1. Introduction
The gold standard in viral disease management is prevention, which is vaccination. Pandemics with new viruses remind us that the development and approval of vaccines takes time. Moreover, vaccines are not 100% efficient, and especially elderly people and immunocompromised individuals may not generate sufficient protection against infectious agents. Therefore, powerful broadly acting antivirals with a more general inhibitory mechanism of action are required.
Non-enveloped enteroviruses are very stable and stay infective on surfaces and in the environment for long periods. Their entry is mostly via the gastro-intestinal tract, but also through the upper respiratory system. They cause a high number of acute infections such as flu, aseptic meningitis and myocarditis, but also contribute to chronic diseases such as dilated cardiomyopathy, asthma or type I diabetes [
1]. For enteroviruses, no approved antivirals exist, and there are vaccines available only against poliovirus and EV71 [
1]. Therefore, it is necessary to reduce the viral load on the surfaces and in the environment and prevent infection in the primary infection sites. An optimal solution to tackle virus infection would be a drug that directly kills virus infectivity and additionally prevents the virus from entering cells.
Nature provides a large variety of potential antimicrobials and antivirals. Those may be of endophyte origin, secreted by the symbiotic fungi or bacteria living in plants [
2]. One example is Resveratrol (3,5,4′-trihydroxy-trans-stilbene), which is a polyphenol, non-flavonoid compound and is produced by plants in response to cellular damage or pathogen attacks [
3]. Resveratrol (RES) is found in strongly pigmented vegetables and fruits such as the skin of grapes, blueberries and peanuts, and it has strong bioactive potential [
3,
4,
5]. Like RES, flavonoids are polyphenols found as secondary metabolites in vegetables, fruits, nuts, red wine and tea. They share great similarity in structure to RES and great promise to be used as antivirals [
6,
7,
8].
In the present study, EGCG and RES were tested for their antiviral efficacy, and the mechanisms of action was elucidated against the stable non-enveloped enterovirus B group viruses. While RES is considered completely non-toxic [
9], high amounts of free EGCG show in vitro hepatotoxic effects, and in particular in cases in vivo [
10,
11]. Therefore, here, free polyphenols as well as polyphenols functionalized onto gold nanoparticles (AuNPs) were studied for their antiviral efficiency, as binding polyphenols to AuNPs can reduce toxicity. The effect of reduced toxicity by nanoparticle binding has been shown previously for molecules with a known hepatotoxic profile [
12]. AuNPs were synthesized by green chemistry using the polyphenols as redox agents to further reduce the toxic components in the preparations. AuNPs synthesized using green chemistry have been previously reported to act as antivirals [
13].
In this work, we show that both RES and EGCG have a direct and long-lived antiviral effect through binding to multiple sites on the virus particles, preventing the virus from opening, reducing the binding to the cell surface and inducing clustering of the virions.
2. Materials and Methods
2.1. Cells
Human alveolar basal epithelial adenocarcinoma (A549) cells and Green Monkey Kidney (GMK) cells were obtained from American type culture collection (ATCC, Manassas, VA, USA). The A549 and GMK cell lines were propagated in Dulbecco’s Modified Eagle Medium (DMEM) (Gibco, Paisley, UK) and Eagle’s Minimum Essential Medium (MEM) (Gibco, Paisley, UK) respectively, supplemented with 10% Fetal Bovine Serum (FBS, Gibco, Paisley, UK), 1% L-GlutaMAX (Gibco, Paisley, UK) and 1% antibiotics (penicillin/streptomycin) (Gibco, Paisley, UK) in a humidified 5% CO2 incubator at 37 °C.
2.2. Viruses
Coxsackievirus B3 (CVB3; Nancy strain), Coxsackie virus B1 (CVB1, CONN 5 strain) and Coxsackievirus A9 strain (CVA9; Griggs strain), obtained from ATCC, were produced and purified as described before [
14,
15,
16], with the only exception of adding 0.1% (
v/
v) TWEEN
® 80 (Sigma-Aldrich, Steinheim, Germany) during the freeze–thaw cycle.
2.3. Polyphenols
Epigallocatechin gallate (EGCG) and Resveratrol (RES) were purchased from Sigma-Aldrich (Munich, Germany). EGCG was dissolved in water and Resveratrol in 0.02 M NaOH. Stock concentrations of EGCG and RES were 6.5 mM (3 mg/mL) and 8.8 mM (2 mg/mL) respectively, and were stored at −20 °C.
2.4. Nanoparticle Preparations
The green chemistry approach using the polyphenolic compounds as redox agents was chosen to reduce the number of potentially toxic molecules present on the surface of the gold nanoparticles (AuNPs). These polyphenols are supposed to form a stable and stabilizing layer on the AuNPs, as it was observed for citrate AuNPs shown by Mandal et al. [
17]. For the preparation of RES-coated gold nanoparticles (RES-AuNPs), we used a modified protocol by Sanna et al. [
18]. In brief, 20.8 mg of HAuCl
4·3 H
2O (Sigma-Aldrich, Milan Italy) was dissolved in 12 mL of bi-distilled water on ice. Next, an ice-cold light-yellow solution of 3.0 mg of RES in 1.5 mL of 0.02 M NaOH solution was quickly added under vortexing. Under continuous vortexing, the temperature of the solution was slowly raised to room temperature (RT). Immediately after RES addition, the solution became brownish, after 10 min reddish, and after vortexing at 500 rpm for 1 h, the color turned into wine red. Unbound RES was removed by a series of washing (first wash: 0.02 M NaOH; second wash: H
2O) and centrifugation steps (2 × 14,000 rpm, 20 min at 4 °C to remove NaOH; 1 × 12,000 rpm, 20 min at 4 °C to remove H
2O), and the pellet was finally resuspended in water. The centrifugation was carried out by a Microfuge 40R with a Fiberlite F21-48 x2 Fixed Angle Rotor, 45°, R
max 97 mm (ThermoFisher, Rome, Italy). The final gold concentration was 5.3 mg/mL, as determined by ICP-MS. The concentration of RES in the stock solution was calculated (
Supplementary Material) to be 34.4 μM.
For the preparation of EGCG-AuNPs, 5.9 mg of EGCG was dissolved in 15 mL of bi-distilled water and cooled on ice. An ice-cold solution of 12.9 mg of HAuCl
4·3 H
2O in 2 mL of H
2O was added quickly and mixed by shaking on ice for another 10 min. After the color changed from bright yellow to red within 1 min, the temperature of the solution was slowly increased to RT for 2 h under continuous vortexing at 500 rpm. The excess EGCG was removed by a series of washing and centrifugation steps (2× centrifugation at 14,000 rpm for 20 min at 4 °C addition of ddH
2O, 1× centrifugation at 12,000 rpm for 20 min at 4 °C). The deep red pellet was resuspended in water. The final gold concentration was 1.64 mg/mL, as determined by ICP-MS. The concentration of EGCG in the stock solution was calculated (
Supplementary Material) to be 5.61 μM.
Quantitative Analysis of EGCG and RES Bound to the AuNP Surface
The amount of EGCG and RES present on the surface of the gold nanoparticles was estimated indirectly by the substitution with
l-cysteine. The thiol group of the cysteine binds stronger to the gold than the aromatic ring system of polyphenols, and therefore will replace it. The colorimetric method finally determines the remaining free thiol groups of the
l-cysteine, assuming a complete substitution of the polyphenols by cysteine [
19]. The residual-free thiol groups of
l-cysteine in solution after polyphenol exchange on the nanoparticle surface were quantified photometrically using Ellman’s reagent (5,5′-dithio-bis (2-nitrobenzoic acid)). The amount of polyphenols was then calculated considering a 1:1 ratio of replacement (1 cysteine replaces 1 polyphenol).
