1. Introduction
Strokes are one of the leading causes of death worldwide, and the lack of effective therapies for acute cerebrovascular injuries is one of the most pressing medical problems [
1]. To date, there are no therapeutic protocols that can be used to reduce the effects of a post-ischaemic brain injury and stimulate revascularisation and neuro-regeneration after stroke in practical medicine. The difficulties in treating stroke are due to drastic necrosis during the first few minutes in the ischaemic core, followed by apoptosis in the surrounding penumbra for a 3–6 h period (therapeutic window) and degeneration in the ischemic area for several weeks [
2]. The current urgent treatment aims to remove the clot and to restore the cerebral blood flow. However, for the preservation of the brain tissue, the stroke pathophysiology suggests that the therapeutic impacts not only on ischaemia but also on excitotoxicity be addressed, along with apoptosis and neuroinflammation to prevent brain cell death [
3]. To date, cell [
4,
5] and gene [
6,
7] therapies are proposed as promising therapeutic strategies for stroke treatment.
It is well known that, during neuro-ontogenesis, the survivability and migration of neural progenitor cells, directed axon growth and synaptogenesis are realised through neurotrophic and growth factors produced in certain neurons or vascular and glial cells. Thus, the general principles of neuro-ontogenesis may also apply to neuro-regeneration. Moreover, the redeployment of the embryonic gene regulatory network during regeneration [
8] also indicates that one could use the genes involved in neurogenesis to stimulate neuro-regeneration in adults with CNS disorders. The delivery of the recombinant cDNA encoding neurotrophic factors, such as nerve growth factor (NGF), vascular endothelial growth factor (VEGF), brain derived growth factor (BDNF), glial-cell-line-derived neurotrophic factor (GDNF), basic fibroblast growth factor (BFGF) and ciliary neurotrophic factor (CNTF), demonstrated positive therapeutic effects on the ischaemic brain [
7,
9].
Recently, to examine stroke gene therapy in the acute phase [
10] and preventive [
11] treatment in a rat model, we employed human umbilical cord blood mononuclear cells (UCB-MC) for the tandem delivery of three therapeutic genes encoding VEGF, GDNF, and the neural cell adhesion molecule (NCAM). We showed the positive effects of the gene-modified UCB-MC on the negative consequences of the ischaemia-induced brain injury. However, in practical medicine, the use of the UCB-MC is restricted not only due to ethics and possible immunogenic problems, but also because it is seriously limited by the small amounts of UCB-MC that can be acquired from one donor and by the deficiency of these types of cells in general. In view of this, we developed a new personalised precision cell-mediated (ex vivo) gene therapy approach based on the intravenous administration of a patient’s leucoconcentrate, simultaneously transduced with three chimeric adenoviral vectors (Ad5/35F) carrying cDNA encoding VEGF, GDNF, and NCAM in equal ratios for the treatment of spinal cord injuries in a mini-pig model [
12,
13].
Many of the studies testing the new approaches to stroke treatment have been performed on rodent models, which have limited translational potential. The pathophysiology and pathomorphology of stroke in rodents with lissencephalic brains are not fully comparable with brain injuries in human stroke [
14]. Therefore, preclinical stroke studies need to be performed on large mammals that have neuroanatomical, physiological and biochemical characteristics that are close to those of a human. Pigs not only have a gyrencephalic brain and white matter volume similar to that of humans [
14], but they are also genetically closer to humans than mice [
15], which suggests the practicality of using a porcine model of ischaemic stroke for the discovery of new approaches to stroke treatment. Thus, swine stroke models are considered to have translational advantages compared to rodents, dogs, and non-human primates.
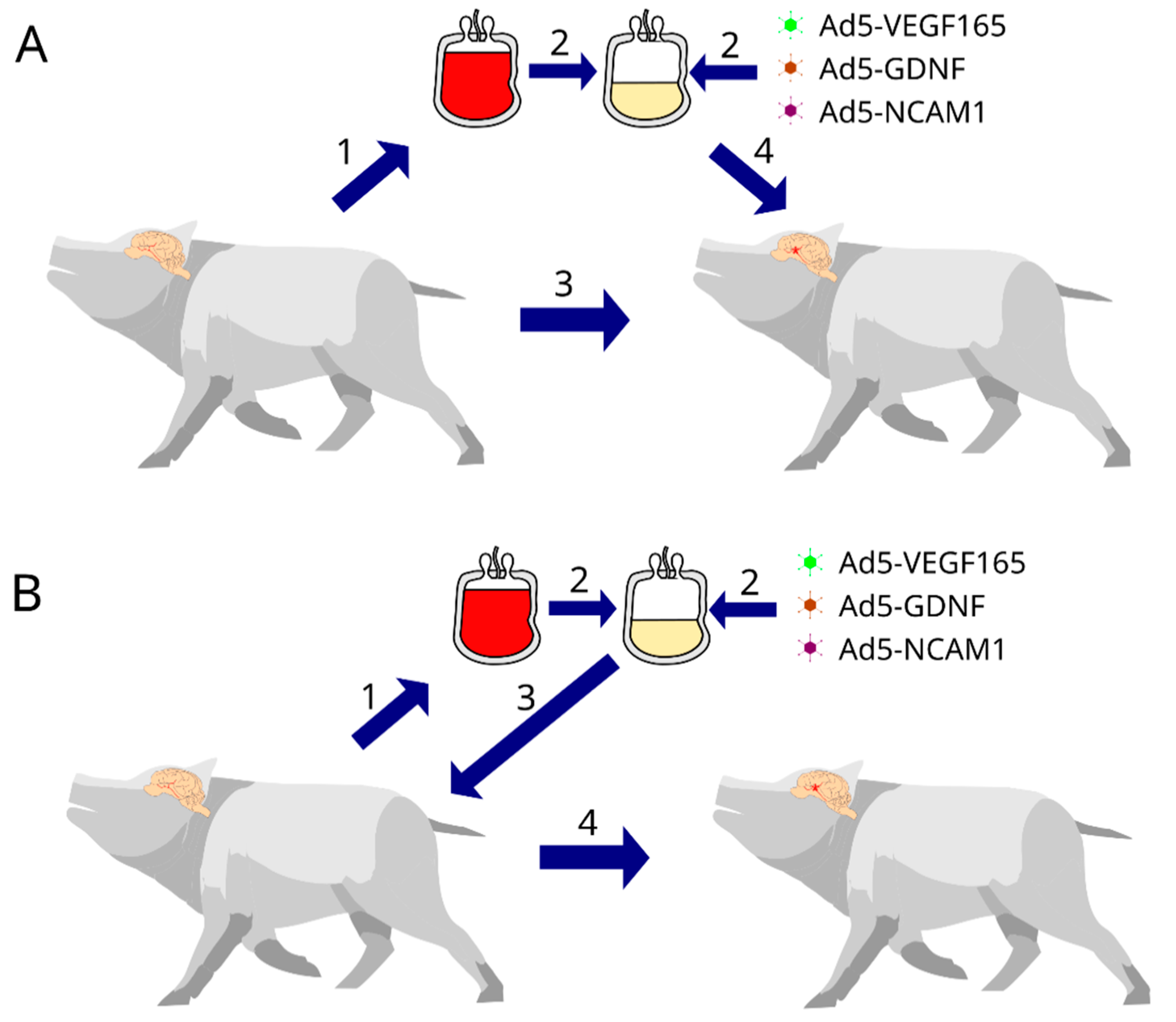
In the present mini-pig study, we developed a protocol for simulating moderate ischemic cerebral stroke and evaluated the efficacy of the autologous, genetically enriched leucoconcentrate (GEL) producing recombinant VEGF, GDNF, and NCAM in the acute phase (intravenous infusion 4 h after stroke modelling) and preventive phase (intravenous infusion of GEL 2 days before stroke modelling) of stroke gene therapy (
Figure 1).
