


Modulation of the mTOR Pathway by Curcumin in the Heart of Septic Mice
 , , and
, , and 
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Experimental Animals
2.2. Polymicrobial Sepsis (Cecal Ligation and Puncture—CLP Model)
2.3. Euthanasia and Collection
2.4. High-Resolution Microscopy
2.5. Transmission Electronic Microscopy
2.6. Western Blotting
2.7. RNA Isolation, Reverse Transcription, and RT-PCR
2.8. Statistical Analysis
3. Results
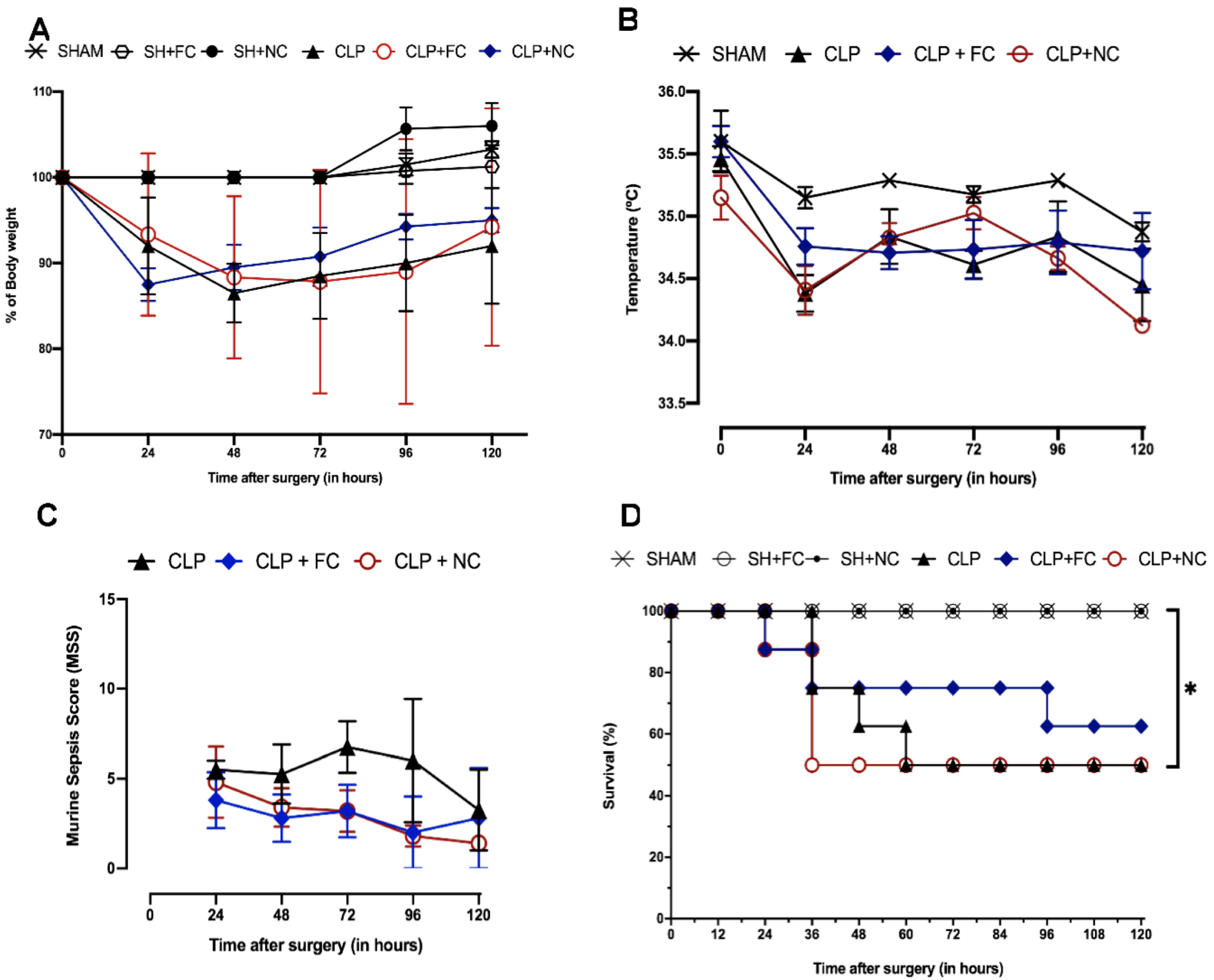
3.1. Characterization of Murine Sepsis Using the CLP Model
3.2. Effects of Curcumin on Cardiac Morphology
3.2.1. Histopathological Evaluation of the Heart
3.2.2. Ultrastructural Analysis of the Myocardium
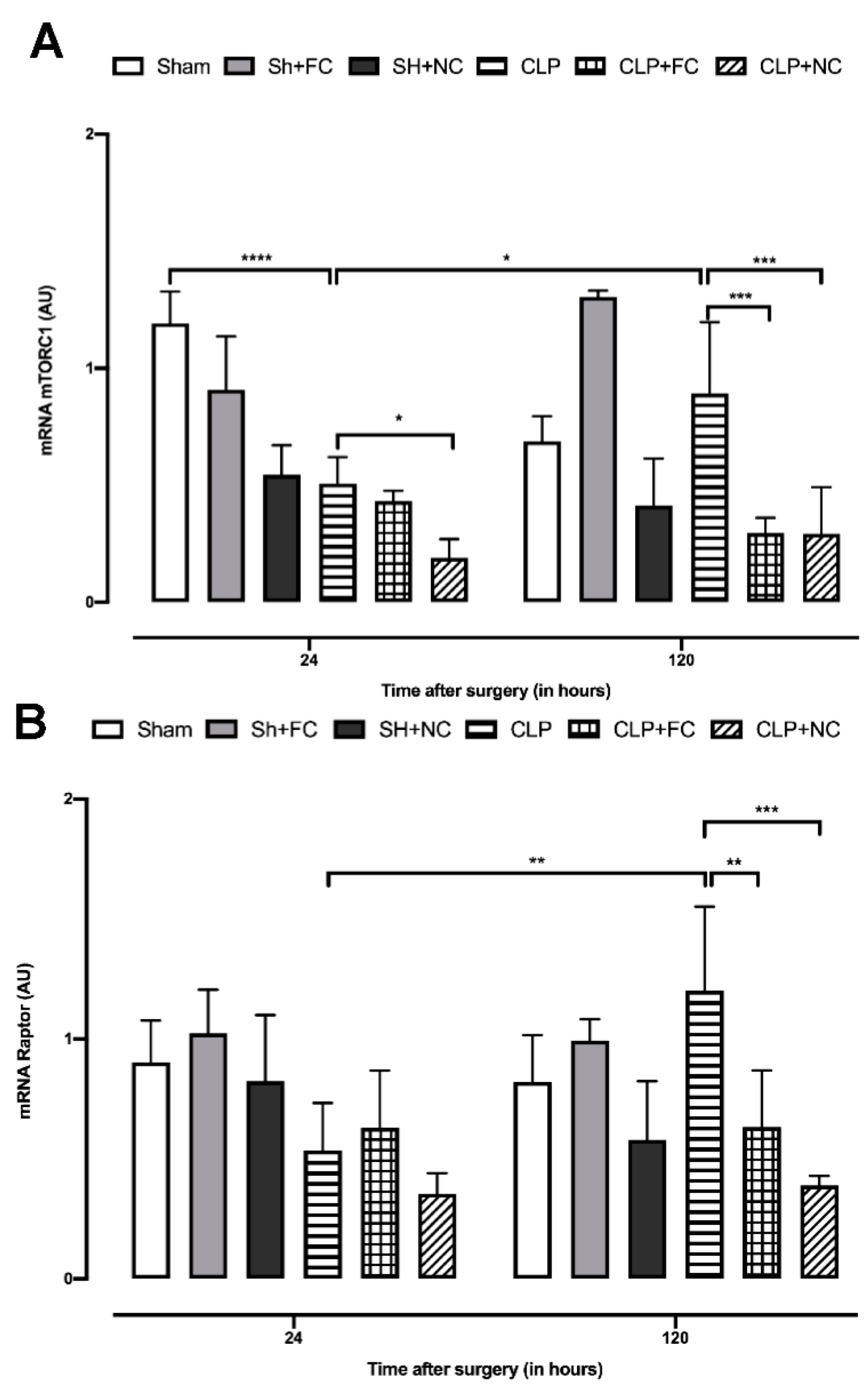
3.3. Gene Expression of mTORC1 Complex Components
3.4. Gene Expression of mTORC2 Complex Components
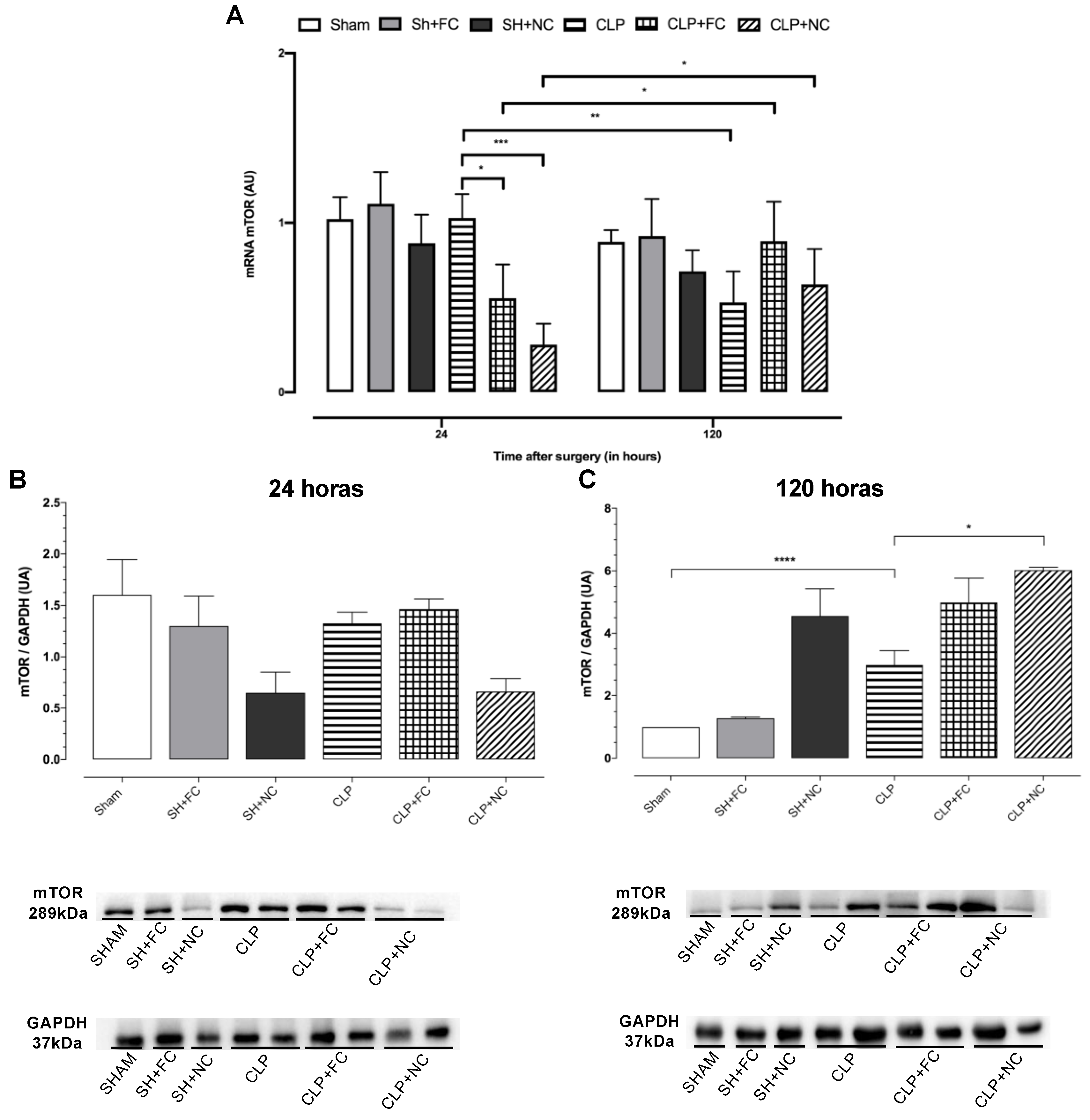
3.5. Gene and Protein Expression of Total mTOR
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
Obtaining and Characterizing Nanocurcumin (NC)
References
- Burton, A.; Waisbren, M.D. Bacteremia due to gram-negative bacilli other than the salmonella. AMA Arch. Intern. Med. 1951, 88, 467–488. [Google Scholar] [CrossRef]
- Parrillo, J.E.; Parker, M.M.; Natanson, C.; Suffredini, A.F.; Danner, R.L.; Cunnion, R.E.; Ognibene, F.P. Septic Shock in Humans Advances in the Understanding of Pathogenesis, Cardiovascular. Ann. Intern. Med. 1990, 113, 227–242. [Google Scholar] [CrossRef] [PubMed]
- Merx, M.W.; Weber, C. Sepsis and the Heart. Circulation 2007, 116, 793–802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Souza Dantas, V.C.; Costa, E.L.V. A Look at the Diastolic Function in Severe Sepsis and Septic Shock. Rev. Bras. Ter. Intensiv. 2015, 27, 307–308. [Google Scholar]
- Celes, M.R.N.; Malvestio, L.M.; Suadicani, S.O.; Prado, C.M.; Figueiredo, M.J.; Campos, E.C.; Freitas, A.C.S.; Spray, D.C.; Tanowitz, H.B.; da Silva, J.S.; et al. Disruption of Calcium Homeostasis in Cardiomyocytes Underlies Cardiac Structural and Functional Changes in Severe Sepsis. PLoS ONE 2013, 8, e68809. [Google Scholar] [CrossRef]