For its determination, a fresh stock solution of 1 mg of cysteine in 1 mL of 0.5 M phosphate buffer, pH 8.0, was diluted to a final concentration in l-cysteine of 102 mM. 100 μL of this solution was mixed with different volumes of EGCG-AuNPs or RES-AuNPs stock solution (ranging from 10 to 100 μL) and 0.5 M phosphate buffer solution at pH 8.0 to reach a final solution volume of 500 μL. This was mixed with 500 mL of Ellman’s reagent solution (3 mg dissolved in 10 mL of 0.5 M phosphate buffer, pH 8.0). The resulting mixture was incubated in the dark and at room temperature for 2 h. After this time, the samples were centrifuged at 13,000 rpm for 5 min (Mikro 22 R, Hettich Zentrifugen, Germany) and 100 µL of each sample was transferred into a microplate reader (PerkinElmer VICTOR X3 Multilabel) to measure absorbance at 450 nm. The amount of thiol groups was estimated from a standard curve of l-cysteine.
2.5. Dynamic Light Scattering Measurements
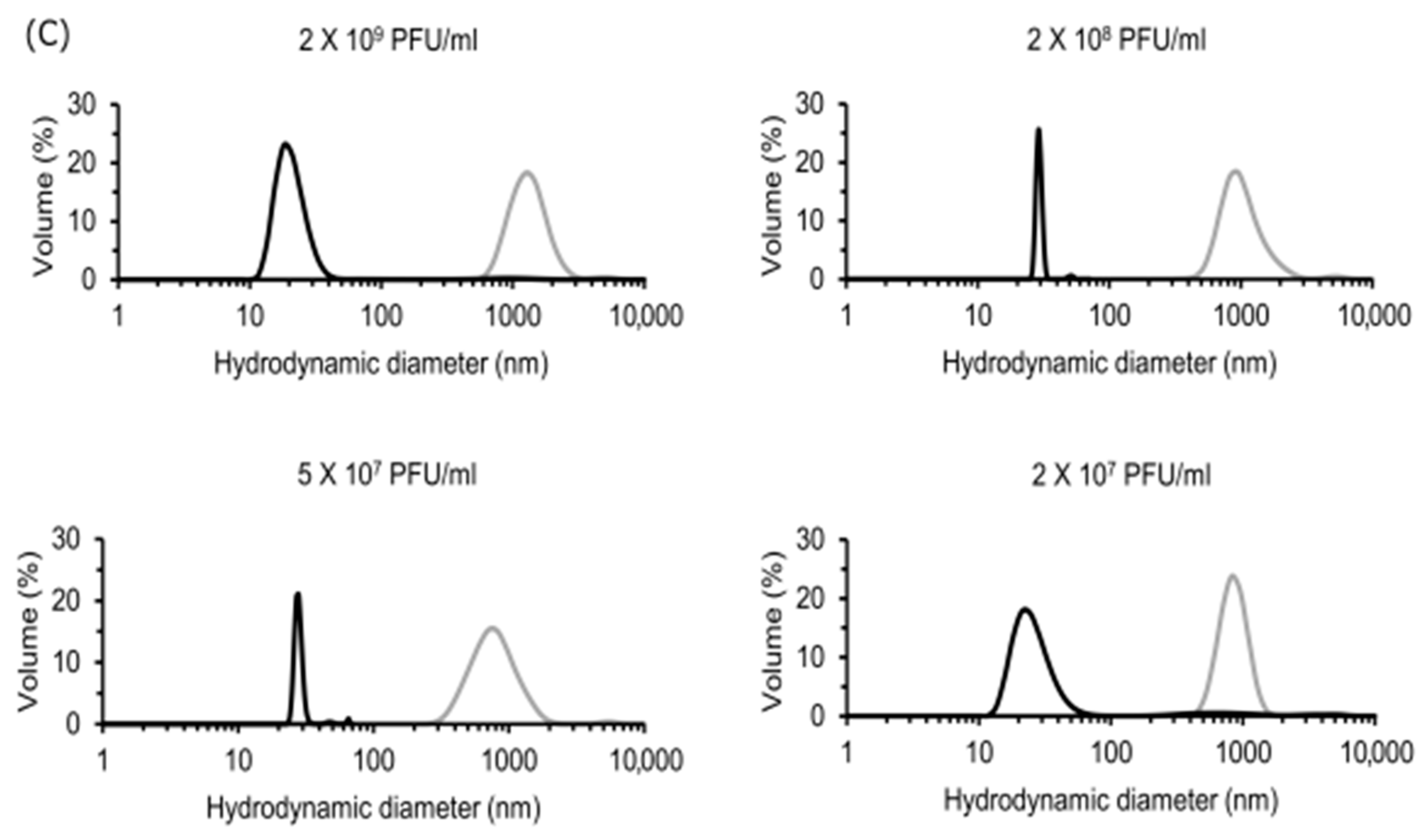
For dynamic light scattering (DLS) measurements, CVA9 virus samples were diluted in PBS from a stock solution (5 × 1010 PFU/mL) to 2 × 109, 2 × 108, 5 × 107 and 2 × 107 PFU/mL. EGCG was added to each of these dilutions such that its final concentration was 6.5 μM. Experimental virus controls with the same dilutions but without the EGCG were also prepared. Both the control virus and EGCG-treated virus were incubated for 1 h at 37 °C. Post-incubation, the virus/virus cluster size distribution was measured by DLS at 25 °C with the Zetasizer Nano ZS instrument (Malvern Instruments, Malvern, UK). The attenuator was set by the system according to scattering intensity of the sample and set measurement position at 3 mm. As controls, we measured both EGCG and PBS buffer alone. As expected, no particles were observed in the controls.
2.6. Antiviral Activity Assay
The antiviral activity of the polyphenols against species B enteroviruses was determined using a cytopathic effect (CPE) inhibition assay, modified from Schmidtke et al. [
20]. Briefly, A549 cells were cultured for 24 h at 37 °C in 96-well flat-bottomed microtiter plate (Sarstedt, Numbrecht, Germany) at a density of 12,000 cells/well in 100 µL of DMEM supplemented with 10% FBS, 1% GlutaMAX and 1% penicillin/streptomycin antibiotics. The following day, each of the viruses (2 × 10
7 PFU/mL for CVB3 and CVB1 and 2 × 10
8 PFU/mL for CVA9) were pre-incubated with the compounds for 1 h at 37 °C. The virus–compound mixture was further diluted 10 times with DMEM to obtain a final MOI of 10 for CVB3 and CVB1 and 100 for CVA9. This mix was then added to cells for 24 h in a humidified 5% CO
2 incubator at 37 °C. A virus without the compound was used as a positive control and a mock infection without the virus and compound as a negative control for the experiments. The development of a cytopathic effect (CPE) was monitored using light microscopy. The next day, the cells were fixed and stained for 10 min with CPE dye (0.03% crystal violet, 2% ethanol and 36.5% formaldehyde) after washing twice with PBS. After two washes with water, the stained viable cells were lysed using a lysis buffer (0.8979 g of sodium citrate and 1N HCl in 47.5% ethanol) to elute the crystal violet. Then, the absorbance was measured spectrophotometrically at 570 nm using the VICTOR
TM X4 multilabel reader (PerkinElmer, Turku, Finland). This assay was performed twice independently for each of the virus–compound concentrations.
2.7. Cytotixicity Assay
Cytotoxicity studies were performed using the CPE inhibition assay by adding the compounds (without any virus) to A549 cells. Mock infection was used as a negative control. This assay was performed twice independently.
2.8. Time of Addition Studies
Time of addition studies were performed using the CPE inhibition assay. Briefly, A549 cells were cultured for 24 h in 96-well flat-bottomed microtiter plate at a density of 12,000 cells/well in 100 µL of DMEM supplemented with 10% FBS, 1% GlutaMAX and 1% penicillin/streptomycin antibiotics. On the subsequent day, cells were infected with CVB3 (MOI 10) for 1 h at 37 °C. Afterwards, excess virus was removed by repeated washing. Then, fresh media (supplemented with 10% FBS, 1% GlutaMAX and 1% penicillin/streptomycin) containing EGCG (65 μM) and RES (880 μM) were added, and cells were incubated for 24 h. Virus control (without the compounds), compound control (without virus) and mock infection were used as controls for the studies. This assay was performed twice independently for each of the virus–compound concentrations, with 5 technical replicates in each experiment.