2. Material and Methods
2.1. Animals
Miniature Vietnamese pot-bellied 8-month-old female pigs (25–30 kg; n = 16) were obtained from the Kazan State Academy of Veterinary Medicine by N.E. Bauman (Kazan, Russia). Throughout the investigation, the mini-pigs were kept with one per housing area under controlled temperatures (24–25 °C), air conditioning, a 12 h light/dark cycle, and properly organised feeding. All experimental procedures were approved by the Kazan State Medical University Animal Care and Use Committee (approval No. 5, dated 26 May 2020).
2.2. Autologous Genetically Enriched Leucoconcentrate
Genetically enriched leucoconcentrate (GEL) was prepared according to our original protocol, as described previously [
13]. With the mini-pigs under anaesthesia following an intramuscular injection of tiletamine/zolazepam (Zoletil 100, Virbac Laboratoires, Carros, France; 10 mg/kg) and xylazine (Xyla, Interchemie werken ‘De Adelaar’ B.V., Castenray, the Netherlands; 40 mg/kg), 50 mL of peripheral blood was collected from
V. subclavia. The blood was collected in a plastic blood bag of 250 mL with anticoagulant CPDA-1 (Green Cross, Yongin-si, Korea) (the volume of preservative in each bag was 35 mL). The leucoconcentrate was prepared in the blood bag according to our original protocol, which sequentially includes the sedimentation of erythrocytes with 6% hydroxyethyl starch, followed by centrifugation at 34×
g for 10 min at 10 °C (DP-2065 R PLUS, Centrifugal Presvac RV; Presvac, Buenos Aires, Argentina) and washing with saline. The supernatant was discarded from the bag, and the remaining solution containing white blood cells (WBC) in the bag was considered to be the leucoconcentrate.
Chimeric (Ad5/35F) viral vectors carrying cDNA encoding VEGF, GDNF, NCAM and reporter green fluorescent protein (GFP) were developed based on the human adenovirus serotype 5 with a fibre derived from adenovirus serotype 35 (Ad5/35) [
13]. The transduction of the obtained leucoconcentrate was conducted using a blood container with an MOI (multiplicity of infection) equal to 10 based on the WBC count and the titres of the adenoviral vectors. An equal ratio of each chimeric adenoviral vector, Ad5/35-VEGF165 (2.0 × 10
9 PFU/mL), Ad5/35-GDNF (7.0 × 10
10 PFU/mL), and Ad5/35-NCAM1 (5.0 × 10
10 PFU/mL), was injected into the blood bag, and the leucoconcentrate was transduced for 12 h under constant rocking on a shaker (ELMI) at room temperature (21–22 °C). After that, 200 mL of sterile saline was added to the blood bag, and it centrifuged at 1000 rpm at 10 °C for 10 min. The supernatant (saline containing cell fragments and free viral particles) was extracted from the container, so that approximately 30 mL of solution remained in the container, including the cell suspension, a leucoconcentrate enriched with genetic material (GEL-VGN). The reporter GEL-GFP was prepared using Ad5/35-GFP (2.5 × 10
9 PFU/mL). The efficacy of the therapeutic (VEGF165, GDNF, NCAM1) and reporter (GFP) transgenes’ expression was confirmed as described previously [
13]. In the present study, the GFP expression in the WBCs in the mini-pig brain was confirmed using fluorescent microscopy 7 days after the intravenous auto-infusion of GML-GFP into the mini-pigs with stroke.
2.3. Ischaemic Stroke Modelling
In clinical practice, middle cerebral artery (MCA) occlusion is the primary cause of cerebral damage in humans. Therefore, the MCA is the main target of in vivo stroke models [
16]. The peculiarities of the cerebral vascular anatomy of pigs, such as the presence of two to four MCAs on each side of the brain and the involvement of the anterior and posterior cerebral arteries in the circulation in the circle of Willis, present certain difficulties in reaching a reproducible ischemic cerebral infarction [
14]. In the study, we employed the protocol which includes the sequential occlusion of the right common carotid artery and the left MCA. The animals were intramuscularly anaesthetised with 3 mg/kg Zoletil
®100 (Virbac Sante Animale, Carros, France) and subsequently connected to an inhalation anaesthesia apparatus (Minor Vet Optima, Zoomed, Moscow, Russia) through which 2.0–2.5% isoflurane (Laboratorios Karizoo, S.A., Barcelona, Spain) mixed with oxygen was administered. All surgical manipulations were performed in accordance with the aseptic and antiseptic rules. In the first step, the right common carotid artery was ligated to reduce the blood flow in the circle of Willis. The next step included the occlusion of the left MCA. Operative access to the MCA was achieved via a trepanation hole in the left temporal bone. After trepanation, the dura mater was dissected, and the distal branches of the MCA were cauterised by electrocoagulation under an operating microscope, YZ-20T9 (Nanjing Redsun Optical Co., LTD, Nanjing, China). In the postoperative period, all animals received antibacterial (Ceftriaxone intramuscularly, 50 mg/kg once daily) and analgesic therapy (Ketorol intramuscularly, 2.5 mg/kg three times daily).
2.4. Experimental Groups
For the stroke gene therapy in the acute phase, GEL-VGN was prepared in the 14–16 h period before stroke modelling and was infused via the auricular vein 4 h after surgery. For the preventive gene therapy, GEL-VGN was prepared and infused 2 days before stroke modelling. In the study of the genetically modified WBCs expressing the reporter GFP homing in the brains of the experimental mini-pigs, two groups of animals were infused with GEL-GFP in the acute phase of stroke and before stroke modelling.
According to the design of the experiment, the animals were divided into 6 groups: (1) Con—the control group (
n = 3, after stroke, animals were infused with saline), (2) TA—therapeutic acute group (
n = 3, after stroke, animals were infused with GEL-VGN), (3) TP—therapeutic preventive group (
n = 3, before stroke, animals were infused with GEL-VGN), (4) RA—reporter acute group (
n = 2, after stroke, animals were infused with GEL-GFP), (5) RP—reporter preventive group (
n = 2, before stroke, animals were infused with GEL-GFP), and (6) Int—intact group (
n = 3, healthy animals were used to obtain basic molecular and immunofluorescence data for the comparative analysis with the results of experimental mini-pigs). In control and therapeutic groups, the animals were euthanised on day 21 after stroke modelling. The endpoint of the experiment was chosen based on studies of the recovery from arm disability within 3 weeks, observed in 80% of patients with stroke [
17], and the 3-week period of adenoviral vector activity (effective expression of transgenes) [
18].
