
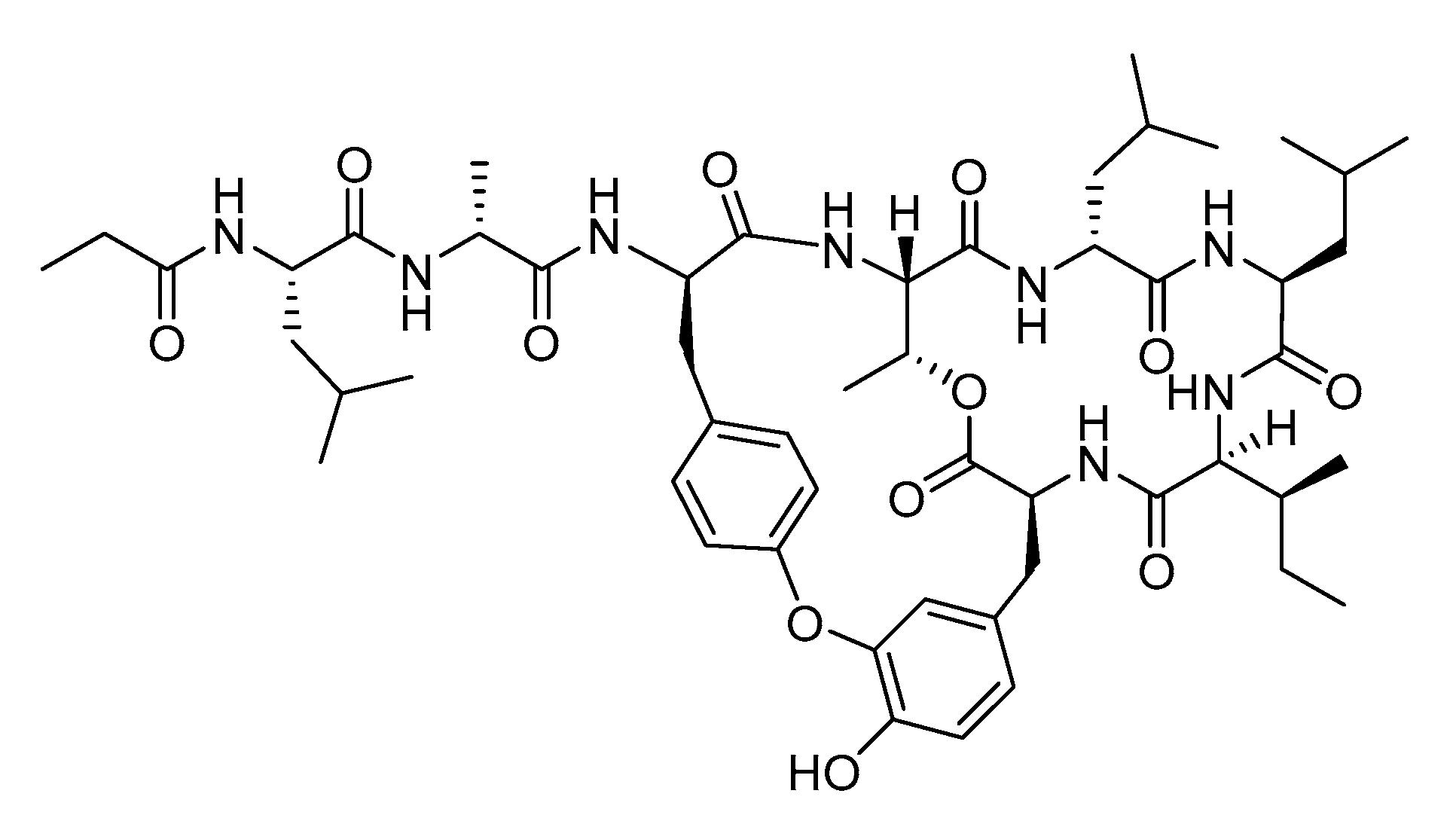
In Vitro Metabolism Study of Seongsanamide A in Human Liver Microsomes Using Non-Targeted Metabolomics and Feature-Based Molecular Networking

 ,
,  and
and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Reagents
2.2. Incubation of Seongsanamide A in HLMs
2.3. Liquid Chromatography-High Resolution Mass Spectrometric Analysis
2.4. Metabolite Identification Using a Metabolomics Approach
2.5. Metabolite Identification Using a FBMN
2.6. Reaction Phenotyping Study of Seongsanamide A Using Recombinant Cytochrome P450 Isoforms
2.7. Inhibitory Potency of Seongsanamide A against Human Cytochrome P450 Activity
3. Results and Discussion
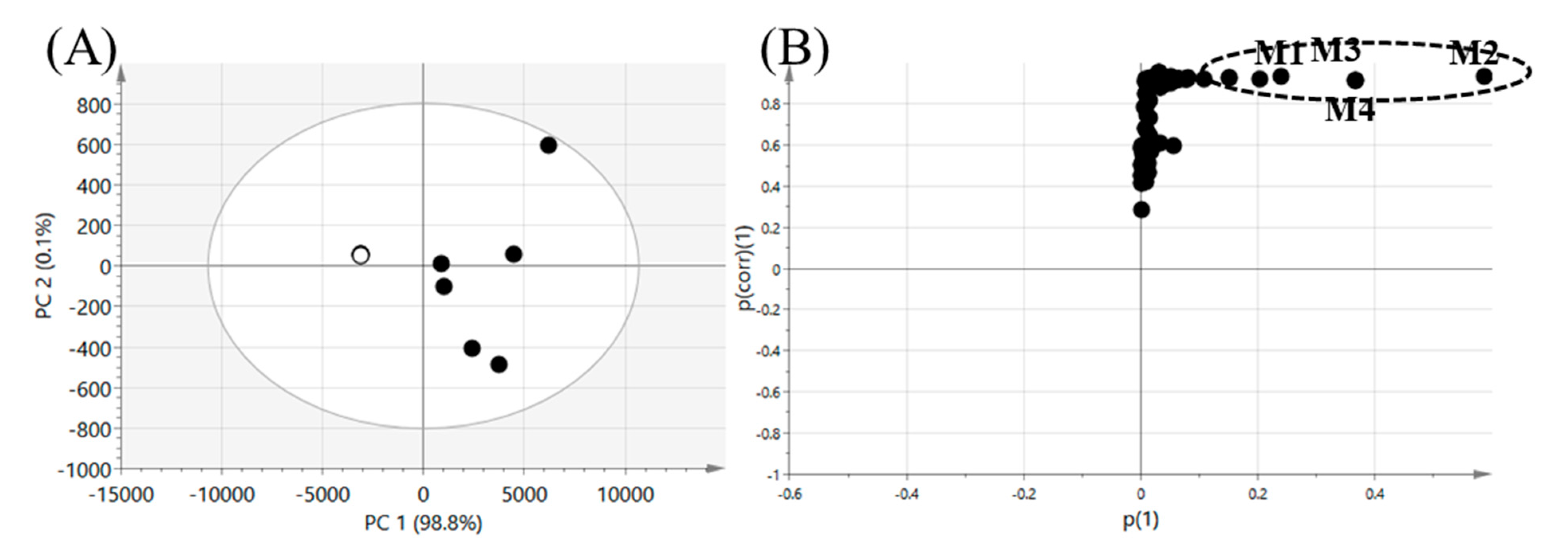
3.1. Profiling of Seongsanamide A Metabolites Using a Metabolomic Approach
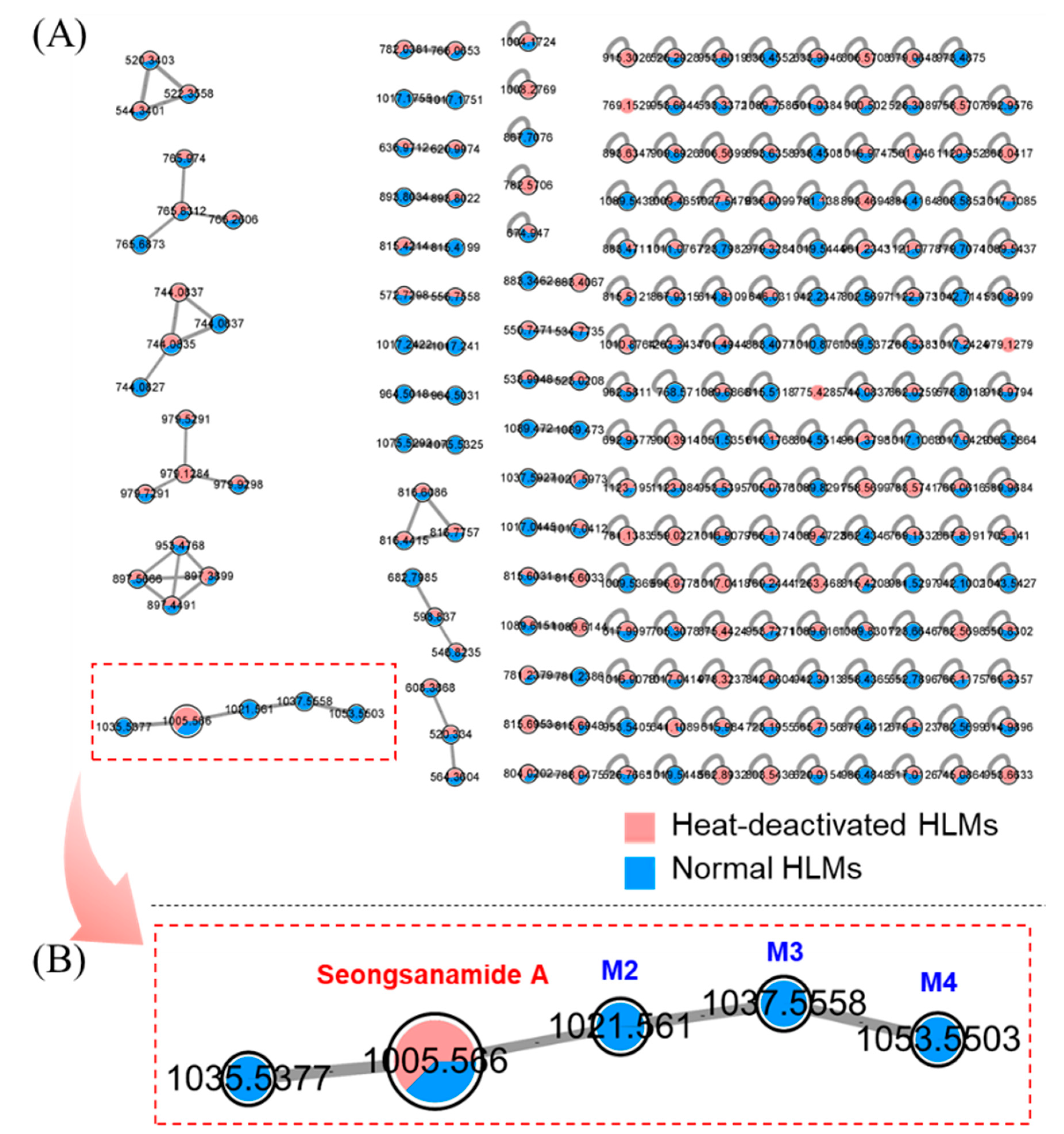
3.2. Profiling of Seongsanamide A Metabolites Using an FBMN
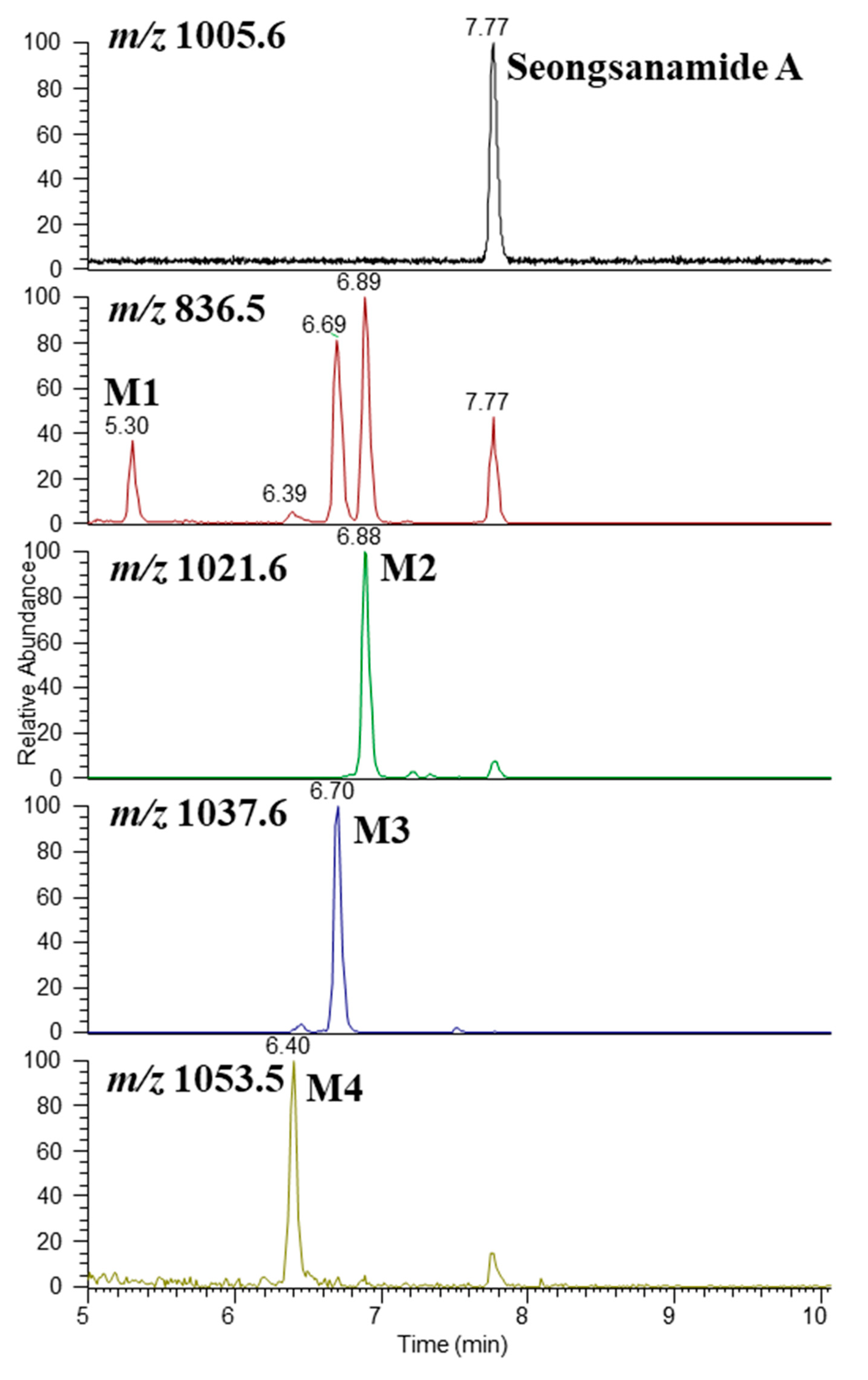
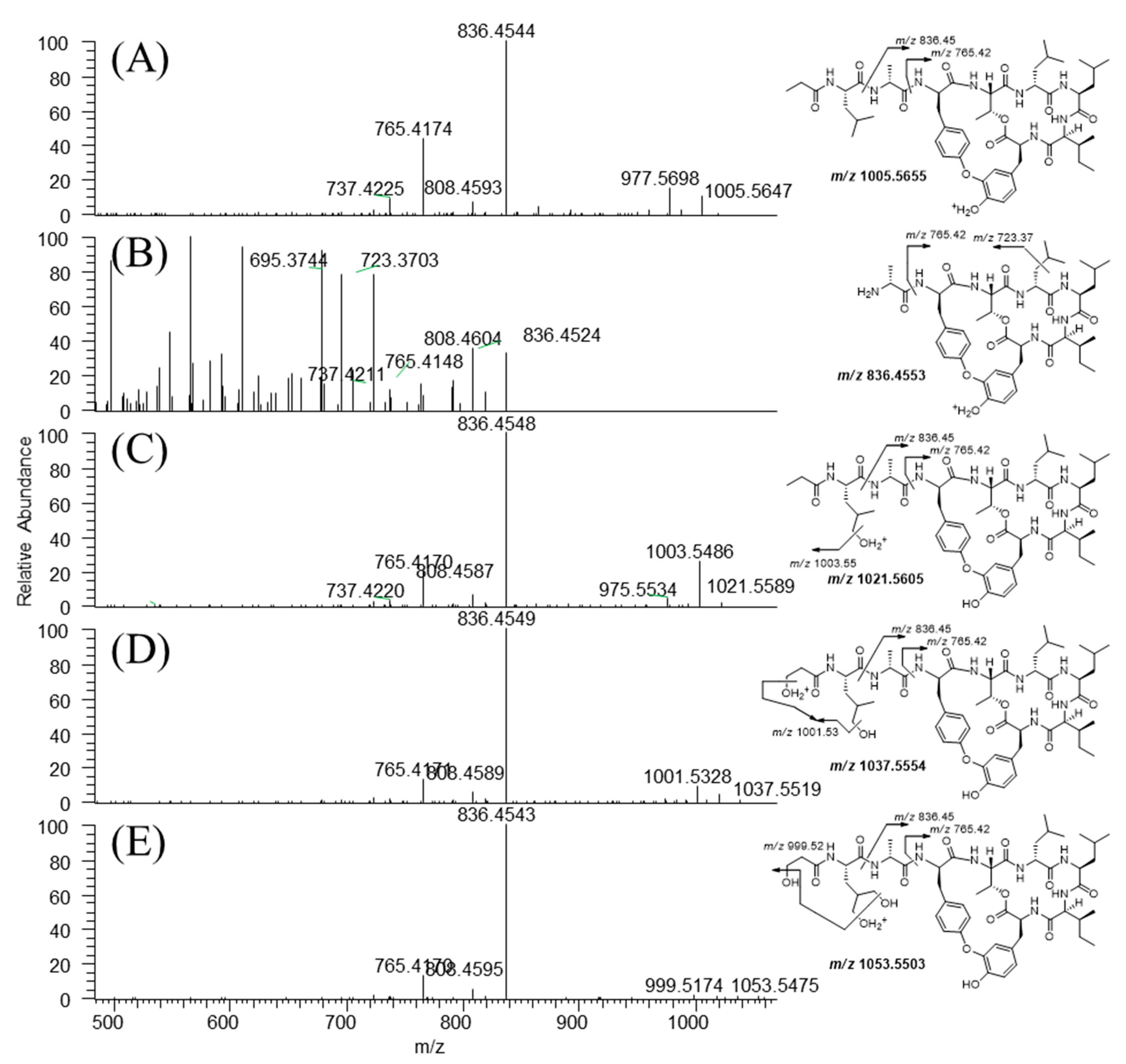
3.3. Metabolite Structure Identification
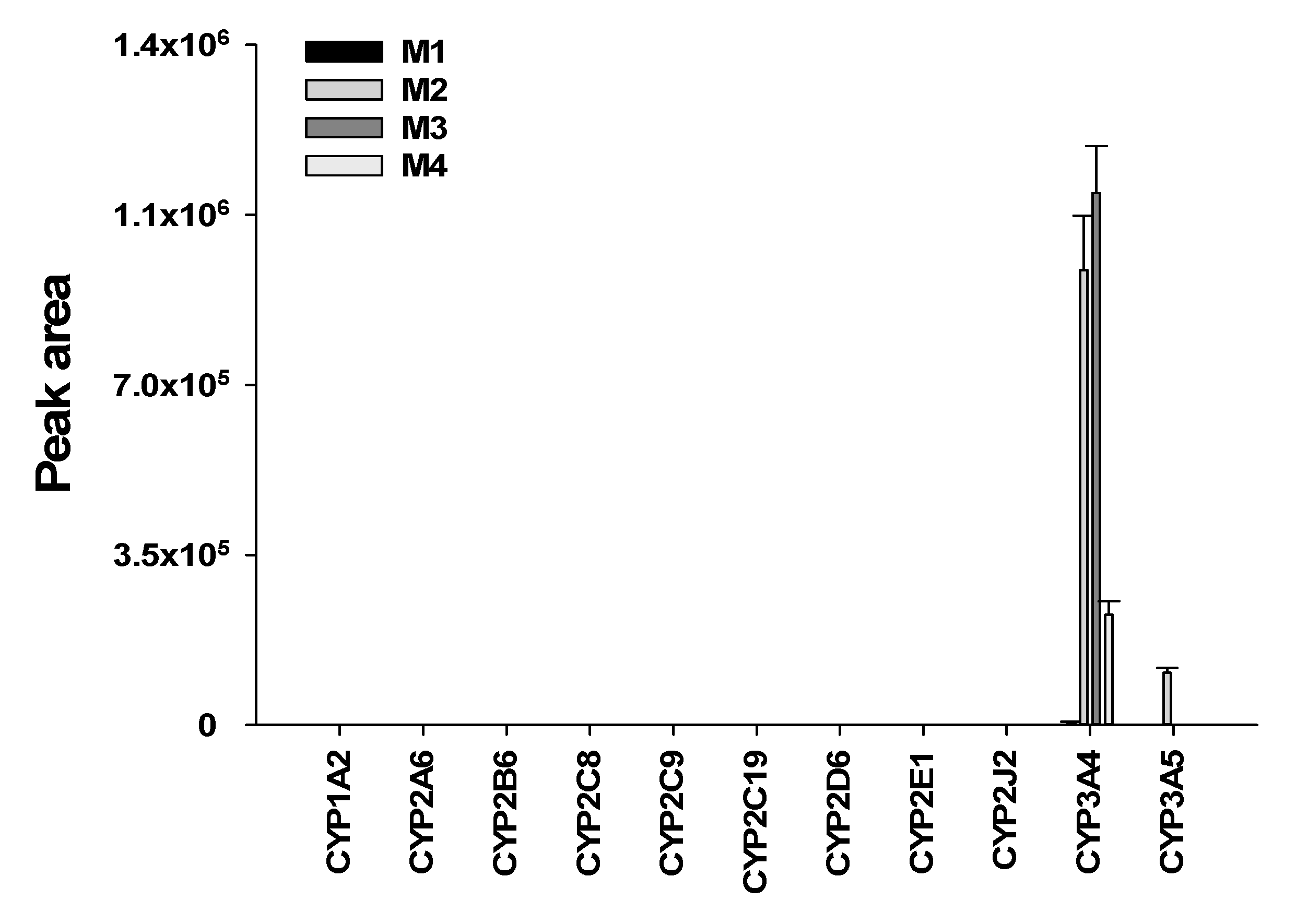
3.4. Characterization of Human Cytochrome P450 Enzymes Responsible for the Formation of Four Metabolites of Seongsanamide A
3.5. In Vitro Inhibition of Cytochrome P450 Enzymes by Seongsanamide A
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Rhodes, C.; Pei, D. Bicyclic Peptides as Next-Generation Therapeutics. Chemistry 2017, 23, 12690–12703. [Google Scholar] [CrossRef]
- Zorzi, A.; Deyle, K.; Heinis, C. Cyclic peptide therapeutics: Past, present and future. Curr. Opin. Chem. Biol. 2017, 38, 24–29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fukuhara, K.; Takada, K.; Watanabe, R.; Suzuki, T.; Okada, S.; Matsunaga, S. Colony-wise Analysis of a Theonella swinhoei Marine Sponge with a Yellow Interior Permitted the Isolation of Theonellamide I. J. Nat. Prod. 2018, 81, 2595–2599. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, S.; Arita, Y.; Honda, M.; Iwamoto, K.; Matsuyama, A.; Shirai, A.; Kawasaki, H.; Kakeya, H.; Kobayashi, T.; Matsunaga, S.; et al. Marine antifungal theonellamides target 3beta-hydroxysterol to activate Rho1 signaling. Nat. Chem. Biol. 2010, 6, 519–526. [Google Scholar] [CrossRef] [PubMed]
