
Protective Effects of Choline against Inflammatory Cytokines and Characterization of Transport in Motor Neuron-like Cell Lines (NSC-34)

Abstract
:1. Introduction
2. Materials and Methods
2.1. Radioisotope and Chemical Compounds
2.2. NSC-34 Cell Lines
2.3. Culture of NSC-34 Cell Lines
2.4. Uptake Study of [3H]Choline on NSC-34 Cell Lines
2.5. Uptake Study under Sodium- and Potassium-Free Conditions
2.6. Kinetic Parameter Estimation of [3H]Choline Uptake in NSC-34 Cell Lines
2.7. Real-Time PCR Analysis
2.8. Cytokines & Nerve Growth Factor Study Methods
3. Results
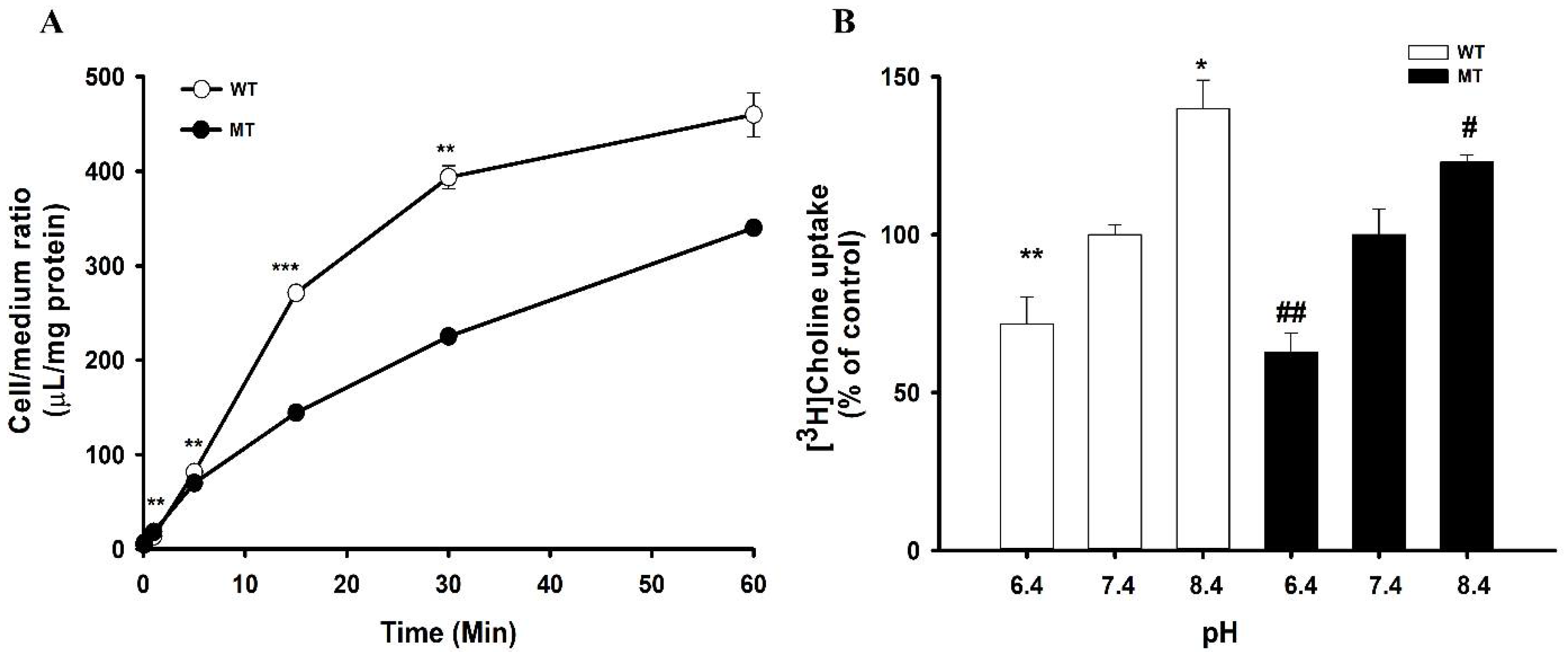
3.1. Time Course and Effect of pH on [3H]Choline Transport in NSC-34 Cell Lines
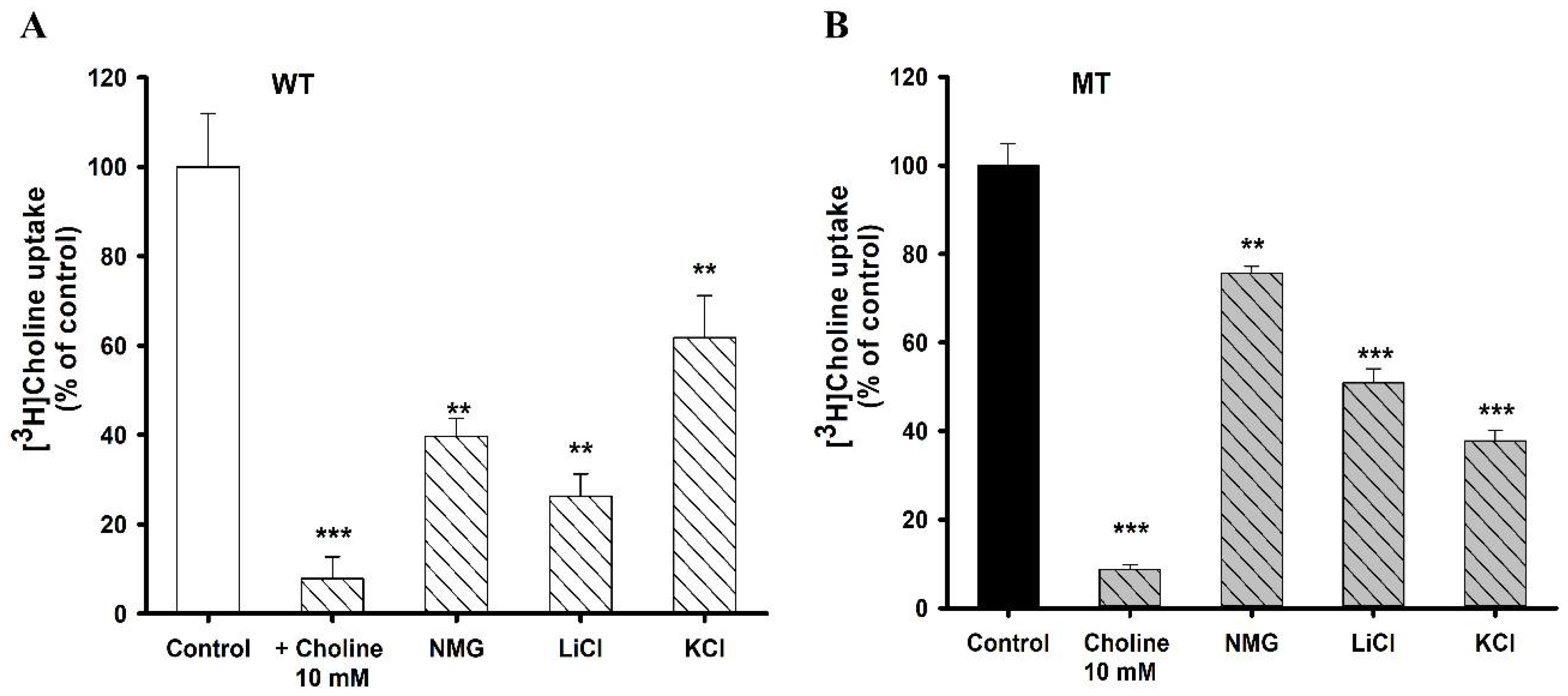
3.2. Na+ and K+ ion Dependence on [3H]Choline Uptake in NSC-34 Cell Lines
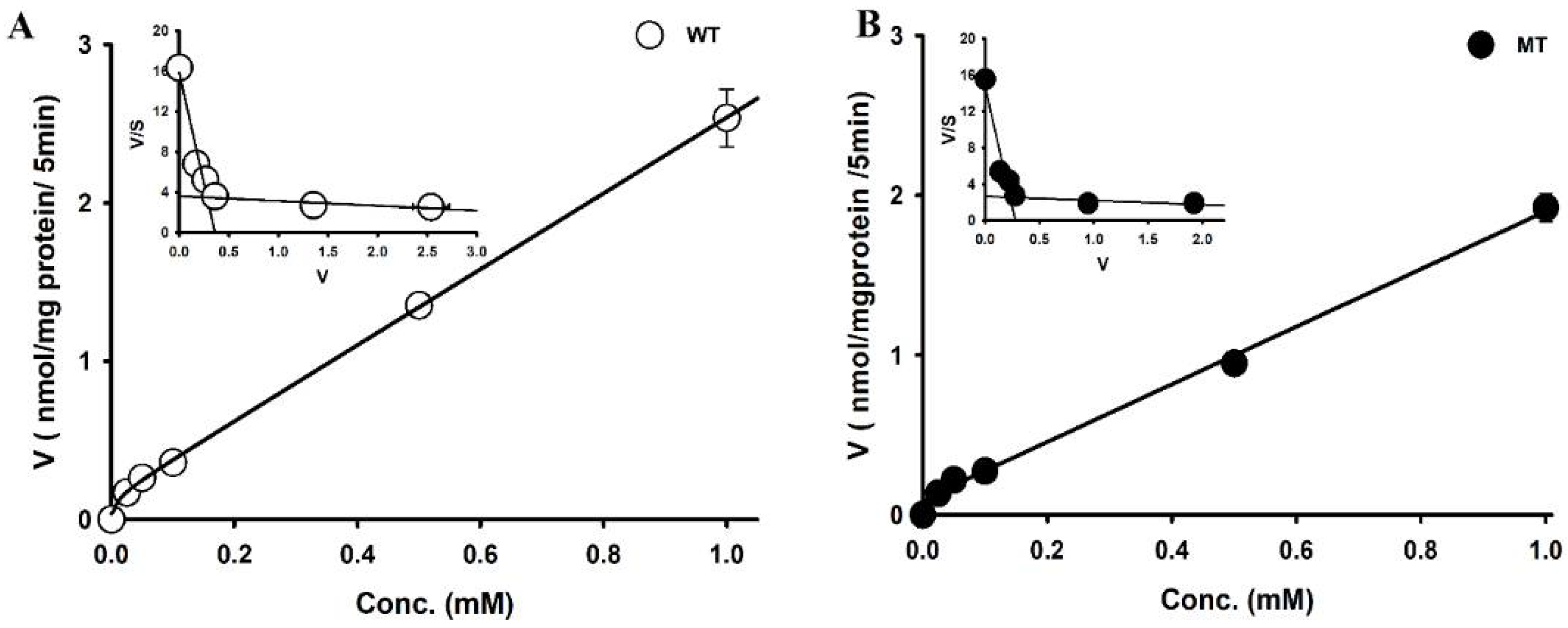
3.3. Transport Kinetics Parameters of [3H]Choline in NSC-34 Cell Line
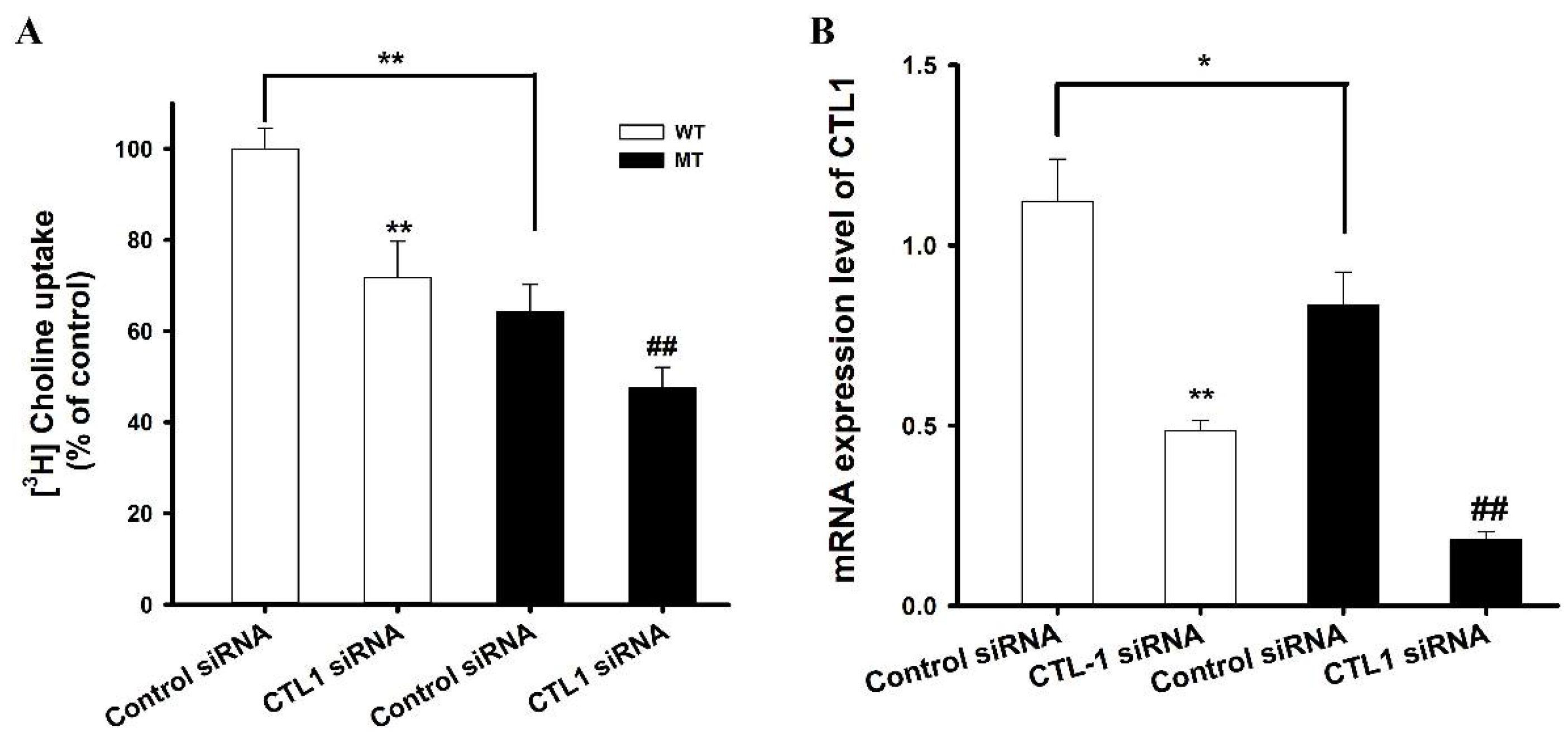
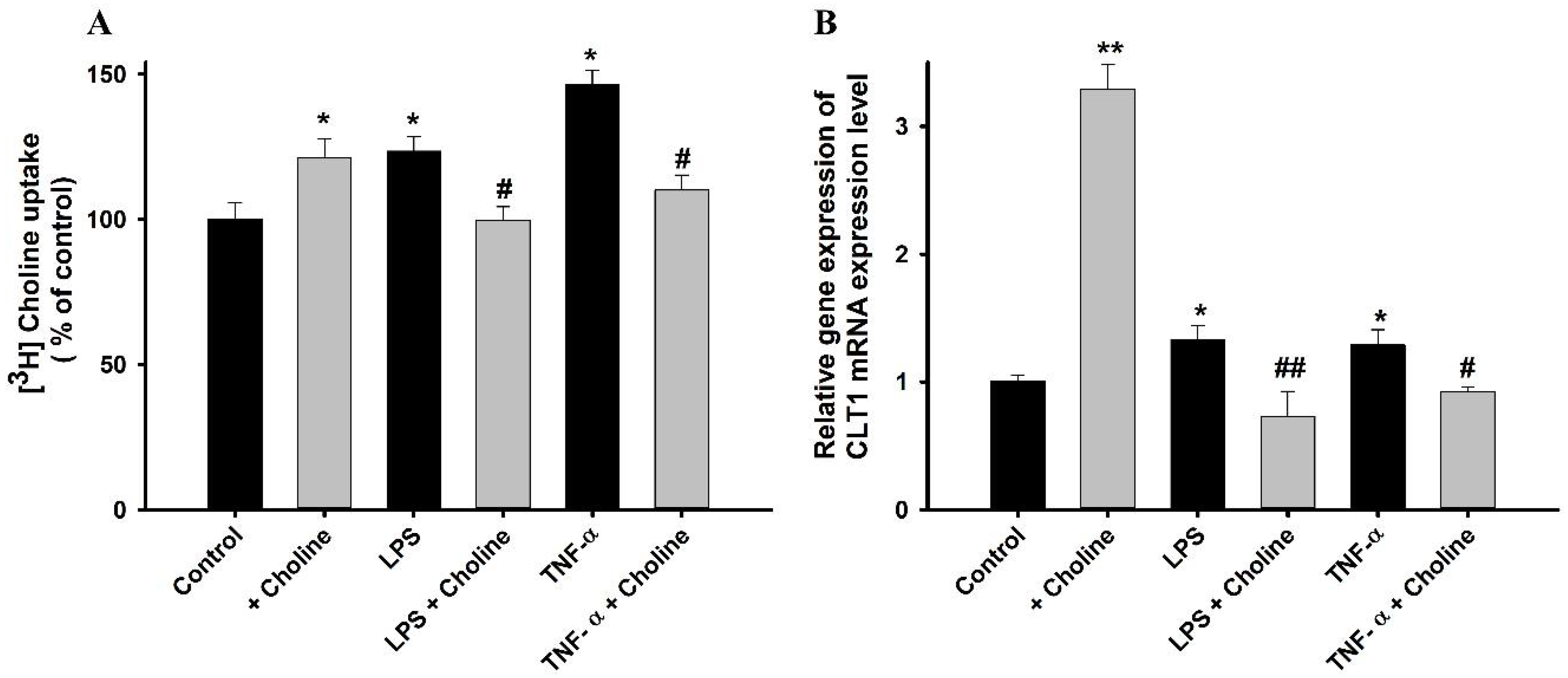
3.4. RT-PCR Analysis of CTL1 Expression in ALS Model Cell Lines
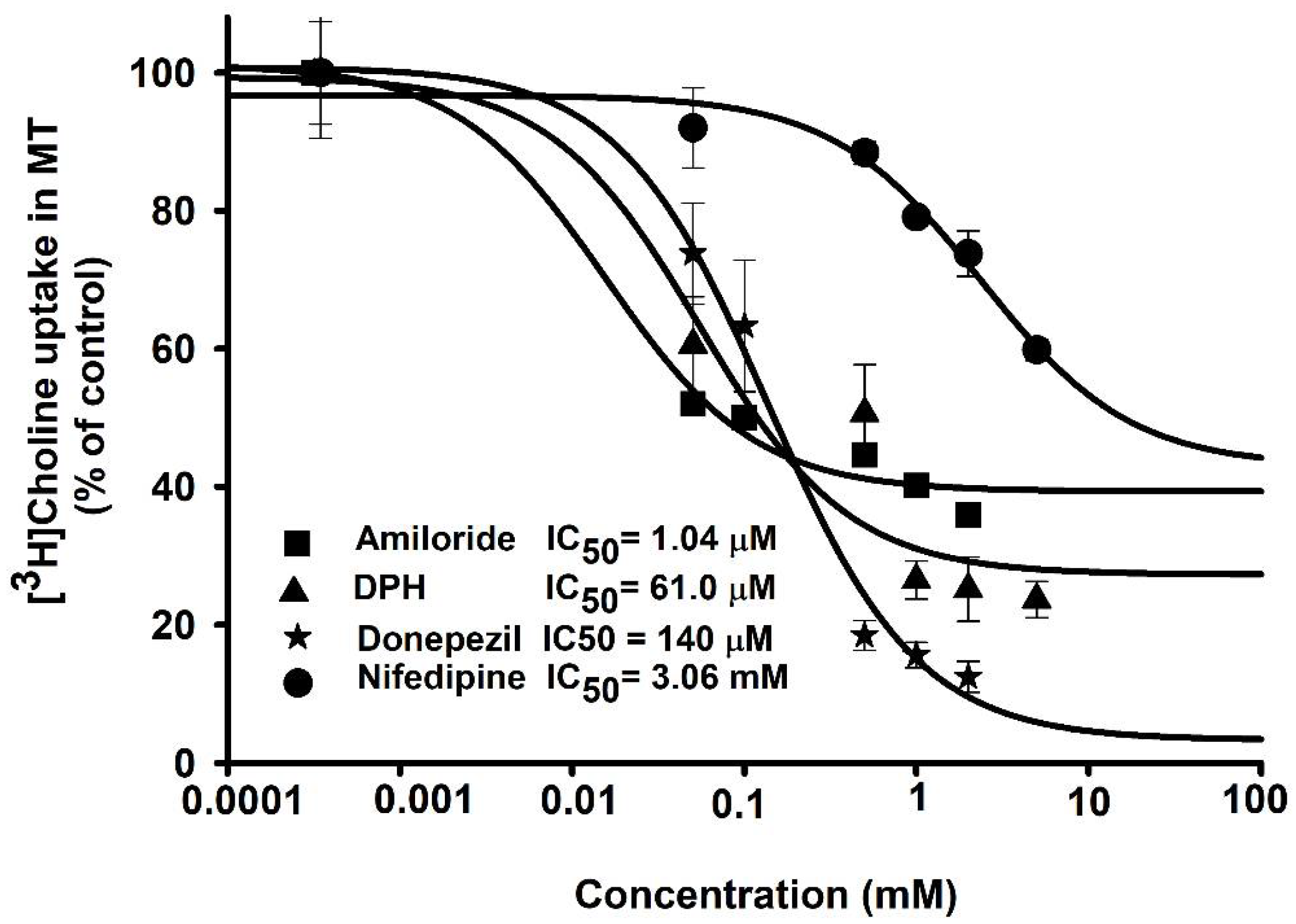
3.5. Inhibitory Effects of Pharmacological Compounds on Choline Transport by ALS Model Cell Lines
3.6. IC50 of Drugs in Disease Model Cell Lines
3.7. Uptake of [3H]Choline Increases with NGF
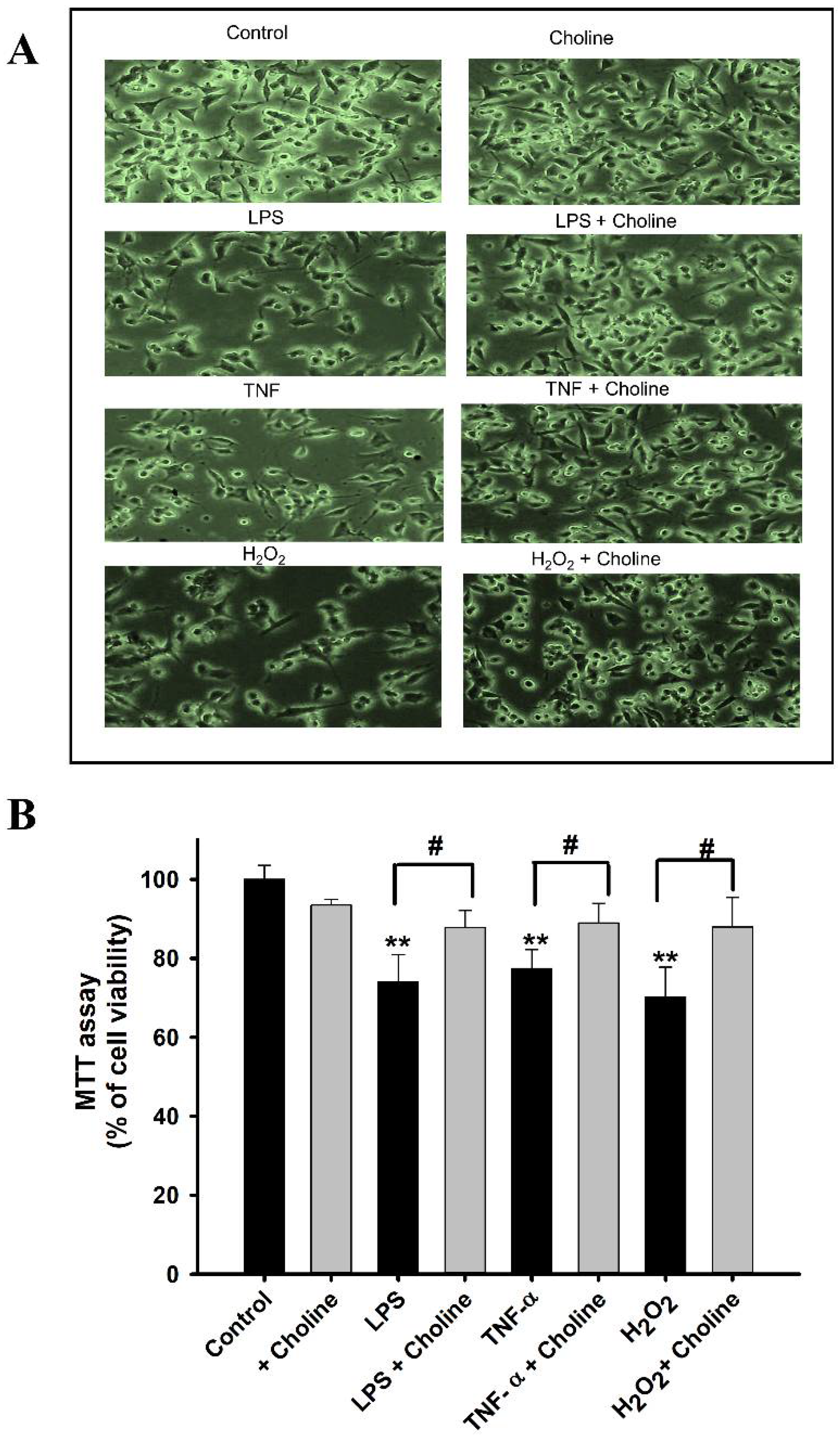
3.8. Effect of Choline on the Cytotoxicity Induced by LPS and TNF-α in ALS Model Cell Lines
3.9. Cell Viability Analysis by MTT Assay
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bekdash, R.A. Choline, the brain and neurodegeneration: Insights from epigenetics. Front. Biosci.