
PAD4 and Its Inhibitors in Cancer Progression and Prognosis

Abstract
:
1. Introduction
2. Peptidyl Arginine Deiminase (PAD) and Citrullination
3. PAD Expression in Cancer and Associated Thrombosis
4. The Structure of PAD4 and Its Mechanism of Action
4.1. PAD4 Structure
4.2. Calcium Dependence and The Catalytic Mechanism of PAD4
5. PAD4-Mediated Protein Citrullination and Gene Regulation Components
5.1. Crosstalk between Components of PAD4-Mediated Histone Citrullination and Other Modifications
5.2. The Roles Played by PAD4 in Cell Differentiation and Apoptosis
5.3. PAD4-Mediated Nonhistone Protein Citrullination
5.4. PAD4 Autocitrullination
6. PAD4-Mediated NET Formation Is Involved in Cancer Progression
6.1. Key Role Played by PAD4 in NET Formation
6.2. Effects of NETs on Cancer Progression and Prognosis
7. PAD4 in Cancers
7.1. Gastric Carcinoma (GC)
7.2. Ovarian Carcinoma (OC)
7.3. Osteosarcoma (OS)
7.4. Nasopharyngeal Carcinoma (NPC)
7.5. Hepatic Carcinoma
7.6. Lung Carcinoma
7.7. Breast Carcinoma
7.8. Leukemia
8. PAD4 and Cancer-Associated Thrombosis (CAT)
9. PAD4 Inhibitors
9.1. Pan-PAD and PAD4 Inhibitors
9.2. Mechanism of Haloacetamidine PAD Inhibitors
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
Abbreviations
| CAT | Cancer-Associated Thrombosis |
| PAD4 | Peptidyl arginine deiminase 4 |
| VTE | Venous thromboembolism |
| ER | Estrogen receptor |
| PTM | Post-translational modification |
| NLS | Nuclear localization sequence |
| CRPC | Castration-resistant prostate cancer |
| HDAC1 | Histone deacetylase 1 |
| HDAC2 | Histone deacetylase 2 |
| ES | ES OCT4-GIP |
| iPS | Induced pluripotent stem cells |
| ATRA | All-trans retinoic acid |
| mTORC1 | Mammalian target of rapamycin complex 1 |
| CK | Cytokeratin |
| AT | Antithrombin |
| EMT | Epithelial–mesenchymal transition |
| GSK3β | Glycogen synthase kinase 3β |
| ETS | Erythroblast transformation-specific |
| EGF | Epidermal growth factor |
| NSCLC | Non-small cell lung cancer |
| CRC | Colorectal cancer |
| ECM | Extracellular matrix |
| NPM1 | Nucleophosmin |
| RPS2 | Ribosomal protein S2 |
| ROS | Reactive oxygen species |
| NETs | Neutrophil extracellular traps |
| NE | Neutrophil elastase |
| MPO | Myeloperoxidase |
| MMP9 | Matrix metalloproteinase 9 |
| CG | Cathepsin G |
| PMA | Phorbol myristate acetate |
| Abs | Antibodies |
| PKC | Protein kinase C |
| TLRs | Toll-like receptors |
| GM-CSF | Granulocyte-macrophage colony stimulating factor |
| LPS | Lipopolysaccharide |
| C5a | Complement factor 5a |
| H3Cit | Citrullinated histone H3 |
| G-CSF | Granulocyte colony-stimulating factor |
| HMGB1 | High-mobility group box 1 |
| CTLs | Cytotoxic T lymphocytes |
| NKs | Natural killer cells |
| cfDNA | Cell-free DNA |
| TFPI | Tissue factor pathway inhibitor |
| PSGL-1 | p-selectin glycoprotein ligand-1 |
| DAMPs | Damage-associated molecular patterns |
| GC | Gastric carcinoma |
| OC | Ovarian carcinoma |
| OS | Osteosarcoma |
| NPC | Nasopharyngeal carcinoma |
| PHC | Primary hepatic carcinoma |
| HCC | Hepatocellular carcinoma |
| ICC | Intrahepatic cholangiocarcinoma |
| EVs | Extracellular vesicles |
| MET | Mesenchymal-epithelial transition |
| CSC | Cancer stem cell |
| TNBC | Triple-negative breast cancer |
| CECNs | Cancer extracellular chromatin networks |
| HIFs | Hypoxia-inducible factors |
| HREs | Hypoxia response elements |
| APL | Acute promyelocytic leukemia |
| HIT | Heparin-induced thrombocytopenia |
| MDSCs | Myeloid suppressor cells |
References
- Cedervall, J.; Zhang, Y.; Olsson, A.-K. Tumor-Induced NETosis as a Risk Factor for Metastasis and Organ Failure. Cancer Res. 2016, 76, 4311–4315. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mehlen, P.; Puisieux, A. Metastasis: A question of life or death. Nat. Cancer 2006, 6, 449–458. [Google Scholar] [CrossRef] [PubMed]
- Fernandes, C.J.; Morinaga LT, K.; Alves, J.L.; Castro, M.A.; Calderaro, D.; Jardim, C.V.P.; Souza, R. Cancer-associated thrombosis: The when, how and why. Eur. Respir. Rev. 2019, 28, 180119. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gottschalk, A.; Sharma, S.; Ford, J.; Durieux, M.E. Review article: The role of the perioperative period in recurrence after cancer surgery. Anesth. Analg. 2010, 110, 1636. [Google Scholar] [CrossRef] [PubMed]
- Horowitz, M.; Neeman, E.; Sharon, E.; Ben-Eliyahu, S. Exploiting the critical perioperative period to improve long-term cancer outcomes. Nat. Rev. Clin. Oncol. 2015, 12, 213–226. [Google Scholar] [CrossRef] [Green Version]
- Khan, S.; Kelly, K.J.; Veerapong, J.; Lowy, A.M.; Baumgartner, J.M. Incidence, Risk Factors, and Prevention Strategies for Venous Thromboembolism after Cytoreductive Surgery and Hyperthermic Intraperitoneal Chemotherapy. Ann. Surg. Oncol. 2019, 26, 2276–2284. [Google Scholar] [CrossRef]
- Blom, J.W.; Vanderschoot JP, M.; Oostindier, M.J.; Osanto, S.; van der Meer, F.J.; Rosendaal, F.R. Incidence of venous thrombosis in a large cohort of 66 329 cancer patients: Results of a record linkage study. J. Thromb. Hemost. 2006, 4, 529–535. [Google Scholar] [CrossRef] [Green Version]
- Hamza, M.S.; Mousa, S.A. Cancer-Associated Thrombosis: Risk Factors, Molecular Mechanisms, Future Management. Clin. Appl. Thromb. Hemost. 2020, 26, 1076029620954282. [Google Scholar] [CrossRef]
- Nasser, N.J.; Fox, J.; Agbarya, A. Potential Mechanisms of Cancer-Related Hypercoagulability. Cancers 2020, 12, 566. [Google Scholar] [CrossRef] [Green Version]
- Alghamdi, M.; Al Ghamdi, K.A.; Khan, R.H.; Uversky, V.N.; Redwan, E.M. An interplay of structure and intrinsic disorder in the functionality of pepti-dylarginine deiminases, a family of key autoimmunity-related enzymes. Cell. Mol. Life Sci. 2019, 76, 4635–4662. [Google Scholar] [CrossRef]
- Anzilotti, C.; Pratesi, F.; Tommasi, C.; Migliorini, P. Peptidylarginine deiminase 4 and citrullination in health and disease. Autoimmun. Rev. 2010, 9, 158–160. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.; Wang, Y. Peptidylarginine deiminases in citrullination, gene regulation, health and pathogenesis. Biochim. Biophys. Acta 2013, 1829, 1126–1135. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clancy, K.W.; Russell, A.-M.; Subramanian, V.; Nguyen, H.; Qian, Y.; Campbell, R.M.; Thompson, P.R. Citrullination/Methylation Crosstalk on Histone H3 Regulates ER-Target Gene Transcription. ACS Chem. Biol. 2017, 12, 1691–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuthbert, G.L.; Daujat, S.; Snowden, A.W.; Erdjument-Bromage, H.; Hagiwara, T.; Yamada, M.; Schneider, R.; Gregory, P.D.; Tempst, P.; Bannister, A.J.; et al. Histone Deimination Antagonizes Arginine Methylation. Cell 2004, 118, 545–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Yao, H.; Zhang, Z.; Li, M.; Luo, Y.; Thompson, P.R.; Gilmour, D.S.; Wang, Y. Regulation of p53 Target Gene Expression by Peptidylarginine Deiminase 4. Mol. Cell. Biol. 2008, 28, 4745–4758. [Google Scholar] [CrossRef] [Green Version]
- Yao, H.; Li, P.; Venters, B.J.; Zheng, S.; Thompson, P.R.; Pugh, B.F.; Wang, Y. Histone Arg Modifications and p53 Regulate the Expression of OKL38, a Mediator of Apoptosis. J. Biol. Chem. 2008, 283, 20060–20068. [Google Scholar] [CrossRef] [Green Version]
- Brinkmann, V.; Reichard, U.; Goosmann, C.; Fauler, B.; Uhlemann, Y.; Weiss, D.S.; Weinrauch, Y.; Zychlinsky, A. Neutrophil extracellular traps kill bacteria. Science 2004, 303, 1532–1535. [Google Scholar] [CrossRef]
- Guimarães-Costa, A.B.; Nascimento MT, C.; Froment, G.S.; Soares, R.P.; Morgado, F.N.; Conceição-Silva, F.; Saraiva, E.M. Leishmania amazonensis promastigotes induce and are killed by neutrophil extracellular traps. Proc. Natl. Acad. Sci. USA 2009, 106, 6748–6753. [Google Scholar] [CrossRef] [Green Version]
- Urban, C.F.; Ermert, D.; Schmid, M.; Abu-Abed, U.; Goosmann, C.; Nacken, W.; Brinkmann, V.; Jungblut, P.R.; Zychlinsky, A. Neutrophil Extracellular Traps Contain Calprotectin, a Cytosolic Protein Complex Involved in Host Defense against Candida albicans. PLoS Pathog. 2009, 5, e1000639. [Google Scholar] [CrossRef] [Green Version]
- Saitoh, T.; Komano, J.; Saitoh, Y.; Misawa, T.; Takahama, M.; Kozaki, T.; Uehata, T.; Iwasaki, H.; Omori, H.; Yamaoka, S.; et al. Neutrophil Extracellular Traps Mediate a Host Defense Response to Human Immunode-ficiency Virus-1. Cell Host Microbe 2012, 12, 109–116. [Google Scholar] [CrossRef]
- Urban, C.F.; Reichard, U.; Brinkmann, V.; Zychlinsky, A. Neutrophil extracellular traps capture and kill Candida albicans yeast and hyphal forms. Cell. Microbiol. 2006, 8, 668–676. [Google Scholar] [CrossRef] [PubMed]
- Kearney, P.L.; Bhatia, M.; Jones, N.G.; Yuan, L.; Glascock, M.C.; Catchings, K.L.; Yamada, M.; Thompson, P.R. Kinetic Characterization of Protein Arginine Deiminase 4: A Transcriptional Core-pressor Implicated in the Onset and Progression of Rheumatoid Arthritis. Biochemistry 2005, 44, 10570–10582. [Google Scholar] [CrossRef]
- Mastronardi, F.G.; Wood, D.D.; Mei, J.; Raijmakers, R.; Tseveleki, V.; Dosch, H.M.; Probert, L.; Casaccia-Bonnefil, P.; Moscarello, M.A. Increased Citrullination of Histone H3 in Multiple Sclerosis Brain and Animal Models of Demyelination: A Role for Tumor Necrosis Factor-Induced Peptidylarginine Deiminase 4 Translocation. J. Neurosci. 2006, 26, 11387–11396. [Google Scholar] [CrossRef] [Green Version]
- Claushuis TA, M.; van der Donk, L.E.H.; Luitse, A.L.; van Veen, H.A.; van der Wel, N.N.; van Vught, L.A.; Roelofs, J.J.T.H.; de Boer, O.J.; Lankelma, J.M.; Boon, L.; et al. Role of Peptidylarginine Deiminase 4 in Neutrophil Extracellular Trap Formation and Host Defense duringKlebsiella pneumoniae–Induced Pneumonia-Derived Sepsis. J. Immunol. 2018, 201, 1241–1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elliott, W.; Guda, M.R.; Asuthkar, S.; Teluguakula, N.; Prasad, D.V.R.; Tsung, A.J.; Velpula, K.K. PAD Inhibitors as a Potential Treatment for SARS-CoV-2 Immunothrombosis. Bio-Med. 2021, 9, 1867. [Google Scholar]
- Wang, Y.; Chen, R.; Gan, Y.; Ying, S. The roles of PAD2- and PAD4-mediated protein citrullination catalysis in cancers. Int. J. Cancer 2021, 148, 267–276. [Google Scholar] [CrossRef]
- Ciesielski, O.; Biesiekierska, M.; Panthu, B.; Soszyński, M.; Pirola, L.; Balcerczyk, A. Citrullination in the pathology of inflammatory and autoimmune disorders: Recent advances and future perspectives. Cell. Mol. Life Sci. 2022, 79, 94. [Google Scholar] [CrossRef]
- Chang, X.; Han, J.; Pang, L.; Zhao, Y.; Yang, Y.; Shen, Z. Increased PADI4 expression in blood and tissues of patients with malignant tumors. BMC Cancer 2009, 9, 40. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Han, J. Expression of peptidylarginine deiminase type 4 (PAD4) in various tumors. Mol. Carcinog. 2005, 45, 183–196. [Google Scholar] [CrossRef]
- Tilvawala, R.; Thompson, P.R. Peptidyl arginine deiminases: Detection and functional analysis of protein citrullination. Curr. Opin. Struct. Biol. 2019, 59, 205–215. [Google Scholar] [CrossRef]
- Fuhrmann, J.; Clancy, K.W.; Thompson, P.R. Chemical Biology of Protein Arginine Modifications in Epigenetic Regulation. Chem. Rev. 2015, 115, 5413–5461. [Google Scholar] [CrossRef] [PubMed]
- Ordóñez, A.; Martínez-Martínez, I.; Corrales, F.J.; Miqueo, C.; Miñano, A.; Vicente, V.; Corral, J. Effect of citrullination on the function and conformation of antithrombin. FEBS J. 2009, 276, 6763–6772. [Google Scholar] [CrossRef] [PubMed]
- Orgován, G.; Noszál, B. The complete microspeciation of arginine and citrulline. J. Pharm. Biomed. Anal. 2011, 54, 965–971. [Google Scholar] [CrossRef] [PubMed]
- Alghamdi, M.; Alasmari, D.; Assiri, A.; Mattar, E.; AlJaddawi, A.A.; Alattas, S.G.; Redwan, E.M. An Overview of the Intrinsic Role of Citrullination in Autoimmune Disorders. J. Immunol. Res. 2019, 2019, 7592851. [Google Scholar] [CrossRef] [Green Version]
- Tarcsa, E.; Marekov, L.N.; Mei, G.; Melino, G.; Lee, S.C.; Steinert, P.M. Protein unfolding by peptidylarginine deiminase. Substrate specificity and structural relationships of the natural substrates trichohyalin and filaggrin. J. Biol. Chem. 1996, 271, 30709–30716. [Google Scholar] [CrossRef] [Green Version]
- György, B.; Tóth, E.; Tarcsa, E.; Falus, A.; Buzás, E.I. Citrullination: A posttranslational modification in health and disease. Int. J. Biochem. Cell Biol. 2006, 38, 1662–1677. [Google Scholar] [CrossRef]
- Lee, C.-Y.; Wang, D.; Wilhelm, M.; Zolg, D.P.; Schmidt, T.; Schnatbaum, K.; Reimer, U.; Pontén, F.; Uhlén, M.; Hahne, H.; et al. Mining the Human Tissue Proteome for Protein Citrullination. Mol. Cell. Proteom. 2018, 17, 1378–1391. [Google Scholar] [CrossRef] [Green Version]