- Celes, M.R.N.; Prado, C.M.; Rossi, M.A. Sepsis: Going to the Heart of the Matter. Pathobiology 2012, 80, 70–86. [Google Scholar] [CrossRef]
- Celes, M.R.N.; Torres-Dueñas, D.; Malvestio, L.M.; Blefari, V.; Campos, E.C.; Ramos, S.G.; Prado, C.M.; Cunha, F.Q.; Rossi, M.A. Disruption of Sarcolemmal Dystrophin and β-Dystroglycan May Be a Potential Mechanism for Myocardial Dysfunction in Severe Sepsis. Lab. Investig. 2010, 90, 531. [Google Scholar] [CrossRef] [Green Version]
- Rattis, B.A.C.; Freitas, A.C.; Oliveira, J.F.; Calandrini-Lima, J.L.A.; Figueiredo, M.J.; Soave, D.F.; Ramos, S.G.; Celes, M.R.N. Effect of Verapamil, an L-Type Calcium Channel Inhibitor, on Caveolin-3 Expression in Septic Mouse Hearts. Oxid. Med. Cell. Longev. 2021, 2021, 6667074. [Google Scholar] [CrossRef]
- Celes, M.R.N.; Torres-Dueñas, D.; Alves-Filho, J.C.; Duarte, D.B.; Cunha, F.Q.; Rossi, M.A. Reduction of Gap and Adherens Junction Proteins and Intercalated Disc Structural Remodeling in the Hearts of Mice Submitted to Severe Cecal Ligation and Puncture Sepsis. Crit. Care Med. 2007, 35, 2176–2185. [Google Scholar] [CrossRef]
- Rossi, M.A.; Celes, M.R.N.; Prado, C.M.; Saggioro, F.P. Myocardial Structural Changes in Long-Term Human Severe Sepsis/Septic Shock May Be Responsible for Cardiac Dysfunction. Shock 2007, 27, 10–18. [Google Scholar] [CrossRef]
- Freitas, A.C.S.; Figueiredo, M.J.; Campos, E.C.; Soave, D.F.; Ramos, S.G.; Tanowitz, H.B.; Celes, M.R.N. Activation of Both the Calpain and Ubiquitin-Proteasome Systems Contributes to Septic Cardiomyopathy through Dystrophin Loss/Disruption and mTOR Inhibition. PLoS ONE 2016, 11, e0166839. [Google Scholar] [CrossRef] [PubMed]
- Laplante, M.; Sabatini, D.M. MTOR Signaling in Growth Control and Disease. Cell 2012, 149, 274–293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walley, K.R. Sepsis-Induced Myocardial Dysfunction and Mammalian Target of Rapamycin Signalling Pathways. Can. J. Cardiol. 2019, 35, 809–812. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fantus, D.; Rogers, N.M.; Grahammer, F.; Huber, T.B.; Thomson, A.W. Roles of mTOR Complexes in the Kidney: Implications for Renal Disease and Transplantation. Nat. Rev. Nephrol. 2016, 12, 587–609. [Google Scholar] [CrossRef] [PubMed]
