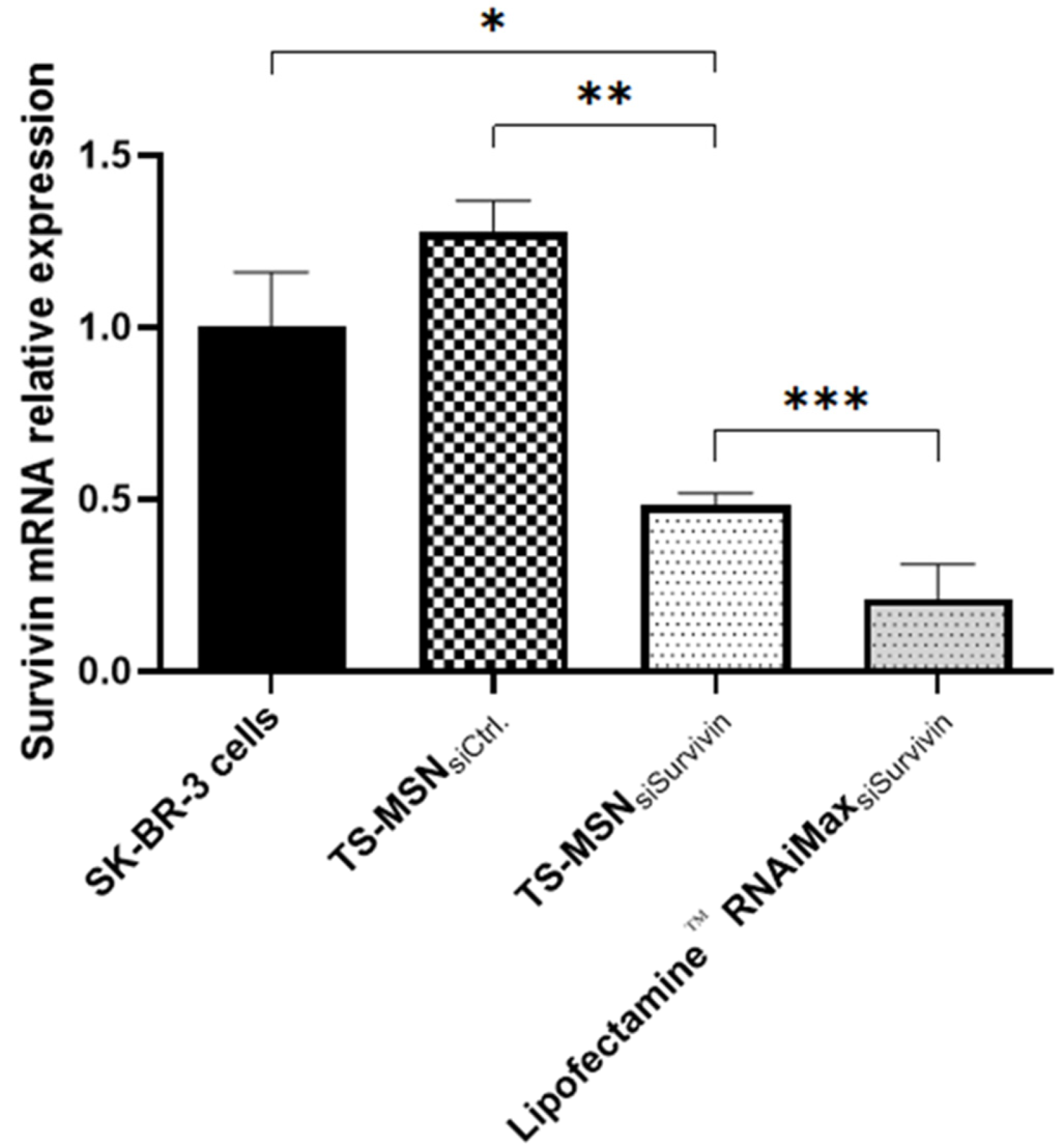
3.2. The Downregulation Efficiency of TS-MSN on HER2+ Breast Cancer Cells
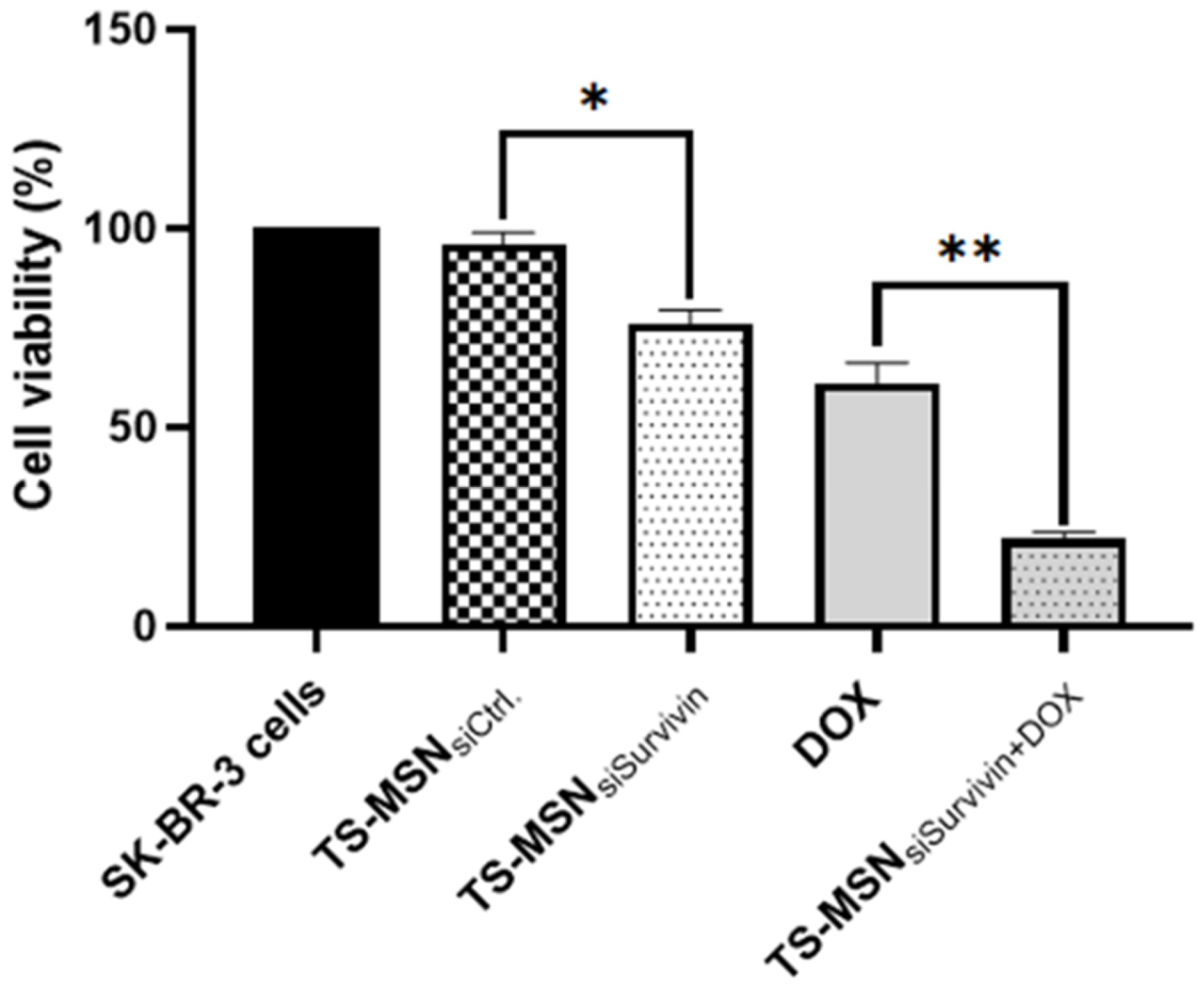
3.3. In Vitro Cytotoxicity of TS-MSN, Doxorubicin, and a Combination of Both on HER2+ Breast Cancer Cells
4. Conclusion and Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ferlay, J.; Colombet, M.; Soerjomataram, I.; Parkin, D.M.; Piñeros, M.; Znaor, A.; Bray, F. Cancer Statistics for the Year 2020: An Overview. Int. J. Cancer 2021, 149, 778–789. [Google Scholar] [CrossRef] [PubMed]
- Sung, H.; Ferlay, J.; Siegel, R.L.; Laversanne, M.; Soerjomataram, I.; Jemal, A.; Bray, F. Global Cancer Statistics 2020: GLOBOCAN Estimates of Incidence and Mortality Worldwide for 36 Cancers in 185 Countries. CA A Cancer J. Clin. 2021, 71, 209–249. [Google Scholar] [CrossRef] [PubMed]
- Kittaneh, M.; Montero, A.J.; Glück, S. Molecular Profiling for Breast Cancer: A Comprehensive Review. Biomark. Cancer 2013, 5, 61–70. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slamon, D.; Eiermann, W.; Robert, N.; Pienkowski, T.; Martin, M.; Press, M.; Mackey, J.; Glaspy, J.; Chan, A.; Pawlicki, M.; et al. Adjuvant Trastuzumab in HER2-Positive Breast Cancer. N. Engl. J. Med. 2011, 365, 1273–1283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thorn, C.F.; Oshiro, C.; Marsh, S.; Hernandez-Boussard, T.; McLeod, H.; Klein, T.E.; Altman, R.B. Doxorubicin Pathways: Pharmacodynamics and Adverse Effects. Pharm. Genom. 2011, 21, 440–446. [Google Scholar] [CrossRef]
- Anders, C.K.; Adamo, B.; Karginova, O.; Deal, A.M.; Rawal, S.; Darr, D.; Schorzman, A.; Santos, C.; Bash, R.; Kafri, T.; et al. Pharmacokinetics and Efficacy of PEGylated Liposomal Doxorubicin in an Intracranial Model of Breast Cancer. PLoS ONE 2013, 8, e61359. [Google Scholar] [CrossRef] [Green Version]