- Li, P.; Wang, D.; Yao, H.; Doret, P.; Hao, G.; Shen, Q.; Qiu, H.; Zhang, X.; Wang, Y.; Chen, G. Coordination of PAD4 and HDAC2 in the regulation of p53-target gene expression. Oncogene 2010, 29, 3153–3162. [Google Scholar] [CrossRef] [Green Version]
- Hagiwara, T.; Nakashima, K.; Hirano, H.; Senshu, T.; Yamada, M. Deimination of Arginine Residues in Nucleophosmin/B23 and Histones in HL-60 Granulocytes. Biochem. Biophys. Res. Commun. 2002, 290, 979–983. [Google Scholar] [CrossRef]
- Mondal, S.; Thompson, P.R. Protein Arginine Deiminases (PADs): Biochemistry and Chemical Biology of Protein Citrullination. Acc. Chem. Res. 2019, 52, 818–832. [Google Scholar] [CrossRef]
- Chavanas, S.; Méchin, M.-C.; Nachat, R.; Adoue, V.; Coudane, F.; Serre, G.; Simon, M. Peptidylarginine deiminases and deimination in biology and pathology: Relevance to skin homeostasis. J. Dermatol. Sci. 2006, 44, 63–72. [Google Scholar] [CrossRef] [PubMed]
- Rus DA, A.; Ikejiri, Y.; Ono, H.; Yonekawa, T.; Shiraiwa, M.; Kawada, A.; Takahara, H. Molecular cloning of cDNAs of mouse peptidylarginine deiminase type I, type III and type IV, and the expression pattern of type I in mouse. Eur. J. Biochem. 1999, 259, 660–669. [Google Scholar]
- Guerrin, M.; Ishigami, A.; Mechin, M.C.; Nachat, R.; Valmary, S.; Sebbag, M.; Simon, M.; Senshu, T.; Serre, G. cDNA cloning, gene organization and expression analysis of human pepti-dylarginine deiminase type I. Biochem. J. 2003, 370 Pt 1, 167–174. [Google Scholar] [CrossRef]
- Qin, H.; Liu, X.; Li, F.; Miao, L.; Li, T.; Xu, B.; An, X.; Muth, A.; Thompson, P.R.; Coonrod, S.A.; et al. PAD1 promotes epithelial-mesenchymal transition and metastasis in triple-negative breast cancer cells by regulating MEK1-ERK1/2-MMP2 signaling. Cancer Lett. 2017, 409, 30–41. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamali, H.; Khan, H.A.; Tjin, C.C.; Ellman, J.A. Cellular Activity of New Small Molecule Protein Arginine Deiminase 3 (PAD3) Inhibitors. ACS Med. Chem. Lett. 2016, 7, 847–851. [Google Scholar] [CrossRef] [Green Version]
- Falcão, A.M.; Meijer, M.; Scaglione, A.; Rinwa, P.; Agirre, E.; Liang, J.; Larsen, S.C.; Heskol, A.; Frawley, R.; Klingener, M.; et al. PAD2-Mediated Citrullination Contributes to Efficient Oligodendrocyte Differen-tiation and Myelination. Cell Rep. 2019, 27, 1090–1102. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hojo-Nakashima, I.; Sato, R.; Nakashima, K.; Hagiwara, T.; Yamada, M. Dynamic Expression of Peptidylarginine Deiminase 2 in Human Monocytic Leukaemia THP-1 Cells During Macrophage Differentiation. J. Biochem. 2009, 146, 471–479. [Google Scholar] [CrossRef]
- Vossenaar, E.R. Expression and activity of citrullinating peptidylarginine deiminase enzymes in monocytes and macrophages. Ann. Rheum. Dis. 2004, 63, 373–381. [Google Scholar] [CrossRef] [Green Version]
- Asaga, H.; Nakashima, K.; Senshu, T.; Ishigami, A.; Yamada, M. Immunocytochemical localization of peptidylarginine deiminase in human eosinophils and neutrophils. J. Leukoc. Biol. 2001, 70, 46–51. [Google Scholar] [CrossRef]
- Nakashima, K.; Hagiwara, T.; Yamada, M. Nuclear Localization of Peptidylarginine Deiminase V and Histone Deimination in Granulocytes. J. Biol. Chem. 2002, 277, 49562–49568. [Google Scholar] [CrossRef] [Green Version]
- Vossenaar, E.R.; Zendman AJ, W.; van Venrooij, W.J.; Pruijn, G.J. PAD, a growing family of citrullinating enzymes: Genes, features and involvement in disease. BioEssays 2003, 25, 1106–1118. [Google Scholar] [CrossRef] [PubMed]
- Guo, Q.; Fast, W. Citrullination of Inhibitor of Growth 4 (ING4) by Peptidylarginine Deminase 4 (PAD4) Disrupts the Interaction between ING4 and p53. J. Biol. Chem. 2011, 286, 17069–17078. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Coonrod, S.A.; Kraus, W.L.; Jelinek, M.A.; Stallcup, M.R. Regulation of coactivator complex assembly and function by protein arginine meth-ylation and demethylimination. Proc. Natl. Acad. Sci. USA 2005, 102, 3611–3616. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zawrotniak, M.; Rapala-Kozik, M. Neutrophil extracellular traps (NETs)—Formation and implications. Acta Biochim. Pol. 2013, 60, 277–284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, L.; Nagar, M.; Maurais, A.J.; Slade, D.J.; Parelkar, S.S.; Coonrod, S.A.; Weerapana, E.; Thompson, P.R. Calcium Regulates the Nuclear Localization of Protein Arginine Deiminase 2. Biochemistry 2019, 58, 3042–3056. [Google Scholar] [CrossRef]
- Zhang, X.; Bolt, M.; Guertin, M.J.; Chen, W.; Zhang, S.; Cherrington, B.D.; Slade, D.J.; Dreyton, C.J.; Subramanian, V.; Bicker, K.L.; et al. Peptidylarginine deiminase 2-catalyzed histone H3 arginine 26 citrullination facilitates estrogen receptor α target gene activation. Proc. Natl. Acad. Sci. USA 2012, 109, 13331–13336. [Google Scholar] [CrossRef] [Green Version]
- Esposito, G.; Vitale, A.M.; Leijten FP, J.; Strik, A.M.; Koonen-Reemst, A.M.; Yurttas, P.; Robben, T.J.; Coonrod, S.; Gossen, J.A. Peptidylarginine deiminase (PAD) 6 is essential for oocyte cytoskeletal sheet formation and female fertility. Mol. Cell. Endocrinol. 2007, 273, 25–31. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Shi, Y.; Fu, J.; Yu, M.; Feng, R.; Sang, Q.; Liang, B.; Chen, B.; Qu, R.; Li, B.; et al. Mutations in PADI6 Cause Female Infertility Characterized by Early Embryonic Arrest. Am. J. Hum. Genet. 2016, 99, 744–752. [Google Scholar] [CrossRef] [Green Version]
- Arita, K.; Hashimoto, H.; Shimizu, T.; Nakashima, K.; Yamada, M.; Sato, M. Structural Basis for Ca(2+)-Induced Activation of Human PAD4. Nat. Struct. Mol. Biol. 2004, 11, 777–783. [Google Scholar] [CrossRef]
- Ishida-Yamamoto, A.; Senshu, T.; Takahashi, H.; Akiyama, K.; Nomura, K.; Iizuka, H. Decreased Deiminated Keratin K1 in Psoriatic Hyperproliferative Epidermis. J. Investig. Dermatol. 2000, 114, 701–705. [Google Scholar] [CrossRef] [Green Version]
- Musse, A.A.; Li, Z.; Ackerley, C.A.; Bienzle, D.; Lei, H.; Poma, R.; Harauz, G.; Moscarello, M.A.; Mastronardi, F.G. Peptidylarginine deiminase 2 (PAD2) overexpression in transgenic mice leads to myelin loss in the central nervous system. Dis. Model. Mech. 2008, 1, 229–240. [Google Scholar] [CrossRef] [PubMed]
- Carrillo-Vico, A.; Leech, M.D.; Anderton, S.M. Contribution of Myelin Autoantigen Citrullination to T Cell Autoaggression in the Central Nervous System. J. Immunol. 2010, 184, 2839–2846. [Google Scholar] [CrossRef] [PubMed]
- Asaga, H.; Ishigami, A. Protein deimination in the rat brain after kainate administration: Citrulline-containing proteins as a novel marker of neurodegeneration. Neurosci. Lett. 2001, 299, 5–8. [Google Scholar] [CrossRef]
- Asaga, H.; Akiyama, K.; Ohsawa, T.; Ishigami, A. Increased and type II-specific expression of peptidylarginine deiminase in activated microglia but not hyperplastic astrocytes following kainic acid-evoked neurodegeneration in the rat brain. Neurosci. Lett. 2002, 326, 129–132. [Google Scholar] [CrossRef]
- van Beers, J.J.; Raijmakers, R.; Alexander, L.; Stammen-Vogelzangs, J.; Lokate, A.M.; Heck, A.J.; Schasfoort, R.B.; Pruijn, G.J. Mapping of citrullinated fibrinogen B-cell epitopes in rheumatoid arthritis by imaging surface plasmon resonance. Arthritis Res. Ther. 2010, 12, R219. [Google Scholar] [CrossRef] [Green Version]
- Kinloch, A.; Lundberg, K.; Wait, R.; Wegner, N.; Lim, N.H.; Zendman, A.J.W.; Saxne, T.; Malmstr, V.; Venables, P.J. Synovial fluid is a site of citrullination of autoantigens in inflammatory arthritis. Arthritis Care Res. 2008, 58, 2287–2295. [Google Scholar] [CrossRef]
- Jang, B.; Ishigami, A.; Maruyama, N.; Carp, R.I.; Kim, Y.-S.; Choi, E.-K. Peptidylarginine deiminase and protein citrullination in prion diseases. Prion 2013, 7, 42–46. [Google Scholar] [CrossRef] [Green Version]
- Jang, B.; Kim, E.; Choi, J.-K.; Jin, J.-K.; Kim, J.-I.; Ishigami, A.; Maruyama, N.; Carp, R.I.; Kim, Y.-S.; Choi, E.-K. Accumulation of Citrullinated Proteins by Up-Regulated Peptidylarginine Deiminase 2 in Brains of Scrapie-Infected Mice: A Possible Role in Pathogenesis. Am. J. Pathol. 2008, 173, 1129–1142. [Google Scholar] [CrossRef] [Green Version]
- Chavanas, S.; Adoue, V.; Méchin, M.-C.; Ying, S.; Dong, S.; Duplan, H.; Charveron, M.; Takahara, H.; Serre, G.; Simon, M. Long-Range Enhancer Associated with Chromatin Looping Allows AP-1 Regulation of the Peptidylarginine Deiminase 3 Gene in Differentiated Keratinocyte. PLoS ONE 2008, 3, e3408. [Google Scholar] [CrossRef] [Green Version]
- Basmanav, F.B.; Cau, L.; Tafazzoli, A.; Méchin, M.C.; Wolf, S.; Romano, M.T.; Valentin, F.; Wiegmann, H.; Huchenq, A.; Kandil, R. Mutations in Three Genes Encoding Proteins Involved in Hair Shaft Formation Cause Uncombable Hair Syndrome. Am. J. Hum. Genet. 2016, 99, 1292–1304. [Google Scholar] [CrossRef] [Green Version]
- Uysal-Onganer, P.; MacLatchy, A.; Mahmoud, R.; Kraev, I.; Thompson, P.R.; Inal, J.M.; Lange, S. Peptidylarginine Deiminase Isozyme-Specific PAD2, PAD3 and PAD4 Inhibitors Differentially Modulate Extracellular Vesicle Signatures and Cell Invasion in Two Glioblastoma Multiforme Cell Lines. Int. J. Mol. Sci. 2020, 21, 1495. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vossenaar, E.R.; Zendman, A.J.; van Venrooij, W.J. Citrullination, a possible functional link between susceptibility genes and rheumatoid arthritis. Arthritis Res. Ther. 2004, 6, 107. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.; Yamada, R.; Suzuki, A.; Sawada, T.; Yoshino, S.; Tokuhiro, S.; Yamamoto, K. Localization of peptidylarginine deiminase 4 (PADI4) and citrullinated protein in synovial tissue of rheumatoid arthritis. Rheumatology 2004, 44, 40–50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Acharya, N.K.; Nagele, E.P.; Han, M.; Coretti, N.J.; DeMarshall, C.; Kosciuk, M.C.; Boulos, P.A.; Nagele, R.G. Neuronal PAD4 expression and protein citrullination: Possible role in production of autoantibodies associated with neurodegenerative disease. J. Autoimmun. 2012, 38, 369–380. [Google Scholar] [CrossRef]
- Chen, C.C.; Isomoto, H.; Narumi, Y.; Sato, K.; Oishi, Y.; Kobayashi, T.; Yanagihara, K.; Mizuta, Y.; Kohno, S.; Tsukamoto, K. Haplotypes of PADI4 susceptible to rheumatoid arthritis are also associated with ulcerative colitis in the Japanese population. Clin. Immunol. 2008, 126, 165–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raup-Konsavage, W.M.; Wang, Y.; Wang, W.W.; Feliers, D.; Ruan, H.; Reeves, W.B. Neutrophil peptidyl arginine deiminase-4 has a pivotal role in ische-mia/reperfusion-induced acute kidney injury. Kidney Int. 2018, 93, 365–374. [Google Scholar] [CrossRef]
- Costa, N.A.; Polegato, B.F.; Pereira, A.G.; Paiva, S.A.R.; Gut, A.L.; Balbi, A.L.; Ponce, D.; Zornoff, L.A.M.; Azevedo, P.S.; Minicucci, M.F. Evaluation of peptidylarginine deiminase 4 and PADI4 polymorphisms in sep-sis-induced acute kidney injury. Rev. Da Assoc. Médica Bras. 2020, 66, 1515–1520. [Google Scholar] [CrossRef]
- Veras, F.P.; Pontelli, M.C.; Silva, C.M.; Toller-Kawahisa, J.E.; de Lima, M.; Nascimento, D.C.; Schneider, A.H.; Caetité, D.; Tavares, L.A.; Paiva, I.M. SARS-CoV-2–triggered neutrophil extracellular traps mediate COVID-19 pathology. J. Exp. Med. 2020, 217, e20201129. [Google Scholar] [CrossRef]
- Qian, J.; Nguyen, N.M.P.; Rezaei, M.; Huang, B.; Tao, Y.; Zhang, X.; Cheng, Q.; Yang, H.; Asangla, A.; Majewski, J.; et al. Biallelic PADI6 variants linking infertility, miscarriages, and hydatidiform moles. Eur. J. Hum. Genet. 2018, 26, 1007–1013. [Google Scholar] [CrossRef] [Green Version]
- Mohanan, S.; Cherrington, B.D.; Horibata, S.; McElwee, J.L.; Thompson, P.R.; Coonrod, S.A. Potential Role of Peptidylarginine Deiminase Enzymes and Protein Citrul-lination in Cancer Pathogenesis. Biochem. Res. Int. 2012, 2012, 895343. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Xu, B.; Zhang, X.; Zheng, Y.; Zhao, Y.; Chang, X. PADI2 gene confers susceptibility to breast cancer and plays tumorigenic role via ACSL4, BINC3 and CA9 signaling. Cancer Cell Int. 2016, 16, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, W.; Zheng, Y.; Xu, B.; Ma, F.; Li, C.; Zhang, X.; Wang, Y.; Chang, X. Investigating the expression, effect and tumorigenic pathway of PADI2 in tumors. Oncol. Targets Ther. 2017, 10, 1475–1485. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, G.; Zhang, X.; Feng, T.; Pan, J.; Chen, W.; Yang, M.; Bai, X.; Pang, Y.; Yu, J.; et al. PADI2-Mediated Citrullination Promotes Prostate Cancer Progression. Cancer Res. 2017, 77, 5755–5768. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cantariño, N.; Musulén, E.; Valero, V.; Peinado, M.A.; Perucho, M.; Moreno, V.; Forcales, S.V.; Douet, J.; Buschbeck, M. Downregulation of the Deiminase PADI2 Is an Early Event in Colorectal Carcino-genesis and Indicates Poor Prognosis. Mol. Cancer Res. 2016, 14, 841–848. [Google Scholar] [CrossRef] [Green Version]