2.9. Radioactive Labeling of CVA9
The radioactive virus was produced in confluent monolayers of GMK cells, which were washed and incubated with PBS for 15 min at 37 °C and infected with CVA9 (Griggs strain; ATCC) diluted in low-methionine-cysteine medium (MP Biomedicals, Illkrich, France) supplemented with 1% FBS and 1% Glutamax. After 3 h of infection, 40 μCi/mL of radioactive [35S] methionine/cysteine (PerkinElmer, Boston, MA, USA) diluted in low-methionine-cysteine medium supplemented with 1% FBS was added to the cells, and virus replication was allowed to proceed for 24 h at 37 °C. After three freeze–thaw cycles, the supernatant was cleared by pelleting at 2500× g for 10 min. TWEEN® 80 (0.1% v/v) was added to the pellet and incubated on ice for 30 min. Post-incubation, centrifugation was performed for 10 min with 4000× g at 4 °C. The collected supernatant was added on a 2 mL sucrose cushion (40%). The cushions were ultracentrifuged with an SW-41 rotor at 151,263× g for 2.5 h (4 °C). All the liquid up until the cushion and first 500 μL fraction from the top were discarded, and the next 3 × 500 μL fractions were collected, which were pelleted with a 70Ti rotor at 90,140× g for 2 h at 4 °C. The concentrated virus was then layered on top of the 10 mL sucrose gradient (5–20%) and ultracentrifuged with a SW-41 rotor at 151,263× g for 2 h (4 °C). Fractions (500 µL) were collected from the top to the bottom of the cushion and small samples from each fraction were mixed with a scintillation cocktail UltimaGoldTM (PerkinElmer, Waltham, MA, USA) for counting the radioactivity (counts per minute) using a Tri-Carb® 2910TR liquid scintillation analyzer (PerkinElmer, Downers Grove, IL, USA). The 160S-containing fractions were pooled and used for the experiments (7000 CPM/μL; 1.4 × 109 PFU/mL).
2.10. Gradient Assay
Metabolically labeled CVA9 (70,000 CPM corresponding to 1.4 × 108 PFU/mL) was pre-treated with EGCG (6.5 μM) and RES (1.8 mM) for 1 h at 37 °C. A virus control (without the compounds but the same treatment) and fresh virus (without the polyphenols and without treatment) were used as positive controls for the assay. The test and control samples were further diluted 10 times using a buffer, PBS-MgCl2 (PBS containing 2 mM MgCl2). The samples were then layered over 10 mL sucrose (5–20%) gradients and ultracentrifuged with a SW-41 rotor at 151,263× g (average g value) for 2 h (4 °C) using the SW-41 rotor. Gradients were fractionated from top to bottom (500 μL fractions) and mixed with a scintillation cocktail for counting the radioactivity using a PerkinElmer Tri-Carb® 2910TR liquid scintillation analyzer.
2.11. Real-Time Fluorescence Uncoating Assay
The real-time fluorescence uncoating assay has been previously described for measuring uncoating of enteroviruses at 37 °C [
16,
21]. The assay is based on the fluorescence emitted by SYBR Green II (SGII) when bound to viral genomic RNA. The experiment was carried out in a 96-well plate with a total reaction volume of 100 μL. Each well contained 0.5 μg of Coxsackievirus B3, 10× SGII (Life Technologies, Eugene, OR, USA) diluted in double-distilled water (ddH
2O), polyphenol-functionalized nanoparticles EGCG-AuNPs (Au concentration 16.4 μg/mL, ligand concentration 5.61 × 10
−2 μM) and RES-AuNPs (Au concentration 53 μg/mL, ligand concentration 3.44 × 10
−1 μM) in storage buffer PBS-MgCl
2 and opening buffer (20 mM NaCl, 6 mM KH
2PO
4, 12 mM K
2HPO
4 and 0.01% faf-BSA). We had two experimental controls: one was a virus in the storage buffer and the other contained a virus in the opening buffer. The samples were prepared on ice to prevent any effect of temperature on uncoating. Additionally, we had controls to exclude fluorescence quenching/enhancement from other molecules in the reaction mix. RNAse at a final concentration of 10 μg/mL was used to distinguish between the fluorescence originating from porous intermediate virions (allowing entry of SGII inside) and the RNA in solution. The fluorescence was recorded every 15 min for an hour at 37 °C using the PerkinElmer Multilabel Reader Victor X4 installed with a F485 lamp filter, F535 emission filter and a counting time of 1 s.
2.12. Particle Stability Thermal Release Assay (PaSTRy)
The PaSTRy assay was performed as described before [
22]. The experiment was carried out in thin-walled PCR plates (Agilent, Amstelveen, Netherlands). A reaction mixture containing 0.5 μg of Coxsackievirus B3 and EGCG (65 μM) in storage buffer or opening buffer was preincubated for 1 h at 37 °C and 5% CO
2. Post-incubation, 10× SGII (Invitrogen) in ddH
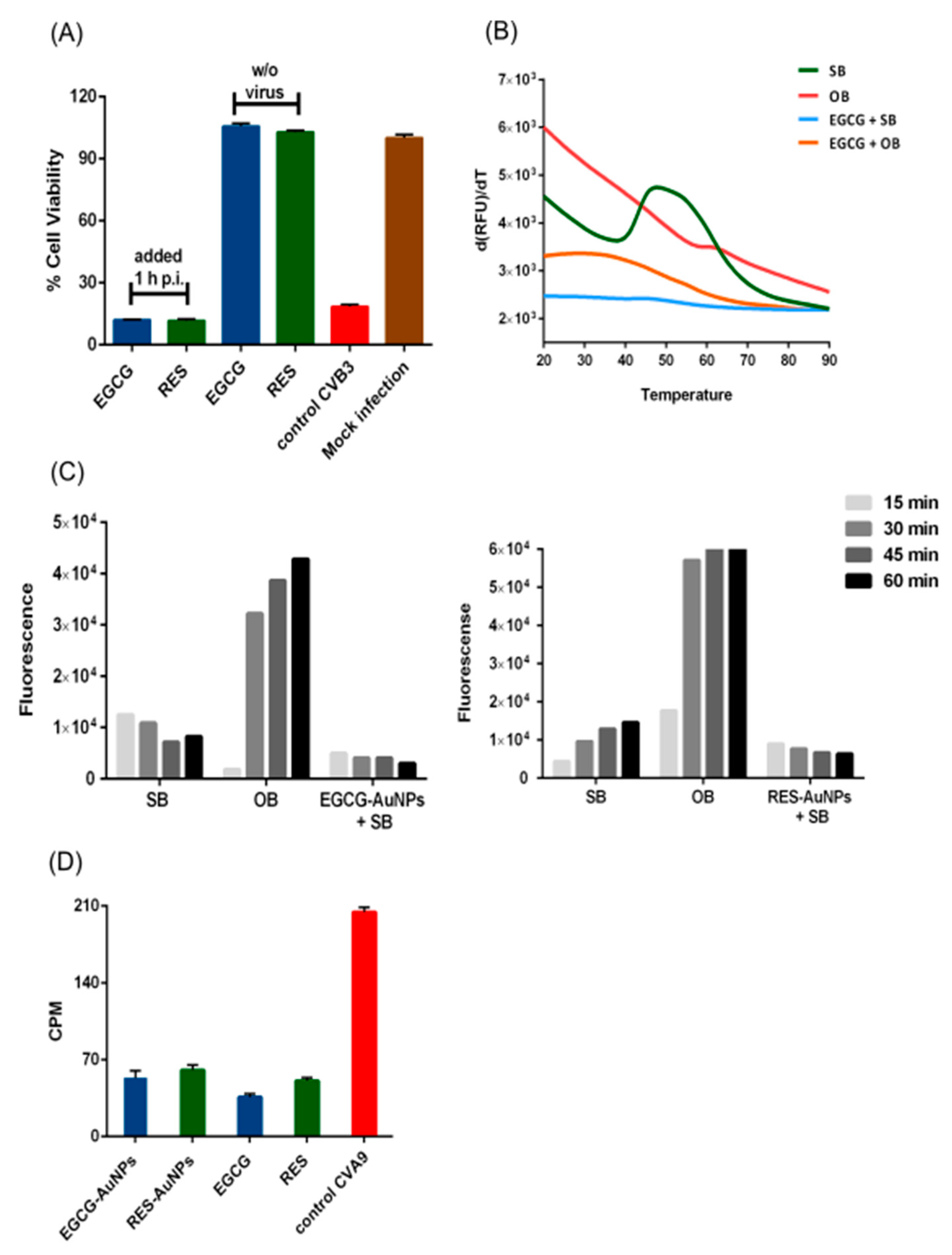
2O was added to the reaction mix, and a final volume of 50 μL was aliquoted into the thin-walled PCR plates. The thermal cycler (BioRad C100, Helsinki, Finland) recorded the fluorescence in quadruple from 20 to 90 °C with 0.5 °C intervals. The fluorescence data output was extracted from the BioRad CFX manager (2.1 software, accessed on 10 October 2020) and processed in Microsoft Excel (2016). The melt curve was obtained by plotting the relative fluorescence units (RFU) values versus temperature. The melting temperature (Tm) was determined from the melt peak, which was plotted using the derivative of the RFU as a function of temperature (d(RFU)/dT).