- Saxton, R.A.; Sabatini, D.M. MTOR Signaling in Growth, Metabolism, and Disease. Cell 2017, 168, 960–976. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.Y.; Sabatini, D.M. mTOR at the Nexus of Nutrition, Growth, Ageing, and Disease. Nat. Rev. Mol. Cell Biol. 2020, 21, 183–203. [Google Scholar] [CrossRef]
- Cheng MM, W.; Long, Y.; Wang, H.; Han MM, W.; Zhang, J.; Cui, N. Role of the mTOR Signalling Pathway in Human Sepsis-Induced Myocardial Dysfunction. Can. J. Cardiol. 2019, 35, 875–883. [Google Scholar] [CrossRef] [Green Version]
- Frost, R.A.; Lang, C.H. MTOR Signaling in Skeletal Muscle during Sepsis and Inflammation: Where Does It All Go Wrong? Physiology 2011, 26, 83–96. [Google Scholar] [CrossRef] [Green Version]
- Pålsson-McDermott, E.M.; O’Neill, L.A.J. Targeting Immunometabolism as an Anti-Inflammatory Strategy. Cell Res. 2020, 30, 300–314. [Google Scholar] [CrossRef] [Green Version]
- Soltani, A.; Bahreyni, A.; Boroumand, N.; Roshan, M.K.; Khazaei, M.; Ryzhikov, M.; Soleimanpour, S.; Avan, A.; Hassanian, S.M. Therapeutic Potency of MTOR Signaling Pharmacological Inhibitors in the Treatment of Proinflammatory Diseases, Current Status, and Perspectives. J. Cell. Physiol. 2018, 233, 4783–4790. [Google Scholar] [CrossRef]
- Sciarretta, S.; Volpe, M.; Sadoshima, J. Mammalian Target of Rapamycin Signaling in Cardiac Physiology and Disease. Circ. Res. 2014, 114, 549–564. [Google Scholar] [CrossRef] [PubMed]
- Zhao, D.; Yang, J.; Yang, L. Insights for Oxidative Stress and mTOR Signaling in Myocardial Ischemia/Reperfusion Injury under Diabetes. Oxid. Med. Cell. Longev. 2017, 2017, 6437467. [Google Scholar] [CrossRef] [Green Version]