- Yuzhalin, A.E. Citrullination in Cancer. Cancer Res. 2019, 79, 1274–1284. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yuzhalin, A.E.; Gordon-Weeks, A.N.; Tognoli, M.L.; Jones, K.; Markelc, B.; Konietzny, R.; Fischer, R.; Muth, A.; O’Neill, E.; Thompson, P.R. Colorectal cancer liver metastatic growth depends on PAD4-driven citrullination of the extracellular matrix. Nat. Commun. 2018, 9, 4783. [Google Scholar] [CrossRef] [Green Version]
- Demers, M.; Wagner, D.D. NETosis: A New Factor in Tumor Progression and Cancer-Associated Thrombosis. Semin. Thromb. Hemost. 2014, 40, 277–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cools-Lartigue, J.; Spicer, J.; Najmeh, S.; Ferri, L. Neutrophil extracellular traps in cancer progression. Cell. Mol. Life Sci. 2014, 71, 4179–4194. [Google Scholar] [CrossRef] [PubMed]
- Stadler, S.C.; Vincent, C.T.; Fedorov, V.D.; Patsialou, A.; Cherrington, B.D.; Wakshlag, J.J.; Mohanan, S.; Zee, B.M.; Zhang, X.; Garcia, B.A.; et al. Dysregulation of PAD4-mediated citrullination of nuclear GSK3β activates TGF-β signaling and induces epithelialto-mesenchymal transition in breast cancer cells. Proc. Natl. Acad. Sci. USA 2013, 110, 16283. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Q.; Song, C.; Liu, X.; Qin, H.; Miao, L.; Zhang, X. Peptidylarginine deiminase 4 overexpression resensitizes MCF-7/ADR breast cancer cells to adriamycin via GSK3β/p53 activation. Cancer Manag. Res. 2019, 11, 625–636. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.; Arfman, T.; Wichapong, K.; Reutelingsperger, C.P.M.; Voorberg, J.; Nicolaes, G.A.F. PAD4 takes charge during neutrophil activation: Impact of PAD4 mediated NET for-mation on immune-mediated disease. J. Thromb. Haemost. 2021, 19, 1607–1617. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-L.; Chiang, Y.-H.; Liu, G.-Y.; Hung, H.-C. Functional Role of Dimerization of Human Peptidylarginine Deiminase 4 (PAD4). PLoS ONE 2011, 6, e21314. [Google Scholar] [CrossRef] [PubMed]
- Teo, C.Y.; Shave, S.; Thean Chor, A.L.; Salleh, A.B.; Rahman, M.B.; Walkinshaw, M.D.; Tejo, B.A. Discovery of a new class of inhibitors for the protein arginine deiminase type 4 (PAD4) by structure-based virtual screening. BMC Bioinform. 2012, 13 (Suppl. S17), S4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jones, J.E.; Causey, C.P.; Knuckley, B.; Slack-Noyes, J.L.; Thompson, P.R. Protein arginine deiminase 4 (PAD4): Current understanding and future therapeutic potential. Curr. Opin. Drug Discov. Dev. 2009, 12, 616. [Google Scholar]
- Darrah, E.; Giles, J.T.; Ols, M.L.; Bull, H.G.; Andrade, F.; Rosen, A. Erosive Rheumatoid Arthritis Is Associated with Antibodies That Activate PAD4 by In-creasing Calcium Sensitivity. Sci. Transl. Med. 2013, 5, 165–186. [Google Scholar] [CrossRef] [Green Version]
- Tour, O.; Adams, S.R.; Kerr, R.A.; Meijer, R.M.; Sejnowski, T.J.; Tsien, R.W.; Tsien, R.Y. Calcium Green FlAsH as a genetically targeted small-molecule calcium indicator. Nat. Chem. Biol. 2007, 3, 423–431. [Google Scholar] [CrossRef] [Green Version]
- Slade, D.J.; Fang, P.; Dreyton, C.J.; Zhang, Y.; Fuhrmann, J.; Rempel, D.; Bax, B.D.; Coonrod, S.A.; Lewis, H.D.; Guo, M.; et al. Protein arginine deiminase 2 binds calcium in an ordered fashion: Implications for inhibitor design. ACS Chem. Biol. 2015, 10, 1043–1053. [Google Scholar] [CrossRef] [Green Version]
- Knuckley, B.; Bhatia, M.; Thompson, P.R. Protein Arginine Deiminase 4: Evidence for a Reverse Protonation Mechanism. Biochemistry 2007, 46, 6578–6587. [Google Scholar] [CrossRef] [Green Version]
- Zhu, D.; Zhang, Y.; Wang, S. Histone citrullination: A new target for tumors. Mol. Cancer 2021, 20, 90. [Google Scholar] [CrossRef]
- Zhai, Q.; Wang, L.; Zhao, P.; Li, T. Role of citrullination modification catalyzed by peptidylarginine deiminase 4 in gene tran-scriptional regulation. Acta Biochim. Biophys. Sin. 2017, 49, 567. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Wysocka, J.; Sayegh, J.; Lee, Y.-H.; Perlin, J.R.; Leonelli, L.; Sonbuchner, L.S.; McDonald, C.H.; Cook, R.G.; Dou, Y.; et al. Human PAD4 Regulates Histone Arginine Methylation Levels via Demethylimination. Science 2004, 306, 279–283. [Google Scholar] [CrossRef] [PubMed]
- Klose, R.J.; Zhang, Y. Regulation of histone methylation by demethylimination and demethylation. Nat. Rev. Mol. Cell Biol. 2007, 8, 307–318. [Google Scholar] [CrossRef] [PubMed]
- Kolodziej, S.; Kuvardina, O.N.; Oellerich, T.; Herglotz, J.; Backert, I.; Kohrs, N.; Buscató, E.; Wittmann, S.K.; Salinas-Riester, G.; Bonig, H.; et al. PADI4 acts as a coactivator of Tal1 by counteracting repressive histone arginine methylation. Nat. Commun. 2014, 5, 3995. [Google Scholar] [CrossRef] [PubMed]
- Sharma, P.; Azebi, S.; England, P.; Christensen, T.; Møller-Larsen, A.; Petersen, T.; Batsché, E.; Muchardt, C. Citrullination of Histone H3 Interferes with HP1-Mediated Transcriptional Repression. PLoS Genet. 2012, 8, e1002934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Denis, H.; Deplus, R.; Putmans, P.; Yamada, M.; Métivier, R.; Fuks, F. Functional Connection between Deimination and Deacetylation of Histones. Mol. Cell. Biol. 2009, 29, 4982–4993. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; David, B.T.; Trawczynski, M.; Fessler, R.G. Advances in Pluripotent Stem Cells: History, Mechanisms, Technologies, and Applications. Stem Cell Rev. Rep. 2020, 16, 3–32. [Google Scholar] [CrossRef] [Green Version]
- Barati, M.; Akhondi, M.; Mousavi, N.S.; Haghparast, N.; Ghodsi, A.; Baharvand, H.; Ebrahimi, M.; Hassani, S.-N. Pluripotent Stem Cells: Cancer Study, Therapy, and Vaccination. Stem Cell Rev. Rep. 2021, 17, 1975–1992. [Google Scholar] [CrossRef]
- Christophorou, M.A.; Castelo-Branco, G.; Halley-Stott, R.P.; Oliveira, C.S.; Loos, R.; Radzisheuskaya, A.; Mowen, K.A.; Bertone, P.; Silva, J.C.; Zernicka-Goetz, M.; et al. Citrullination regulates pluripotency and histone H1 binding to chromatin. Nature 2014, 507, 104–108. [Google Scholar] [CrossRef] [Green Version]
- Song, G.; Shi, L.; Guo, Y.; Yu, L.; Wang, L.; Zhang, X.; Li, L.; Han, Y.; Ren, X.; Guo, Q.; et al. A novel PAD4/SOX4/PU.1 signaling pathway is involved in the committed differentiation of acute promyelocytic leukemia cells into granulocytic cells. Oncotarget 2015, 7, 3144–3157. [Google Scholar] [CrossRef] [Green Version]
- Slack, J.L.; Causey, C.P.; Thompson, P.R. Protein arginine deiminase 4: A target for an epigenetic cancer therapy. Cell. Mol. Life Sci. 2011, 68, 709–720. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Li, P.; Wang, S.; Hu, J.; Chen, X.A.; Wu, J.; Fisher, M.; Oshaben, K.; Zhao, N.; Gu, Y.; et al. Anticancer Peptidylarginine Deiminase (PAD) Inhibitors Regulate the Autophagy Flux and the Mammalian Target of Rapamycin Complex 1 Activity. J. Biol. Chem. 2012, 287, 25941–25953. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanikawa, C.; Espinosa, M.; Suzuki, A.; Masuda, K.; Yamamoto, K.; Tsuchiya, E.; Ueda, K.; Daigo, Y.; Nakamura, Y.; Matsuda, K. Regulation of histone modification and chromatin structure by the p53–PADI4 pathway. Nat. Commun. 2012, 3, 676. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, G.Y.; Liao, Y.F.; Chang, W.H.; Liu, C.C.; Hsieh, M.C.; Hsu, P.C.; Tsay, G.J.; Hung, H.C. Overexpression of peptidylarginine deiminase IV features in apoptosis of haematopoietic cells. Apoptosis 2006, 11, 183–196. [Google Scholar] [CrossRef]
- Tsopanoglou, N.E.; Maragoudakis, M.E. On the Mechanism of Thrombin-induced Angiogenesis. J. Biol. Chem. 1999, 274, 23969–23976. [Google Scholar] [CrossRef] [Green Version]
- De Meyer, S.F.; Savchenko, A.S.; Haas, M.S.; Schatzberg, D.; Carroll, M.C.; Schiviz, A.; Dietrich, B.; Rottensteiner, H.; Scheiflinger, F.; Wagner, D.D. Protective anti-inflammatory effect of ADAMTS13 on myocardial is-chemia/reperfusion injury in mice. Blood 2012, 120, 5217–5223. [Google Scholar] [CrossRef]
- Sorvillo, N.; Mizurini, D.M.; Coxon, C.; Martinod, K.; Tilvawala, R.; Cherpokova, D.; Salinger, A.J.; Seward, R.J.; Staudinger, C.; Weerapana, E.; et al. Plasma Peptidylarginine Deiminase IV Promotes VWF-Platelet String For-mation and Accelerates Thrombosis After Vessel Injury. Circ. Res. 2019, 125, 507–519. [Google Scholar] [CrossRef] [PubMed]
- Saxena, K.; Jolly, M.K.; Balamurugan, K. Hypoxia, partial EMT and collective migration: Emerging culprits in metastasis. Transl. Oncol. 2020, 13, 100845. [Google Scholar] [CrossRef]
- Mancinelli, R.; Carpino, G.; Petrungaro, S.; Mammola, C.L.; Tomaipitinca, L.; Filippini, A.; Facchiano, A.; Ziparo, E.; Giampietri, C. Multifaceted Roles of GSK-3 in Cancer and Autophagy-Related Diseases. Oxidative Med. Cell. Longev. 2017, 2017, 4629495. [Google Scholar] [CrossRef] [Green Version]
- Zhou, B.P.; Deng, J.; Xia, W.; Xu, J.; Li, Y.M.; Gunduz, M.; Hung, M.C. Dual regulation of Snail by GSK-3β-mediated phosphorylation in control of epithelial–mesenchymal transition. Nat. Cell Biol. 2004, 6, 931–940. [Google Scholar] [CrossRef]
- Kawahara, T.; Shareef, H.K.; Aljarah, A.K.; Ide, H.; Li, Y.; Kashiwagi, E.; Netto, G.J.; Zheng, Y.; Miyamoto, H. ELK1 is up-regulated by androgen in bladder cancer cells and promotes tumor progression. Oncotarget 2015, 6, 29860–29876. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.; Gamble, M.J.; Stadler, S.; Cherrington, B.D.; Causey, C.P.; Thompson, P.R.; Roberson, M.S.; Kraus, W.L.; Coonrod, S.A. Genome-Wide Analysis Reveals PADI4 Cooperates with Elk-1 to Activate c-Fos Expression in Breast Cancer Cells. PLoS Genet. 2011, 7, e1002112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Duan, Q.; Pang, C.; Chang, N.; Zhang, J.; Liu, W. Overexpression of PAD4 suppresses drug resistance of NSCLC cell lines to gefitinib through inhibiting Elk1-mediated epithelial-mesenchymal transition. Oncol. Rep. 2016, 36, 551–558. [Google Scholar] [CrossRef] [Green Version]
- Tsai, J.H.; Donaher, J.L.; Murphy, D.A.; Chau, S.; Yang, J. Spatiotemporal Regulation of Epithelial-Mesenchymal Transition Is Essential for Squamous Cell Carcinoma Metastasis. Cancer Cell 2012, 22, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Tsai, J.H.; Yang, J. Epithelial–mesenchymal plasticity in carcinoma metastasis. Genes Dev. 2013, 27, 2192–2206. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Yamada, R.; Suzuki, A.; Kochi, Y.; Sawada, T.; Yamamoto, K. Citrullination of fibronectin in rheumatoid arthritis synovial tissue. Rheumatology 2005, 44, 1374–1382. [Google Scholar] [CrossRef] [Green Version]
- Stefanelli, V.L.; Choudhury, S.; Hu, P.; Liu, Y.; Schwenzer, A.; Yeh, C.R.; Chambers, D.M.; Pesson, K.; Li, W.; Segura, T.; et al. Citrullination of fibronectin alters integrin clustering and focal adhesion stability promoting stromal cell invasion. Matrix Biol. 2019, 82, 86–104. [Google Scholar] [CrossRef]
- Tanikawa, C.; Ueda, K.; Nakagawa, H.; Yoshida, N.; Nakamura, Y.; Matsuda, K. Regulation of Protein Citrullination through p53/PADI4 Network in DNA Damage Response. Cancer Res. 2009, 69, 8761–8769. [Google Scholar] [CrossRef] [Green Version]
- Guo, Q.; Bedford, M.T.; Fast, W. Discovery of peptidylarginine deiminase-4 substrates by protein array: Antagonistic citrullination and methylation of human ribosomal protein S2. Mol. BioSyst. 2011, 7, 2286–2295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deplus, R.; Denis, H.; Putmans, P.; Calonne, E.; Fourrez, M.; Yamamoto, K.; Suzuki, A.; Fuks, F. Citrullination of DNMT3A by PADI4 regulates its stability and controls DNA methylation. Nucleic Acids Res. 2014, 42, 8285–8296. [Google Scholar] [CrossRef] [Green Version]
- Tanikawa, C.; Ueda, K.; Suzuki, A.; Iida, A.; Nakamura, R.; Atsuta, N.; Tohnai, G.; Sobue, G.; Saichi, N.; Momozawa, Y. Citrullination of RGG Motifs in FET Proteins by PAD4 Regulates Protein Aggregation and ALS Susceptibility. Cell Rep. 2018, 22, 1473–1483. [Google Scholar] [CrossRef] [Green Version]
- Andrade, F.; Darrah, E.; Gucek, M.; Cole, R.N.; Rosen, A.; Zhu, X. Autocitrullination of human peptidyl arginine deiminase type 4 regulates protein citrullination during cell activation. Arthritis Care Res. 2010, 62, 1630–1640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Slack, J.L.; Jones, L.E.; Bhatia, M.M.; Thompson, P.R. Autodeimination of Protein Arginine Deiminase 4 Alters Protein–Protein Interactions but Not Activity. Biochemistry 2011, 50, 3997–4010. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Wichapong, K.; Lamers, S.; Reutelingsperger, C.P.; Nicolaes, G.A. Autocitrullination of PAD4 does not alter its enzymatic activity: In vitro and in silico studies. Int. J. Biochem. Cell Biol. 2021, 134, 105938. [Google Scholar] [CrossRef]