-Landmark 2018, 23, 1113–1143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Boillée, S.; Vande Velde, C.; Cleveland, D.W.W. ALS: A Disease of Motor Neurons and Their Nonneuronal Neighbors. Neuron 2006, 52, 39–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Xu, G.; Borchelt, D.R. High molecular weight complexes of mutant superoxide dismutase 1: Age-dependent and tissue-specific accumulation. Neurobiol. Dis. 2002, 9, 139–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tateno, M.; Sadakata, H.; Tanaka, M.; Itohara, S.; Shin, R.M.; Miura, M.; Masuda, M.; Aosaki, T.; Urushitani, M.; Misawa, H.; et al. Calcium-permeable AMPA receptors promote misfolding of mutant SOD1 protein and development of amyotrophic lateral sclerosis in a transgenic mouse model. Hum. Mol. Genet. 2004, 13, 2183–2196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tateno, M.; Kato, S.; Sakurai, T.; Nukina, N.; Takahashi, R.; Araki, T. Mutant SOD1 impairs axonal transport of choline acetyltransferase and acetylcholine release by sequestering KAP3. Hum. Mol. Genet. 2009, 18, 942–955. [Google Scholar] [CrossRef]
- Goncharova, P.S.; Davydova, T.K.; Popova, T.E.; Novitsky, M.A.; Petrova, M.M.; Gavrilyuk, O.A.; Al-Zamil, M.; Zhukova, N.G.; Nasyrova, R.F.; Shnayder, N.A. Nutrient effects on motor neurons and the risk of amyotrophic lateral sclerosis. Nutrients 2021, 13, 3804. [Google Scholar] [CrossRef]
- Nowicka, N.; Juranek, J.; Juranek, J.K.; Wojtkiewicz, J. Risk factors and emerging therapies in amyotrophic lateral sclerosis. Int. J. Mol. Sci. 2019, 20, 2616. [Google Scholar] [CrossRef] [Green Version]
- Ray, B.; Bailey, J.A.; Simon, J.R.; Lahiri, D.K. High-affinity choline uptake (HACU) and choline acetyltransferase (ChAT) activity in neuronal cultures for mechanistic and drug discovery studies. Curr. Protoc. Neurosci. 2012, 1, 1–24. [Google Scholar] [CrossRef] [Green Version]
- Zeisel, S.H.; Da Costa, K.A. Choline: An essential nutrient for public health. Nutr. Rev. 2009, 67, 615–623. [Google Scholar] [CrossRef] [Green Version]
- Bekdash, R.A. Neuroprotective Effects of Choline and Other Methyl Donors. Nutrients 2019, 11, 2995. [Google Scholar] [CrossRef]