- Narita, K.; Katoh, T. Total Synthesis of Thailandepsin B, a Potent HDAC Inhibitor Isolated from a Microorganism. Chem. Pharm. Bull. 2016, 64, 913–917. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieland, T.; Faulstich, H. Amatoxins, Phallotoxins, Phallolysin, and Antamanide: The Biologically Active Components of PoisonousAmanitaMushroom. CRC Crit. Rev. Biochem. 1978, 5, 185–260. [Google Scholar] [CrossRef]
- Kim, G.J.; Li, X.; Kim, S.H.; Yang, I.; Hahn, D.; Chin, J.; Nam, S.J.; Nam, J.W.; Nam, D.H.; Oh, D.C.; et al. Seongsanamides A-D: Antiallergic Bicyclic Peptides from Bacillus safensis KCTC 12796BP. Org. Lett. 2018, 20, 7539–7543. [Google Scholar] [CrossRef] [PubMed]
- Han, F.; Guo, Y.; Ye, T. Total synthesis of antiallergic bicyclic peptide seongsanamide A. Org. Chem. Front. 2020, 7, 1658–1662. [Google Scholar] [CrossRef]
- Eliopoulos, G.M.; Willey, S.; Reiszner, E.; Spitzer, P.G.; Caputo, G.; Moellering, R.C. In vitro and in vivo activity of LY 146032, a new cyclic lipopeptide antibiotic. Antimicrob. Agents Chemother. 1986, 30, 532–535. [Google Scholar] [CrossRef] [Green Version]
- D’Costa, V.M.; Mukhtar, T.A.; Patel, T.; Koteva, K.; Waglechner, N.; Hughes, D.W.; Wright, G.D.; De Pascale, G. Inactivation of the Lipopeptide Antibiotic Daptomycin by Hydrolytic Mechanisms. Antimicrob. Agents Chemother. 2011, 56, 757–764. [Google Scholar] [CrossRef] [Green Version]
- Amundsen, R.; Åsberg, A.; Ohm, I.K.; Christensen, H. Cyclosporine A- and Tacrolimus-Mediated Inhibition of CYP3A4 and CYP3A5 in Vitro. Drug Metab. Dispos. 2011, 40, 655–661. [Google Scholar] [CrossRef]