- Amulic, B.; Cazalet, C.; Hayes, G.L.; Metzler, K.D.; Zychlinsky, A. Neutrophil Function: From Mechanisms to Disease. Annu. Rev. Immunol. 2012, 30, 459–489. [Google Scholar] [CrossRef]
- Scapini, P.; Cassatella, M.A. Social networking of human neutrophils within the immune system. Blood 2014, 124, 710–719. [Google Scholar] [CrossRef]
- Mantovani, A.; Cassatella, M.A.; Costantini, C.; Jaillon, S. Neutrophils in the activation and regulation of innate and adaptive immunity. Nat. Rev. Immunol. 2011, 11, 519–531. [Google Scholar] [CrossRef] [PubMed]
- DeLeo, F.R.; Allen, L.H. Phagocytosis and neutrophil extracellular traps. Fac. Rev. 2020, 9, 25. [Google Scholar] [CrossRef]
- Branzk, N.; Lubojemska, A.; Hardison, S.E.; Wang, Q.; Gutierrez, M.G.; Brown, G.D.; Papayannopoulos, V. Neutrophils sense microbe size and selectively release neutrophil extra-cellular traps in response to large pathogens. Nat. Immunol. 2014, 15, 1017–1025. [Google Scholar] [CrossRef] [Green Version]
- Burgener, S.S.; Schroder, K. Neutrophil Extracellular Traps in Host Defense. Cold Spring Harb. Perspect. Biol. 2020, 12, a37028. [Google Scholar] [CrossRef]
- Delgado-Rizo, V.; Martínez-Guzmán, M.A.; Iñiguez-Gutierrez, L.; García-Orozco, A.; Alvarado-Navarro, A.; Fafutis-Morris, M. Neutrophil Extracellular Traps and Its Implications in Inflammation: An Overview. Front. Immunol. 2017, 8, 81. [Google Scholar] [CrossRef] [Green Version]
- Bronze-Da-Rocha, E.; Santos-Silva, A. Neutrophil Elastase Inhibitors and Chronic Kidney Disease. Int. J. Biol. Sci. 2018, 14, 1343–1360. [Google Scholar] [CrossRef] [Green Version]
- Takei, H.; Araki, A.; Watanabe, H.; Ichinose, A.; Sendo, F. Rapid killing of human neutrophils by the potent activator phorbol 12-myristate 13-acetate (PMA) accompanied by changes different from typical apoptosis or necrosis. J. Leukoc. Biol. 1996, 59, 229–240. [Google Scholar] [CrossRef]
- Garcia-Romo, G.S.; Caielli, S.; Vega, B.; Connolly, J.; Allantaz, F.; Xu, Z.; Punaro, M.; Baisch, J.; Guiducci, C.; Coffman, R.L.; et al. Netting Neutrophils Are Major Inducers of Type I IFN Production in Pediatric Systemic Lupus Erythematosus. Sci. Transl. Med. 2011, 3, 73ra20. [Google Scholar] [CrossRef] [PubMed]
- Behnen, M.; Leschczyk, C.; Möller, S.; Batel, T.; Klinger, M.; Solbach, M.D.W.; Laskay, T. Immobilized Immune Complexes Induce Neutrophil Extracellular Trap Release by Human Neutrophil Granulocytes via FcγRIIIB and Mac-1. J. Immunol. 2014, 193, 1954–1965. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Warnatsch, A.; Ioannou, M.; Wang, Q.; Papayannopoulos, V. Neutrophil extracellular traps license macrophages for cytokine production in atherosclerosis. Science 2015, 349, 316–320. [Google Scholar] [CrossRef] [Green Version]
- Neeli, I.; Radic, M. Opposition between PKC isoforms regulates histone deimination and neutrophil extracellular chromatin release. Front. Immunol. 2013, 4, 38. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, T.A.; Abed, U.; Goosmann, C.; Hurwitz, R.; Schulze, I.; Wahn, V.; Weinrauch, Y.; Brinkmann, V.; Zychlinsky, A. Novel cell death program leads to neutrophil extracellular traps. J. Cell Biol. 2007, 176, 231–241. [Google Scholar] [CrossRef] [PubMed]
- Hakkim, A.; Fuchs, T.A.; Martinez, N.E.; Hess, S.; Prinz, H.; Zychlinsky, A.; Waldmann, H. Activation of the Raf-MEK-ERK pathway is required for neutrophil extra-cellular trap formation. Nat. Chem. Biol. 2011, 7, 75–77. [Google Scholar] [CrossRef] [PubMed]
- Sollberger, G.; Tilley, D.O.; Zychlinsky, A. Neutrophil Extracellular Traps: The Biology of Chromatin Externalization. Dev. Cell 2018, 44, 542–553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yipp, B.G.; Kubes, P. NETosis: How vital is it? Blood 2013, 122, 2784–2794. [Google Scholar] [CrossRef]
- Konig, M.F.; Andrade, F. A Critical Reappraisal of Neutrophil Extracellular Traps and NETosis Mimics Based on Differ-ential Requirements for Protein Citrullination. Front. Immunol. 2016, 7, 461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masuda, S.; Nakazawa, D.; Shida, H.; Miyoshi, A.; Kusunoki, Y.; Tomaru, U.; Ishizu, A. NETosis markers: Quest for specific, objective, and quantitative markers. Clin. Chim. Acta 2016, 459, 89–93. [Google Scholar] [CrossRef] [PubMed]
- Clark, S.R.; Ma, A.C.; Tavener, S.A.; McDonald, B.; Goodarzi, Z.; Kelly, M.M.; Patel, K.D.; Chakrabarti, S.; McAvoy, E.; Sinclair, G.D.; et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat. Med. 2007, 13, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Yipp, B.G.; Petri, B.; Salina, D.; Jenne, C.N.; Scott, B.N.; Zbytnuik, L.D.; Pittman, K.; Asaduzzaman, M.; Wu, K.; Meijndert, H.C.; et al. Infection-induced NETosis is a dynamic process involving neutrophil multitasking in vivo. Nat. Med. 2012, 18, 1386–1393. [Google Scholar] [CrossRef] [Green Version]
- Byrd, A.S.; O’Brien, X.M.; Johnson, C.M.; Lavigne, L.M.; Reichner, J.S. An extracellular matrix-based mechanism of rapid neutrophil extracellular trap formation in response to Candida albicans. J. Immunol. 2013, 190, 4136–4148. [Google Scholar] [CrossRef] [Green Version]
- Carestia, A.; Kaufman, T.; Rivadeneyra, L.; Landoni, V.I.; Pozner, R.G.; Negrotto, S.; D’Atri, L.P.; Gómez, R.M.; Schattner, M. Mediators and molecular pathways involved in the regulation of neutrophil extracellular trap formation mediated by activated platelets. J. Leukoc. Biol. 2015, 99, 153–162. [Google Scholar] [CrossRef] [Green Version]
- Jorch, S.K.; Kubes, P. An emerging role for neutrophil extracellular traps in noninfectious disease. Nat. Med. 2017, 23, 279–287. [Google Scholar] [CrossRef]
- Pilsczek, F.H.; Salina, D.; Poon, K.K.H.; Fahey, C.; Yipp, B.G.; Sibley, C.D.; Robbins, S.M.; Green, F.H.; Surette, M.G.; Sugai, M.; et al. A Novel Mechanism of Rapid Nuclear Neutrophil Extracellular Trap Formation in Response to Staphylococcus aureus. J. Immunol. 2010, 185, 7413–7425. [Google Scholar] [CrossRef] [Green Version]
- Yousefi, S.; Mihalache, C.; Kozlowski, E.; Schmid, I.; Simon, H.U. Viable neutrophils release mitochondrial DNA to form neutrophil extra-cellular traps. Cell Death Differ. 2009, 16, 1438–1444. [Google Scholar] [CrossRef]
- Thiam, H.R.; Wong, S.L.; Wagner, D.D.; Waterman, C.M. Cellular Mechanisms of NETosis. Annu. Rev. Cell Dev. Biol. 2020, 36, 191–218. [Google Scholar] [CrossRef]
- Papayannopoulos, V.; Metzler, K.D.; Hakkim, A.; Zychlinsky, A. Neutrophil elastase and myeloperoxidase regulate the formation of neutrophil extracellular traps. J. Cell Biol. 2010, 191, 677–691. [Google Scholar] [CrossRef] [Green Version]
- Metzler, K.D.; Goosmann, C.; Lubojemska, A.; Zychlinsky, A.; Papayannopoulos, V. A Myeloperoxidase-Containing Complex Regulates Neutrophil Elastase Release and Actin Dynamics during NETosis. Cell Rep. 2014, 8, 883–896. [Google Scholar] [CrossRef] [Green Version]
- Kenny, E.F.; Herzig, A.; Krüger, R.; Muth, A.; Mondal, S.; Thompson, P.R.; Brinkmann, V.; Bernuth, H.V.; Zychlinsky, A. Diverse stimuli engage different neutrophil extracellular trap pathways. eLife 2017, 6, e24437. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Li, M.; Stadler, S.; Correll, S.; Li, P.; Wang, D.; Hayama, R.; Leonelli, L.; Han, H.; Grigoryev, S.A.; et al. Histone hypercitrullination mediates chromatin decondensation and neutrophil extra-cellular trap formation. J. Cell Biol. 2009, 184, 205–213. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Neeli, I.; Khan, S.; Radic, M. Histone Deimination as a Response to Inflammatory Stimuli in Neutrophils. J. Immunol. 2008, 180, 1895–1902. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, P.; Li, M.; Lindberg, M.R.; Kennett, M.J.; Xiong, N.; Wang, Y. PAD4 is essential for antibacterial innate immunity mediated by neutrophil extracellular traps. J. Exp. Med. 2010, 207, 1853–1862. [Google Scholar] [CrossRef]
- Martinod, K.; Demers, M.; Fuchs, T.A.; Wong, S.L.; Brill, A.; Gallant, M.; Hu, J.; Wang, Y.; Wagner, D.D. Neutrophil histone modification by peptidylarginine deiminase 4 is critical for deep vein thrombosis in mice. Proc. Natl. Acad. Sci. USA 2013, 110, 8674–8679. [Google Scholar] [CrossRef] [Green Version]
- Lewis, H.D.; Liddle, J.; Coote, J.E.; Atkinson, S.J.; Barker, M.D.; Bax, B.D.; Bicker, K.L.; Bingham, R.P.; Campbell, M.; Chen, Y.H.; et al. Inhibition of PAD4 activity is sufficient to disrupt mouse and human NET formation. Nat. Chem. Biol. 2015, 11, 189–191. [Google Scholar] [CrossRef]
- Leshner, M.; Wang, S.; Lewis, C.; Zheng, H.; Chen, X.A.; Santy, L.; Wang, Y. PAD4 mediated histone hypercitrullination induces heterochromatin decondensation and chromatin unfolding to form neutrophil extracellular trap-like structures. Front. Immunol. 2012, 3, 307. [Google Scholar] [CrossRef] [Green Version]
- Guiducci, E.; Lemberg, C.; Küng, N.; Schraner, E.; Theocharides, A.; LeibundGut-Landmann, S. Candida albicans-Induced NETosis Is Independent of Peptidylarginine Deiminase 4. Front. Immunol. 2018, 9, 1573. [Google Scholar] [CrossRef]
- Tatsiy, O.; McDonald, P.P. Physiological Stimuli Induce PAD4-Dependent, ROS-Independent NETosis, With Early and Late Events Controlled by Discrete Signaling Pathways. Front. Immunol. 2018, 9, 2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rohrbach, A.S.; Slade, D.J.; Thompson, P.R.; Mowen, K.A. Activation of PAD4 in NET formation. Front. Immunol. 2012, 3, 360. [Google Scholar] [CrossRef] [Green Version]
- Thiam, H.R.; Wong, S.L.; Qiu, R.; Kittisopikul, M.; Vahabikashi, A.; Goldman, A.E.; Goldman, R.D.; Wagner, D.D.; Waterman, C.M. NETosis proceeds by cytoskeleton and endomembrane disassembly and PAD4-mediated chromatin decondensation and nuclear envelope rupture. Proc. Natl. Acad. Sci. USA 2020, 117, 7326–7337. [Google Scholar] [CrossRef] [PubMed]
- Dwivedi, N.; Neeli, I.; Schall, N.; Wan, H.; Desiderio, D.M.; Csernok, E.; Thompson, P.R.; Dali, H.; Briand, J.-P.; Muller, S.; et al. Deimination of linker histones links neutrophil extracellular trap release with autoantibodies in systemic autoimmunity. FASEB J. 2014, 28, 2840–2851. [Google Scholar] [CrossRef] [Green Version]
- Chapman, E.A.; Lyon, M.; Simpson, D.; Mason, D.; Beynon, R.J.; Moots, R.J.; Wright, H.L. Caught in a Trap? Proteomic Analysis of Neutrophil Extracellular Traps in Rheumatoid Arthritis and Systemic Lupus Erythematosus. Front. Immunol. 2019, 10, 423. [Google Scholar] [CrossRef]
- Barbu, E.A.; Dominical, V.M.; Mendelsohn, L.; Thein, S.L. Detection and Quantification of Histone H4 Citrullination in Early NETosis With Image Flow Cytometry Version 4. Front. Immunol. 2020, 11, 1335. [Google Scholar] [CrossRef] [PubMed]
- Mauracher, L.M.; Posch, F.; Martinod, K.; Grilz, E.; Däullary, T.; Hell, L.; Brostjan, C.; Zielinski, C.; Ay, C.; Wagner, D.D.; et al. Citrullinated histone H3, a biomarker of neutrophil extracellular trap for-mation, predicts the risk of venous thromboembolism in cancer patients. J. Thromb. Haemost. 2018, 16, 508–518. [Google Scholar] [CrossRef] [Green Version]
- Wu, Z.; Deng, Q.; Pan, B.; Alam, H.B.; Tian, Y.; Bhatti, U.F.; Liu, B.; Mondal, S.; Thompson, P.R.; Li, Y. Inhibition of PAD2 Improves Survival in a Mouse Model of Lethal LPS-Induced Endotoxic Shock. Inflammation 2020, 43, 1436–1445. [Google Scholar] [CrossRef]
- Bawadekar, M.; Shim, D.; Johnson, C.J.; Warner, T.F.; Rebernick, R.; Damgaard, D.; Nielsen, C.H.; Pruijn, G.J.M.; Nett, J.E.; Shelef, M.A. Peptidylarginine deiminase 2 is required for tumor necrosis factor al-pha-induced citrullination and arthritis, but not neutrophil extracellular trap formation. J. Autoimmun. 2017, 80, 39–47. [Google Scholar] [CrossRef]
- Holmes, C.L.; Shim, D.; Kernien, J.; Johnson, C.J.; Nett, J.E.; Shelef, M.A. Insight into Neutrophil Extracellular Traps through Systematic Evaluation of Citrullination and Peptidylarginine Deiminases. J. Immunol. Res. 2019, 2019, 2160192. [Google Scholar] [CrossRef]
- Mutua, V.; Gershwin, L.J. A Review of Neutrophil Extracellular Traps (NETs) in Disease: Potential Anti-NETs Therapeutics. Clin. Rev. Allergy Immunol. 2021, 61, 194–211. [Google Scholar] [CrossRef]
- Kaplan, M.J.; Radic, M. Neutrophil Extracellular Traps: Double-Edged Swords of Innate Immunity. J. Immunol. 2012, 189, 2689–2695. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Papayannopoulos, V. Neutrophil extracellular traps in immunity and disease. Nat. Rev. Immunol. 2017, 18, 134–147. [Google Scholar] [CrossRef] [PubMed]