2.5. Behavioural Tests
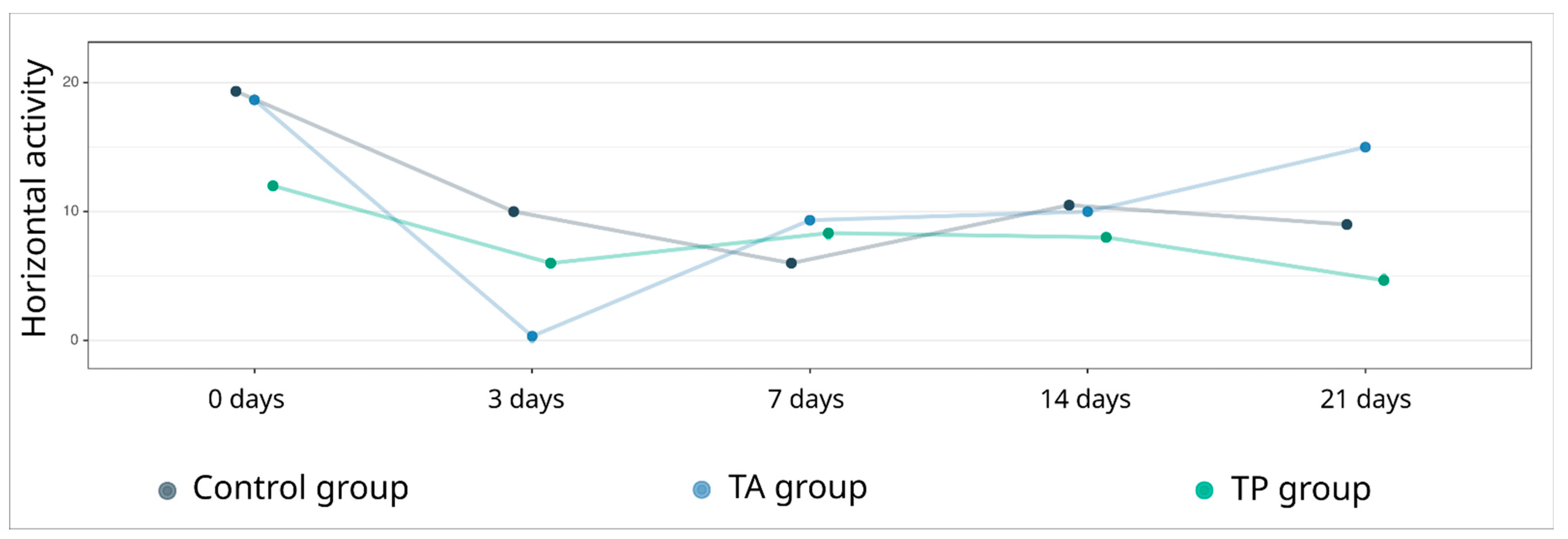
The “open field” test was used to evaluate the general locomotor activity levels of the experimental animals before and on days 3, 7, 14, and 21 after stroke modelling. The mini-pigs were placed in an arena (3 × 3 m) divided into 6 squares, and the horizontal activity was assessed by counting the number of lines crossed in 10 min, conducted by two supervisors in a blind manner in relation to the test animals.
2.6. Post-Ischaemic Changes in the Brain
2.6.1. Sample Collection
At 21 days post-surgery, the control (Con) and treated animals (TA and TP) were anaesthetised, as described above, and were infused with potassium chloride (150 mg/kg) via the auricular vein. The brains were extracted from the cranial cavity and fixed in 4% paraformaldehyde solution. The brains of mini-pigs from the RA and RP groups were harvested 7 days after infusion with the GEL-GFP.
2.6.2. Morphological Analysis of the Cerebral Infarction
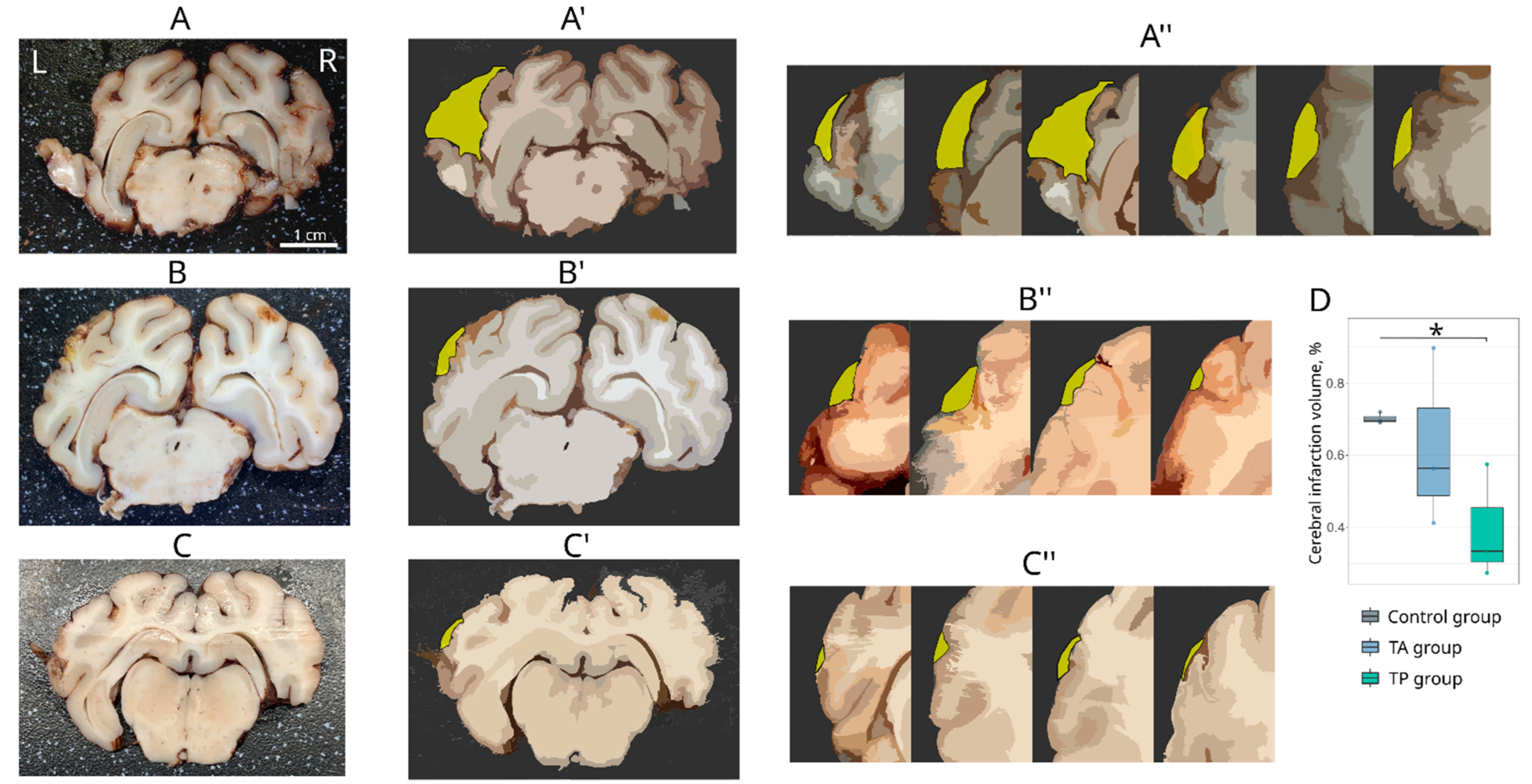
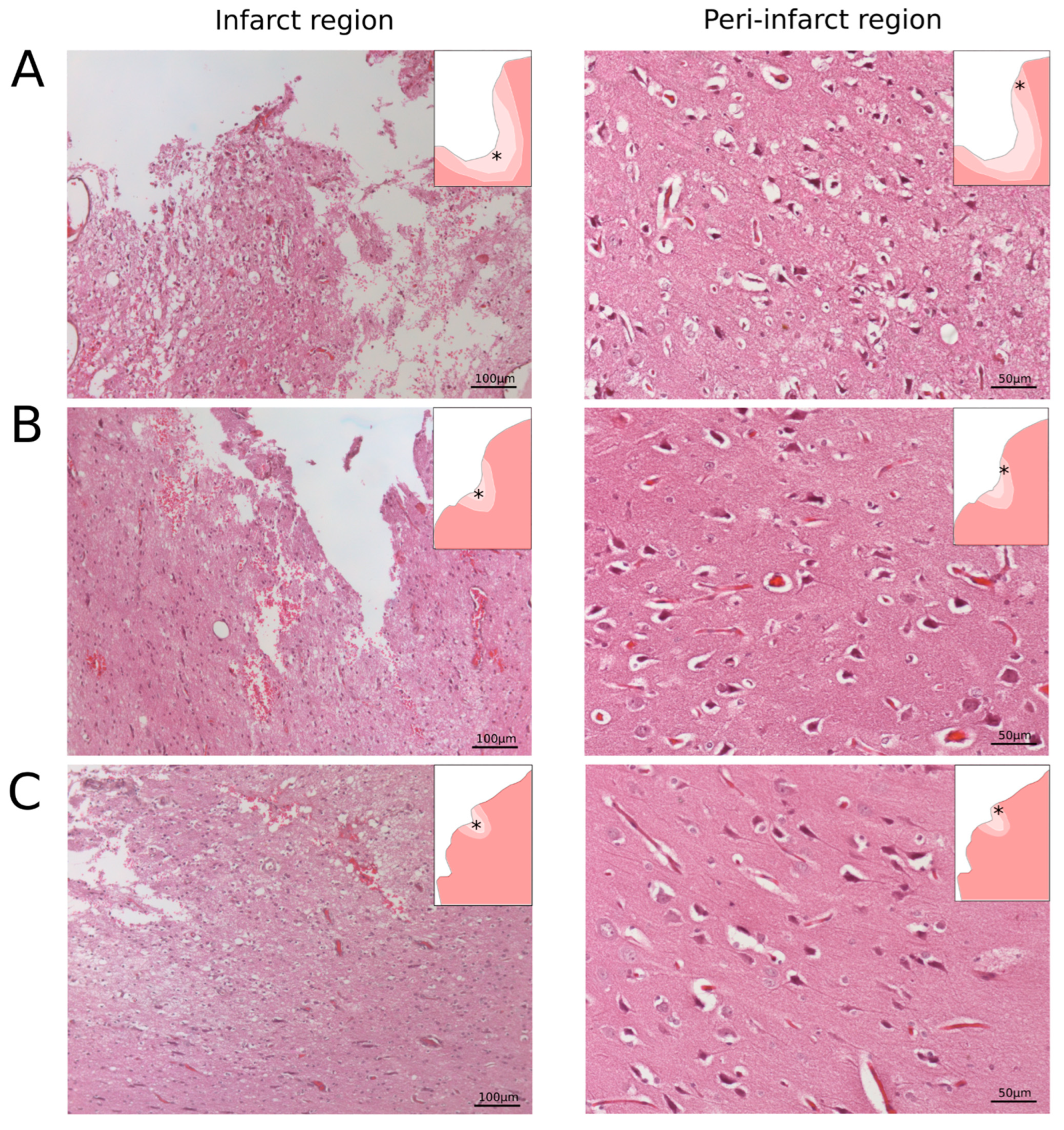
To analyse the area of the infarction, digital images of the left hemispheres were captured. Afterwards, the whole brains were cut into 3 mm-thick frontal slices. The number of obtained slices varied from 4 to 7 and depended on the size of the ischemic injury area. For the morphometric analysis of the infarct volume, each 3 mm slice was photographed from both (frontal and caudal) sides. The absolute infarct volume (AIV) was calculated using digital images of each 3 mm-thick slice obtained from the infarct