- Sharma, R.; McDonald, T.S.; Eng, H.; Limberakis, C.; Stevens, B.D.; Patel, S.; Kalgutkar, A.S. In vitro metabolism of the glucagon-like peptide-1 (GLP-1)-derived metabolites GLP-1(9-36)amide and GLP-1(28-36)amide in mouse and human hepatocytes. Drug Metab. Dispos. 2013, 41, 2148–2157. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.; Lu, Y.; Guan, X.; Dong, B.; Chavan, H.; Wang, J.; Zhang, Y.; Krishnamurthy, P.; Li, F. Metabolomics reveals the formation of aldehydes and iminium in gefitinib metabolism. Biochem. Pharmacol. 2015, 97, 111–121. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Choi, W.-G.; Moon, J.-Y.; Lee, J.Y.; Lee, S.; Lee, H.S. Metabolomics-assisted metabolite profiling of itraconazole in human liver preparations. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2018, 1083, 68–74. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-H.; Choi, W.-G.; Lee, S.; Lee, H.S. Revisiting the Metabolism and Bioactivation of Ketoconazole in Human and Mouse Using Liquid Chromatography–Mass Spectrometry-Based Metabolomics. Int. J. Mol. Sci. 2017, 18, 621. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Fang, Z.-Z.; Zheng, Y.; Zhou, K.; Hu, C.; Krausz, K.W.; Sun, D.; Idle, J.; Gonzalez, F.J. Metabolic profiling of praziquantel enantiomers. Biochem. Pharmacol. 2014, 90, 166–178. [Google Scholar] [CrossRef] [Green Version]
- Wang, M.; Carver, J.J.; Phelan, V.V.; Sanchez, L.M.; Garg, N.; Peng, Y.; Nguyen, D.D.; Watrous, J.; Kapono, C.A.; Luzzatto-Knaan, T.; et al. Sharing and community curation of mass spectrometry data with Global Natural Products Social Molecular Networking. Nat. Biotechnol. 2016, 34, 828–837. [Google Scholar] [CrossRef] [Green Version]
- Allard, P.-M.; Genta-Jouve, G.; Wolfender, J.-L. Deep metabolome annotation in natural products research: Towards a virtuous cycle in metabolite identification. Curr. Opin. Chem. Biol. 2017, 36, 40–49. [Google Scholar] [CrossRef]
- Nothias, L.F.; Petras, D.; Schmid, R.; Duhrkop, K.; Rainer, J.; Sarvepalli, A.; Protsyuk, I.; Ernst, M.; Tsugawa, H.; Fleischauer, M.; et al. Feature-based molecular networking in the GNPS analysis environment. Nat. Methods 2020, 17, 905–908. [Google Scholar] [CrossRef]