- Pommier, Y.; Leo, E.; Zhang, H.; Marchand, C. DNA Topoisomerases and Their Poisoning by Anticancer and Antibacterial Drugs. Chem. Biol. 2010, 17, 421–433. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.-Y.; Kim, S.-J.; Kim, B.-J.; Rah, S.-Y.; Chung, S.M.; Im, M.-J.; Kim, U.-H. Doxorubicin-Induced Reactive Oxygen Species Generation and Intracellular Ca2+increase Are Reciprocally Modulated in Rat Cardiomyocytes. Exp. Mol. Med. 2006, 38, 535–545. [Google Scholar] [CrossRef] [Green Version]
- Hager, S.; Fittler, F.J.; Wagner, E.; Bros, M. Nucleic Acid-Based Approaches for Tumor Therapy. Cells 2020, 9, 2061. [Google Scholar] [CrossRef]
- Sen, M.; Grandis, J.R. Nucleic Acid-Based Approaches to STAT Inhibition. Jak-Stat 2012, 1, 285–291. [Google Scholar] [CrossRef]
- Arami, S.; Rashidi, M.; Mahdavi, M.; Fathi, M.; Entezami, A. Synthesis and Characterization of Fe3O4 -PEG-LAC-Chitosan-PEI Nanoparticle as a Survivin SiRNA Delivery System. Hum. Exp. Toxicol. 2017, 36, 227–237. [Google Scholar] [CrossRef]
- Bruniaux, J.; Allard-Vannier, E.; Aubrey, N.; Lakhrif, Z.; Ben Djemaa, S.; Eljack, S.; Marchais, H.; Hervé-Aubert, K.; Chourpa, I.; David, S. Magnetic Nanocarriers for the Specific Delivery of SiRNA: Contribution of Breast Cancer Cells Active Targeting for down-Regulation Efficiency. Int. J. Pharm. 2019, 569, 118572. [Google Scholar] [CrossRef]
- Alric, C.; Aubrey, N.; Allard-Vannier, É.; di Tommaso, A.; Blondy, T.; Dimier-Poisson, I.; Chourpa, I.; Hervé-Aubert, K. Covalent Conjugation of Cysteine-Engineered ScFv to PEGylated Magnetic Nanoprobes for Immunotargeting of Breast Cancer Cells. RSC Adv. 2016, 6, 37099–37109. [Google Scholar] [CrossRef]
- Janko, C.; Ratschker, T.; Nguyen, K.; Zschiesche, L.; Tietze, R.; Lyer, S.; Alexiou, C. Functionalized Superparamagnetic Iron Oxide Nanoparticles (SPIONs) as Platform for the Targeted Multimodal Tumor Therapy. Front. Oncol. 2019, 9, 59. [Google Scholar] [CrossRef] [Green Version]