region in the left cerebral hemisphere. The relative infarct volume (RIV) was calculated as AIV/(AIV + V of brain). All morphometric analyses were performed using ImageJ software (NIH). Paraffin embedded brain tissue was cut with a rotary microtome HM 325 (Thermo Scientific, Waltham, WA, USA). The obtained frontal slices were stained with haematoxylin and eosin. For the histological analysis, digitised images of the brain cortex were taken with a microscope, Axioscope A1 (Carl Zeiss, Oberkochen, Germany).
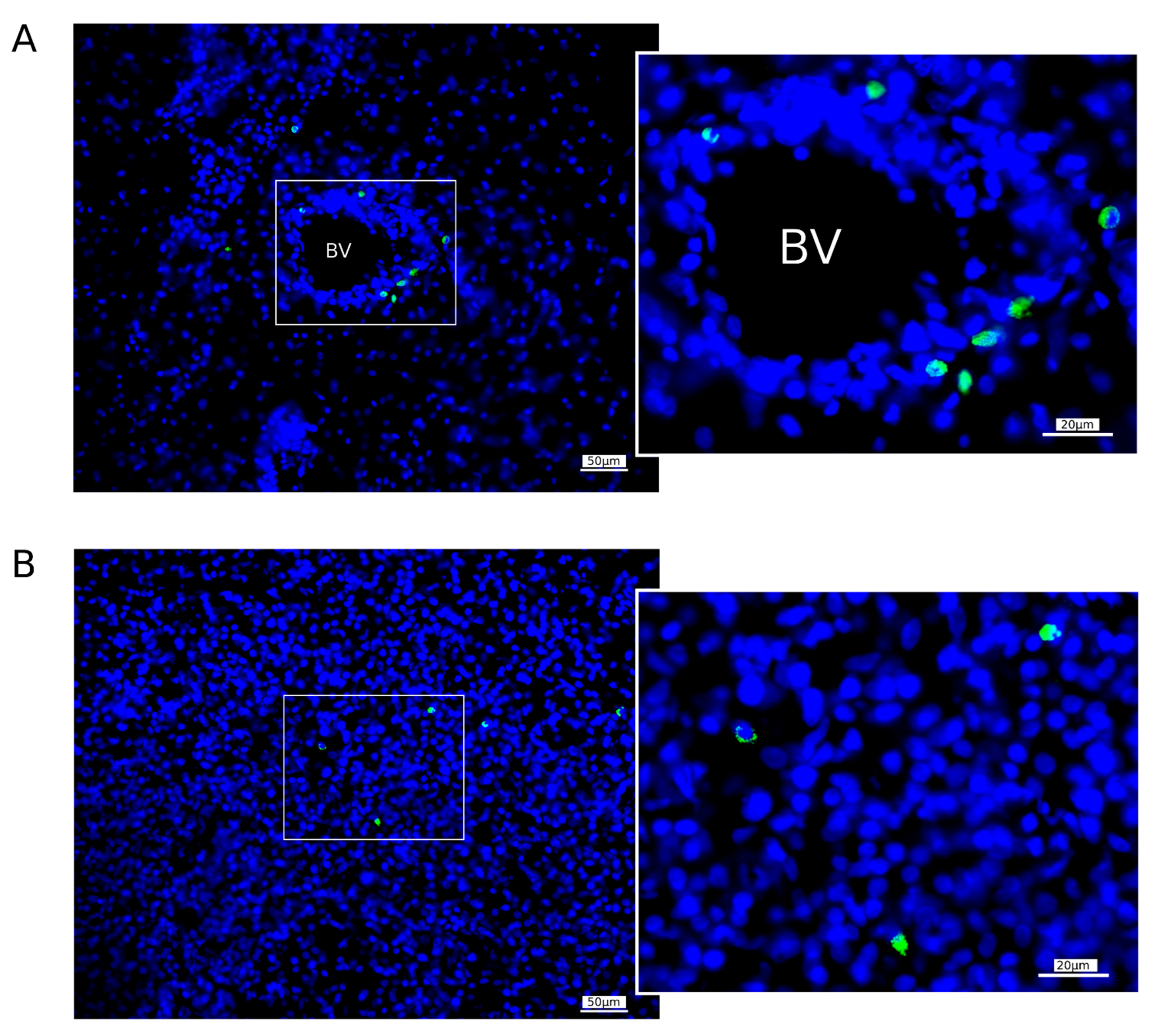
2.6.3. Homing of WBCs Expressing GFP in the Brain of Mini-Pigs
The migration of WBCs producing GFP in the brains was studied using fluorescent microscopy 7 days after intravenous the infusion of GML-GFP into the mini-pigs with stroke in the RA and RP groups. Samples from the left (peri-infarct region) hemisphere were fixed in 4% paraformaldehyde solution, cryoprotected in 30% sucrose, and embedded in tissue-freezing medium, and frozen 20 μm sections were prepared using a Microm HM 560 cryostat (Thermo Fisher Scientific, Waltham, WA, USA). Slide-mounted sections were incubated in DAPI solution (10 μg/mL in PBS; Sigma) and embedded in antifade mounting medium, and digital images were taken with a luminescence microscope (Axioscope A1, Carl Zeiss, Germany). GFP-positive WBCs were counted in 10 sections of the peri-infarct region of each mini-pig. The cell counting was performed based on digital images in an area of 600 × 500 μm. Average cell counts for each animal were standardised according to the area of 1 mm2.