- Blusztajn, J.; Slack, B.; Mellott, T. Neuroprotective Actions of Dietary Choline. Nutrients 2017, 9, 815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choline transporters. Br. J. Pharmacol. 2009, 158, 8767. [CrossRef]
- O’Regan, S.; Traiffort, E.; Ruat, M.; Cha, N.; Compaoré, D.; Meunier, F.M. An electric lobe suppressor for a yeast choline transport mutation belongs to a new family of transporter-like proteins. Proc. Natl. Acad. Sci. USA 2000, 97, 1835–1840. [Google Scholar] [CrossRef] [Green Version]
- Nakamura, T.; Fujiwara, R.; Ishiguro, N.; Oyabu, M.; Nakanishi, T.; Shirasaka, Y.; Maeda, T.; Tamai, I. Involvement of choline transporter-like proteins, CTL1 and CTL2, in glucocorticoid-induced acceleration of phosphatidylcholine synthesis via increased choline uptake. Biol. Pharm. Bull. 2010, 33, 691–696. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kang, Y.; Lee, K.; Lee, N.; Terasakp, T. Donepezil, Tacrine and -Phenyl-n-tert-Butyl Nitrone ( PBN ) Inhibit Choline Transport by Conditionally Immortalized Rat. Arch. Pharm. Res. 2005, 28, 443–450. [Google Scholar] [CrossRef] [PubMed]
- Lee, N.Y.; Choi, H.M.; Kang, Y.S. Choline Transport via Choline Transporter-like Protein 1 in Conditionally Immortalized Rat Syncytiotrophoblast Cell Lines TR-TBT. Placenta 2009, 30, 368–374. [Google Scholar] [CrossRef]
- Martínez, H.J.; Dreyfus, C.F.; Miller Jonakait, G.; Black, I.B. Nerve growth factor selectively increases cholinergic markers but not neuropeptides in rat basal forebrain in culture. Brain Res. 1987, 412, 295–301. [Google Scholar] [CrossRef]
- Chen, L.; Chen, Y.; Zhao, M.; Zheng, L.; Fan, D. Changes in the concentrations of trimethylamine N-oxide (TMAO) and its precursors in patients with amyotrophic lateral sclerosis. Sci. Rep. 2020, 10, 15198. [Google Scholar] [CrossRef]