- Palanisamy, S.; Wang, Y.-M. Superparamagnetic Iron Oxide Nanoparticulate System: Synthesis, Targeting, Drug Delivery and Therapy in Cancer. Dalton. Trans. 2019, 48, 9490–9515. [Google Scholar] [CrossRef]
- Hervé, K.; Douziech-Eyrolles, L.; Munnier, E.; Cohen-Jonathan, S.; Soucé, M.; Marchais, H.; Limelette, P.; Warmont, F.; Saboungi, M.L.; Dubois, P.; et al. The Development of Stable Aqueous Suspensions of PEGylated SPIONs for Biomedical Applications. Nanotechnology 2008, 19, 465608. [Google Scholar] [CrossRef]
- Allard-Vannier, E.; Cohen-Jonathan, S.; Gautier, J.; Hervé-Aubert, K.; Munnier, E.; Soucé, M.; Legras, P.; Passirani, C.; Chourpa, I. Pegylated Magnetic Nanocarriers for Doxorubicin Delivery: A Quantitative Determination of Stealthiness in Vitro and in Vivo. Eur. J. Pharm. Biopharm. 2012, 81, 498–505. [Google Scholar] [CrossRef]
- Alric, C.; Hervé-Aubert, K.; Aubrey, N.; Melouk, S.; Lajoie, L.; Même, W.; Même, S.; Courbebaisse, Y.; Ignatova, A.A.; Feofanov, A.V.; et al. Targeting HER2-Breast Tumors with ScFv-Decorated Bimodal Nanoprobes. J. Nanobiotechnol. 2018, 16, 18. [Google Scholar] [CrossRef] [Green Version]
- Bruniaux, J.; Djemaa, S.B.; Hervé-Aubert, K.; Marchais, H.; Chourpa, I.; David, S. Stealth Magnetic Nanocarriers of SiRNA as Platform for Breast Cancer Theranostics. Int. J. Pharm. 2017, 532, 660–668. [Google Scholar] [CrossRef]
- Altieri, D.C. Validating Survivin as a Cancer Therapeutic Target. Nat. Rev. Cancer. 2003, 3, 46–54. [Google Scholar] [CrossRef]
- Altieri, D.C. Targeting Survivin in Cancer. Cancer. Lett. 2013, 332, 225–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lv, Y.-G.; Yu, F.; Yao, Q.; Chen, J.-H.; Wang, L. The Role of Survivin in Diagnosis, Prognosis and Treatment of Breast Cancer. J. Thorac. Dis. 2010, 2, 100–110. [Google Scholar] [PubMed]