- Han, W.; Wang, H.; Su, L.; Long, Y.; Cui, N.; Liu, D. Inhibition of the mTOR Pathway Exerts Cardioprotective Effects Partly through Autophagy in CLP Rats. Mediators Inflamm. 2018, 2018, 4798209. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.C.; Patchva, S.; Koh, W.; Aggarwal, B.B. Discovery of Curcumin, a Component of the Golden Spic, and Its Micraculous Biological Activities. Clin. Exp. Pharmacol. Physiol. 2012, 39, 283–299. [Google Scholar] [CrossRef] [PubMed]
- Paciello, F.; Rita Fetoni, A.; Mezzogori, D.; Rolesi, R.; Di Pino, A.; Paludetti, G.; Grassi, C.; Troiani, D. The Dual Role of Curcumin and Ferulic Acid in Counteracting Chemoresistance and Cisplatin-Induced Ototoxicity. Sci. Rep. 2020, 10, 1063. [Google Scholar] [CrossRef] [Green Version]
- Rattis, B.A.C.; Ramos, S.G.; Celes, M.R.N. Curcumin as a Potential Treatment for COVID-19. Front. Pharmacol. 2021, 12, 2085–2087. [Google Scholar] [CrossRef]
- Johnson, S.M.; Gulhati, P.; Arrieta, I.; Wang, X.; Uchida, T.; Gao, T.; Evers, B.M. Curcumin Inhibits Proliferation of Colorectal Carcinoma by Modulating Akt/MTOR Signaling. Anticancer Res. 2009, 29, 3185–3190. [Google Scholar]
- Beevers, C.S.; Chen, L.; Liu, L.; Luo, Y.; Webster, N.J.G.; Huang, S. Curcumin Disrupts the Mammalian Target of Rapamycin-Raptor Complex. Cancer Res. 2009, 69, 1000–1008. [Google Scholar] [CrossRef] [Green Version]
- Wichterman, K.A.; Baue, A.E.; Chaudry, I.H. Sepsis and Septic Shock—A Review of Laboratory Models and a Proposal. J. Surg. Res. 1980, 29, 189–201. [Google Scholar] [CrossRef]
- Rittirsch, D.; Huber-Lang, M.S.; Flierl, M.A.; Ward, P.A. Immunodesign of Experimental Sepsis by Cecal Ligation and Puncture. Nat. Protoc. 2009, 4, 31–36. [Google Scholar] [CrossRef] [Green Version]
- Mai, S.H.C.; Sharma, N.; Kwong, A.C.; Dwivedi, D.J.; Khan, M.; Grin, P.M.; Fox-Robichaud, A.E.; Liaw, P.C. Body Temperature and Mouse Scoring Systems as Surrogate Markers of Death in Cecal Ligation and Puncture Sepsis. Intensive Care Med. Exp. 2018, 6, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Silva, L.S.; Catalão, C.H.R.; Felippotti, T.T.; De Oliveira-Pelegrin, G.R.; Petenusci, S.; De Freitas, L.A.P.; Rocha, M.J.A. Curcumin Suppresses Inflammatory Cytokines and Heat Shock Protein 70 Release and Improves Metabolic Parameters during Experimental Sepsis. Pharm. Biol. 2017, 55, 269–276. [Google Scholar] [CrossRef]