- Le Daré, B.; Ferron, P.-J.; Allard, P.-M.; Clément, B.; Morel, I.; Gicquel, T. New insights into quetiapine metabolism using molecular networking. Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Allard, S.; Allard, P.M.; Morel, I.; Gicquel, T. Application of a molecular networking approach for clinical and forensic toxicology exemplified in three cases involving 3-MeO-PCP, doxylamine, and chlormequat. Drug Test. Anal. 2019, 11, 669–677. [Google Scholar] [CrossRef]
- Chambers, M.C.; Maclean, B.; Burke, R.; Amodei, D.; Ruderman, D.L.; Neumann, S.; Gatto, L.; Fischer, B.; Pratt, B.; Egertson, J.; et al. A cross-platform toolkit for mass spectrometry and proteomics. Nat. Biotechnol. 2012, 30, 918–920. [Google Scholar] [CrossRef]
- Kang, K.B.; Gao, M.; Kim, G.J.; Choi, H.; Sung, S.H. Rhamnellosides A and B, omega-Phenylpentaene Fatty Acid Amide Diglycosides from the Fruits of Rhamnella franguloides. Molecules 2018, 23, 752. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pluskal, T.; Castillo, S.; Villar-Briones, A.; Orešič, M. MZmine 2: Modular framework for processing, visualizing, and analyzing mass spectrometry-based molecular profile data. BMC Bioinform. 2010, 11, 395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A Software Environment for Integrated Models of Biomolecular Interaction Networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef]
- Park, S.-Y.; Nguyen, P.-H.; Kim, G.; Jang, S.-N.; Lee, G.-H.; Phuc, N.M.; Wu, Z.; Liu, K.-H. Strong and Selective Inhibitory Effects of the Biflavonoid Selamariscina A against CYP2C8 and CYP2C9 Enzyme Activities in Human Liver Microsomes. Pharmaceutics 2020, 12, 343. [Google Scholar] [CrossRef]
- Kim, M.J.; Kim, H.; Cha, I.J.; Park, J.S.; Shon, J.H.; Liu, K.H.; Shin, J.G. High-throughput screening of inhibitory potential of nine cytochrome P450 enzymes in vitro using liquid chromatography/tandem mass spectrometry. Rapid. Commun. Mass Spectrom. 2005, 19, 2651–2658. [Google Scholar] [CrossRef] [PubMed]