- Cui, M.; Au, J.L.-S.; Wientjes, M.G.; O’Donnell, M.A.; Loughlin, K.R.; Lu, Z. Intravenous SiRNA Silencing of Survivin Enhances Activity of Mitomycin C in Human Bladder RT4 Xenografts. J. Urol. 2015, 194, 230–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, X.; Zheng, J.; Shi, H.; Zhang, Z.; Wang, W. Survivin Knockdown Enhances Gastric Cancer Cell Sensitivity to Radiation and Chemotherapy In Vitro and in Nude Mice. Am. J. Med. Sci. 2012, 344, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Dong, H.; Yao, L.; Bi, W.; Wang, F.; Song, W.; Lv, Y. Combination of Survivin SiRNA with Neoadjuvant Chemotherapy Enhances Apoptosis and Reverses Drug Resistance in Breast Cancer MCF-7 Cells. J. Can. Res. 2015, 11, 717. [Google Scholar] [CrossRef]
- Liu, W.; Zhu, F.; Jiang, Y.; Sun, D.; Yang, B.; Yan, H. SiRNA Targeting Survivin Inhibits the Growth and Enhances the Chemosensitivity of Hepatocellular Carcinoma Cells. Oncol. Rep. 2013, 29, 1183–1188. [Google Scholar] [CrossRef] [Green Version]
- Massart, R. Preparation of Aqueous Magnetic Liquids in Alkaline and Acidic Media. IEEE Trans. Magn. 1981, 17, 1247–1248. [Google Scholar] [CrossRef]
- Perillo, E.; Hervé-Aubert, K.; Allard-Vannier, E.; Falanga, A.; Galdiero, S.; Chourpa, I. Synthesis and in Vitro Evaluation of Fluorescent and Magnetic Nanoparticles Functionalized with a Cell Penetrating Peptide for Cancer Theranosis. J. Colloid Interface Sci. 2017, 499, 209–217. [Google Scholar] [CrossRef]
- Bustin, S.A. A-Z of Quantitative PCR; International University Line: La Jolla, CA, USA, 2004. [Google Scholar]
- David, S.; Marchais, H.; Bedin, D.; Chourpa, I. Modelling the Response Surface to Predict the Hydrodynamic Diameters of Theranostic Magnetic SiRNA Nanovectors. Int. J. Pharm. 2015, 478, 409–415. [Google Scholar] [CrossRef]
- Chong, Z.X.; Yeap, S.K.; Ho, W.Y. Transfection Types, Methods and Strategies: A Technical Review. PeerJ 2021, 9, e11165. [Google Scholar] [CrossRef]