- Xiao, X.; Yang, M.; Sun, D.; Sun, S. Curcumin Protects against Sepsis-Induced Acute Lung Injury in Rats. J. Surg. Res. 2012, 176, e31–e39. [Google Scholar] [CrossRef]
- Chen, L.; Lu, Y.; Zhao, L.; Hu, L.; Qiu, Q.; Zhang, Z.; Li, M.; Hong, G.; Wu, B.; Zhao, G.; et al. Curcumin Attenuates Sepsis-Induced Acute Organ Dysfunction by Preventing Inflammation and Enhancing the Suppressive Function of Tregs. Int. Immunopharmacol. 2018, 61, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Siddiqui, A.M.; Cui, X.; Wu, R.; Dong, W.; Zhou, M.; Hu, M.; Simms, H.H.; Wang, P. The Anti-Inflammatory Effect of Curcumin in an Experimental Model of Sepsis Is Mediated by up-Regulation of Peroxisome Proliferator-Activated Receptor-γ. Crit. Care Med. 2006, 34, 1874–1882. [Google Scholar] [CrossRef] [PubMed]
- Kakihana, Y.; Ito, T.; Nakahara, M.; Yamaguchi, K.; Yasuda, T. Sepsis-Induced Myocardial Dysfunction: Pathophysiology and Management. J. Intensive Care 2016, 4, 22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arshad, L.; Areeful Haque, M.; Bukhari, S.N.A.; Jantan, I. An Overview of Structure-Activity Relationship Studies of Curcumin Analogs as Antioxidant and Anti-Inflammatory Agents. Future Med. Chem. 2017, 9, 605–626. [Google Scholar] [CrossRef]
- Lu, W.; Jiang, J.P.; Hu, J.; Wang, J.; Zheng, M.Z. Curcumin Protects against Lipopolysaccharide-Induced Vasoconstriction Dysfunction via Inhibition of Thrombospondin-1 and Transforming Growth Factor-SS1. Exp. Ther. Med. 2015, 9, 377–383. [Google Scholar] [CrossRef] [Green Version]
- Karunaweera, N.; Raju, R.; Gyengesi, E.; Munch, G. Plant Polyphenols as Inhibitors of Nf-Kb Induced Cytokine Production—A Potential Anti-Inflammatory Treatment for Alzheimer’s Disease? Front. Mol. Neurosci. 2015, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Yin, H.; Guo, Q.; Li, X.; Tang, T.; Li, C.; Wang, H.; Sun, Y.; Feng, Q.; Ma, C.; Gao, C.; et al. Curcumin Suppresses IL-1β Secretion and Prevents Inflammation through Inhibition of the NLRP3 Inflammasome. J. Immunol. 