- Cashman, N.R.; Durham, H.D.; Blusztajn, J.K.; Oda, K.; Tabira, T.; Shaw, I.T.; Dahrouge, S.; Antel, J.P. Neuroblastoma × spinal cord (NSC) hybrid cell lines resemble developing motor neurons. Dev. Dyn. 1992, 194, 209–221. [Google Scholar] [CrossRef]
- Gomes, C.; Palma, A.S.; Almeida, R.; Regalla, M.; McCluskey, L.F.; Trojanowski, J.Q.; Costa, J. Establishment of a cell model of ALS disease: Golgi apparatus disruption occurs independently from apoptosis. Biotechnol. Lett. 2008, 30, 603–610. [Google Scholar] [CrossRef]
- Jung, M.K.; Kim, K.Y.; Lee, N.Y.; Kang, Y.S.; Hwang, Y.J.; Kim, Y.; Sung, J.J.; McKee, A.; Kowall, N.; Lee, J.; et al. Expression of taurine transporter (TauT) is modulated by heat shock factor 1 (HSF1) in motor neurons of ALS. Mol. Neurobiol. 2013, 47, 699–710. [Google Scholar] [CrossRef] [PubMed]
- Latif, S.; Kang, Y. Change in cationic amino acid transport system and effect of lysine pretreatment on inflammatory state in amyotrophic lateral sclerosis cell model. Biomol. Ther. 2021, 29, 498–505. [Google Scholar] [CrossRef] [PubMed]
- Latif, S.; Kang, Y.-S. Differences of Transport Activity of Arginine and Regulation on Neuronal Nitric Oxide Synthase and Oxidative Stress in Amyotrophic Lateral Sclerosis Model Cell Lines. Cells 2021, 10, 3554. [Google Scholar] [CrossRef] [PubMed]
- Gyawali, A.; Gautam, S.; Hyeon, S.J.; Ryu, H.; Kang, Y.S. L-Citrulline level and transporter activity are altered in experimental models of amyotrophic lateral sclerosis. Mol. Neurobiol. 2021, 58, 647–657. [Google Scholar] [CrossRef]
- Gyawali, A.; Kang, Y.-S. Pretreatment Effect of Inflammatory Stimuli and Characteristics of Tryptophan Transport on Brain Capillary Endothelial (TR-BBB) and Motor Neuron Like (NSC-34) Cell Lines. Biomedicines 2020, 9, 9. [Google Scholar] [CrossRef]
- Kang, Y.S.; Terasaki, T.; Ohnishi, T.; Tsuji, A. In vivo and in vitro evidence for a common carrier mediated transport of choline and basic drugs through the blood-brain barrier. J. Pharmacobiodyn. 1990, 6, 353–360. [Google Scholar] [CrossRef] [Green Version]