- Horibe, T.; Torisawa, A.; Akiyoshi, R.; Hatta-Ohashi, Y.; Suzuki, H.; Kawakami, K. Transfection Efficiency of Normal and Cancer Cell Lines and Monitoring of Promoter Activity by Single-cell Bioluminescence Imaging. Luminescence 2014, 29, 96–100. [Google Scholar] [CrossRef]
- Arami, S.; Mahdavi, M.; Rashidi, M.-R.; Yekta, R.; Rahnamay, M.; Molavi, L.; Hejazi, M.-S.; Samadi, N. Apoptosis Induction Activity and Molecular Docking Studies of Survivin SiRNA Carried by Fe3O4-PEG-LAC-Chitosan-PEI Nanoparticles in MCF-7 Human Breast Cancer Cells. J. Pharm. Biomed. Anal. 2017, 142, 145–154. [Google Scholar] [CrossRef]
- Schindler, C.; Collinson, A.; Matthews, C.; Pointon, A.; Jenkinson, L.; Minter, R.R.; Vaughan, T.J.; Tigue, N.J. Exosomal Delivery of Doxorubicin Enables Rapid Cell Entry and Enhanced in Vitro Potency. PLoS ONE 2019, 14, e0214545. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, P.L.; Elkamhawy, A.; Choi, Y.H.; Lee, C.H.; Lee, K.; Cho, J. Suppression of Tumor Growth and Cell Migration by Indole-Based Benzenesulfonamides and Their Synergistic Effects in Combination with Doxorubicin. Int. J. Mol. Sci. 2022, 23, 9903. [Google Scholar] [CrossRef]
- Xu, D.; Li, D.W.; Xie, J.; Chen, X.W. Effect and Mechanism of Survivin on Hypoxia-Induced Multidrug Resistance of Human Laryngeal Carcinoma Cells. BioMed Res. Int. 2019, 2019, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Ganesh, S.; Iyer, A.K.; Morrissey, D.V.; Amiji, M.M. Hyaluronic Acid Based Self-Assembling Nanosystems for CD44 Target Mediated SiRNA Delivery to Solid Tumors. Biomaterials 2013, 34, 3489–3502. [Google Scholar] [CrossRef] [Green Version]
- Li, Q.-X.; Zhao, J.; Liu, J.-Y.; Jia, L.-T.; Huang, H.-Y.; Xu, Y.-M.; Zhang, Y.; Zhang, R.; Wang, C.-J.; Yao, L.-B.; et al. Survivin Stable Knockdown by SiRNA Inhibits Tumor Cell Growth and Angiogenesis in Breast and Cervical Cancers. Cancer. Biol. Ther. 2006, 5, 860–866. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| siRNA | Sequence |