2.13. Transmission Electron Microscopy
A reaction mixture containing the CVA9 (1.6 × 1010 PFU/mL), polyphenols (EGCG, 650 μM; RES, 880 μM), polyphenol-functionalized nanoparticles EGCG-AuNPs (Au concentration 164 μg/mL, ligand concentration 5.61 × 10−1 μM) and RES-AuNPs (Au concentration 530 μg/mL, ligand concentration 3.44 μM) in buffer (PBS-MgCl2) was incubated for 1 h at 37 °C and 5% CO2. During the incubation, the Formvar-coated copper grids were glow-discharged using the EMS/SC7620 Mini sputter coater (Electron Microscopy Sciences, Hatfield, PA, USA). Post-incubation, 5 μL of the reaction mixture was pipetted on the glow-discharged grids for 20 s and then blotted away using Whatman paper. To negatively stain the sample, 5 μL of 1% (w/v) Phosphotungstic acid (PTA in water, pH adjusted to neutrality) (Sigma-Aldrich, Shinagawa-ku, Japan) or 2% (w/v) Uranyl acetate (Electron Microscopy Sciences, Hatfield, PA, USA) was pipetted on grids and incubated for 30 s, and blotted away using Whatman paper. The samples were visualized in a JEOL JEM-1400 transmission electron microscope (JEOl, Tokyo, Japan). The diameter of functionalized gold nanoparticles from the images taken using the TEM was measured by automated counting using the BioImageXD software (www.bioimagexd.net, accessed on 16 January 2021).
2.14. Radioactive Binding Assay
A549 cells were cultured for 24 h in 96-well flat-bottomed microtiter plates at a density of 12,000 cells/well in 100 μL of DMEM supplemented with 10% FBS, 1% GlutaMAX and 1% penicillin/streptomycin antibiotics. On day 2, 35SMet/Cys-CVA9 (28,000 CPM corresponding to 1.38 × 108 PFU/mL) was pre-treated with the polyphenols and polyphenol-functionalized nanoparticles EGCG (65 μM), RES (880 μM), EGCG-AuNPs (Au concentration 16.4 μg/mL, ligand concentration 5.61 × 10−2 μM) and RES-AuNPs (Au concentration 53 μg/mL, ligand concentration 3.44 × 10−1 μM) in buffer (PBS-MgCl2), and incubated at 37 °C for 1 h. Virus control without the compound was also prepared and incubated. Post-incubation, the sample tubes were placed on ice and 1% DMEM was added to make the final volume reach 100 μL. Following this, the samples with DMEM were added to the cells. Then, the 96-well plate was kept on ice while rocking for 1 h to ensure uniform binding of the virus on the cell surface. Next, the media was aspirated from each well and three washes with ice-cold PBS were performed to remove any unbound virus from the cell surface. To detach the cells, 150 μL of 1% Triton was added to the wells and incubated for 30 min. The entire liquid was transferred to a tube containing 4 mL of scintillation cocktail UltimaGoldTM and measured using the Tri-Carb® 2910TR liquid scintillation analyzer.
2.15. Dilution and Temperature Assays
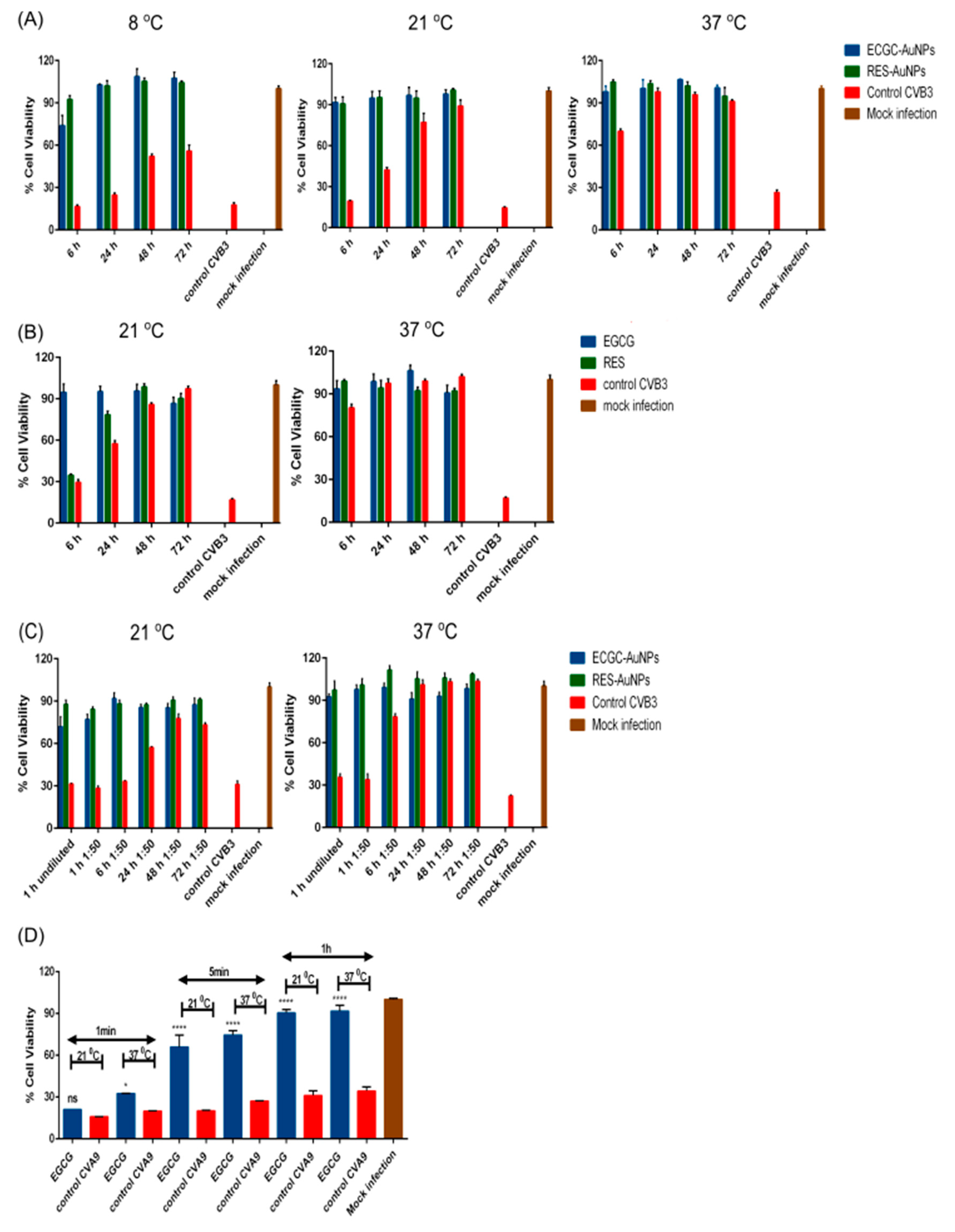
Dilution and temperature assays were performed using the CPE inhibition assay. For the dilution assay, CVB3 (4.4 × 108 PFU/mL), polyphenol-functionalized nanoparticles EGCG-AuNPs (Au concentration 164 μg/mL, ligand concentration 5.61 × 10−1 μM) and RES-AuNPs (Au concentration 530 μg/mL, ligand concentration 3.44 μM) in buffer (PBS-MgCl2) were incubated at 37 °C for 1 h. An experimental virus control (without the compounds) and fresh virus control (without the compounds and without any incubation) were used as two controls for the assay. Post-incubation, each of the test samples and the experimental virus control were divided equally into two tubes, one was diluted 50 times using a buffer (PBS-MgCl2) and the other part was stored at −20 °C (as an undiluted sample). The diluted samples were incubated over 72 h at 37 °C, such that equal fractions from them were collected at regular time-intervals (1, 6, 24, 48 and 72 h) and stored at −20 °C. At the end of 72 h, all the diluted and undiluted samples were further diluted with DMEM and added onto the cells, to obtain a final MOI of 15. On the subsequent day, the plates were stained for CPE as explained before.