- Shimon, M.; Egozi, Y.; Kloog, Y.; Sokolovsky, M.; Cohen, S. Kinetics of Choline Uptake into Isolated Rat Forebrain Microvessels: Evidence of Endocrine Modulation. J. Neurochem. 1988, 50, 1719–1724. [Google Scholar] [CrossRef]
- Kang, Y.S.; Terasaki, T.; Tsuji, A. Dysfunction of choline transport ststem through Blood-Brain Barrier in the stroke-prone spontaneously hypertensvie rat. J. Pharmacobio-Dyn. 1990, 13, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Yuan, Z.; Wagner, L.; Poloumienko, A.; Bakovic, M. Identification and expression of a mouse muscle-specific CTL1 gene. Gene 2004, 341, 305–312. [Google Scholar] [CrossRef]
- Wille, S.; Szekeres, A.; Majdic, O.; Prager, E.; Staffler, G.; Stöckl, J.; Kunthalert, D.; Prieschl, E.E.; Baumruker, T.; Burtscher, H.; et al. Characterization of CDw92 as a Member of the Choline Transporter-Like Protein Family Regulated Specifically on Dendritic Cells. J. Immunol. 2001, 167, 5795–5804. [Google Scholar] [CrossRef]
- Okuda, T.; Haga, T.; Kanai, Y.; Endou, H.; Ishihara, T.; Katsura, I. Identification and characterization of the high-affinity choline transporter. Nat. Neurosci. 2000, 3, 120–125. [Google Scholar] [CrossRef] [PubMed]
- Okada, T.; Muto, E.; Yamanaka, T.; Uchino, H.; Inazu, M. Functional Expression of Choline Transporters in Microglia and Their Regulation of Microglial M1/M2 Polarization. Int. J. Mol. Sci. 2022, 23, 8924. [Google Scholar] [CrossRef] [PubMed]
- Tuček, S. Problems in the organization and control of acetylcholine synthesis in brain neurons. Prog. Biophys. Mol. Biol. 1984, 44, 1–46. [Google Scholar] [CrossRef]