2018, 200, 2835–2846. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Tang, Q.; Duan, P.; Yang, L. Curcumin as a Therapeutic Agent for Blocking NF-ΚB Activation in Ulcerative Colitis. Immunopharmacol. Immunotoxicol. 2018, 40, 476–482. [Google Scholar] [CrossRef] [PubMed]
- Solt, L.A.; May, M.J. The IκB Kinase Complex: Master Regulator of NF-ΚB Signaling. Immunol. Res. 2008, 42, 3–18. [Google Scholar] [CrossRef]
- Mantzarlis, K.; Tsolaki, V.; Zakynthinos, E. Role of Oxidative Stress and Mitochondrial Dysfunction in Sepsis and Potential Therapies. Oxid. Med. Cell. Longev. 2017, 2017, 5985209. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nimse, S.B.; Pal, D. Free Radicals, Natural Antioxidants, and Their Reaction Mechanisms. RSC Adv. 2015, 5, 27986–28006. [Google Scholar] [CrossRef] [Green Version]
- Yang, C.; Wu, K.; Li, S.; You, Q. Protective Effect of Curcumin against Cardiac Dysfunction in Sepsis Rats. Pharm. Biol. 2013, 51, 482–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Priyadarsini, K.I.; Maity, D.K.; Naik, G.H.; Kumar, M.S.; Unnikrishnan, M.K.; Satav, J.G.; Mohan, H. Role of Phenolic O-H and Methylene Hydrogen on the Free Radical Reactions and Antioxidant Activity of Curcumin. Free Radic. Biol. Med. 2003, 35, 475–484. [Google Scholar] [CrossRef]
- Ahmadabady, S.; Beheshti, F.; Shahidpour, F.; Khordad, E.; Hosseini, M. A Protective Effect of Curcumin on Cardiovascular Oxidative Stress Indicators in Systemic Inflammation Induced by Lipopolysaccharide in Rats. Biochem. Biophys. Rep. 2021, 25, 100908. [Google Scholar] [CrossRef]
- Lin, H.; Wang, W.; Lee, M.; Meng, Q.; Ren, H. Current Status of Septic Cardiomyopathy: Basic Science and Clinical Progress. Front. Pharmacol. 2020, 11, 210. [Google Scholar] [CrossRef] [Green Version]
- Chong, Z.Z.; Shang, Y.C.; Maiese, K. Cardiovascular Disease and mTOR Signaling. Trends Cardiovasc. Med. 2011, 21, 151–155. [Google Scholar] [CrossRef] [Green Version]