When testing the effect of temperatures, two approaches were used. One with longer and the other with shorter times of incubation. For testing the longer periods of incubation, CVB3 (4.4 × 108 PFU/mL) was pre-treated with EGCG (650 μM), RES (880 μM), EGCG-AuNPs (Au concentration 164 μg/mL, ligand concentration 5.61 × 10−1 μM) and RES-AuNPs (Au concentration 530 μg/mL, ligand concentration 3.44 μM) in buffer (PBS-MgCl2) at RT (21 °C), 8 and 37 °C for 72 h. An experimental virus control (without the compounds) and fresh virus control (without the compounds and without any incubation) were used as two controls for the assay. Test samples and the experimental virus control were collected after incubating at the above-mentioned temperatures for 6, 24, 48 and 72 h and stored at −20 °C. After 72 h, all the samples were further diluted with DMEM and added to the cells, to obtain a final MOI of 15. On the subsequent day, the plates were stained for CPE, as explained before. In case of shorter incubation times, CVA9 (2 × 107 PFU/mL) was pre-treated with EGCG (6.5 μM) at 21 or 37 °C for 1 min, 5 min and 1 h and then added to the cells.
2.16. Blind Docking of Polyphenolic Compounds against CVA9 and CVB3
2.16.1. The Experimental Design
For the in silico docking experiment, the polyphenols Epigallocatechin gallate (EGCG) and Resveratrol (RES) were to be blindly docked over large external surfaces of the viral capsids of CVA9 and CVB3. Due to the overly large surfaces to be scanned, the capsids were segmented into relatively large grid boxes centered around their fold axes (2, 3 and 5) of rotational symmetry. This strategy was used in order to obtain overlapping segments spanning as much of the viral capsid surfaces as possible.
2.16.2. Ligand Preparation
The molecular structures of the polyphenolic compounds, Epigallocatechin gallate (Drug Bank ID: DB03823) and Resveratrol (Drug Bank ID: DB02709), were reconstituted and protonated from their SMILE strings using the RDKit library. The MGLTools software [
23] was then used to assign Gasteiger partial charges and merge non-polar hydrogen atoms.
2.16.3. Capsid Preparation
Full capsids for the CVA9 Griggs strain (PDB ID: 1D4M) [
24] and CVB3 Nancy strain (PDB ID: 6GZV) [
25] were reconstituted from their first biological assemblies using the PyMOL software. While the CVA9 capsid was already protonated, the PDB2PQR tool was used to protonate the CVB3 capsid to pH 7. However, as this altered the chain labels and segment identifiers, an in-house Python script (capsidockprep.py) was designed to restore these labels. In order to screen the maximum outer surface area from the capsid, the grid box center was placed at each individual capsid fold axis of rotational symmetry, with the box y-z plane being perpendicular to the axis of rotation and partially embedded within the capsid. In order to do so, centroids (x, y and z coordinates measured in angstroms) for each of the fold axes were selected, before shifting the box along the
x-axis by 10 Å to the right. Internally, before shifting, the capsid centroid is centered at the origin (0, 0, 0) prior to rotating the capsid centroid position vector to be along the
x-axis. For simplicity, the PDB2PQR tool is wrapped within an in-house Python script to enable protonation when the “ph” parameter is set. Similar to AutoDock Vina, a grid interval size of 0.375 Å is used. The “gridsize” parameter is specified in grid points. The grid parameters for all of the fold axes are provided in
Table 1. The AutoDock Vina plugin from PyMOL was used to visualize the effects of shifting and grid sizing.
The preprocessing by the in-house Python script greatly reduced the number of atoms by including protein chains belonging to any atom within the grid box, whilst keeping chain labels and segment IDs. Due to the high number of atoms required to capture the complete protomers in their protonated form for the 5-fold axis (beyond the limit of the PDB format), the experiment was focused around the 2- and 3-fold axes of rotation symmetry. As performed for the ligands, charges were assigned, and non-polar hydrogen atoms were merged using MGLTools.
2.16.4. Capsid Ligand Docking
QuickVina-W [
26] was used for docking the polyphenolic compounds, with a grid center of (0, 0, 0) and grid dimensions as defined in
Table 1. The runs were performed at an exhaustiveness of 1000 with 24 cores per job, using GNU Parallel [
27] at the Centre for High-Performance Computing (CHPC). A maximum of 20 top ligand binding poses were retained in each case run.
2.16.5. Analysis of Docking Results
The Arpeggio tool [
28] was then used to estimate the protein–ligand interactions around a subset of the complex trimmed at a maximum radius of 14 Å around the ligand atoms. An in-house Python script (without any external library dependency) was used to extract this subset, retaining the chain label and segment ID. Hydrogen bonding, hydrophobic interactions and the binding energy scores were then used to characterize ligand binding. The distributions of these measurements across the entire docking experiment were arranged according to the strains and the detected binding sites. In order to differentiate the binding sites from existing ones, both the canonical hydrophobic pocket [
24] and the VP1-VP3 druggable interprotomer pocket (the Butcher Neyts’ pocket) from Abdelnabi and co-workers [
25] were carefully mapped on all 3D structures, as their compositions and locations were not identical in each strain. In the case of the hydrophobic pocket, the PyVOL plugin [
29] in PyMOL was used, specifying VP1 as the search space and a minimal pocket volume of 400 Å, in addition to the default parameters. The minimum volume had been determined beforehand using the palmitic acid-contacting residues (any protein atom within 4 Å of each ligand atom) from a 3D structure of the CVB3 Woodruff strain (PDB ID: 1COV) [
30] to guide the cavity search. In the case of the interprotomer pocket, the PROMALS3D web server [
31] was used to align the VP1 sequences and correct for any misaligned residue positions using 3D structural information. The ligand-interacting residues (binding the benzene sulfonamide derivative) from CVB3 (6GZV) identified by PDB were used as a reference in a sequence alignment with the CVA9 (1D4M) strain to obtain homologous positions for the latter [
32]. No correction was needed for VP3 residues of the interprotomer surface, as the alignment contained no gaps. Unless specified otherwise, residues have been numbered according to the respective crystal structures.