- Müller, J.; Born, I.; Neubert, R.H.; Brandsch, M. Apical uptake of choline and cationic drugs in epithelial cell lines derived from human placenta. Placenta 2005, 26, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Chen, S.; Lu, K.; Wang, F.; Deng, J.; Xu, Z.; Wang, X.; Zhou, Q.; Le, W.; Zhao, Y. Verapamil ameliorates motor neuron degeneration and improves lifespan in the SOD1G93A mouse model of als by enhancing autophagic flux. Aging Dis. 2019, 10, 1159–1173. [Google Scholar] [CrossRef] [Green Version]
- Thompson, E. Pharmacist considerations for treating patients with ALS. U.S. Pharm. 2019, 44, HS-2–HS-10. [Google Scholar]
- Tsai, C.P.; Lin, F.C.; Lee, J.K.W.; Lee, C.T.C. Aspirin use associated with amyotrophic lateral sclerosis: A total population-based case-control study. J. Epidemiol. 2015, 25, 172–177. [Google Scholar] [CrossRef] [Green Version]
- De Angelis, F.; Connick, P.; Parker, R.A.; Plantone, D.; Doshi, A.; John, N.; Stutters, J.; MacManus, D.; Prados, F.; Marshall, I.; et al. Amiloride, fluoxetine or riluzole to reduce brain volume loss in secondary progressive multiple sclerosis: The MS-SMART four-arm RCT. Effic. Mech. Eval. 2020, 7, 1–72. [Google Scholar] [CrossRef]
- Emon, M.A.E.K.; Kodamullil, A.T.; Karki, R.; Younesi, E.; Hofmann-Apitius, M. Using Drugs as Molecular Probes: A Computational Chemical Biology Approach in Neurodegenerative Diseases. J. Alzheimer’s Dis. 2017, 56, 677–686. [Google Scholar] [CrossRef] [Green Version]
- Szutowicz, A. Aluminum, NO, and nerve growth factor neurotoxicity in cholinergic Neurons. J. Neurosci. Res. 2001, 66, 1009–1018. [Google Scholar] [CrossRef]
- Szutowicz, A.; Madziar, B.; Pawełczyk, T.; Tomaszewicz, M.; Bielarczyk, H. Effects of NGF on acetylcholine, acetyl-CoA metabolism, and viability of differentiated and non-differentiated cholinergic neuroblastoma cells. J. Neurochem. 2004, 90, 952–961. [Google Scholar] [CrossRef] [PubMed]
- Rocco, M.L.; Soligo, M.; Manni, L.; Aloe, L. Nerve Growth Factor: Early Studies and Recent Clinical Trials. Curr. Neuropharmacol. 2018, 16, 1455–1465. [Google Scholar] [CrossRef]