2.6.4. Immunofluorescence Staining
The immunofluorescence analysis of the molecular and cellular changes in the peri-infarct area was performed using the 20 μm slide-mounted sections, using primary antibodies (Ab) to specific cell markers (
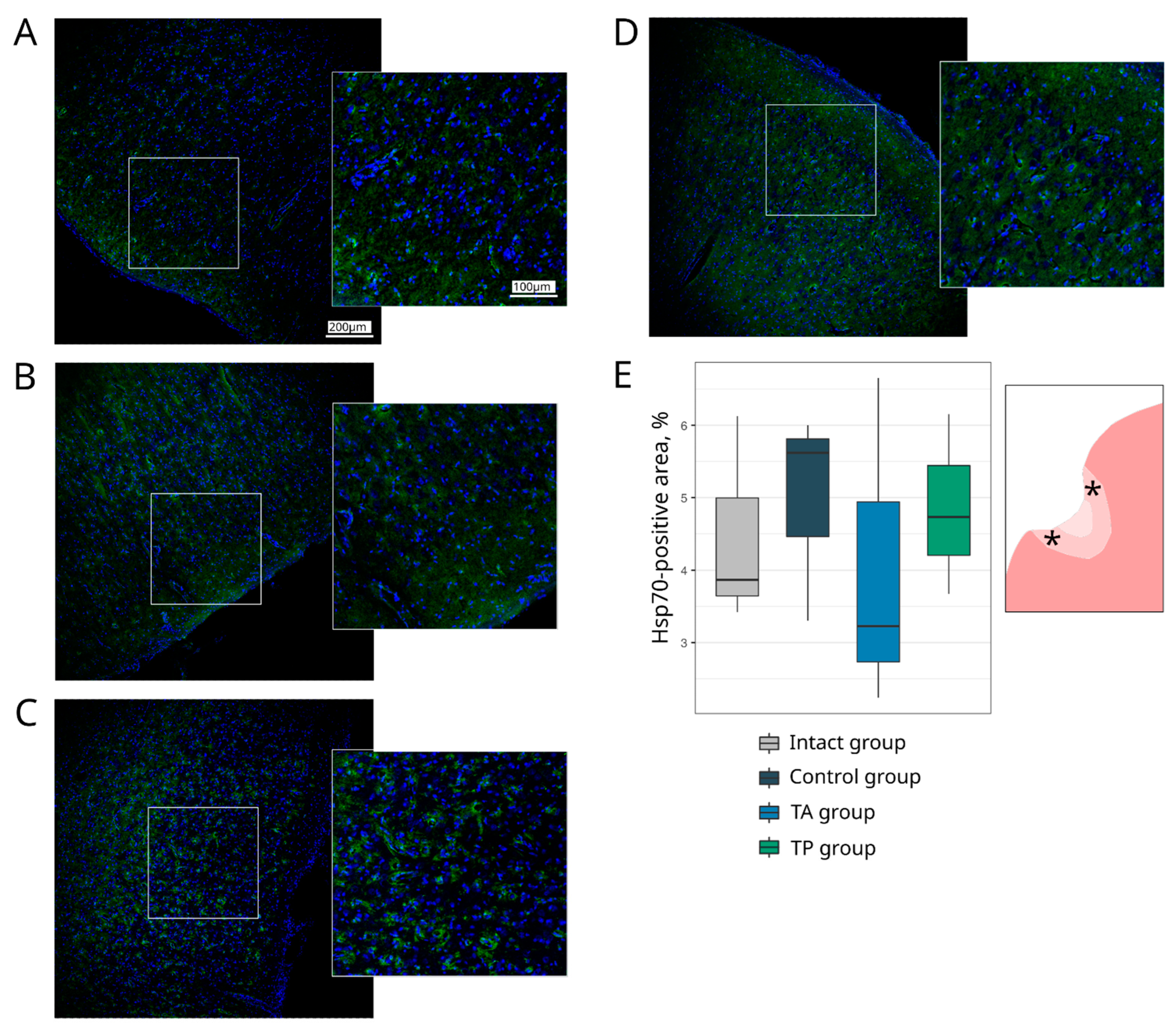
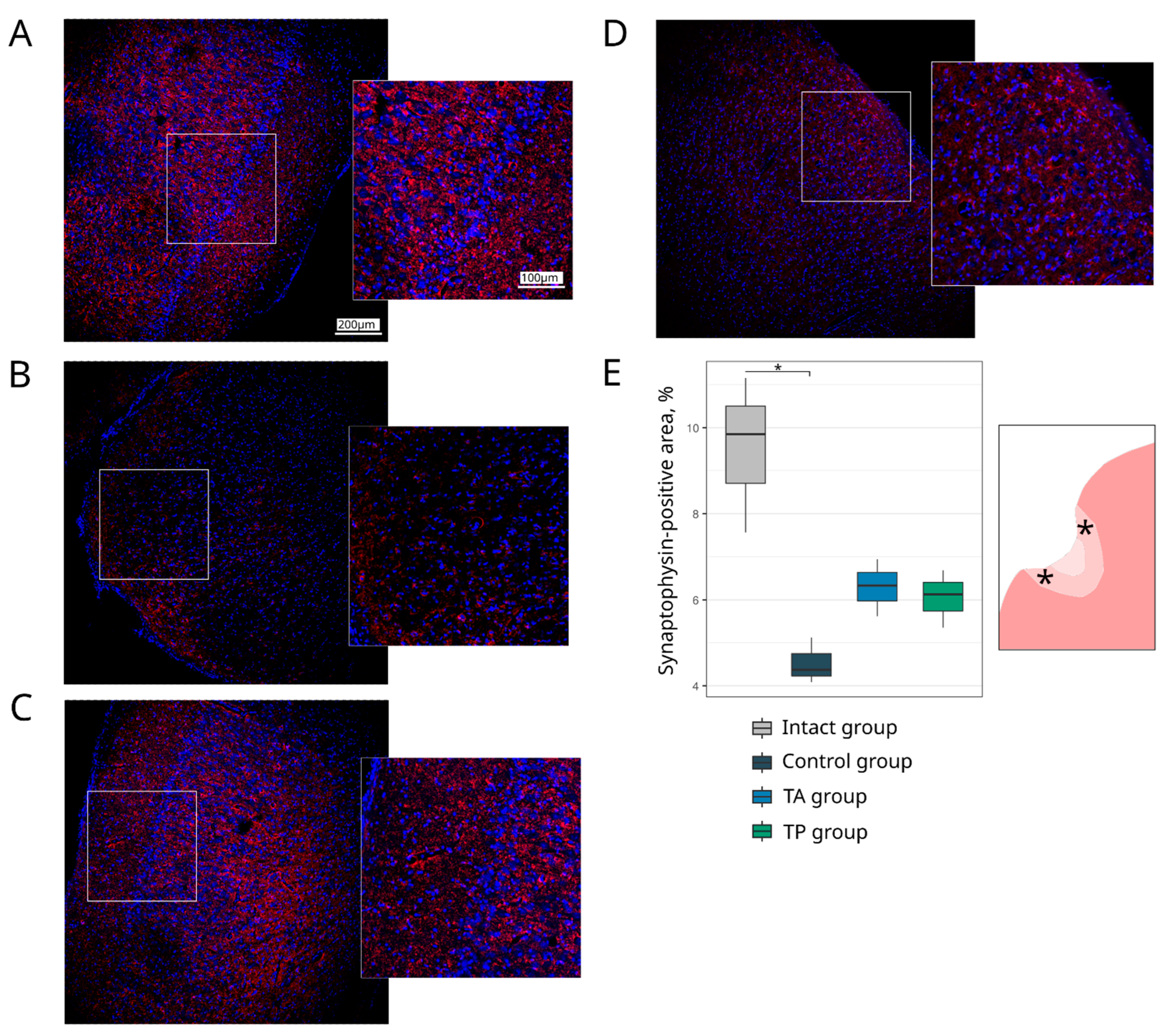
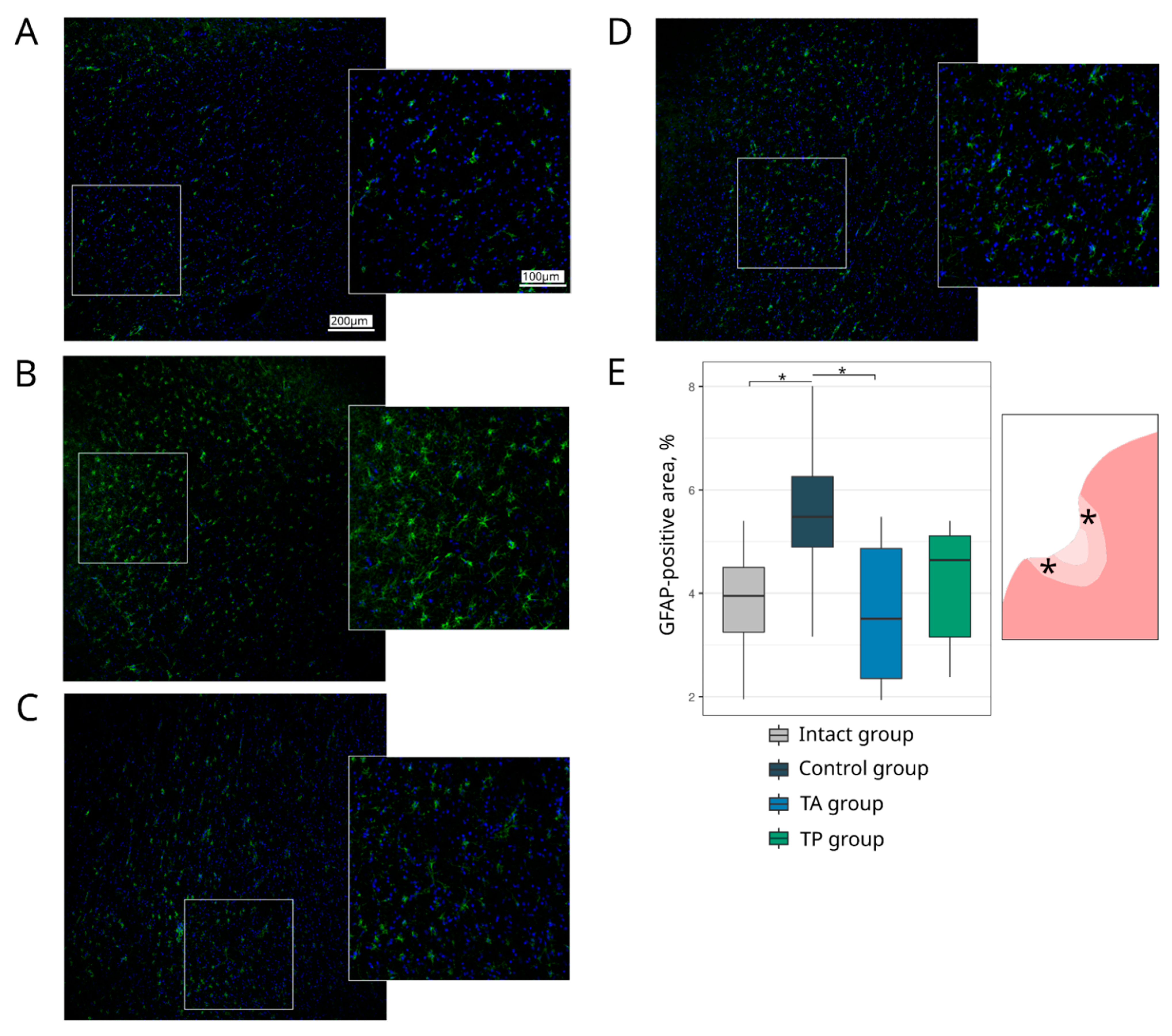
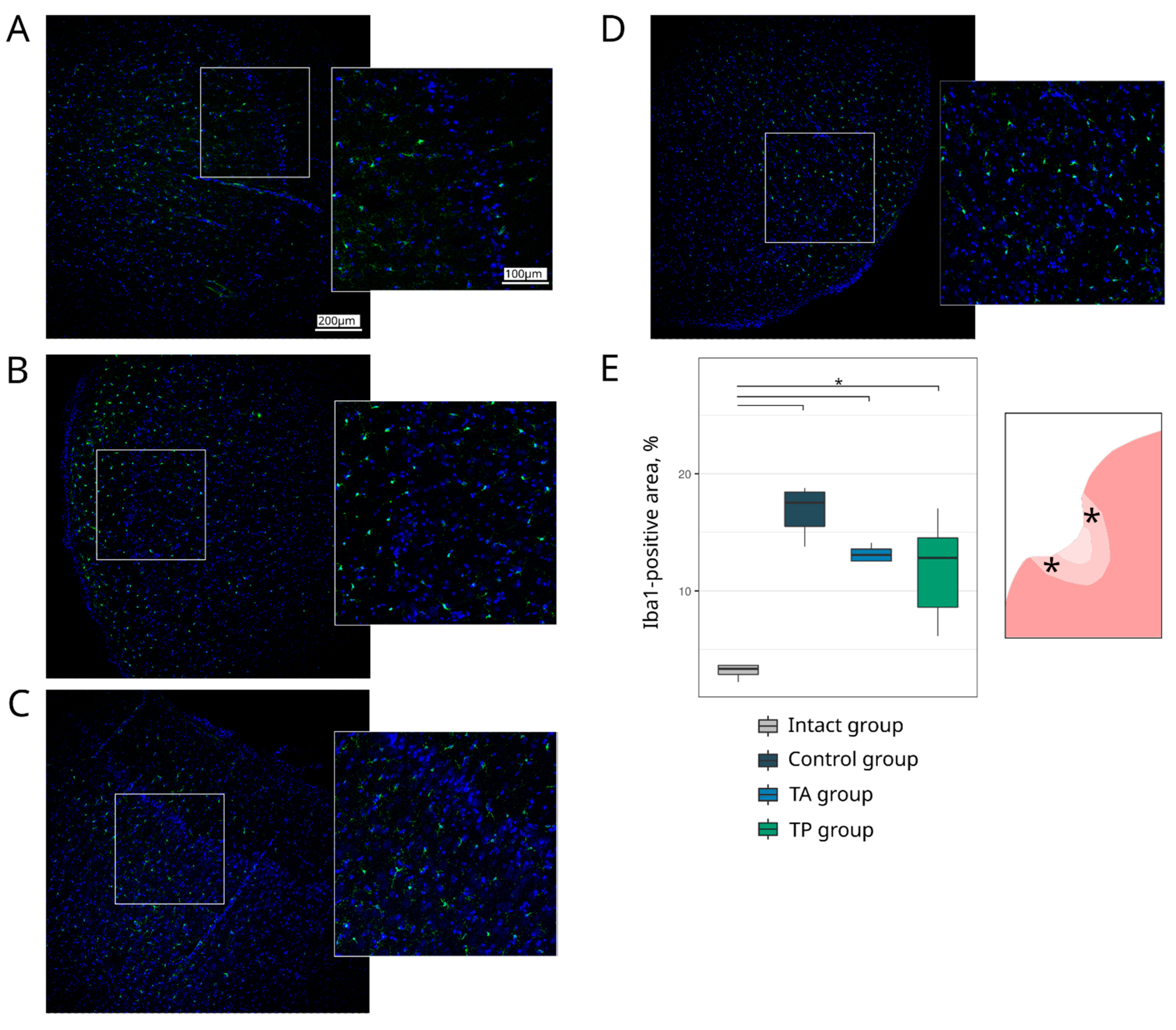
Table 1). The survivability of brain cells was evaluated using the Ab to pro-apoptotic protein (Caspase3) and heat shock protein 70 kDa (Hsp70). The expression of the synaptic proteins was assessed with the Abs against synaptic vesicle protein (Synaptophysin) and postsynaptic density protein 95 kDa (PSD95). The reaction of the neuroglial cells was evaluated using the Ab to glial fibrillary acidic protein (GFAP) for astrocytes, the Ab to oligodendrocyte transcription factor 2 (Olig2) for oligodendroglial cells, and the Ab to ionised calcium binding adaptor molecule 1 (Iba1) for microglial cells.