2.17. Statistical Analysis
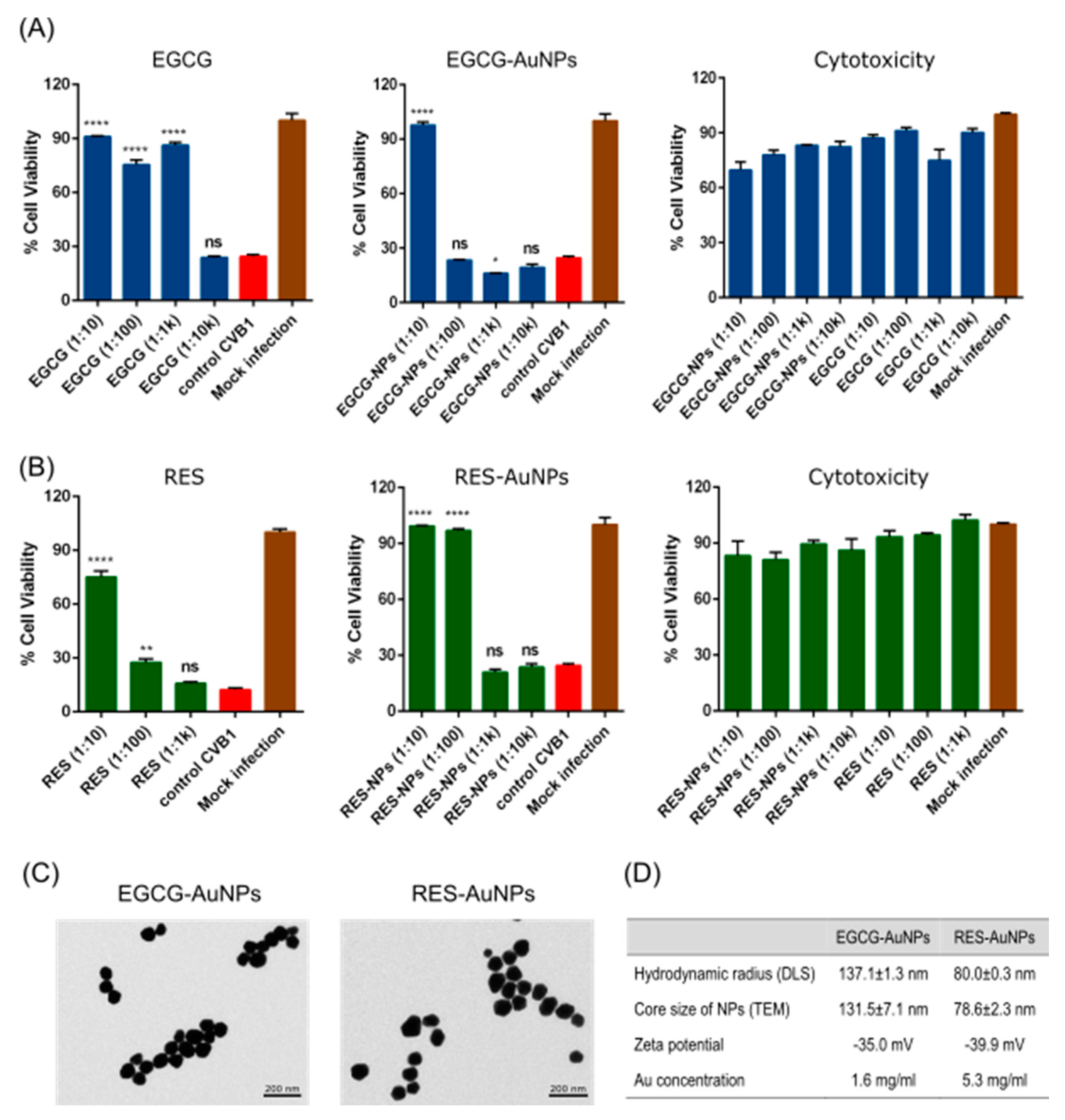
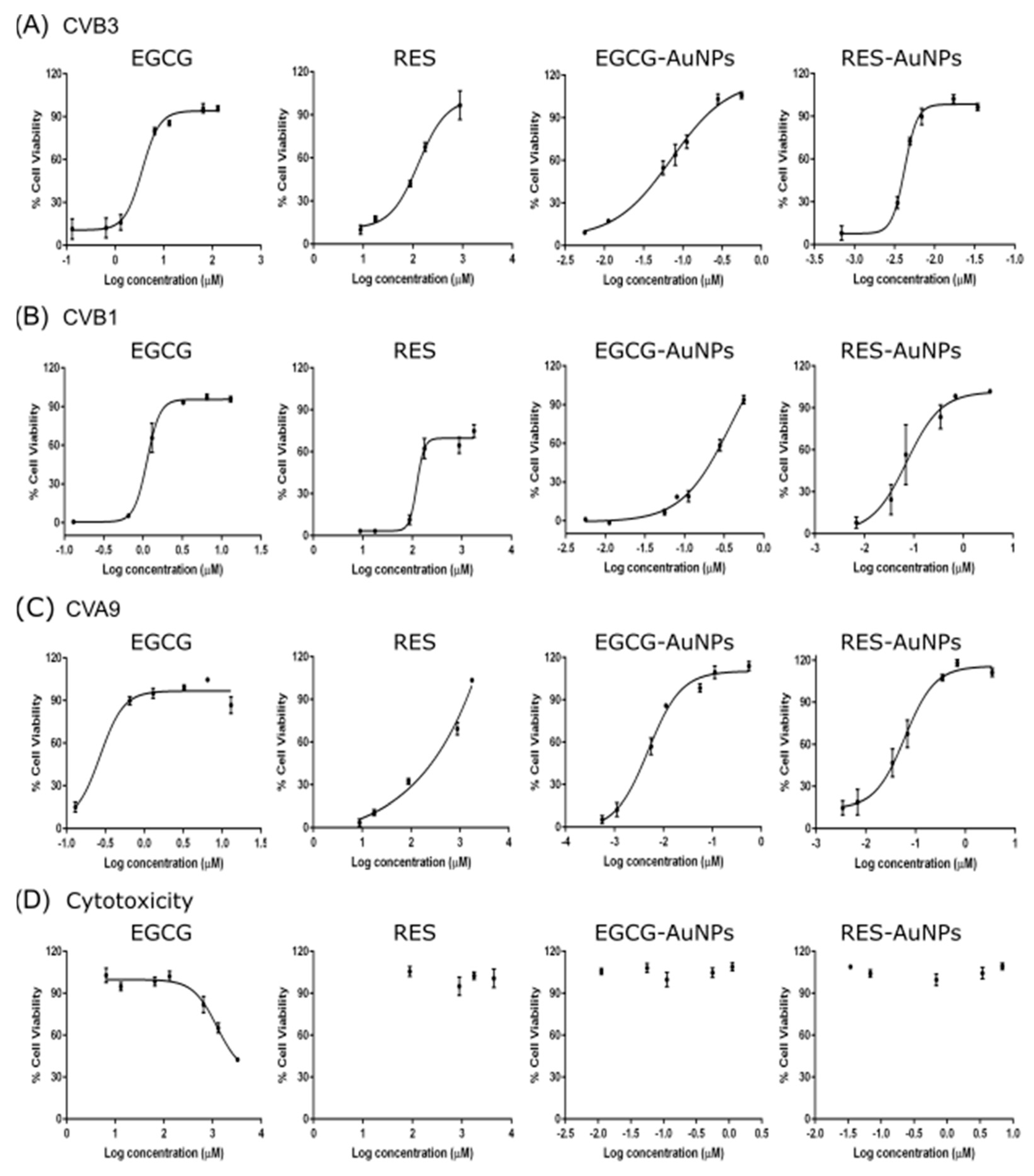
Statistical analysis was performed using GraphPad Prism 6 (GraphPad Software, San Diego, CA, USA). Data are presented as mean ± standard error (SEM). The 50% effective concentrations (EC50) and 50% cytotoxic concentrations (CC50) were calculated by regression analysis of the dose–response curves generated from the experimental data using the software. Statistical significance was calculated by performing one-way/two-way ANOVA followed by the Bonferroni test (* p < 0.05, ** p < 0.01, *** p < 0.001 and **** p < 0.0001).
4. Discussion
There is a great need for broad-acting non-toxic antivirals that reduce the virus load in the body or on surfaces. Present and future pandemics as well as the yearly occurring epidemics cause high economic costs due to hospitalizations and absence from workplaces and from schools. Enteroviruses still lack a clinically accepted antiviral for treatment or disinfection of hands and surfaces. Therefore, we studied two polyphenols, EGCG from green tea and RES from red grapes and other fruits, as soluble polyphenols and polyphenol-functionalized gold nanoparticles and explored the mechanism of action for the molecules on three very stable, non-enveloped enteroviruses (CVA9, CVB1 and CVB3).
Our results showed that EGCG and RES have strong antiviral efficacy against the three enterovirus serotypes. Antiviral efficacy in general was already known for the two polyphenols before [
34], but not for enteroviruses. The EC
50 values were in the range of 0.266–3.411 µM for EGCG and 107.894–330.369 µM for RES, thus suggesting a good efficacy already at low concentrations. The CC
50 showed no cytotoxicity. The polyphenols bound to the AuNPs showed EC
50 values in the range of 0.004–0.237 µM for EGCG and 0.004–0.068 µM for RES.