- De Chaves, E.P.; Vance, D.E.; Campenot, R.B.; Vance, J.E. Axonal synthesis of phosphatidylcholine is required for normal axonal growth in rat sympathetic neurons. J. Cell Biol. 1995, 128, 913–918. [Google Scholar] [CrossRef] [PubMed]
- Araki, W.; Wurtman, R.J. Control of membrane phosphatidylcholine biosynthesis by diacylglycerol levels in neuronal cells undergoing neurite outgrowth. Proc. Natl. Acad. Sci. USA 1997, 94, 11946–11950. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oh, J.D.; Chartisathian, K.; Chase, T.N.; Butcher, L.L. Overexpression of neurotrophin receptor p75 contributes to the excitotoxin-induced cholinergic neuronal death in rat basal forebrain. Brain Res. 2000, 853, 174–185. [Google Scholar] [CrossRef]
- Turner, M.R.; Goldacre, R.; Ramagopalan, S.; Talbot, K.; Goldacre, M.J. Autoimmune disease preceding amyotrophic lateral sclerosis: An epidemiologic study. Neurology 2013, 81, 1222–1225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tortelli, R.; Zecca, C.; Piccininni, M.; Benmahamed, S.; Dell’Abate, M.T.; Barulli, M.R.; Capozzo, R.; Battista, P.; Logroscino, G. Plasma Inflammatory Cytokines Are Elevated in ALS. Front. Neurol. 2020, 11, 1–8. [Google Scholar] [CrossRef]
- Gyawali, A.; Hyeon, S.J.; Ryu, H.; Kang, Y.-S. The Alteration of L-Carnitine Transport and Pretreatment Effect under Glutamate Cytotoxicity on Motor Neuron-Like NSC-34 Lines. Pharmaceutics 2021, 13, 551. [Google Scholar] [CrossRef]
- Lee, N.Y.; Kang, Y.S. Taurine protects glutamate neurotoxicity in motor neuron cells. Adv. Exp. Med. Biol. 2017, 975, 887–895. [Google Scholar] [CrossRef]
- Rowley, T.J.; McKinstry, A.; Greenidge, E.; Smith, W.; Flood, P. Antinociceptive and anti-inflammatory effects of choline in a mouse model of postoperative pain. Br. J. Anaesth. 2010, 105, 201–207. [Google Scholar] [CrossRef] [Green Version]