- Joo, J.; Liu, K.-H. Inhibitory effect of honokiol and magnolol on cytochrome P450 enzyme activities in human liver microsomes. Mass Spectrom. Lett. 2013, 4, 34–37. [Google Scholar] [CrossRef] [Green Version]
- Xie, C.; Gao, X.; Sun, D.; Zhang, Y.; Krausz, K.W.; Qin, X.; Gonzalez, F.J. Metabolic Profiling of the Novel Hypoxia-Inducible Factor 2α Inhibitor PT2385 In Vivo and In Vitro. Drug Metab. Dispos. 2018, 46, 336–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shon, J.C.; Shin, H.-S.; Seo, Y.K.; Yoon, Y.-R.; Shin, H.; Liu, K.-H. Direct Infusion MS-Based Lipid Profiling Reveals the Pharmacological Effects of Compound K-Reinforced Ginsenosides in High-Fat Diet Induced Obese Mice. J. Agric. Food Chem. 2015, 63, 2919–2929. [Google Scholar] [CrossRef]
- Kim, S.E.; Ji, S.B.; Kim, E.; Jeong, M.; Kim, J.; Lee, G.M.; Seo, H.J.; Bae, S.; Jeong, Y.; Lee, S.; et al. Nontargeted Metabolomics by High-Resolution Mass Spectrometry to Study the In Vitro Metabolism of a Dual Inverse Agonist of Estrogen-Related Receptors beta and gamma, DN203368. Pharmaceutics 2021, 13, 776. [Google Scholar] [CrossRef]
- Watrous, J.; Roach, P.; Alexandrov, T.; Heath, B.S.; Yang, J.Y.; Kersten, R.D.; van der Voort, M.; Pogliano, K.; Gross, H.; Raaijmakers, J.M.; et al. Mass spectral molecular networking of living microbial colonies. Proc. Natl. Acad. Sci. USA 2012, 109, E1743–E1752. [Google Scholar] [CrossRef] [Green Version]
- Guthals, A.; Watrous, J.D.; Dorrestein, P.C.; Bandeira, N. The spectral networks paradigm in high throughput mass spectrometry. Mol. BioSyst. 2012, 8, 2535–2544. [Google Scholar] [CrossRef] [Green Version]
- Hegazi, N.M.; Radwan, R.A.; Bakry, S.M.; Saad, H.H. Molecular networking aided metabolomic profiling of beet leaves using three extraction solvents and in relation to its anti-obesity effects. J. Adv. Res. 2020, 24, 545–555. [Google Scholar] [CrossRef]