Most antivirals target cellular replication steps of the viruses, such as promoting apoptosis and via inhibiting Nf-kB or MAPK pathways and replication [
5,
35,
36]. Most studies on the polyphenols as antivirals for enteroviruses focus on EV71 and observe antiviral action past entry and during replication of the viruses. Our results reveal that both EGCG and Resveratrol act as entry and attachment inhibitors by binding to enterovirus surfaces. This was confirmed by the time of addition CPE assay when polyphenols pre-incubated with viruses showed inhibition, while drugs added after entry to cells had no effect. In silico docking data further showed that these polyphenols could bind to multiple areas on the virion surface. Other drugs binding directly to enterovirus capsids, such as pleconaril, target the hydrophobic pocket that is normally housed by an aliphatic lipid such as palmitic or oleic acid [
37,
38]. The hydrophobic pocket has been associated with capsid stability and suggested to release its lipid upon viral RNA release [
39]. We have recently shown that albumin treatment of enteroviruses Echovirus 1 and CVA9 do expand the virion, which is typically an event associated with emptying of the pocket. However, we showed that treatment with albumin, which may steal fatty acids from the virion, does not yet lead to virus uncoating but rather virus priming to an intermediary form, leading to virus RNA release only upon further cues, such as changes in concentration of key ions [
16,
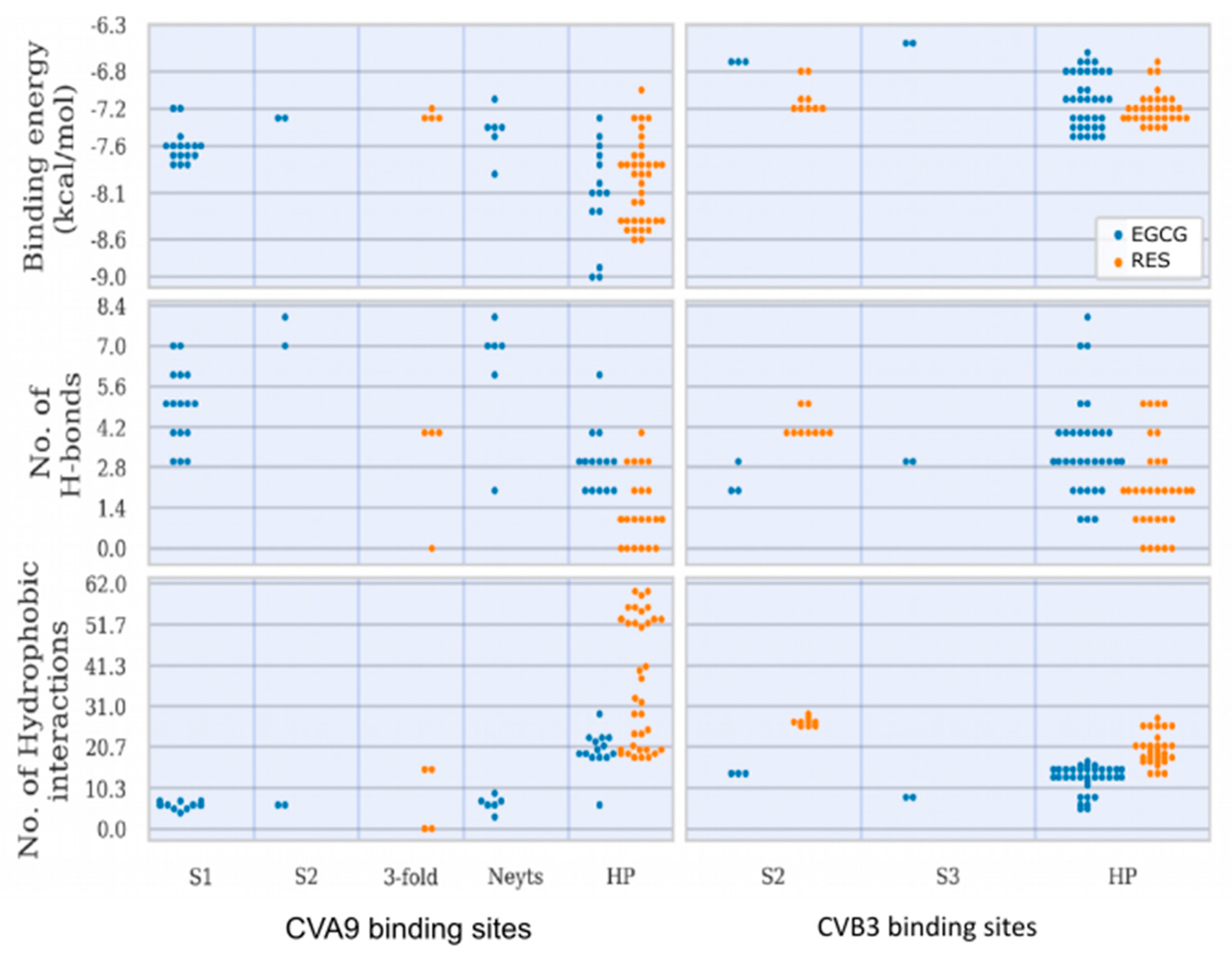
21]. Here, we have studied three viruses, two of which show similarly spacious pockets, whereas the CVB3 Nancy strain has a collapsed pocket that cannot house large molecules. Consequently, CVA9 showed the highest estimated binding energies for both drugs in the HP. In addition, simulations showed that the HP is the preferable docking site for the drugs with the 73% of the docking poses associated with it.
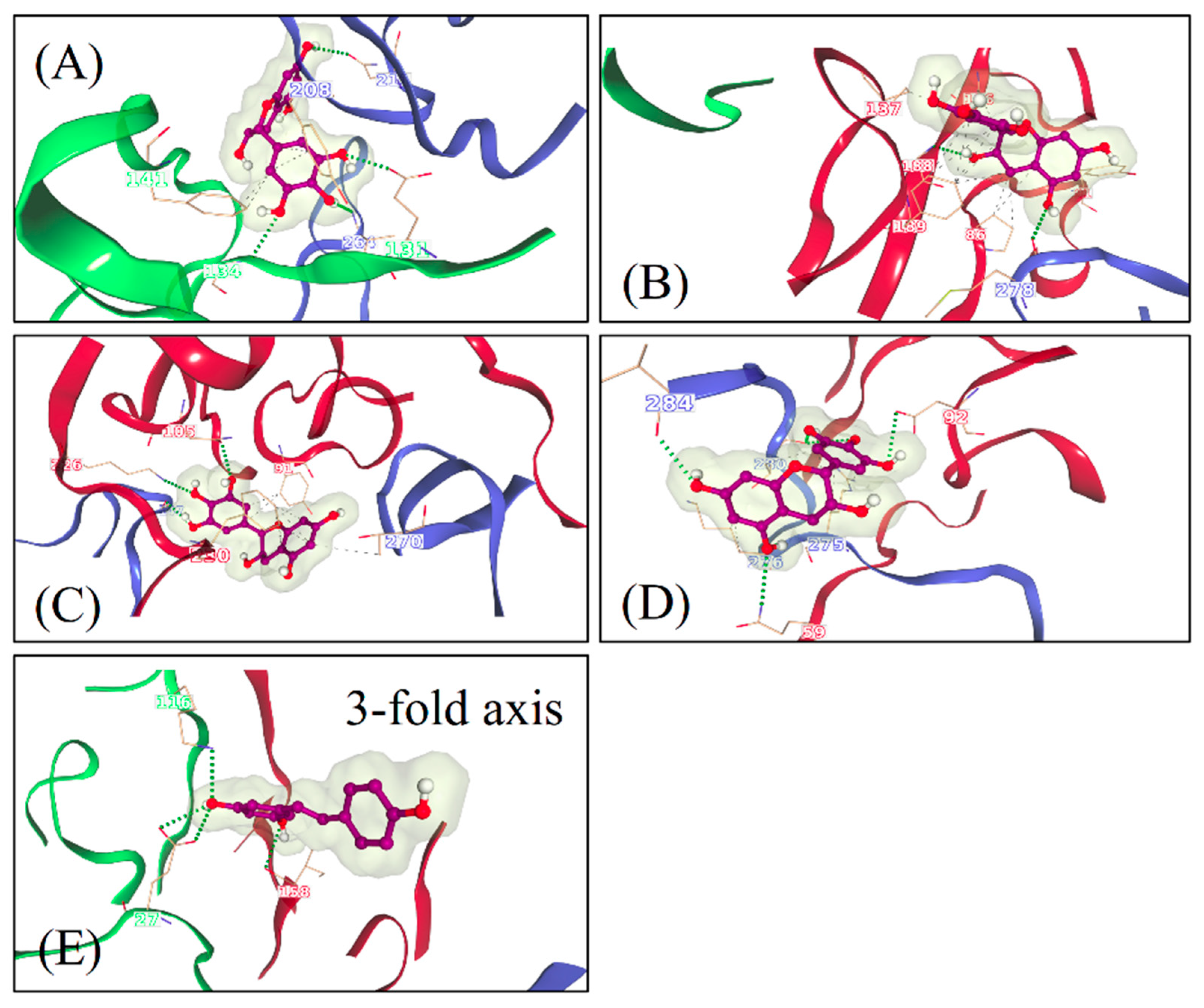
A novel algorithm that was developed for this study to reveal the docking of the molecules on enterovirus surfaces considered larger areas around the 2- and 3-fold symmetry axis. Remarkably, the docking poses for the polyphenols, RES and EGCG on the viral capsids suggested the existence of multiple binding sites, as seen from the 2-fold (
Figure 6) and 3-fold (
Figure 7) axes of symmetry, respectively. It seemed that the binding locations were influenced by both the physico-chemical characteristics of the polyphenols, the geometric landscapes and partial charge distributions present within the capsid assemblies for each strain. While both compounds are polyphenolic, Resveratrol is classified by PubChem as smaller and more hydrophobic (2 heterocycles, an XLogP3 value of 3.1 and 3 H-bond acceptors) than EGCG (4 heterocycles, an XLogP3 value of 1.2 and 11 H-bond acceptors) [
40], which we believe to be a main factor influencing the distribution of the ligand binding sites. The fact that the HPs detected in CVA9 are much deeper than those of CVB3 may explain the better fit of the smaller Resveratrol further inside the capsid compared to EGCG. Altogether, three new sites were found, in addition to the known HP and the recently discovered pocket that we call the Butcher-Neyts pocket [
25].