- Donskov, F. Immunomonitoring and prognostic relevance of neutrophils in clinical trials. Semin. Cancer Biol. 2013, 23, 200–207. [Google Scholar] [CrossRef] [Green Version]
- Sionov, R.V.; Fridlender, Z.G.; Granot, Z. The Multifaceted Roles Neutrophils Play in the Tumor Microenvironment. Cancer Microenviron. 2014, 8, 125–158. [Google Scholar] [CrossRef] [Green Version]
- Demers, M.; Wong, S.L.; Martinod, K.; Gallant, M.; Cabral, J.E.; Wang, Y.; Wagner, D.D. Priming of neutrophils toward NETosis promotes tumor growth. OncoImmunology 2016, 5, e1134073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaltenmeier, C.; Simmons, R.L.; Tohme, S.; Yazdani, H.O. Neutrophil Extracellular Traps (NETs) in Cancer Metastasis. Cancers 2021, 13, 6131. [Google Scholar] [CrossRef]
- Yazdani, H.O.; Roy, E.; Comerci, A.J.; van der Windt, D.J.; Zhang, H.; Huang, H.; Loughran, P.; Shiva, S.; Geller, D.A.; Bartlett, D.L.; et al. Neutrophil Extracellular Traps Drive Mitochondrial Homeostasis in Tumors to Augment Growth. Cancer Res. 2019, 79, 5626–5639. [Google Scholar] [CrossRef] [Green Version]
- Tohme, S.; Yazdani, H.O.; Al-Khafaji, A.B.; Chidi, A.P.; Loughran, P.; Mowen, K.; Wang, Y.; Simmons, R.L.; Huang, H.; Tsung, A. Neutrophil Extracellular Traps Promote the Development and Progression of Liver Metastases after Surgical Stress. Cancer Res. 2016, 76, 1367–1380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Najmeh, S.; Cools-Lartigue, J.; Rayes, R.F.; Gowing, S.; Vourtzoumis, P.; Bourdeau, F.; Giannias, B.; Berube, J.; Rousseau, S.; Ferri, L.E.; et al. Neutrophil extracellular traps sequester circulating tumor cells via β1-integrin mediated interactions. Int. J. Cancer 2017, 140, 2321–2330. [Google Scholar] [CrossRef] [Green Version]
- Monti, M.; De Rosa, V.; Iommelli, F.; Carriero, M.V.; Terlizzi, C.; Camerlingo, R.; Belli, S.; Fonti, R.; Di Minno, G.; Del Vecchio, S. Neutrophil Extracellular Traps as an Adhesion Substrate for Different Tumor Cells Expressing RGD-Binding Integrins. Int. J. Mol. Sci. 2018, 19, 2350. [Google Scholar] [CrossRef] [Green Version]
- Cools-Lartigue, J.; Spicer, J.; McDonald, B.; Gowing, S.; Chow, S.; Giannias, B.; Bourdeau, F.; Kubes, P.; Ferri, L. Neutrophil extracellular traps sequester circulating tumor cells and promote metastasis. J. Clin. Investig. 2013, 123, 3446–3458. [Google Scholar] [CrossRef] [PubMed]
- Park, J.; Wysocki, R.W.; Amoozgar, Z.; Maiorino, L.; Fein, M.R.; Jorns, J.; Schott, A.F.; Kinugasa-Katayama, Y.; Lee, Y.; Won, N.H.; et al. Cancer cells induce metastasis-supporting neutrophil extracellular DNA traps. Sci. Transl. Med. 2016, 8, 361ra138. [Google Scholar] [CrossRef] [PubMed]
- Teijeira, Á.; Garasa, S.; Gato, M.; Alfaro, C.; Migueliz, I.; Cirella, A.; de Andrea, C.; Ochoa, M.C.; Otano, I.; Etxeberria, I.; et al. CXCR1 and CXCR2 Chemokine Receptor Agonists Produced by Tumors Induce Neutrophil Extracellular Traps that Interfere with Immune Cytotoxicity. Immunity 2020, 52, 856–871.e8. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.-Y.; Luo, Q.; Lu, L.; Zhu, W.-W.; Sun, H.-T.; Wei, R.; Lin, Z.-F.; Wang, X.-Y.; Wang, C.-Q.; Lu, M.; et al. Increased neutrophil extracellular traps promote metastasis potential of hepatocellular carcinoma via provoking tumorous inflammatory response. J. Hematol. Oncol. 2020, 13, 3. [Google Scholar] [CrossRef] [Green Version]
- Zha, C.; Meng, X.; Li, E.a.L.; Mi, S.; Qian, D.; Li, Z.; Wu, P.; Hu, S.; Zhao, S.; Cai, J.; et al. Neutrophil extracellular traps mediate the crosstalk between glioma progression and the tumor microenvironment via the HMGB1/RAGE/IL-8 axis. Cancer Biol. Med. 2020, 17, 154–168. [Google Scholar] [CrossRef] [PubMed]
- Martins-Cardoso, K.; Almeida, V.H.; Bagri, K.M.; Rossi, M.I.D.; Mermelstein, C.S.; König, S.; Monteiro, R.Q. Neutrophil Extracellular Traps (NETs) Promote Pro-Metastatic Phenotype in Human Breast Cancer Cells through Epithelial–Mesenchymal Transition. Cancers 2020, 12, 1542. [Google Scholar] [CrossRef]
- Stehr, A.M.; Wang, G.; Demmler, R.; Stemmler, M.P.; Krug, J.; Tripal, P.; Schmid, B.; Geppert, C.I.; Hartmann, A.; Muñoz, L.E.; et al. Neutrophil extracellular traps drive epithelial–mesenchymal transition of human colon cancer. J. Pathol. 2022, 256, 455–467. [Google Scholar] [CrossRef]
- Zhu, T.; Zou, X.; Yang, C.; Li, L.; Wang, B.; Li, R.; Li, H.; Xu, Z.; Huang, D.; Wu, Q. Neutrophil extracellular traps promote gastric cancer metastasis by inducing epitheli-al-mesenchymal transition. Int. J. Mol. Med. 2021, 48, 127. [Google Scholar] [CrossRef]
- Albrengues, J.; Shields, M.A.; Ng, D.; Park, C.G.; Ambrico, A.; Poindexter, M.E.; Upadhyay, P.; Uyeminami, D.L.; Pommier, A.; Küttner, V.; et al. Neutrophil extracellular traps produced during inflammation awaken dormant cancer cells in mice. Science 2018, 361, 4227. [Google Scholar] [CrossRef] [Green Version]
- Demers, M.; Wagner, D.D. Neutrophil extracellular traps. OncoImmunology 2014, 2, e22946. [Google Scholar] [CrossRef] [Green Version]
- von Brühl, M.; Stark, K.; Steinhart, A.; Chandraratne, S.; Konrad, I.; Lorenz, M.; Khandoga, A.; Tirniceriu, A.; Coletti, R.; Köllnberger, M. Monocytes, neutrophils, and platelets cooperate to initiate and propagate venous thrombosis in mice in vivo. J. Exp. Med. 2012, 209, 819–835. [Google Scholar] [CrossRef] [PubMed]
- Swystun, L.L.; Mukherjee, S.; Liaw, P.C. Breast cancer chemotherapy induces the release of cell-free DNA, a novel procoagulant stimulus. J. Thromb. Haemost. 2011, 9, 2313–2321. [Google Scholar] [CrossRef] [PubMed]
- Gould, T.J.; Vu, T.T.; Swystun, L.L.; Dwivedi, D.J.; Mai, S.H.; Weitz, J.I.; Liaw, P.C. Neutrophil Extracellular Traps Promote Thrombin Generation Through Plate-let-Dependent and Platelet-Independent Mechanisms. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1977–1984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Masucci, M.T.; Minopoli, M.; Del Vecchio, S.; Carriero, M.V. The Emerging Role of Neutrophil Extracellular Traps (NETs) in Tumor Progression and Metastasis. Front. Immunol. 2020, 11, 1749. [Google Scholar] [CrossRef]
- Jung, H.S.; Gu, J.; Kim, J.; Nam, Y.; Song, J.W.; Kim, H.K. Cancer cell–induced neutrophil extracellular traps promote both hypercoagulability and cancer progression. PLoS ONE 2019, 14, e216055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, J.; Zhang, X.; Pelayo, R.; Monestier, M.; Ammollo, C.T.; Semeraro, F.; Taylor, F.B.; Esmon, N.L.; Lupu, F.; Esmon, C.T. Extracellular histones are major mediators of death in sepsis. Nat. Med. 2009, 15, 1318–1321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saffarzadeh, M.; Juenemann, C.; Queisser, M.A.; Lochnit, G.; Barreto, G.; Galuska, S.P.; Lohmeyer, J.; Preissner, K.T. Neutrophil Extracellular Traps Directly Induce Epithelial and Endo-thelial Cell Death: A Predominant Role of Histones. PLoS ONE 2012, 7, e32366. [Google Scholar] [CrossRef]
- Semeraro, F.; Ammollo, C.T.; Morrissey, J.H.; Dale, G.L.; Friese, P.; Esmon, N.L.; Esmon, C.T. Extracellular histones promote thrombin generation through plate-let-dependent mechanisms: Involvement of platelet TLR2 and TLR4. Blood 2011, 118, 1952–1961. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, T.A.; Bhandari, A.A.; Wagner, D.D. Histones induce rapid and profound thrombocytopenia in mice. Blood 2011, 118, 3708–3714. [Google Scholar] [CrossRef] [Green Version]
- Ammollo, C.T.; Semeraro, F.; Xu, J.; Esmon, N.L.; Esmon, C.T. Extracellular histones increase plasma thrombin generation by impairing thrombomodulin-dependent protein C activation. J. Thromb. Haemost. 2011, 9, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Noubouossie, D.F.; Whelihan, M.F.; Yu, Y.; Sparkenbaugh, E.; Pawlinski, R.; Monroe, D.M.; Key, N.S. In vitro activation of coagulation by human neutrophil DNA and histone proteins but not neutrophil extracellular traps. Blood 2017, 129, 1021–1029. [Google Scholar] [CrossRef] [PubMed]
- Massberg, S.; Grahl, L.; von Bruehl, M.-L.; Manukyan, D.; Pfeiler, S.; Goosmann, C.; Brinkmann, V.; Lorenz, M.; Bidzhekov, K.; Khandagale, A.B.; et al. Reciprocal coupling of coagulation and innate immunity via neutrophil serine proteases. Nat. Med. 2010, 16, 887–896. [Google Scholar] [CrossRef]
- Sorvillo, N.; Cherpokova, D.; Martinod, K.; Wagner, D.D. Extracellular DNA NET-Works with Dire Consequences for Health. Circ. Res. 2019, 125, 470–488. [Google Scholar] [CrossRef] [PubMed]
- McDonald, B.; Davis, R.P.; Kim, S.-J.; Tse, M.; Esmon, C.T.; Kolaczkowska, E.; Jenne, C.N. Platelets and neutrophil extracellular traps collaborate to promote intravascular coagulation during sepsis in mice. Blood 2017, 129, 1357–1367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Etulain, J.; Martinod, K.; Wong, S.L.; Cifuni, S.M.; Schattner, M.; Wagner, D.D. P-selectin promotes neutrophil extracellular trap formation in mice. Blood 2015, 126, 242–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sreeramkumar, V.; Adrover, J.M.; Ballesteros, I.; Cuartero, M.I.; Rossaint, J.; Bilbao, I.; Nácher, M.; Pitaval, C.; Radovanovic, I.; Fukui, Y.; et al. Neutrophils scan for activated platelets to initiate inflammation. Science 2014, 346, 1234–1238. [Google Scholar] [CrossRef] [Green Version]
- Panicker, S.R.; Mehta-DSouza, P.; Zhang, N.; Klopocki, A.G.; Shao, B.; McEver, R.P. Circulating soluble P-selectin must dimerize to promote inflammation and coagulation in mice. Blood 2017, 130, 181–191. [Google Scholar] [CrossRef] [Green Version]
- Maugeri, N.; Campana, L.; Gavina, M.; Covino, C.; De Metrio, M.; Panciroli, C.; Maiuri, L.; Maseri, A.; D’Angelo, A.; Bianchi, M.E.; et al. Activated platelets present high mobility group box 1 to neutrophils, inducing autophagy and promoting the extrusion of neutrophil extracellular traps. J. Thromb. Haemost. 2014, 12, 2074–2088. [Google Scholar] [CrossRef]
- Dyer, M.R.; Chen, Q.; Haldeman, S.; Yazdani, H.; Hoffman, R.; Loughran, P.; Tsung, A.; Zuckerbraun, B.S.; Simmons, R.L.; Neal, M.D. Deep vein thrombosis in mice is regulated by platelet HMGB1 through release of neutrophil-extracellular traps and DNA. Sci. Rep. 2018, 8, 2068. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.W.; Lee, J.K. Role of HMGB1 in the Interplay between NETosis and Thrombosis in Ischemic Stroke: A Review. Cells 2020, 9, 1794. [Google Scholar] [CrossRef] [PubMed]
- Shi, L.; Yao, H.; Liu, Z.; Xu, M.; Tsung, A.; Wang, Y. Endogenous PAD4 in Breast Cancer Cells Mediates Cancer Extracellular Chromatin Network Formation and Promotes Lung Metastasis. Mol. Cancer Res. 2020, 18, 735–747. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Berger-Achituv, S.; Brinkmann, V.; Abed, U.A.; Kühn, L.I.; Ben-Ezra, J.; Elhasid, R.; Zychlinsky, A. A proposed role for neutrophil extracellular traps in cancer immunoediting. Front. Immunol. 2013, 4, 48. [Google Scholar] [CrossRef] [PubMed]
- Boone, B.A.; Murthy, P.; Miller-Ocuin, J.; Doerfler, W.R.; Ellis, J.T.; Liang, X.; Ross, M.A.; Wallace, C.T.; Sperry, J.L.; Lotze, M.T.; et al. Chloroquine reduces hypercoagulability in pancreatic cancer through inhi-bition of neutrophil extracellular traps. BMC Cancer 2018, 18, 678. [Google Scholar] [CrossRef] [Green Version]
- Wong, S.L.; Wagner, D.D. Peptidylarginine deiminase 4: A nuclear button triggering neutrophil extracellular traps in inflammatory diseases and aging. FASEB J. 2018, 32, 6258–6370. [Google Scholar] [CrossRef]
- Kolaczkowska, E.; Jenne, C.N.; Surewaard, B.G.J.; Thanabalasuriar, A.; Lee, W.Y.; Sanz, M.J.; Mowen, K.; Opdenakker, G.; Kubes, P. Molecular mechanisms of NET formation and degradation revealed by intravital imaging in the liver vasculature. Nat. Commun. 2015, 6, 6673. [Google Scholar] [CrossRef] [Green Version]
- Alasmari, S.Z. In Vivo Imaging of Neutrophil Extracellular Traps (NETs): Visualization Methods and Outcomes. BioMed Res. Int. 2020, 2020, 4192745. [Google Scholar] [CrossRef] [Green Version]
- Chang, X.; Fang, K. PADI4 and tumourigenesis. Cancer Cell Int. 2010, 10, 7. [Google Scholar] [CrossRef] [Green Version]
- Thålin, C.; Lundström, S.; Seignez, C.; Daleskog, M.; Lundström, A.; Henriksson, P.; Helleday, T.; Phillipson, M.; Wallén, H.; Demers, M. Citrullinated histone H3 as a novel prognostic blood marker in patients with advanced cancer. PLoS ONE 2018, 13, e191231. [Google Scholar] [CrossRef]
- Visconti, L.; Nelissen, K.; Deckx, L.; Van Den Akker, M.; Adriaensen, W.; Daniels, L.; Matheï, C.; Linsen, L.; Hellings, N.; Stinissen, P.; et al. Prognostic value of circulating cytokines on overall survival and disease-free survival in cancer patients. Biomark. Med. 2014, 8, 297–306. [Google Scholar] [CrossRef]