| Ctrl. sense | 5′-GGAAGAUCAUAAUGGACAG[dT][dT]-3′ |
| Ctrl. antisense | 5′-CUGUCCAUUAUGAUCUUCC[dT][dT]-3′ |
| Survivin sense | 5′-GUCUGGACCUCAUGUUGUU[dT][dT]-3′ |
| Survivin antisense | 5′-AACAACAUGAGGUCCAGAC[dT][dT]-3′ |
| primer | Sequence |
| GAPDH reverse | 5′-AGTTGTCATGGATGACCTTGG-3′ |
| GAPDH forward | 5′-CAAAAGGGTCATCATCTCTGC-3′ |
| Survivin reverse | 5′-CCGGACGAATGCTTTTTATG-3′ |
| Survivin forward | 5′-GCCCAGTGTTTCTTCTGCTT-3′ |
| Experiment | A: Number of Cells | B: Serum Concentration |
|---|---|---|
| 1 | 300,000 | 10% |
| 2 | 500,000 | 10% |
| 3 | 300,000 | 20% |
| 4 | 500,000 | 20% |
| Hydrodynamic Diameter (nm) | Polydispersity Index | Zeta Potential (mV) | |
|---|---|---|---|
| TS-MSNsiCtrl. | 131 ± 18 | 0.302 ± 0.038 | +12.44 ± 07.34 |
| TS-MSNsiSurvivin | 144 ± 30 | 0.288 ± 0.030 | +10.56 ± 05.70 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Eljack, S.; Allard-Vannier, E.; Misericordia, Y.; Hervé-Aubert, K.; Aubrey, N.; Chourpa, I.; Faggad, A.; David, S. Combination of Nanovectorized siRNA Directed against Survivin with Doxorubicin for Efficient Anti-Cancer Activity in HER2+ Breast Cancer Cells. Pharmaceutics 2022, 14, 2537. https://doi.org/10.3390/pharmaceutics14112537
Eljack S, Allard-Vannier E, Misericordia Y, Hervé-Aubert K, Aubrey N, Chourpa I, Faggad A, David S. Combination of Nanovectorized siRNA Directed against Survivin with Doxorubicin for Efficient Anti-Cancer Activity in HER2+ Breast Cancer Cells. Pharmaceutics. 2022; 14(11):2537. https://doi.org/10.3390/pharmaceutics14112537
Chicago/Turabian StyleEljack, Sahar, Emilie Allard-Vannier, Yoann Misericordia, Katel Hervé-Aubert, Nicolas Aubrey, Igor Chourpa, Areeg Faggad, and Stephanie David. 2022. "Combination of Nanovectorized siRNA Directed against Survivin with Doxorubicin for Efficient Anti-Cancer Activity in HER2+ Breast Cancer Cells" Pharmaceutics 14, no. 11: 2537. https://doi.org/10.3390/pharmaceutics14112537