- Xie, H.; Kong, Y.-S.; Li, R.-Z.; Nothias, L.-F.; Melnik, A.V.; Zhang, H.; Liu, L.-L.; An, T.-T.; Liu, R.; Yang, Z.; et al. Feature-Based Molecular Networking Analysis of the Metabolites Produced by In Vitro Solid-State Fermentation Reveals Pathways for the Bioconversion of Epigallocatechin Gallate. J. Agric. Food Chem. 2020, 68, 7995–8007. [Google Scholar] [CrossRef] [PubMed]
- Aron, A.T.; Gentry, E.C.; McPhail, K.L.; Nothias, L.-F.; Nothias-Esposito, M.; Bouslimani, A.; Petras, D.; Gauglitz, J.M.; Sikora, N.; Vargas, F.; et al. Reproducible molecular networking of untargeted mass spectrometry data using GNPS. Nat. Protoc. 2020, 15, 1954–1991. [Google Scholar] [CrossRef]
- Guan, F.; Uboh, C.E.; Soma, L.R.; Rudy, J. Sequence Elucidation of an Unknown Cyclic Peptide of High Doping Potential by ETD and CID Tandem Mass Spectrometry. J. Am. Soc. Mass Spectrom. 2011, 22, 718–730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smyth, W.F.; McClean, S.; Ramachandran, V.N. A study of the electrospray ionisation of pharmacologically significant 1,4-benzodiazepines and their subsequent fragmentation using an ion-trap mass spectrometer. Rapid Commun. Mass Spectrom. 2000, 14, 2061–2069. [Google Scholar] [CrossRef]
- Chen, G.; Li, X.; Saleri, F.; Guo, M. Analysis of Flavonoids in Rhamnus davurica and Its Antiproliferative Activities. Molecules 2016, 21, 1275. [Google Scholar] [CrossRef] [PubMed]
- Gao, R.; Zhong, D.; Liu, K.; Xia, Y.; Shi, R.; Li, H.; Chen, X. Simultaneous determination of morinidazole, its N-oxide, sulfate, and diastereoisomeric N+-glucuronides in human plasma by liquid chromatography–tandem mass spectrometry. J. Chromatogr. B Analyt. Technol. Biomed. Life Sci. 2012, 908, 52–58. [Google Scholar] [CrossRef]
- Dai, W.; Yin, P.; Zeng, Z.; Kong, H.; Tong, H.; Xu, Z.; Lu, X.; Lehmann, R.; Xu, G. Nontargeted modification-specific metabolomics study based on liquid chromatography-high-resolution mass spectrometry. Anal. Chem. 2014, 86, 9146–9153. [Google Scholar] [CrossRef]