- Sanmamed, M.F.; Carranza-Rua, O.; Alfaro, C.; Oñate, C.; Martín-Algarra, S.; Perez, G.; Landazuri, S.F.; Gonzalez, A.; Gross, S.; Rodriguez, I.; et al. Serum Interleukin-8 Reflects Tumor Burden and Treatment Response across Malignancies of Multiple Tissue Origins. Clin. Cancer Res. 2014, 20, 5697–5707. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Y.; Zhao, G.; Xu, B.; Liu, C.; Li, C.; Zhang, X.; Chang, X. PADI4 has genetic susceptibility to gastric carcinoma and upregulates CXCR2, KRT14 and TNF-α expression levels. Oncotarget 2016, 7, 62159–62176. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xin, J.; Song, X. Role of peptidylarginine deiminase type 4 in gastric cancer. Exp. Ther. Med. 2016, 12, 3155–3160. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.T.; Wu, H.; Li, H.L.; Zheng, Y.B. PADI4 promotes epithelial-mesenchymal transition (EMT) in gastric cancer via the upregulation of interleukin 8. BMC Gastroenterol. 2022, 22, 25. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Chang, X.; Yuan, G.; Zhao, Y.; Wang, P. Expression of Peptidylarginine Deiminase Type 4 in Ovarian Tumors. Int. J. Biol. Sci. 2010, 6, 454–464. [Google Scholar] [CrossRef] [PubMed]
- Cui, Y.-Y.; Yan, L.; Zhou, J.; Zhao, S.; Zheng, Y.-B.; Sun, B.-H.; Lv, H.-T.; Rong, F.-N.; Chang, X.-T. The role of peptidylarginine deiminase 4 in ovarian cancer cell tumorigenesis and invasion. Tumor Biol. 2015, 37, 5375–5383. [Google Scholar] [CrossRef]
- Lee, W.; Ko, S.Y.; Mohamed, M.S.; Kenny, H.A.; Lengyel, E.; Naora, H. Neutrophils facilitate ovarian cancer premetastatic niche formation in the omentum. J. Exp. Med. 2019, 216, 176–194. [Google Scholar] [CrossRef]
- Zhai, Q.; Qin, J.; Jin, X.; Sun, X.; Wang, L.; Du, W.; Li, T.; Xiang, X. PADI4 modulates the invasion and migration of osteosarcoma cells by down-regulation of epithelial-mesenchymal transition. Life Sci. 2020, 256, 117968. [Google Scholar] [CrossRef]
- Guo, J.; Yin, L.; Zhang, X.; Su, P.; Zhai, Q. Factors Associated with Promoted Proliferation of Osteosarcoma by Peptidylarginine Deiminase 4. BioMed Res. Int. 2021, 2021, 5596014. [Google Scholar] [CrossRef]
- Chen, H.; Luo, M.; Wang, X.; Liang, T.; Huang, C.; Huang, C.; Wei, L. Inhibition of PAD4 enhances radiosensitivity and inhibits aggressive phenotypes of nasopharyngeal carcinoma cells. Cell. Mol. Biol. Lett. 2021, 26, 9. [Google Scholar] [CrossRef]
- Chen, H.; Wei, L.; Luo, M.; Wang, X.; Zhan, Y.; Mao, Y.; Huang, C.; Li, J.; Lu, H. PAD4 inhibitor promotes DNA damage and radiosensitivity of nasopharyngeal carcinoma cells. Environ. Toxicol. 2021, 36, 2291–2301. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Wei, L.; Luo, M.; Wang, X.; Zhu, C.; Huang, H.; Liu, X.; Lu, H.; Zhong, Y. LINC00324 suppresses apoptosis and autophagy in nasopharyngeal carcinoma through upregulation of PAD4 and activation of the PI3K/AKT signaling pathway. Cell Biol. Toxicol. 2021, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Fan, T.; Zhang, C.; Zong, M.; Zhao, Q.; Yang, X.; Hao, C.; Zhang, H.; Yu, S.; Guo, J.; Gong, R.; et al. Peptidylarginine deiminase IV promotes the development of chemoresistance through inducing autophagy in hepatocellular carcinoma. Cell Biosci. 2014, 4, 49. [Google Scholar] [CrossRef] [PubMed]
- Lu, M.; Zhang, X.; Xu, Y.; He, G.; Liu, Q.; Zhu, J.; Zhang, C. Elevated histone H3 citrullination is associated with increased Beclin1 expression in HBV-related hepatocellular carcinoma. J. Med. Virol. 2020, 92, 1221–1230. [Google Scholar] [CrossRef]
- van der Windt, D.J.; Sud, V.; Zhang, H.; Varley, P.R.; Goswami, J.; Yazdani, H.O.; Tohme, S.; Loughran, P.; O’Doherty, R.M.; Minervini, M.I.; et al. Neutrophil extracellular traps promote inflammation and development of hepatocellular carcinoma in nonalcoholic steatohepatitis. Hepatology 2018, 68, 1347–1360. [Google Scholar] [CrossRef] [Green Version]
- Guan, X.; Lu, Y.; Zhu, H.; Yu, S.; Zhao, W.; Chi, X.; Xie, C.; Yin, Z. The Crosstalk Between Cancer Cells and Neutrophils Enhances Hepatocellular Carcinoma Metastasis via Neutrophil Extracellular Traps-Associated Cathepsin G Component: A Potential Therapeutic Target. J. Hepatocell. Carcinoma 2021, 8, 451–465. [Google Scholar] [CrossRef]
- Arelaki, S.; Arampatzioglou, A.; Kambas, K.; Papagoras, C.; Miltiades, P.; Angelidou, I.; Mitsios, A.; Kotsianidis, I.; Skendros, P.; Sivridis, E.; et al. Gradient Infiltration of Neutrophil Extracellular Traps in Colon Cancer and Evidence for Their Involvement in Tumour Growth. PLoS ONE 2016, 11, e0154484. [Google Scholar] [CrossRef] [Green Version]
- Baka, Z.; Barta, P.; Losonczy, G.; Krenács, T.; Pápay, J.; Szarka, E.; Sármay, G.; Babos, F.; Magyar, A.; Géher, P.; et al. Specific expression of PAD4 and citrullinated proteins in lung cancer is not associated with anti-CCP antibody production. Int. Immunol. 2011, 23, 405–414. [Google Scholar] [CrossRef]
- Ulivi, P.; Mercatali, L.; Casoni, G.L.; Scarpi, E.; Bucchi, L.; Silvestrini, R.; Sanna, S.; Monteverde, M.; Amadori, D.; Poletti, V.; et al. Multiple Marker Detection in Peripheral Blood for NSCLC Diagnosis. PLoS ONE 2013, 8, e57401. [Google Scholar] [CrossRef] [Green Version]
- Lu, Y.; Peng, Z.; Zhu, D.; Jia, Y.; Taledaohan, A.; Li, Y.; Liu, J.; Wang, Y.; Wang, Y. RGD Peptide and PAD4 Inhibitor-Loaded Gold Nanorods for Chemo-Photothermal Combined Therapy to Inhibit Tumor Growth, Prevent Lung Metastasis and Improve Biosafety. Int. J. Nanomed. 2021, 16, 5565–5580. [Google Scholar] [CrossRef]
- Moshkovich, N.; Ochoa, H.J.; Tang, B.; Yang, H.H.; Yang, Y.; Huang, J.; Lee, M.P.; Wakefield, L.M. Peptidylarginine deiminase IV (PADI4) regulates breast cancer stem cells via a novel tumor cell-autonomous suppressor role. Cancer Res. 2020, 80, 2125–2137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zheng, Q.; Osunsade, A.; David, Y. Protein arginine deiminase 4 antagonizes methylglyoxal-induced histone glycation. Nat. Commun. 2020, 11, 3241. [Google Scholar] [CrossRef] [PubMed]
- Wei, L.; Wang, X.; Luo, M.; Wang, H.; Chen, H.; Huang, C. The PAD4 inhibitor GSK484 enhances the radiosensitivity of triple-negative breast cancer. Hum. Exp. Toxicol. 2020, 40, 1074–1083. [Google Scholar] [CrossRef]
- Wang, Y.; Lyu, Y.; Tu, K.; Xu, Q.; Yang, Y.; Salman, S.; Le, N.; Lu, H.; Chen, C.; Zhu, Y.; et al. Histone citrullination by PADI4 is required for HIF-dependent transcriptional responses to hypoxia and tumor vascularization. Sci. Adv. 2021, 7, eabe3771. [Google Scholar] [CrossRef]
- Rocks, N.; Vanwinge, C.; Radermecker, C.; Blacher, S.; Gilles, C.; Marée, R.; Gillard, A.; Evrard, B.; Pequeux, C.; Marichal, T.; et al. Ozone-primed neutrophils promote early steps of tumour cell metastasis to lungs by enhancing their NET production. Thorax 2019, 74, 768–779. [Google Scholar] [CrossRef] [PubMed]
- Nakashima, K.; Hagiwara, T.; Ishigami, A.; Nagata, S.; Asaga, H.; Kuramoto, M.; Senshu, T.; Yamada, M. Molecular Characterization of Peptidylarginine Deiminase in HL-60 Cells Induced by Retinoic Acid and 1α,25-Dihydroxyvitamin D3*. J. Biol. Chem. 1999, 274, 27786–27792. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nakashima, K.; Arai, S.; Suzuki, A.; Nariai, Y.; Urano, T.; Nakayama, M.; Ohara, O.; Yamamura, K.-I.; Yamamoto, K.; Miyazaki, T. PAD4 regulates proliferation of multipotent haematopoietic cells by controlling c-myc expression. Nat. Commun. 2013, 4, 1836–1838. [Google Scholar] [CrossRef] [Green Version]
- Prior, J.C. Ovarian Aging and the Perimenopausal Transition: The Paradox of Endogenous Ovarian Hyperstimulation. Endocrine 2005, 26, 297–300. [Google Scholar] [CrossRef]
- Xu, Z.; Han, X.; Ou, D.; Liu, T.; Li, Z.; Jiang, G.; Liu, J.; Zhang, J. Targeting PI3K/AKT/mTOR-mediated autophagy for tumor therapy. Appl. Microbiol. Biotechnol. 2020, 104, 575–587. [Google Scholar] [CrossRef]
- Liu, R.; Chen, Y.; Liu, G.; Li, C.; Song, Y.; Cao, Z.; Li, W.; Hu, J.; Lu, C.; Liu, Y. PI3K/AKT pathway as a key link modulates the multidrug resistance of cancers. Cell Death Dis. 2020, 11, 797. [Google Scholar] [CrossRef]
- Wang, X.; Zhang, A.; Sun, H. Power of metabolomics in diagnosis and biomarker discovery of hepatocellular carcinoma. Hepatology 2012, 57, 2072–2077. [Google Scholar] [CrossRef] [PubMed]
- Kopetz, S.; Chang, G.J.; Overman, M.J.; Eng, C.; Sargent, D.J.; Larson, D.W.; Grothey, A.; Vauthey, J.N.; Nagorney, D.M.; McWilliams, R.R. Improved Survival in Metastatic Colorectal Cancer Is Associated with Adoption of Hepatic Resection and Improved Chemotherapy. J. Clin. Oncol. 2009, 27, 3677–3683. [Google Scholar] [CrossRef] [PubMed]
- Wolpin, B.M.; Mayer, R.J. Systemic Treatment of Colorectal Cancer. Gastroenterology 2008, 134, 1296–1310. [Google Scholar] [CrossRef] [PubMed]
- Reck, M.; Rabe, K.F. Precision Diagnosis and Treatment for Advanced Non-Small-Cell Lung Cancer. N. Engl. J. Med. 2017, 377, 849–861. [Google Scholar] [CrossRef] [Green Version]
- Siegel, R.L.; Miller, K.D.; Fuchs, H.E.; Jemal, A. Cancer statistics, 2022. CA A Cancer J. Clin. 2022, 72, 7–33. [Google Scholar] [CrossRef]
- Dong, S.; Zhang, Z.; Takahara, H. Estrogen-Enhanced Peptidylarginine Deiminase Type IV Gene (PADI4) Expression in MCF-7 Cells Is Mediated by Estrogen Receptor-α-Promoted Transfactors Activator Protein-1, Nuclear Factor-Y, and Sp1. Mol. Endocrinol. 2007, 21, 1617–1629. [Google Scholar] [CrossRef] [Green Version]
- Voduc, K.D.; Cheang MC, U.; Tyldesley, S.; Gelmon, K.; Nielsen, T.O.; Kennecke, H. Breast Cancer Subtypes and the Risk of Local and Regional Relapse. J. Clin. Oncol. 2010, 28, 1684–1691. [Google Scholar] [CrossRef] [Green Version]
- Chen, X.; Yu, X.; Chen, J.; Zhang, Z.; Tuan, J.; Shao, Z.; Guo, X.; Feng, Y. Analysis in early stage triple-negative breast cancer treated with mastectomy without adjuvant radiotherapy: Patterns of failure and prognostic factors. Cancer 2013, 119, 2366–2374. [Google Scholar] [CrossRef]
- Hung, H.-C.; Lin, C.-Y.; Liao, Y.-F.; Hsu, P.-C.; Tsay, G.J.; Liu, G.-Y. The functional haplotype of peptidylarginine deiminase IV (S55G, A82V and A112G) associated with susceptibility to rheumatoid arthritis dominates apoptosis of acute T leukemia Jurkat cells. Apoptosis 2007, 12, 475–487. [Google Scholar] [CrossRef]
- Blom, J.W.; Doggen, C.J.; Osanto, S.; Rosendaal, F.R. Malignancies, prothrombotic mutations, and the risk of venous thrombosis. J. Vasc. Surg. 2005, 293, 715–722. [Google Scholar]
- Ikushima, S.; Ono, R.; Fukuda, K.; Sakayori, M.; Awano, N.; Kondo, K. Trousseau’s syndrome: Cancer-associated thrombosis. Jpn. J. Clin. Oncol. 2016, 46, 204–208. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Demers, M.; Krause, D.S.; Schatzberg, D.; Martinod, K.; Voorhees, J.R.; Fuchs, T.A.; Scadden, D.T.; Wagner, D.D. Cancers predispose neutrophils to release extracellular DNA traps that contribute to cancer-associated thrombosis. Proc. Natl. Acad. Sci. USA 2012, 109, 13076–13081. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, Y.; Tao, W.; Shen, F.; Du, W.; Xu, Z.; Liu, Z. The Emerging Role of Neutrophil Extracellular Traps in Arterial, Venous and Cancer-Associated Thrombosis. Front. Cardiovasc. Med. 2021, 8. [Google Scholar] [CrossRef] [PubMed]
- Leal, A.C.; Mizurini, D.M.; Gomes, T.; Rochael, N.C.; Saraiva, E.M.; Dias, M.S.; Werneck, C.C.; Sielski, M.S.; Vicente, C.P.; Monteiro, R.Q. Tumor-Derived Exosomes Induce the Formation of Neutrophil Extracellular Traps: Implications for The Establishment of Cancer-Associated Thrombosis. Sci. Rep. 2017, 7, 6438. [Google Scholar] [CrossRef] [PubMed]
- Snoderly, H.T.; Boone, B.A.; Bennewitz, M.F. Neutrophil extracellular traps in breast cancer and beyond: Current perspectives on NET stimuli, thrombosis and metastasis, and clinical utility for diagnosis and treatment. Breast Cancer Res. 2019, 21, 145. [Google Scholar] [CrossRef] [Green Version]
- Laridan, E.; Martinod, K.; De Meyer, S.F. Neutrophil Extracellular Traps in Arterial and Venous Thrombosis. Semin. Thromb. Hemost. 2019, 45, 086–093. [Google Scholar] [CrossRef]
- Kimball, A.S.; Obi, A.T.; Diaz, J.A.; Henke, P.K. The Emerging Role of NETs in Venous Thrombosis and Immunothrombosis. Front. Immunol. 2016, 7, 236. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, T.A.; Brill, A.; Duerschmied, D.; Schatzberg, D.; Monestier, M.; Myers, D.D., Jr.; Wrobleski, S.K.; Wakefield, T.W.; Hartwig, J.H.; Wagner, D.D. Extracellular DNA traps promote thrombosis. Proc. Natl. Acad. Sci. USA 2010, 107, 15880–15885. [Google Scholar] [CrossRef] [Green Version]
- Gould, T.J.; Lysov, Z.; Liaw, P.C. Extracellular DNA and histones: Double-edged swords in immunothrombosis. J. Thromb. Haemost. 2015, 13, S82–S91. [Google Scholar] [CrossRef]
- Sørensen, O.E.; Borregaard, N. Neutrophil extracellular traps—The dark side of neutrophils. J. Clin. Investig. 2016, 126, 1612–1620. [Google Scholar] [CrossRef] [Green Version]