Appropriate secondary Abs were employed for an immune reaction with the primary Abs (
Table 1). The cell nuclei were stained with DAPI (10 μg/mL in PBS, Sigma, St. Louis, MO, USA). Slices were embedded in glycerol (GalenoPharm, Saint Petersburg, Russia) and observed under a confocal LEICA TCS SP5 MP microscope (Leica Microsystems, Wetzlar, Germany). The images taken were used to analyse the counts of Caspase3 and Olig2 immunopositive nuclei (calculated taking into account the DAPI staining) and the immunopositive areas for Hsp70, Synaptophysin, PSD95, GFAP, and Iba1 (presented in percentages).
Regarding the quantitative evaluation, a total of 60 sections of the brain cortex were obtained by acquiring two adjacent 3 mm slices from the epicentre of the ischemic lesion from each animal. For the immunofluorescence staining, 6 sections were taken for each molecular marker. The immunoreactivity pattern of each target antigen in a single section was assessed over a total area of 0.5 mm2 using ImageJ software (NIH).
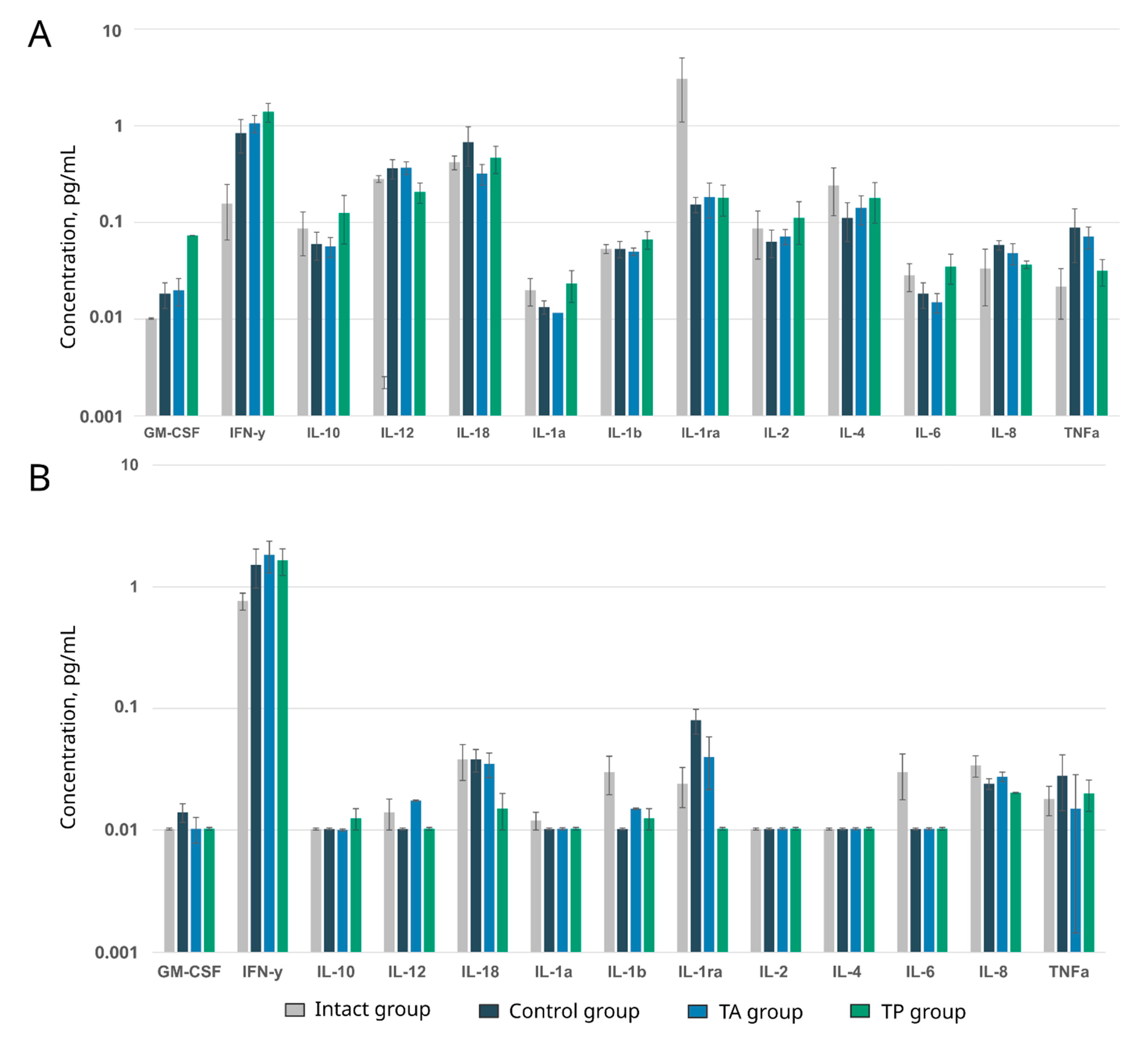
2.7. Analysis of Multiplex Cytokines, Chemokines, and Growth Factors
The multiplex analysis was performed using the Porcine Cytokine/Chemokine MILLIPLEX® magnetic-bead-based multi-analyte panel, containing the following target analytes: GM-CSF, IL-1ra, IL-2, IL-6, IL-10, IL-12, IL-18, TNFα, IFNγ, IL-1β, IL-1α, IL-4, and IL-8 (Millipore, Burlington, MA, USA). Blood and cerebrospinal fluid samples were obtained from the intact mini-pigs and each animal in the control and therapeutic TA and TP groups 3 weeks after stroke modelling and were appropriately processed, frozen at a temperature of −80 °C, and stored until the time of the study. The data were analysed using a MAGPIX® system and the Analyst 5.1 software (Merck, Kenilworth, NJ, USA).
2.8. Statistical Analysis
The data processing, analysis, and visualisation were performed using R 4.1.0 (R Foundation for Statistical Computing, Vienna, Austria). Descriptive statistics are presented as the mean (standard deviation) for the behavioural tests and as the median (1st–3rd quartiles) for the morphometric analysis. The Kruskal–Wallis test, combined with Dunn’s test, as a post hoc method, was used for the group comparison. The results were visualized using box plots. Differences were considered statistically significant when p < 0.05.
4. Discussion
Various viral vectors are used to deliver therapeutic genes to the CNS [
21]. However, the problems of mutagenesis, immunogenicity, and the duration of gene expression in direct (in vivo) gene therapy remain relevant to this day. For the temporary expression of transgenes, the cell-mediated approach to gene delivery is considered an effective and safe method of gene therapy. In stroke gene therapy, for the delivery of the recombinant neurotrophic factors to the ischaemic brain, mesenchymal stem cells [
22,
23,
24] and neural stem cells [
25] have been used. In our previous studies on the rat MCA occlusion model, we employed ex vivo gene-engineered umbilical cord blood mononuclear cells (UCB-MC), simultaneously transduced with three adenoviral vectors (Ad5) carrying
vegf165, gdnf, and
ncam1, for cell-mediated gene therapy in the acute phase [
10] and for the preventive treatment [
11] of stroke. Our results demonstrated the positive effect of the intrathecal injection of UCB-MC, expressing recombinant VEGF, GDNF, and NCAM, on the reduction in the negative consequences of ischaemia-induced brain injury. Recently, for the delivery of the same therapeutic gene combination to the mini-pig post-traumatic spinal cord, for the first time, we used an intravenous administration of the autologous GEL as a complementary therapy to the supra- and sub-lesional epidural electrical stimulation [
13]. The study demonstrated the prospects of using genetically enriched leucoconcentrate for the stimulation of neuro-regeneration in the CNS. The rationale for the preparation of the autologous, genetically enriched leucoconcentrate for cell-mediated gene therapy is based on its simplicity and biosafety. In the current study, we employed this strategy for stroke treatment in the acute phase and for preventive gene therapy in a mini-pig stroke model.