Our results suggest that there are both specific and shared binding sites for EGCG and RES, with the strain also playing a role. HP showed good binding by both viruses and both drugs, suggesting that it is the main attraction on the virion surface. Instead, S1 was only detected with EGCG in CVA9, while EGCG bound S2 in both CVA9 and CVB3. Resveratrol bound to S2 only in CVB3 but to the 3-fold axis only in CVA9, further suggesting that the structural differences of the drugs as well as of the virions dictate the docking to the virions.
The predicted binding energies were lower in the newly found docking sites in comparison to the canonical HP, probably due to its deep invagination and charges present in the pocket. In silico-determined binding energies only provide a general estimation of the affinity of ligand binding and are generally useful for doing preliminary ranking of docked ligands. However, the frequency of binding to a site can additionally assist in the evaluation of lower-scoring poses. Out of the 159 docking poses, the HP ranked first with 73% of poses assigned to it, followed by S1 (10%), S2 (9%), the Butcher-Neyts pocket (4%), the pore at the 3-fold axis (3%) and the S3 site (1%). As the Neyts pocket has already been independently proven experimentally, there are valid reasons to believe that S1, S2 and the 3-fold pore are actual novel binding sites, based on the closeness in percentage clustering, a partial overlap in the range of binding energies and the number of H-bonding interactions. There is less support for the S3 site and S4, even though both may occur.
It is no wonder that the multiple ligand binding sites on the capsid led to a strong antiviral effect directly on the virion. Our studies on the molecular mechanisms of the action showed that stabilization rather than premature RNA release occurred for enteroviruses. This was shown by several methods: (1) thermal assay, (2) our previously developed real-time uncoating assay, (3) radioactive gradient assay and (4) TEM. Stabilization and prevention of virus uncoating was evident even at higher temperatures. Our results also showed that ligand binding was accompanied by strong clustering of the virions in addition to prevention of virion uncoating in the clusters.
The fact that EGCG is a bigger molecule with more hydroxy groups can be the reason why, in comparison to RES, it is more efficient in inhibiting the enteroviruses. The number of hydroxy groups has been addressed before for the difference in antiviral action of the polyphenols, apigenin and luteolin [
36]. Interestingly, Resveratrol being a nearly flat molecule (
Supplementary Figure S3) is more closely attached to the surface of a gold nanoparticle. Previous experiments indicate that aromatic rings are oriented parallel to the gold surface [
17]. EGCG has a nearly planar aromatic ring system with one ring perpendicular to the base (
Supplementary Figure S3).
This indicates that partial immersion of the EGCG is possible and can be responsible for the increase in stability and the subsequent virus aggregation. On the other hand, the three-dimensional structure of the free EGCG molecule can cross-link several viruses by immersing into their binding pockets and inducing cross-linking of two viruses, explaining why the free EGCG is more efficient than the RES molecule. RES is six times more concentrated when bound to AuNPs as compared to EGCG. Moreover, the molecules are presented on smaller nanoparticles that have a strong surface curvature. Even if we assume that RES only penetrates slightly into the pockets because it is nearly parallel to the surface, the sheer number of interactions between virus and RES and the RES presentation by the more curved surface of nanoparticles can work as a cross-linker between multiple binding pockets on several viruses. This could explain why RES bound to nanoparticles is more efficient than the free molecule.
Another interesting fact that we observed in our infection experiments was that RES was slower to inhibit virus infection. After 6 h, its antiviral efficacy was still moderate, whereas after 24 h and later, it had reached full efficiency, while EGCG was very efficient already at earlier time points. This may be directly because RES has fewer docking sites on the virion surface. However, it may also be because it has less hydroxylic groups on its structure, leading to lower binding affinity.
Altogether, the two studied polyphenols showed great antiviral efficacy for all three studied enteroviruses. The discovered mechanism of action is that the polyphenols immerse in several binding sites on the virion surface, resulting in stabilization of the virions against opening, even in harsh conditions. In addition, formation of large clusters was observed, which might be another efficient way to lower the incidence of virions entering cells. Clustering and the resulting precipitation of the virus–polyphenol or polyphenol–AuNP clusters indicate a potential use for safe water clearing applications and for decontamination of surfaces. Future studies will focus on clarifying if the clustered viruses will be prevented from penetrating the mucus layer or be used as vaccines. Previous studies have also demonstrated the therapeutic potential of polyphenols such as EGCG and Curcumin, for treating protein aggregation diseases associated with peripheral neuropathy [
41]. The mechanism by which EGCG works, to prevent the disease progression in case of amyloid fibril formation, is by stabilizing the protein after binding to it and forming soluble nontoxic aggregates [
42,
43]. EGCG interacts with amyloid-forming proteins through hydrophobic interactions or H-bonds, forming stable oligomers [
44]. Similar interactions between the virus capsid and polyphenols have been described in our computational studies. Whether the mechanism behind virus stability/loss of infectivity and prevention of formation of amyloid aggregates reported previously is the same remains to be shown.
In general, it can be concluded that especially the AuNP-bound polyphenols might have interesting potential in future applications for (i) antiviral surface coatings, which can already reduce the viral load before it is transferred to the body surface and unintentionally up-taken, and (ii) as oral antivirals. The oral administration of the polyphenol-coated nanoparticles is especially interesting as the nanoparticle and the bound polyphenol are localized in the primary infection site for enteroviruses, in the gut, before they likely pass through the intestinal cell layer and hence can significantly reduce the viral load of the intestine. The systemic toxicity of these nanoparticles will be determined in future animal experiments. It requires consideration as the polyphenol-AuNPs due to their size and nearly neutral surface charge have optimal properties to penetrate the mucus layer of the intestinal tract [
45,
46].

 ,
, 













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}