- Fuchs, T.A.; Kremer Hovinga, J.A.; Schatzberg, D.; Wagner, D.D.; Lämmle, B. Circulating DNA and myeloperoxidase indicate disease activity in patients with thrombotic microangiopathies. Blood 2012, 120, 1157–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brill, A.; Fuchs, T.A.; Savchenko, A.S.; Chausheva, S.; Alimohammadi, A.; Artner, T.; Panzenboeck, A.; Rinderer, J.; Shafran, I.; Mangold, A.; et al. Neutrophil extracellular traps promote deep vein thrombosis in mice. J. Thromb. Haemost. 2012, 10, 136–144. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martinod, K.; Wagner, D.D. Thrombosis: Tangled up in NETs. Blood 2014, 123, 2768–2776. [Google Scholar] [CrossRef]
- Hisada, Y.; Grover, S.P.; Maqsood, A.; Houston, R.; Ay, C.; Noubouossie, D.F.; Cooley, B.C.; Wallén, H.; Key, N.S.; Thålin, C.; et al. Neutrophils and neutrophil extracellular traps enhance venous thrombosis in mice bearing human pancreatic tumors. Haematologica 2019, 105, 218–225. [Google Scholar] [CrossRef]
- Thålin, C.; Demers, M.; Blomgren, B.; Wong, S.L.; von Arbin, M.; von Heijne, A.; Laska, A.C.; Wallén, H.; Wagner, D.D.; Aspberg, S. NETosis promotes cancer-associated arterial microthrombosis presenting as ischemic stroke with troponin elevation. Thromb. Res. 2016, 139, 56–64. [Google Scholar] [CrossRef] [Green Version]
- Oklu, R.; Sheth, R.A.; Wong KH, K.; Jahromi, A.H.; Albadawi, H. Neutrophil extracellular traps are increased in cancer patients but does not associate with venous thrombosis. Cardiovasc. Diagn. Ther. 2017, 7 (Suppl. S3), S140–S149. [Google Scholar] [CrossRef]
- Perdomo, J.; Leung, H.H.L.; Ahmadi, Z.; Yan, F.; Chong, J.J.H.; Passam, F.H.; Chong, B.H. Neutrophil activation and NETosis are the major drivers of thrombosis in heparin-induced thrombocytopenia. Nat. Commun. 2019, 10, 1322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tilvawala, R.; Nguyen, S.H.; Maurais, A.J.; Nemmara, V.V.; Nagar, M.; Salinger, A.J.; Nagpal, S.; Weerapana, E.; Thompson, P.R. The Rheumatoid Arthritis-Associated Citrullinome. Cell Chem. Biol. 2018, 25, 691–704. [Google Scholar] [CrossRef] [Green Version]
- Morrell, C.N.; Hilt, Z.T.; Pariser, D.N.; Maurya, P. PAD4 and von Willebrand Factor Link Inflammation and Thrombosis. Circ. Res. 2019, 125, 520–522. [Google Scholar] [CrossRef]
- Pritzker, L.B.; Moscarello, M.A. A novel microtubule independent effect of paclitaxel: The inhibition of peptidylarginine deiminase from bovine brain. Biochim. Biophys. Acta 1998, 1388, 154–160. [Google Scholar] [CrossRef]
- Hidaka, Y.; Hagiwara, T.; Yamada, M. Methylation of the guanidino group of arginine residues prevents citrullination by peptidylarginine deiminase IV. FEBS Lett. 2005, 579, 4088–4092. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knuckley, B.; Luo, Y.; Thompson, P.R. Profiling Protein Arginine Deiminase 4 (PAD4): A novel screen to identify PAD4 inhibitors. Bioorganic Med. Chem. 2008, 16, 739–745. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Teo, C.Y.; Tejo, B.A.; Leow AT, C.; Salleh, A.B.; Abdul Rahman, M.B. Novel furan-containing peptide-based inhibitors of protein arginine deiminase type IV (PAD4). Chem. Biol. Drug Des. 2017, 90, 1134–1146. [Google Scholar] [CrossRef] [PubMed]
- Aliko, A.; Kamińska, M.; Falkowski, K.; Bielecka, E.; Benedyk-Machaczka, M.; Malicki, S.; Kozieł, J.; Wong, A.; Bryzek, D.; Kantyka, T.; et al. Discovery of Novel Potential Reversible Peptidyl Arginine Deiminase Inhibitor. Int. J. Mol. Sci. 2019, 20, 2174. [Google Scholar] [CrossRef]
- Lewallen, D.M.; Bicker, K.L.; Madoux, F.; Chase, P.; Anguish, L.; Coonrod, S.; Hodder, P.; Thompson, P.R. A FluoPol-ABPP PAD2 High-Throughput Screen Identifies the First Calcium Site Inhibitor Targeting the PADs. ACS Chem. Biol. 2014, 9, 913–921. [Google Scholar] [CrossRef]
- Knuckley, B.; Jones, J.E.; Bachovchin, D.A.; Slack, J.; Causey, C.P.; Brown, S.J.; Rosen, H.; Cravatt, B.F.; Thompson, P.R. A fluopol-ABPP HTS assay to identify PAD inhibitors. Chem. Commun. 2010, 46, 7175. [Google Scholar] [CrossRef]
- Luo, Y.; Arita, K.; Bhatia, M.; Knuckley, B.; Lee, Y.-H.; Stallcup, M.R.; Sato, M.; Thompson, P.R. Inhibitors and Inactivators of Protein Arginine Deiminase 4: Functional and Structural Characterization. Biochemistry 2006, 45, 11727–11736. [Google Scholar] [CrossRef] [Green Version]
- Causey, C.P.; Jones, J.E.; Slack, J.L.; Kamei, D.; Jones, L.E.; Subramanian, V.; Knuckley, B.; Ebrahimi, P.; Chumanevich, A.A.; Luo, Y.; et al. The development of N-α-(2-carboxyl)benzoyl-N(5)-(2-fluoro-1-iminoethyl)-l-ornithine amide (o-F-amidine) and N-α-(2-carboxyl)benzoyl-N(5)-(2-chloro-1-iminoethyl)-l-ornithine amide (o-Cl-amidine) as second generation protein ar-ginine deiminase (PAD) inhibitors. J. Med. Chem. 2011, 54, 6919–6935. [Google Scholar]
- Muth, A.; Subramanian, V.; Beaumont, E.; Nagar, M.; Kerry, P.; McEwan, P.; Srinath, H.; Clancy, K.; Parelkar, S.; Thompson, P.R. Development of a Selective Inhibitor of Protein Arginine Deiminase 2. J. Med. Chem. 2017, 60, 3198–3211. [Google Scholar] [CrossRef] [Green Version]
- Jones, J.E.; Slack, J.L.; Fang, P.; Zhang, X.; Subramanian, V.; Causey, C.P.; Coonrod, S.A.; Guo, M.; Thompson, P.R. Synthesis and Screening of a Haloacetamidine Containing Library to Identify PAD4 Selective Inhibitors. ACS Chem. Biol. 2012, 7, 160–165. [Google Scholar] [CrossRef]
- Zhu, D.; Lu, Y.; Gui, L.; Wang, W.; Hu, X.; Chen, S.; Wang, Y.; Wang, Y. Self-assembling, pH-responsive nanoflowers for inhibiting PAD4 and neutrophil extracellular trap formation and improving the tumor immune microenvironment. Acta Pharm. Sin. B 2022, 12, 2592–2608. [Google Scholar] [CrossRef] [PubMed]
- Dreyton, C.J.; Anderson, E.D.; Subramanian, V.; Boger, D.L.; Thompson, P.R. Insights into the mechanism of streptonigrin-induced protein arginine deiminase inactivation. Bioorganic Med. Chem. 2014, 22, 1362–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Knuckley, B.; Lee, Y.H.; Stallcup, M.R.; Thompson, P.R. A Fluoroacetamidine-Based Inactivator of Protein Arginine Deiminase 4: Design, Synthesis, and in Vitro and in Vivo Evaluation. J. Am. Chem. Soc. 2006, 128, 1092–1093. [Google Scholar] [CrossRef] [Green Version]
- Biron, B.M.; Chung, C.; O’Brien, X.M.; Chen, Y.; Reichner, J.S.; Ayala, A. Cl-Amidine Prevents Histone 3 Citrullination and Neutrophil Extracellular Trap Formation, and Improves Survival in a Murine Sepsis Model. J. Innate Immun. 2017, 9, 22–32. [Google Scholar] [CrossRef]
- Knight, J.S.; Subramanian, V.; O’Dell, A.A.; Yalavarthi, S.; Zhao, W.; Smith, C.K.; Hodgin, J.B.; Thompson, P.R.; Kaplan, M.J. Peptidylarginine deiminase inhibition disrupts NET formation and protects against kidney, skin and vascular disease in lupus-prone MRL/lpr mice. Ann. Rheum. Dis. 2015, 74, 2199–2206. [Google Scholar] [CrossRef]
- Willis, V.C.; Gizinski, A.M.; Banda, N.K.; Causey, C.P.; Knuckley, B.; Cordova, K.N.; Luo, Y.; Levitt, B.; Glogowska, M.; Chandra, P.; et al. N-α-benzoyl-N5-(2-chloro-1-iminoethyl)-L-ornithine amide, a protein arginine deiminase inhibitor, reduces the severity of murine collagen-induced arthritis. J. Immunol. 2011, 186, 4396–4404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ham, A.; Rabadi, M.; Kim, M.; Brown, K.M.; Ma, Z.; D’Agati, V.; Lee, H.T. Peptidyl arginine deiminase-4 activation exacerbates kidney ischemia-reperfusion injury. Am. J. Physiol. Physiol. 2014, 307, F1052–F1062. [Google Scholar] [CrossRef] [Green Version]
- Budanov, A.V.; Karin, M. p53 Target Genes Sestrin1 and Sestrin2 Connect Genotoxic Stress and mTOR Signaling. Cell 2008, 134, 451–460. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Dong, Z.-Z.; Zhang, J.; Teng, D.; Luo, X.; Li, D.; Zhou, Y. Peptidylarginine deiminases 4 as a promising target in drug discovery. Eur. J. Med. Chem. 2021, 226, 113840. [Google Scholar] [CrossRef]
- Luo, Y.; Knuckley, B.; Bhatia, M.; Pellechia, P.J.; Thompson, P.R. Activity-Based Protein Profiling Reagents for Protein Arginine Deiminase 4 (PAD4): Synthesis and in vitro Evaluation of a Fluorescently Labeled Probe. J. Am. Chem. Soc. 2006, 128, 14468–14469. [Google Scholar] [CrossRef] [Green Version]
- Bicker, K.L.; Thompson, P.R. The protein arginine deiminases: Structure, function, inhibition, and disease. Biopolymers 2013, 99, 155–163. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Knuckley, B.; Causey, C.P.; Pellechia, P.J.; Cook, P.F.; Thompson, P.R. Haloacetamidine-Based Inactivators of Protein Arginine Deiminase 4 (PAD4): Evidence that General Acid Catalysis Promotes Efficient Inactivation. ChemBioChem 2010, 11, 161–165. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Liu, C.; Lin, J. Theoretical study of the mechanism of protein arginine deiminase 4 (PAD4) inhibition by F-amidine. J. Mol. Graph. Model. 2015, 55, 25–32. [Google Scholar] [CrossRef] [PubMed]








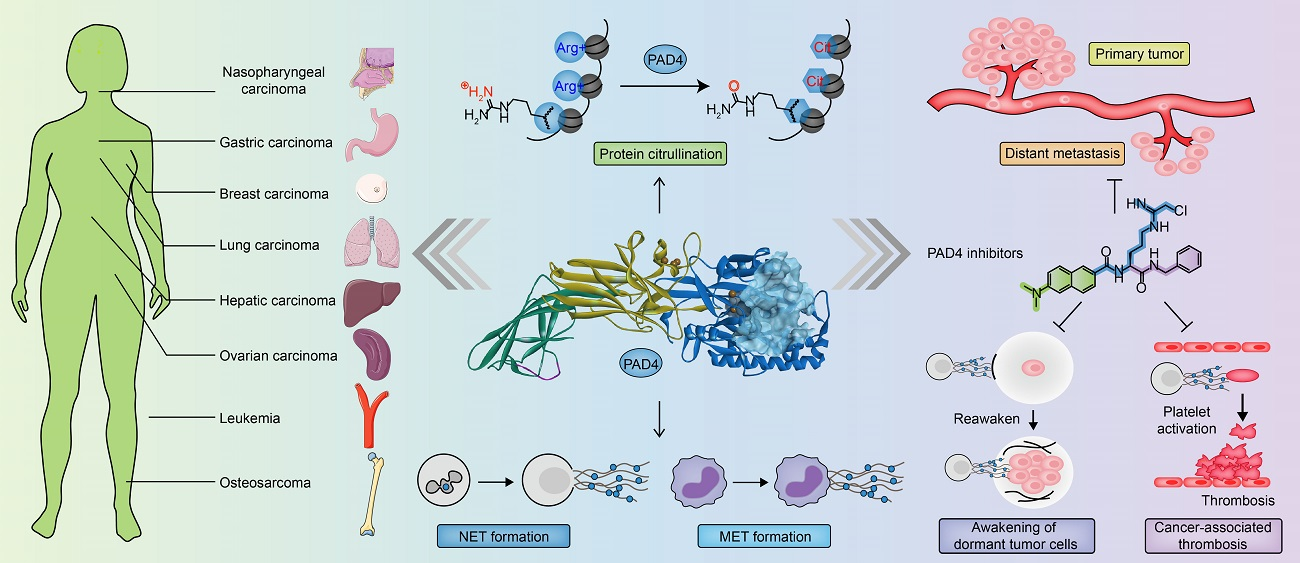{kind=link}
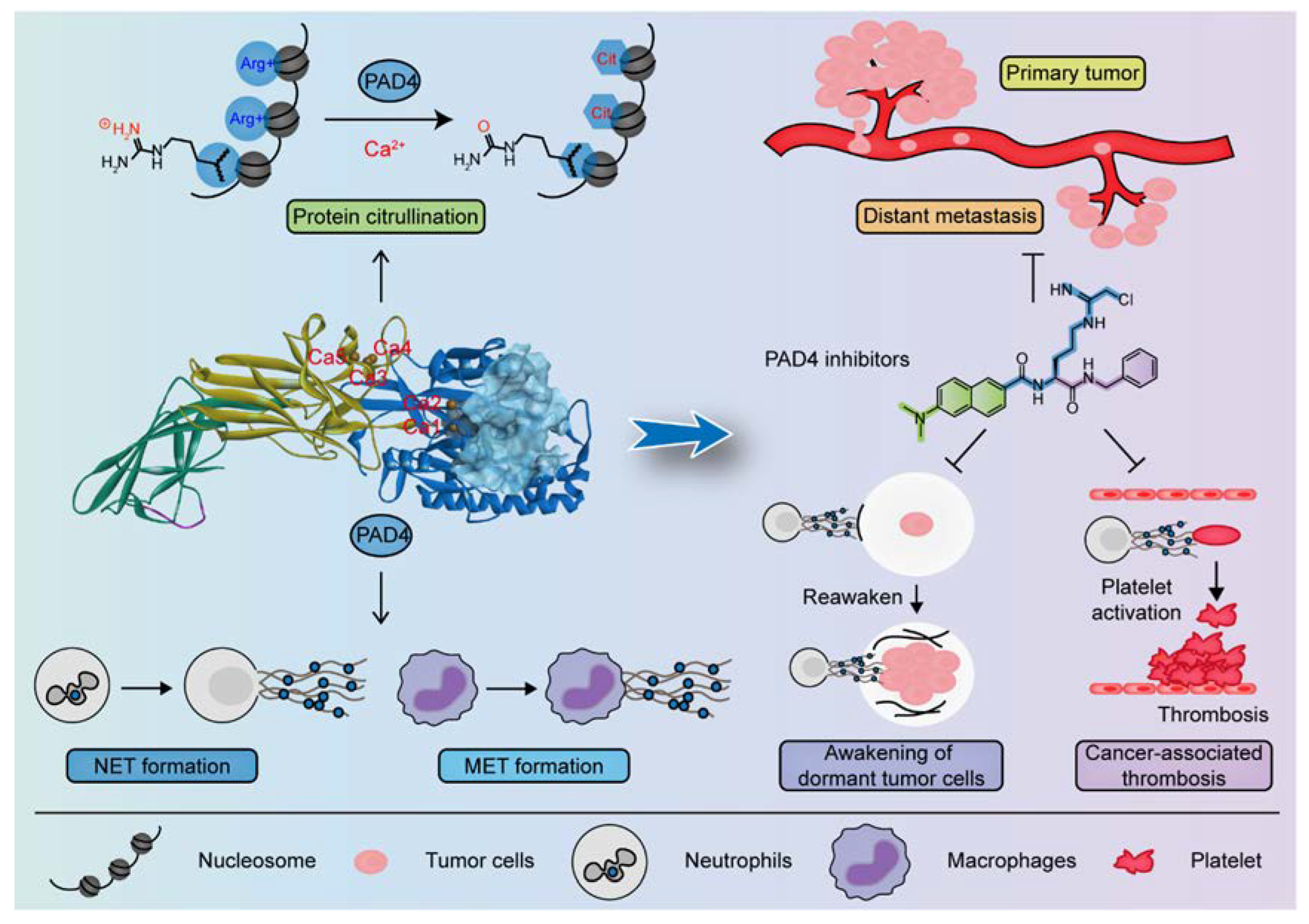{kind=link}
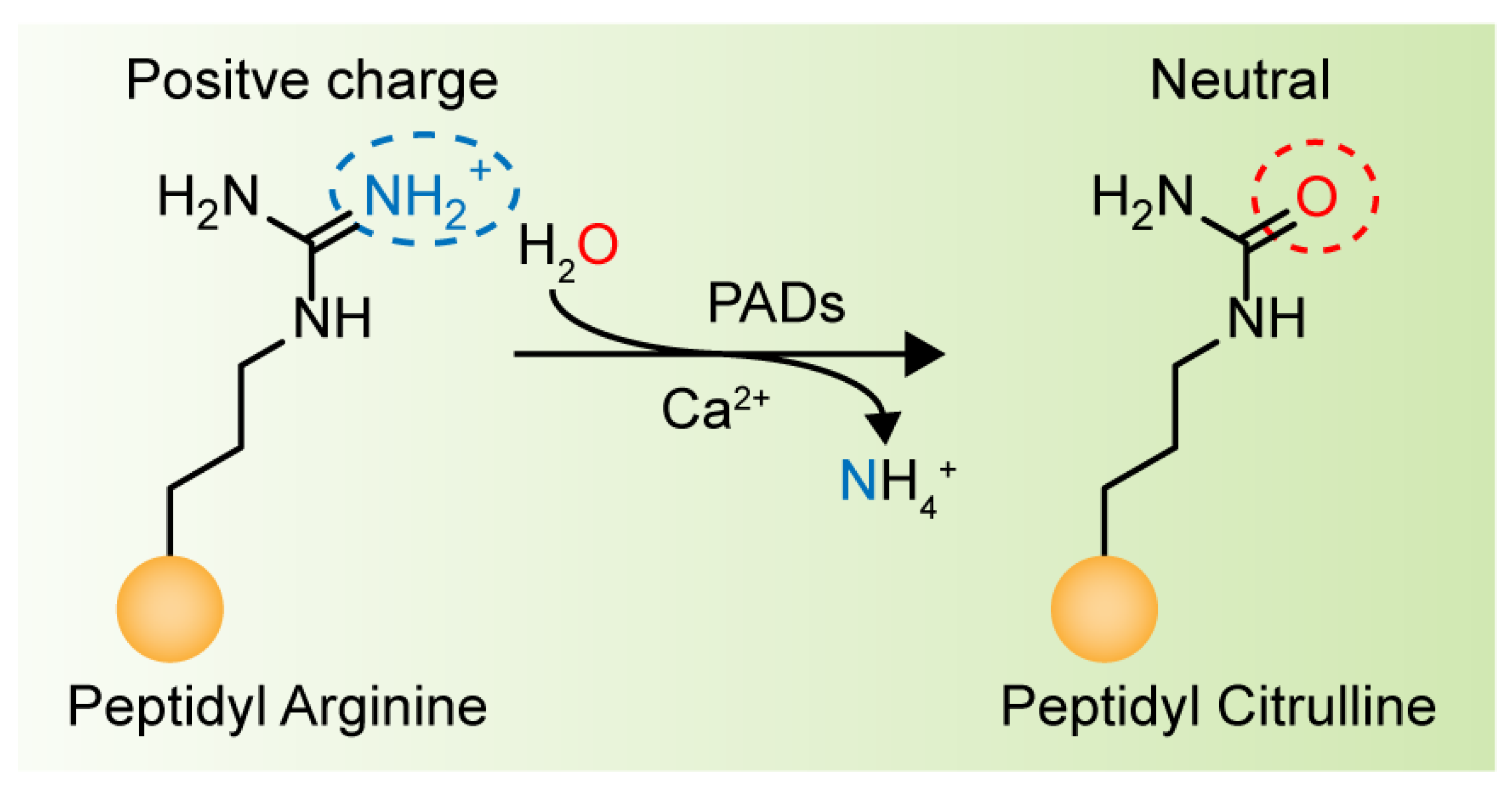{kind=link}
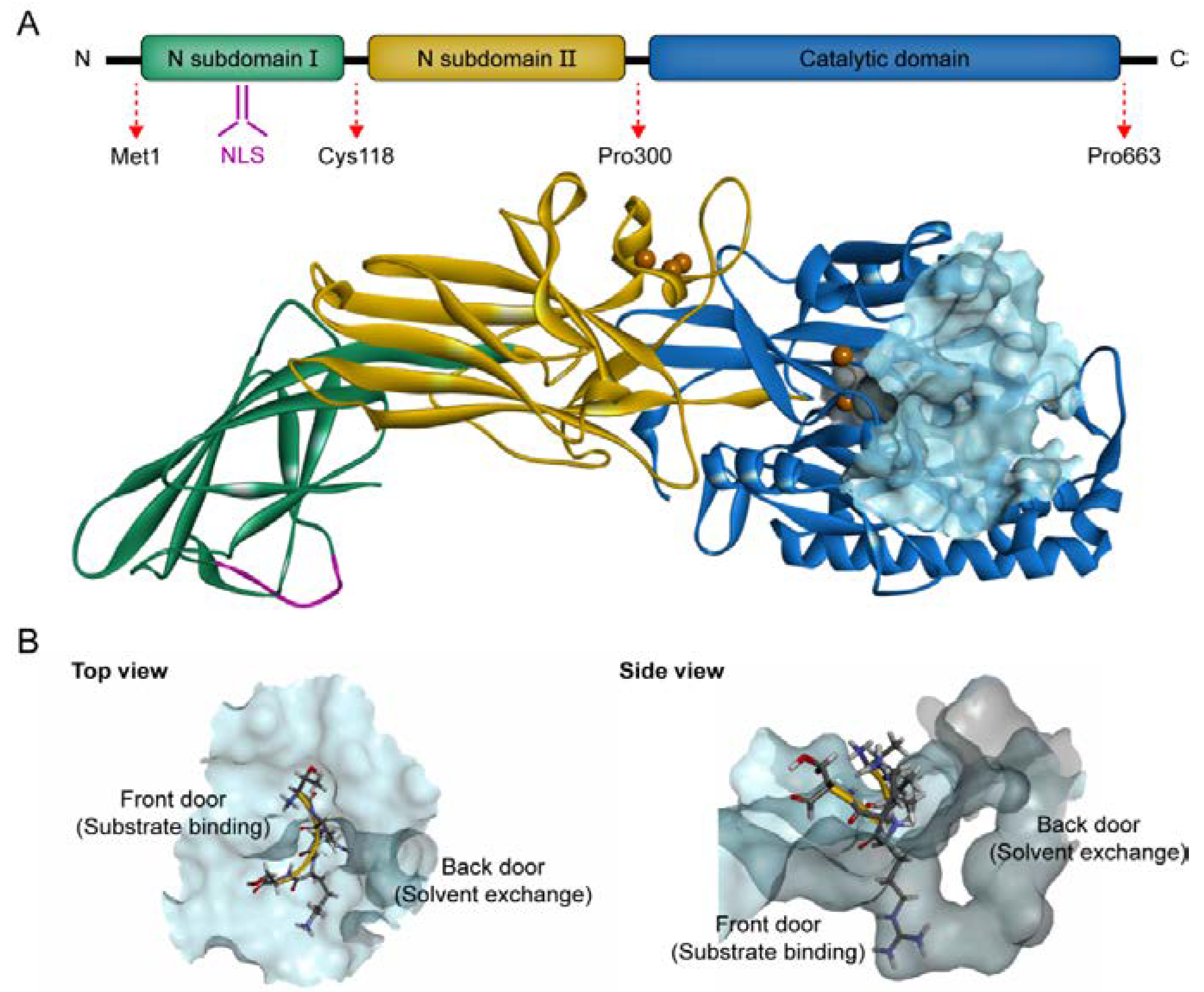{kind=link}
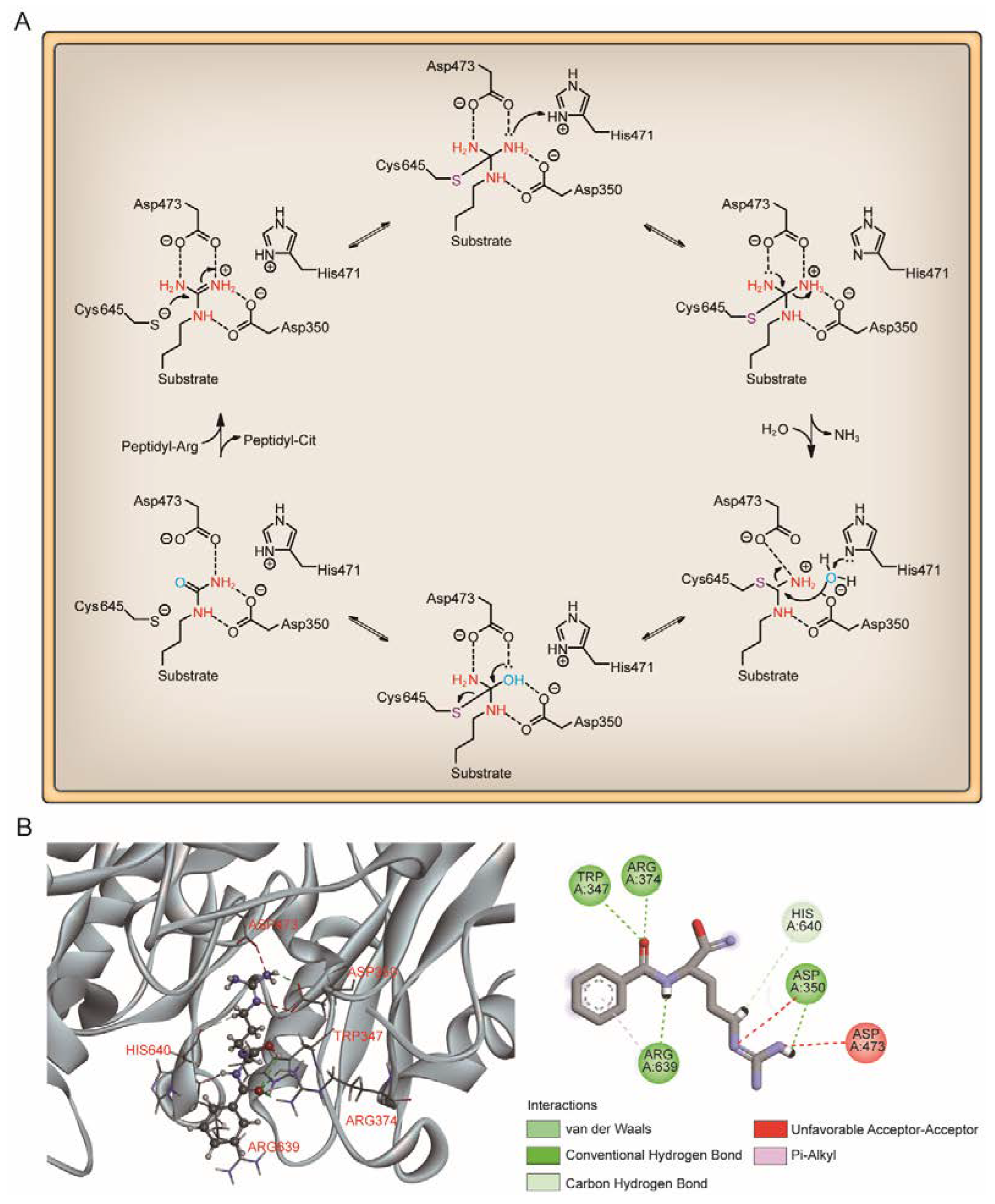{kind=link}
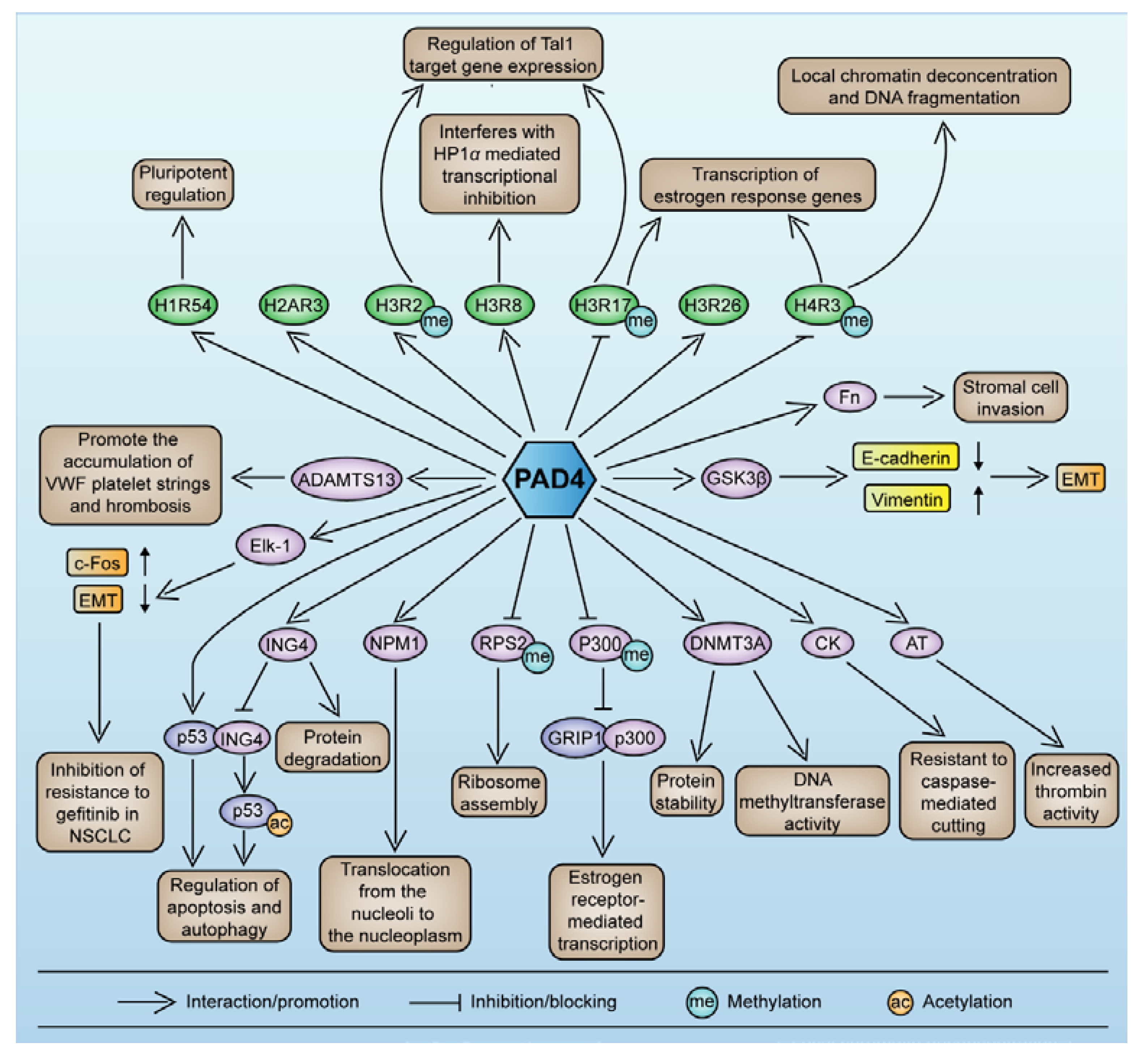{kind=link}
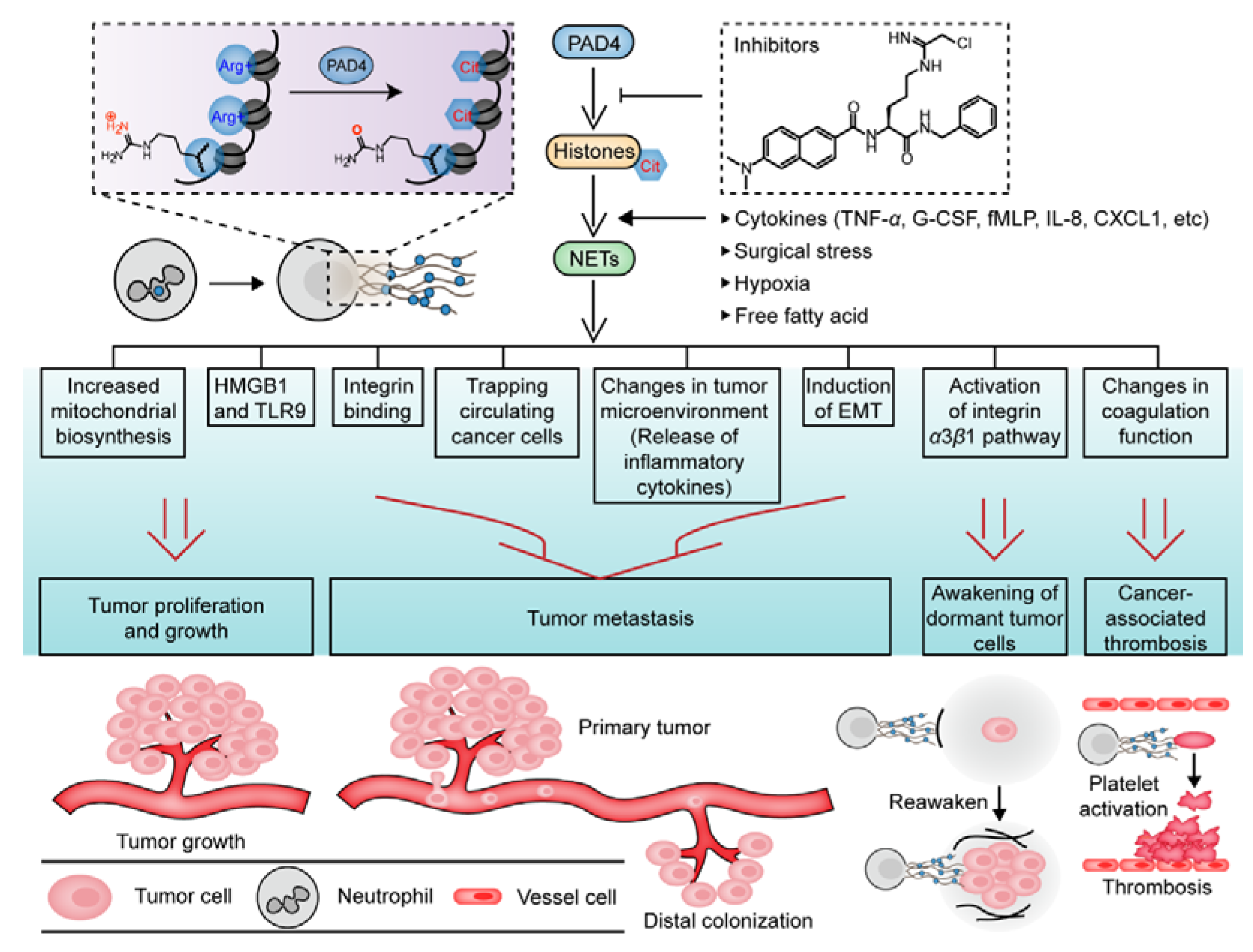{kind=link}
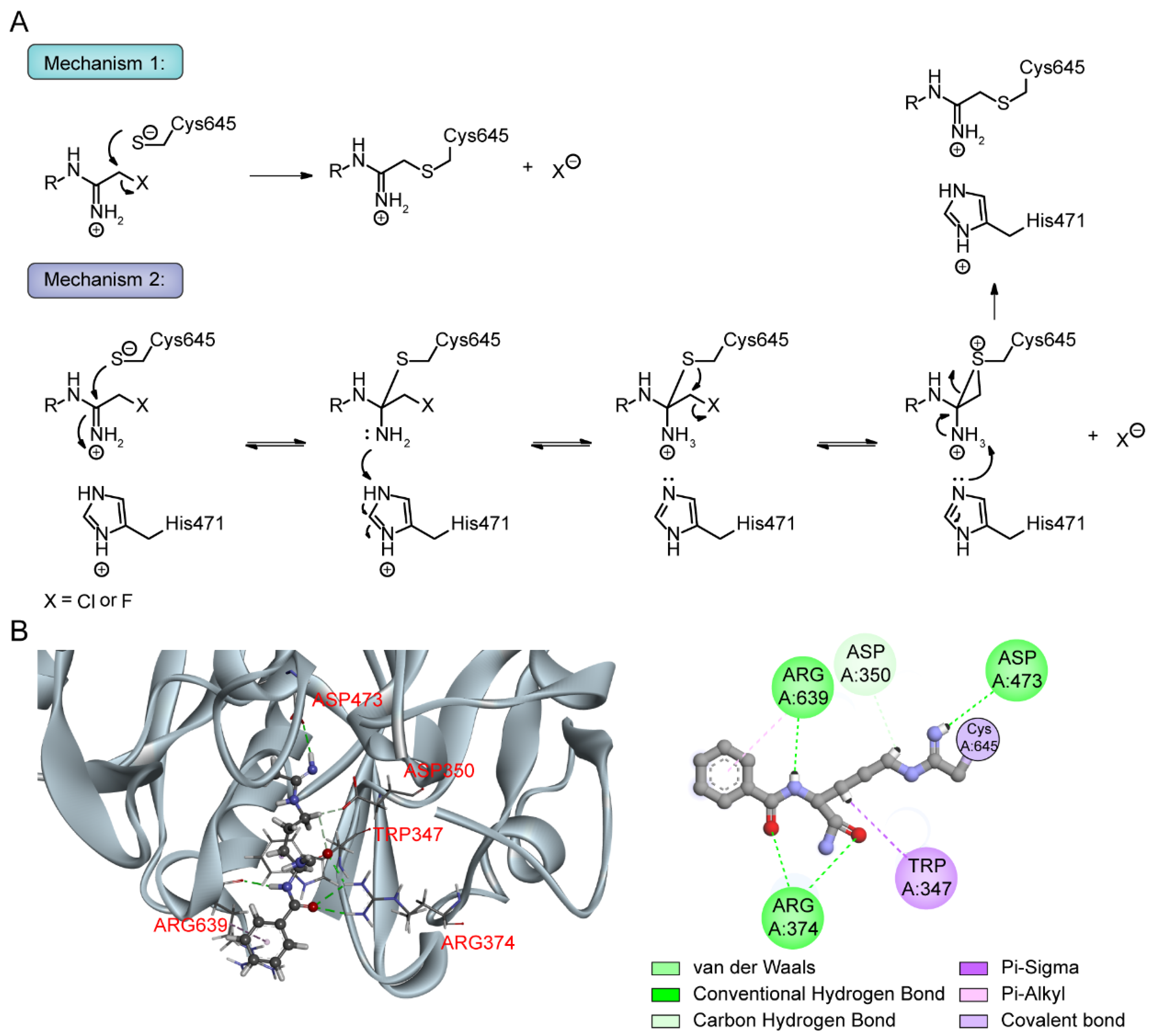{kind=link}
| PAD Isozyme | Expression Profile | Substrates | Physiological Roles | Pathological Roles |
|---|---|---|---|---|
| PAD1 | Immune cells, keratinocytes, hair follicles, epidermis, uterus | Keratin, filaggrin, S100A3, MEK1-ERK1/2-MMP2 signaling enzymes | Skin differentiation, terminal keratinocyte differentiation, the epithelial-mesenchymal transition (EMT) | Psoriasis [60], breast carcinomas [44] |
| PAD2 | Brain, skeletal muscle, spinal cord, glial cells, colon, pancreas, mammary gland, salivary gland, secretory gland, thymus, spleen, immune cells, bone marrow, yolk sac, epidermis, eye | MBP, GFAP, S100A3, CXCL10, CXCL11, Vimentin, β/γ-actin, histones (H3 and H4) | Oligodendrocyte differentiation, myelination, central nervous system (CNS) plasticity, transcription regulation, innate immunity, female reproduction | Multiple sclerosis [61,62], Alzheimer’s disease [63,64], rheumatoid arthritis [65,66], prion disease [67,68] and cancer |
| PAD3 | Epidermis, hair follicles | Filaggrin, vimentin, trichohyalin, S100A3 | Skin differentiation, hair follicle formation, terminal keratinocyte differentiation | Loss of barrier function and epidermal homeostasis [69], uncombable hair syndrome [70], glioblastoma multiforme [71] |
| PAD4 | Immune cells (neutrophils, monocytes, and macrophages), brain, uterus, joints, bone marrow, breast, epithelial cells, cancerous tissues | Histone H1 (R54Cit), H2A (R3Cit), H3 (R2Cit, R8Cit, R17Cit, R26Cit) and H4 (R3Cit, R23Cit), Lamin C, ING4, cytokeratin, antithrombin, ADAMTS13, GSK3β, Elk-1, collagen type I, fibronectin, p300/CBP, NPM1, RPS2, DNMT3A, FUS, EWS, NCF1, NCF2, S100A3 | Chromatin depolymerization, gene expression regulation, cell differentiation and apoptosis, innate immune response, NETosis process, tumor progression | rheumatoid arthritis [72,73], Alzheimer’s disease [74], multiple sclerosis [23], sepsis [24], inflammatory bowel disease [75], acute kidney injury [76,77], COVID-19 [25,78], cancer and thrombosis |
| PAD6 | Ovary, early embryo, ovum, oocyte, testis, thymus | α-Tubulin | Cytoskeletal recombination in eggs and early embryos, preimplantation cleavage, early embryonic development, female fertility, contraceptive drug targets | Infertility [58,79] |
| Cancer Type | Biological Model | Regulatory Mechanisms | Biological Effects | References |
|---|---|---|---|---|
| Gastric carcinoma (GC) | 1. Human gastric adenocarcinoma cell lines: MNK-45, SGC-7901, and AGS | PAD4 upregulates the expression of CXCR2, KRT14, TNF-α, and matrix metalloproteinases MMP2 and MMP9 | Suppression of PADI4 decreases cell proliferation and invasion, results in S-phase arrest and fewer GC cells in the G2/M-phase. The combination of PADI4 short interfering RNA (siRNA) and 5-FU significantly inhibits GC cell proliferation. | [28,29,232,233,234] |
| 2. Human GC cell line: MGC80-3 | PAD4 regulates IL-8 expression, upregulates Slug and vimentin expression, and downregulates E-cadherin expression | PAD4-induced epithelial-mesenchymal transition (EMT) and migration of GC cells are attenuated by inhibition of IL-8 expression. | ||
| Ovarian carcinoma (OC) | 1. Human OC cell lines: A2780 (p53 wild-type), and SKOV3 (p53-null) | Thirteen genes are related to cell proliferation, invasion, migration and apoptosis and their expression is differentially regulated | Silencing of PADI4 decreases the proliferation, invasion, and migration of p53-wt A2780 cells and increases the apoptosis rate, but only proliferation inhibition is observed with p53-null SKOV3 cells. | [28,29,235,236,237] |