- Das, A.; Reis, F.; Mishra, P.K. MTOR Signaling in Cardiometabolic Disease, Cancer, and Aging 2018. Oxid. Med. Cell. Longev. 2019, 2019, 9692528. [Google Scholar] [CrossRef]
- Luck, C.; Demarco, V.G.; Mahmood, A.; Gavini, M.P.; Pulakat, L. Differential Regulation of Cardiac Function and Intracardiac Cytokines by Rapamycin in Healthy and Diabetic Rats. Oxid. Med. Cell. Longev. 2017, 2017, 5724046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, S.C.; Patchva, S.; Aggarwal, B.B. Therapeutic Roles of Curcumin: Lessons Learned from Clinical Trials. AAPS J. 2013, 15, 195–218. [Google Scholar] [CrossRef] [PubMed]
- Qiu, Z.; Zhang, S.; Li, A.; Yu, J.; Li, N.; Huang, F.; Liu, B. The Role of Curcumin in Disruption of HIF-1α Accumulation to Alleviate Adipose Fibrosis via AMPK-Mediated MTOR Pathway in High-Fat Diet Fed Mice. J. Funct. Foods 2017, 33, 155–165. [Google Scholar] [CrossRef]
- Mendoza, M.C.; Er, E.E.; Blenis, J. The Ras-ERK and PI3K-MTOR Pathways: Cross-Talk and Compensation. Trends Biochem. Sci. 2011, 36, 320–328. [Google Scholar] [CrossRef] [Green Version]
- Xiao, K.; Jiang, J.; Guan, C.; Dong, C.; Wang, G.; Bai, L.; Sun, J.; Hu, C.; Bai, C. Curcumin Induces Autophagy via Activating the AMPK Signaling Pathway in Lung Adenocarcinoma Cells. J. Pharmacol. Sci. 2013, 123, 102–109. [Google Scholar] [CrossRef] [Green Version]
- Zhao, X.; Lu, S.; Nie, J.; Hu, X.; Luo, W.; Wu, X.; Liu, H.; Feng, Q.; Chang, Z.; Liu, Y.; et al. Phosphoinositide-Dependent Kinase 1 and MTORC2 Synergistically Maintain Postnatal Heart Growth and Heart Function in Mice. Mol. Cell. Biol. 2014, 34, 1966–1975. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.D.; Shao, Y.; Liu, D.; Liu, N.Y.; Zhu, D.Y. Rictor/MTORC2 Involves Mitochondrial Function in ES Cells Derived Cardiomyocytes via Mitochondrial Connexin 43. Acta Pharmacol. Sin. 2021, 42, 1790–1797. [Google Scholar] [CrossRef]
- Völkers, M.; Konstandin, M.H.; Doroudgar, S.; Toko, H.; Quijada, P.; Din, S.; Joyo, A.; Ornelas, L.; Samse, K.; Thuerauf, D.J.; et al. Mechanistic Target of Rapamycin Complex 2 Protects the Heart from Ischemic Damage. Circulation 2013, 128, 2132–2144. [Google Scholar] [CrossRef] [Green Version]