- Joo, J.; Lee, D.; Wu, Z.; Shin, J.H.; Lee, H.S.; Kwon, B.M.; Huh, T.L.; Kim, Y.W.; Lee, S.J.; Kim, T.W.; et al. In vitro metabolism of obovatol and its effect on cytochrome P450 enzyme activities in human liver microsomes. Biopharm. Drug Dispos. 2013, 34, 195–202. [Google Scholar] [CrossRef]
- Wang, Z.; Fang, Y.; Rock, D.; Ma, J. Rapid screening and characterization of glutathione-trapped reactive metabolites using a polarity switch-based approach on a high-resolution quadrupole orbitrap mass spectrometer. Anal. Bioanal. Chem. 2017, 410, 1595–1606. [Google Scholar] [CrossRef]
- Liu, C.-W.; Lu, Y.-Y.; Yang, Z.-Z.; Xing, Y.-Y.; Xi, T. Rapid screening and characterization of metabolites from a marine-derived actinomycete by high-performance liquid chromatography coupled with electrospray ionization quadrupole time-of-flight mass spectrometry. Rapid Commun. Mass Spectrom. 2010, 24, 3413–3418. [Google Scholar] [CrossRef] [PubMed]
- Kubwabo, C.; Vais, N.; Benoit, F.M. Characterization of microcystins using in-source collision-induced dissociation. Rapid Commun. Mass Spectrom. 2005, 19, 597–604. [Google Scholar] [CrossRef] [PubMed]
- Wacher, V.J.; Silverman, J.A.; Zhang, Y.; Benet, L.Z. Role of P-Glycoprotein and Cytochrome P450 3A in Limiting Oral Absorption of Peptides and Peptidomimetics. J. Pharm. Sci. 1998, 87, 1322–1330. [Google Scholar] [CrossRef] [PubMed]
- Fitzsimmons, M.E.; Collins, J.M. Selective biotransformation of the human immunodeficiency virus protease inhibitor saquinavir by human small-intestinal cytochrome P4503A4: Potential contribution to high first-pass metabolism. Drug Metab. Dispos. 1997, 25, 256–266. [Google Scholar] [PubMed]
- Eagling, V.A.; Back, D.J.; Barry, M.G. Differential inhibition of cytochrome P450 isoforms by the protease inhibitors, ritonavir, saquinavir and indinavir. Br. J. Clin. Pharmacol. 1997, 44, 190–194. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Metabolic Pathway | tR (min) | m/z ((M + H)+) | Error (ppm) | Formula (Neutral) | |