To test the utility of the autologous, genetically enriched leucoconcentrate for stroke gene therapy, we developed an original, reliable, and reproducible model of ischaemic stroke in mini-pigs based on previously reported methods. In the study, we hypothesized two possible mechanisms for the positive effect of GEL-VGN on the post-ischemic brain recovery: (1) genetically modified WBCs can pass through the disrupted blood–brain barrier [
26] and occupy the ischemic region, so that secreted recombinant VEGF, GDNF, and NCAM can act on the target cells locally through a paracrine mechanism; and (2) recombinant VEGF, GDNF, and NCAM secreted by the genetically modified WBCs into the bloodstream can diffuse through the leaky blood–brain barrier into the brain tissue and modulate its neuroplasticity through an endocrine mechanism.
The model of focal cerebral infarction in mini-pigs, using the method of the occlusion of the MCA through the transorbital approach, was examined for the first time by Sakoh et al. [
27]. Imai et al. were the first to use a craniotomy to access the MCA instead of a transorbital approach [
28]. In this study, we combined the occlusion of the distal branches of the MCA via a trepanation hole with the occlusion of the right common carotid artery to reduce the blood flow in the circle of Willis and observed a reproducible focus of ischemic cerebral cortex infarction in the temporal lobe of the left hemisphere, accompanied by moderate locomotor dysfunction. At day 21 after surgery, an immunofluorescence study of the peri-infarct region revealed apoptotic cells; an increase in the area occupied by microglia and astrocytes, which eventually leads to the formation of a glial scar; a decrease in the number of oligodendroglial cells responsible for myelination; and the decrease in the synaptic protein expression by neural cells. The intravenous infusion of the autologous leucoconcentrate enriched with
vegf165,
gdnf, and
ncam1 genes into the mini-pigs in the acute phase of the stroke or 2 days before the modelling of the stroke (preventive gene therapy) resulted in the positive remodelling of the peri-infarct region, including: (1) a smaller infarct volume and number of apoptotic cells; (2) restraining of the organisation of the glial scar; (3) a higher number of myelin-forming oligodendroglial cells; and (4) the restoration of the functional activity of neurons, according to the levels of the synaptic vesicle protein Synaptophysin and postsynaptic density protein (PSD95) expression. The results of the cytokine, chemokine, and growth factor profiling of the blood and cerebrospinal fluid in the experimental mini-pigs should also be considered positive. We found no difference in the levels of main pro-inflammatory (IL-1, IL-6, and TNF-α) and anti-inflammatory (IL-10, IL-12) cytokines in the blood and cerebrospinal fluid of the control and treated mini-pigs when compared with the intact animals. These data demonstrate the termination of the inflammatory response 3 weeks after stroke modelling, although the higher contents of IL-1ra (proinflammatory inhibitor IL1β) in the blood and IL-6 (neuropoietic cytokine) in the cerebrospinal fluid in the intact animals, compared to the experimental mini-pigs, show the incomplete recovery of the animals with stroke. In addition, the similar levels of investigated cytokines, chemokines, and growth factors in the control and treated animals may indicate the absence of immunogenic effects of the genetically modified WBCs at the local and systemic levels.
One of the important issues regarding the strategy of using gene therapy for stroke is the targeted delivery of recombinant genes to the site of ischemic damage. WBCs are a cell type with a high synthetic and secretory activity and the ability to migrate successfully from the bloodstream to the affected tissues. In the study, we demonstrated the potential of genetically modified WBCs to pass through the disrupted blood–brain barrier and occupy the ischemic area. The presence of the gene-modified WBCs expressing GFP in the peri-infarct region of the experimental mini-pigs one week after the administration of the GEL-GFP in the acute phase of stroke or 2 days before the stroke modelling supports our hypothesis, stating that after intravenous infusion of the autologous GEL-VGN, gene-modified WBCs migrate to the brain ischaemic injury region and locally produce the recombinant therapeutic molecules. In the acute phase of stroke, the compromised blood–brain barrier leads not only to the migration of gene-modified WBCs but is also permeable and subject to the infusion of recombinant VEGF, GDNF, and NCAM secreted by gene-modified WBCs into the bloodstream. Thus, the positive effect of GEL-VGN on post-ischemic brain recovery may be due to the action of the therapeutic molecules through paracrine and endocrine mechanisms. VEGF and GDNF have known neuroprotective properties and are widely used in gene therapy studies of neurodegenerative diseases, neurotrauma, and stroke. It is assumed that NCAM plays a role not only as a stimulator of regeneration [
29] but also in ensuring the homing and survival of genetically modified leucocytes in the brain tissue. Due to the lack of ability of the adenoviral vector to integrate into the recipient’s genome, the temporary secretion of the recombinant VEGF, GDNF, and NCAM continues for approximately 3 weeks [
18].
The issue of stroke prevention in patients at high risk of ischemic stroke, combined with transient ischemic attacks of different origins, requires constant clinical examination and treatment with anticoagulants. Our concept of preventive personalized gene therapy is based on the intravenous infusion of GEL-VGN in patients at high risk of stroke. An autologous leucoconcentrate, enriched with genetic material, may be prepared in hospital blood centres using a routine unit of a patient’s peripheral blood and chimeric adenoviral vectors (Ad5/35) carrying recombinant therapeutic genes. Importantly, the procedure for preparing the genetically enriched leucoconcentrate is performed in a standard blood bag, without the use of any animal products, antibiotics, or other chemicals for culturing cells in vitro. This approach can reduce the mass death of neurons in the ischemic penumbra area during the 3–6 h period of the therapeutic window at the onset of stroke, and it could become an important component of emergency therapy [
30].
For the first time, in this study, we demonstrated a new approach to the use of personalized ex vivo gene therapy for stroke based on a simple, economical, and safe platform for the delivery of the recombinant therapeutic gene/genes using an autologous leucoconcentrate, prepared from the patient’s peripheral blood. The transduction of WBCs to the leucoconcentrate was performed in a blood container using a chimeric adenoviral vector (Ad5/35), which has a modified fibre with a high affinity for the CD46 membrane molecule, expressed in all WBCs [
31], without culturing in vitro and without any risks stemming from the use of animal-origin bioproducts and antibiotics.
Other important issues regarding stroke therapy include the development of treatments under the threat of stroke. Preventive gene therapy can restrain ischemic brain damage after the onset of a stroke. The presence of the recombinant neuroprotective molecules can increase the survivability of neurons in the ischemic area immediately after the onset of a stroke. The results obtained in the study confirmed our hypothesis of the rationality of preventive gene therapy based on the intravenous infusion of autologous GEL-VGN under the threat of stroke.
A comparative analysis of the present results and data from our previous studies on triple-gene therapy for stroke in rats with MCO, using the intrathecal injection of human genetically engineered UCB-MC expressing recombinant molecules VEGF, GDNF, and NCAM in the acute phase [
10] and for preventive treatment [
11], revealed similar effects of gene therapy on the cortical remodelling in rats and mini-pigs. These data demonstrate, on the one hand, the consistency of the findings on the efficacy of triple-gene therapy for stroke treatment in small and large animals and, on the other hand, the efficacy of the intravenous infusion of autologous genetically modified WBCs. Thus, based on our previous investigations and the present study, we propose a new strategy for personalized cell-mediated gene therapy for stroke treatment based on the intravenous infusion of the autologous GEL temporally secreting neuroprotective factors (VEGF, GDNF, and NCAM).

 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}