| 2. Mouse OC cell line: ID8 Human OC cell line: ES2 | PAD4 mediates the formation of NETs and promotes the premetastatic omental niche to facilitate the implantation of ovarian cancer cells | Inhibition of PAD4 blocks NET formation and reduces omental colonization of OC cells. | ||
| Osteosarcoma (OS) | 1. Human OS cell lines: U2OS and Saos-2 2. Xenograft nude mice model (injected with cells containing a PADI4-expressing lentivirus vector) | PAD4 activates the Wnt/β-catenin and MEK/ERK signaling pathways and promotes cell proliferation | Inhibition of PAD4 can reduce the proliferation and colony formation ability of OS cells. | [28,29,38,238,239] |
| PAD4 downregulates the expression of E-cadherin in OS cells and induces the EMT | The expression of PAD4 is positively correlated with pulmonary metastasis in patients with OS. Inhibition of PAD4 expression interferes with the EMT of OS cells and inhibits their invasion and migration. | |||
| PAD4 and HDAC2 simultaneously bind to the p21 promoter and inhibit tumor cell apoptosis. | PAD4 inhibitor Cl-amidine or YW3-56 combined with the HDAC inhibitor SAHA exerts an additive effect in inhibiting the proliferation of U2OS cells. | |||
| Nasopharyngeal carcinoma (NPC) | 1. Human NPC cell lines: C666-1, 6-10B, and 5 2. Xenograft nude mice model (injected with C666-1 cells) | The LINC00324/miR-3164/PAD4 axis activates the PI3K/AKT pathway in NPC cells and promotes apoptosis and autophagy. | Silencing of PADI4 inhibits the proliferation and increases the apoptosis rates of NPC cells and induces autophagy. | [240,241,242] |
| PAD4 downregulates the proapoptotic factors Bax, Bak, Bim and Bad; upregulates the anti-apoptotic factors Bcl-2, Bcl-xl and Mcl-1; decreases the proportion of cells in the G0/G1 phase; and promotes cell cycle progression. | PAD4 inhibitors GSK484 and YW3-56 restore the radiosensitivity of NPC cells; suppress cell proliferation, invasion, and migration; and significantly inhibit tumor growth in mice. | |||
| Hepatic carcinoma | 1. Human hepatocellular carcinoma (HCC) cell lines: SMMC-7721 and HepG2 2. Xenograft nude mice model (injected with SMMC-7721 cells) | PAD4 causes chemotherapy resistance of HCC cells by inducing autophagy. | Autophagy inhibitor effectively restores the sensitivity of HCC cells to chemotherapy in vitro and in vivo. | [28,29,186,243,244,245,246] |
| 3. The STAM model of nonalcoholic steatohepatitis (NASH)/HCC | Elevated free fatty acids stimulate NET formation in vitro | NET inhibition by DNase Ι or PADI4 knockout (KO) changes the pattern of liver inflammation and reduces the progression of liver cancer in NASH patients. | ||
|
4. Human HCC cell lines: HepG2 and MHCC97H 5. Mouse HCC cell line: Hepa1-6 C57BL/6 model mice (injected with Hepa1-6) with lipopolysaccharide (LPS)-induced NET formation | NETs activate the TLR4/9-COX2 signaling pathway in HCC cells and induce the inflammatory response, change the tumor inflammatory microenvironment, and promote tumorigenesis and metastasis cascades | The combination of DNase Ι with the anti-inflammatory drugs aspirin/hydroxychloroquine (HCQ) effectively attenuates HCC metastasis in mouse models. | ||
|
6. Human HCC cell lines: HuH7 and MHCC97L 7. Mouse HCC cell line: Hepa1-6 Xenograft nude mice model (injected with HuH7 or 97L cells) 8. In a coinjection experiment (injected intravenously with HuH7 cells and human neutrophils) | HCC cell-derived IL-8 triggers NET formation mediated through the NADPH oxidase pathway | NET-associated cathepsin G promotes HCC metastasis in vitro and in vivo. | ||
| Colon Cancer |
1. Human colon cancer cell lines: SW480, SW620, DLD-1, HT29, HCT116, and LoVo 2. Mouse colon cancer cell line: MC38 3. SCID mouse models of hepatic metastases (injected with HT29, HCT116 or LoVo cells) 4. C57BL/6 mouse models of hepatic metastases (injected with MC38 cells) | PAD4 mediates citrullination of extracellular matrix (ECM) components; promotes colorectal cancer (CRC) cell adhesion; inhibits CRC cell mobility; increases the expression of the epithelial marker E-cadherin; and promotes the mesenchymal–epithelial transition (MET) | The PAD inhibitor BB-Cl-amidine or PADI4 KO inhibits ECM citrullination, enhances mesenchymal cell marker expression, and inhibits liver metastasis growth. DNase Ι or PAD4 inhibitor YW4-03 inhibits NETs-mediated proliferation, adhesion, migration, and invasion of cancer cells. PAD4-KO tumors exhibit decreased mitochondrial density, mitochondrial DNA, ATP production and mitochondrial biogenesis proteins, causing decreased cell proliferation, an increased apoptosis rate and increased oxidative stress. | [86,188,189,247] |
| Lung carcinoma | Non-small-cell lung cancer (NSCLC) cell lines: 95D and A549 Xenograft nude mice model (injected with A549 cells) | Surgical stress promotes NET formation, alters the inflammatory microenvironment, and releases HMGB1 to activate TLR9-dependent pathways in cancer cells | The Au nanodrug delivery system YW3-56 targets tumors and when combined with photothermal therapy significantly inhibits tumor growth and lung metastasis in mice, showing good biosafety. | [28,29,122,248,249,250] |
| NSCLC cell lines: HCC827 and H1650 Gefitinib-resistant cell lines: HCC827/G and H1650/G | NE released from NETs activates the TLR4-p38-PGC-1α axis in cancer cells to increase mitochondrial biosynthesis. | PAD4 expression is decreased in gefitinib-resistant cell lines, and the upregulation of PAD4 expression reverses the resistance of NSCLC cell lines to gefitinib by inhibiting Elk-1 expression and the EMT. | ||
| Breast carcinoma | 1. Human breast carcinoma cell lines: MCF-7 and T47D | PAD4 citrullinates the N-terminal domain of GSK3β, promoting nuclear GSK3β accumulation, inhibiting Smad stability and TGF-β signaling, thereby ensuring maintenance of epithelial phenotypes | Silencing of PADI4 in MCF-7 cells induces the EMT and greater invasiveness of tumors, as shown in xenograft experiments. | [28,29,89,90,251,252] |
| 2. Human breast carcinoma cell lines: MCF-7 and MCF-7/ADR | PAD4 increases the nuclear levels of GSK3β and p53, which activate proapoptotic gene expression and downregulate MDR1 expression | Restoring PAD4 expression in MCF-7/ADR cells reverses drug resistance by inducing apoptosis. | ||
|
3. Human breast carcinoma cell lines: MCF10CA1h, MCF10CA1a, MCF-7E, and MDA-MB-231 LM2 4. Female athymic NCI nu/nu mouse model (mammary fat pad injection with MCF-7 cells) | PAD4 downregulates the expression of stemness master transcription factors NANOG and OCT4 by reducing the transcriptional activation mediated through H3R17me2a marked loci | Endogenous PAD4 activity limits the amount of cancer stem cells (CSCs) residing in multiple breast cancer models in vitro and in vivo. | ||
|
5. Mouse triple-negative breast cancer (TNBC) cell lines: 4T1 and 67NR 6. Allograft female BALB/c mice model (with 4T1 cells injected into mammary fat pad) 7. BALB/c mouse model of pulmonary metastasis (with 4T1 cells injected intravenously) | PAD4 mediates histone hypercitrullination, and cancer extracellular chromatin network (CECN) release from 4T1 cells | PADI4 KO in 4T1 cells inhibits tumor growth and lung metastasis in mice. Degradation of extracellular DNA by DNase Ι inhibits pulmonary metastasis of PADI4-induced 4T1 cells. | [222,253,254] | |
|
8. Human TNBC cell lines: MDA-MB-231 and BT-549 9. Xenograft nude mouse model (injected with MDA-MB-231 cells) | GSK484 facilitates TNBC cell apoptosis, offsets irradiation (IR)-induced upregulation of ATG5 and ATG7 expression and downregulation of p62 expression, while enhancing DNA damage. | GSK484 pretreatment enhances IR-induced inhibition of cell proliferation, migration, and invasion. The combination of IR and GSK484 significantly inhibits tumor growth and metastasis. | ||
|
10. Human TNBC cell line: MDA-MB-231 11. Xenograft SCID mouse model (MDA-MB-231 cells injected with into the mammary fat pad) | Hypoxia-inducible factors (HIFs) recruit PAD4 to the hypoxia response elements (HREs) of HIF target genes under hypoxic conditions and stimulate transcriptional activation | PAD4 promotes breast cancer angiogenesis and tumor growth. | ||
|
12. Dormancy model (inflammation induced through intranasal administration of LPS): 13. BALB/c mice (injected intravenously with mCherry-luciferase-D2.0R cells), nude mice (injected intravenously with MCF-7 cells) 14. BALB/c mouse model of pulmonary metastasis (injected intravenously with 4T1 or MDA-MB-231 cells) | NETs release NE and MMP9 and reshape laminin, activating the integrin α3β1 pathway and FAK/ERK/MLCK/YAP signaling to awaken dormant cancer cells. | Sustained inflammation induces NET formation and promotes the awakening of dormant cancer cells. The PAD4 inhibitor GSK484 and DNase I prevent the reawakening of dormant D2.0R and MCF-7 cancer cells. Ozone exposure increases lung neutrophil inflammation in mice, promoting NET release and cancer cells colonization of the lung. | [200,255] | |
| Leukemia | 1. Acute promyelocytic leukemia (APL) cells: HL-60 (PML-RARα- negative) and NB4 (PML-RARα- positive) | PAD4 inhibition in APL inhibits cell differentiation mediated through the functional PAD4/SOX4/PU.1 axis | All-trans-retinoic acid (ATRA) treatment restores PAD4 expression to promote its nuclear translocation in APL cells, and it promotes the differentiation of leukemia cells by regulating PU.1 expression. | [109,113,256,257] |
| 2. Bone marrow cells isolated from PAD4-KO mice and wt C57BL/6 mice | PAD4 regulates the proliferation of MPP cells by binding LEF1 and HDAC1 to control c-myc expression. | Compared with cells from wt mice, LSK cells, especially MPP cells, proliferate abnormally in PAD4-KO mice. | ||
| 3. Human cell lines: HL-60, Jurkat T, and Jurkat-Tet-On | PAD4 overexpression increases the accumulation of p53 in hematopoietic cells, stimulates the transcription of P21 and Bax, and induces apoptosis and cycle arrest. | PAD4 reduces HL-60 and Jurkat cellular activity in a dose- and time-dependent manner |
| Type | Inhibitor | Structure | IC50 | Reference |
|---|---|---|---|---|
| Reversible inhibitors | Taxol |  | IC50 = ≈5 mM | [94,290] |
| Bz-ADMA |  | IC50 = 0.4 mM | [31,291] | |
| Streptomycin |  | IC50 = 1.8 mM | [31,292] | |
| Minocycline |  | IC50 = 0.62 mM | [31,40,292] | |
| Chlortetracycline |  | IC50 = 100 μM | [31,40,292] | |
| GSK199 |  | IC50 (0 mM Ca) = 200 nM; IC50 (2 mM Ca) = 1.0 μM | [31,40,168] | |
| GSK484 |  | IC50 (0 mM Ca) = 50 nM; IC50 (2 mM Ca) = 250 nM | [31,40,168] | |
| Inh-Dap |  | IC50 = 243.2 ± 2.4 μM | [293] | |
| SC97362 |  | IC50 = 1.88 ± 0.26 μM | [294] | |
| Irreversible inhibitors | NSC95397 |  | kinact/KI (M-1·min−1) PAD1: 175 PAD2: 1600 PAD3: 9150 PAD4: 4530 | [40,295] |
| Streptonigrin |  | kinact/KI (M−1·min−1) PAD1: 3700 PAD2: 12,000 PAD3: 3500 PAD4: 440,000 | [40,296] | |
| Cl-amidine |  | IC50 = 6 μM kinact/KI (M−1 min−1) PAD1: 37,000 PAD2: 1200 PAD3: 2000 PAD4: 13,000 | [31,40,297,298] | |
| F-amidine |  | IC50 = 22 μM kinact/KI (M−1·min−1) PAD1: 2800 PAD2: 380 PAD3: 170 PAD4: 3000 | [31,40,297,298] | |
| o-Cl-amidine |  | IC50 = 2.2 μM kinact/KI (M−1·min−1) PAD1: 106,400 PAD2: 14,100 PAD3: 10,345 PAD4: 38,000 | [31,40,298] | |
| o-F-amidine |  | IC50 = 1.9 μM kinact/KI (M−1·min−1) PAD1: 180,900 PAD2: 7500 PAD3: 6700 PAD4: 32,500 | [31,40,298] | |
| BB-Cl-amidine |  | kinact/KI (M−1·min−1) PAD1: 16,100 PAD2: 4100 PAD3: 6800 PAD4: 13,300 | [31,40,299] | |
| BB-F-amidine |  | kinact/KI (M−1·min−1) PAD1: 900 PAD2: 1200 PAD3: 3400 PAD4: 3750 | [31,40,299] | |
| TDFA |  | IC50 = 2.3 μM kinact/KI (M−1·min−1) PAD1: 1700 PAD2: 500 PAD3: 400 PAD4: 26,000 | [31,40,300] | |
| TDCA |  | IC50 = 3.4 μM kinact/KI (M−1·min−1) PAD1: 21,000 PAD2: 300 PAD3: 920 PAD4: 24,000 | [31,40,300] | |
| YW3-56 |  | IC50 = 1–5 μM | [111] | |
| ZD-E-1M |  | IC50 = 2.39 μM | [301] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhu, D.; Lu, Y.; Wang, Y.; Wang, Y. PAD4 and Its Inhibitors in Cancer Progression and Prognosis. Pharmaceutics 2022, 14, 2414. https://doi.org/10.3390/pharmaceutics14112414
Zhu D, Lu Y, Wang Y, Wang Y. PAD4 and Its Inhibitors in Cancer Progression and Prognosis. Pharmaceutics. 2022; 14(11):2414. https://doi.org/10.3390/pharmaceutics14112414
Chicago/Turabian StyleZhu, Di, Yu Lu, Yanming Wang, and Yuji Wang. 2022. "PAD4 and Its Inhibitors in Cancer Progression and Prognosis" Pharmaceutics 14, no. 11: 2414. https://doi.org/10.3390/pharmaceutics14112414