- Gyawali, A.; Latif, S.; Choi, S.-H.; Hyeon, S.J.; Ryu, H.; Kang, Y.-S. Monocarboxylate transporter functions and neuroprotective effects of valproic acid in experimental models of amyotrophic lateral sclerosis. J. Biomed. Sci. 2022, 29, 2. [Google Scholar] [CrossRef] [PubMed]
- Latif, S.; Choi, S.-H.; Gyawali, A.; Hyeon, S.J.; Kang, Y.-S.; Ryu, H. Antioxidant and Neuroprotective Effects of Paeonol against Oxidative Stress and Altered Carrier-Mediated Transport System on NSC-34 Cell Lines. Antioxidants 2022, 11, 1392. [Google Scholar] [CrossRef] [PubMed]
- Wu, P.; Jiang, W.D.; Liu, Y.; Chen, G.F.; Jiang, J.; Li, S.H.; Feng, L.; Zhou, X.Q. Effect of choline on antioxidant defenses and gene expressions of Nrf2 signaling molecule in the spleen and head kidney of juvenile Jian carp (Cyprinus carpio var. Jian). Fish Shellfish Immunol. 2014, 38, 374–382. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | WT | MT |
|---|---|---|
| Km1 (µM) | 10.1 ± 2.5 | 4.4 ± 0.64 * |
| Km2 (µM) | 280 ± 21 | 43.7 ± 1.2 *** |
| Vmax1 (pmol/mg protein/min) | 101 ± 20 | 55 ± 11 * |
| Vmax2 (pmol/mg protein/min) | 107 ± 81 | 153 ± 16 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Latif, S.; Kang, Y.-S. Protective Effects of Choline against Inflammatory Cytokines and Characterization of Transport in Motor Neuron-like Cell Lines (NSC-34). Pharmaceutics 2022, 14, 2374. https://doi.org/10.3390/pharmaceutics14112374
Latif S, Kang Y-S. Protective Effects of Choline against Inflammatory Cytokines and Characterization of Transport in Motor Neuron-like Cell Lines (NSC-34). Pharmaceutics. 2022; 14(11):2374. https://doi.org/10.3390/pharmaceutics14112374
Chicago/Turabian StyleLatif, Sana, and Young-Sook Kang. 2022. "Protective Effects of Choline against Inflammatory Cytokines and Characterization of Transport in Motor Neuron-like Cell Lines (NSC-34)" Pharmaceutics 14, no. 11: 2374. https://doi.org/10.3390/pharmaceutics14112374