- Yano, T.; Ferlito, M.; Aponte, A.; Kuno, A.; Miura, T.; Murphy, E.; Steenbergen, C. Pivotal Role of MTORC2 and Involvement of Ribosomal Protein S6 in Cardioprotective Signaling. Circ. Res. 2014, 114, 1268–1280. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Groups | |
|---|---|
| SHAM | group of sham-operated animals; |
| SH + FC | group of sham-operated animals treated with free Curcumin; |
| SH + NC | group of sham-operated animals treated with nanocurcumin; |
| CLP | group of animals submitted to a severe septic stimulus; |
| CLP + FC | group of animals submitted to severe septic stimulus treated with free Curcumin; |
| CLP + NC | group of animals submitted to severe septic stimulus treated with nanocurcumin; |
| Gene | Forward | Reverse |
|---|---|---|
| GAPDH | CTTTGTCAAGCTCATTTCCTGG | TCTTGCTCAGTGTCCTTGC |
| mTORC1 | TCGATGAATGTGGGATTGTGG | TGCCTTCGCTGGAGAATATC |
| mTORC2 | ATCTCCGTGTTTATGCTGTCC | CACCGTTTCTCCATTGAGAAC |
| Raptor | GCAGAGCTGGAGAATGAAGG | GTCGAGGCTCTGCTTGTACC |
| Rictor | ATGGAAATAAGGCGAGGTCTG | AAAGCCTCCAACTGTCCTG |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rattis, B.A.C.; Piva, H.L.; Duarte, A.; Gomes, F.G.F.L.R.; Lellis, J.R.; Soave, D.F.; Ramos, S.G.; Tedesco, A.C.; Celes, M.R.N. Modulation of the mTOR Pathway by Curcumin in the Heart of Septic Mice. Pharmaceutics 2022, 14, 2277. https://doi.org/10.3390/pharmaceutics14112277
Rattis BAC, Piva HL, Duarte A, Gomes FGFLR, Lellis JR, Soave DF, Ramos SG, Tedesco AC, Celes MRN. Modulation of the mTOR Pathway by Curcumin in the Heart of Septic Mice. Pharmaceutics. 2022; 14(11):2277. https://doi.org/10.3390/pharmaceutics14112277
Chicago/Turabian StyleRattis, Bruna A. C., Henrique L. Piva, Andressa Duarte, Frederico G. F. L. R. Gomes, Janaína R. Lellis, Danilo F. Soave, Simone G. Ramos, Antonio C. Tedesco, and Mara R. N. Celes. 2022. "Modulation of the mTOR Pathway by Curcumin in the Heart of Septic Mice" Pharmaceutics 14, no. 11: 2277. https://doi.org/10.3390/pharmaceutics14112277