|---|---|---|---|---|---|---|
| Measured | Theoretical | |||||
| P | Seongsanamide A | 7.8 | 1005.5647 | 1005.5655 | –0.822 | C52H77O12N8 |
| M1 | Hydrolysis | 5.3 | 836.4524 | 836.4553 | –3.368 | C43H62O10N7 |
| M2 | Mono-hydroxylation | 6.9 | 1021.5589 | 1021.5605 | –1.459 | C52H77O13N8 |
| M3 | Di-hydroxylation | 6.7 | 1037.5519 | 1037.5554 | –3.263 | C52H77O14N8 |
| M4 | Tri-hydroxylation | 6.4 | 1053.5475 | 1053.5503 | –2.639 | C52H77O15N8 |
| Activity | P450 Isoforms | IC50 (µM) |
|---|---|---|
| Phenacetin O-deethylation | CYP1A2 | >50 |
| Coumarin 7-hydroxylation | CYP2A6 | >50 |
| Bupropion hydroxylation | CYP2B6 | 21.9 |
| Amodiaquine N-deethylation | CYP2C8 | >50 |
| Tolbutamide 4-methylhydroxylation | CYP2C9 | >50 |
| Omeprazole 5-Hydroxylation | CYP2C19 | 25.5 |
| Dextromethorphan O-demethylation | CYP2D6 | 23.7 |
| Chlorzoxazone 6-hydroxylation | CYP2E1 | >50 |
| Midazolam 1′-hydroxylation | CYP3A | 13.1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Z.; Kim, G.J.; Park, S.-Y.; Shon, J.C.; Liu, K.-H.; Choi, H. In Vitro Metabolism Study of Seongsanamide A in Human Liver Microsomes Using Non-Targeted Metabolomics and Feature-Based Molecular Networking. Pharmaceutics 2021, 13, 1031. https://doi.org/10.3390/pharmaceutics13071031
Wu Z, Kim GJ, Park S-Y, Shon JC, Liu K-H, Choi H. In Vitro Metabolism Study of Seongsanamide A in Human Liver Microsomes Using Non-Targeted Metabolomics and Feature-Based Molecular Networking. Pharmaceutics. 2021; 13(7):1031. https://doi.org/10.3390/pharmaceutics13071031
Chicago/Turabian StyleWu, Zhexue, Geum Jin Kim, So-Young Park, Jong Cheol Shon, Kwang-Hyeon Liu, and Hyukjae Choi. 2021. "In Vitro Metabolism Study of Seongsanamide A in Human Liver Microsomes Using Non-Targeted Metabolomics and Feature-Based Molecular Networking" Pharmaceutics 13, no. 7: 1031. https://doi.org/10.3390/pharmaceutics13071031