
Preclinical Assessment of Ursolic Acid Loaded into Nanostructured Lipid Carriers in Experimental Visceral Leishmaniasis


 , , ,
, , ,
Abstract
:
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Preparation Method of Nanostructured Lipid Carriers (NLCs)
2.3. Physical-Chemical Characterization of Nanoparticles
2.3.1. Determination of Encapsulation Efficiency
2.3.2. Determination of Size, Polydispersity, and Zeta Potential
2.3.3. Morphological Analysis of UA-NLC
2.4. Animal and Ethical Considerations
2.5. Histological and Biochemical Changes of Healthy Hamsters Treated with UA-NLC
2.6. Analysis of the Therapeutic Potential of UA-NLC
2.6.1. Determination of Parasite Load
2.6.2. Analysis of Cellular and Humoral Immune Responses
2.7. Statistical Analyses
3. Results
3.1. Physical Characterization of NLC
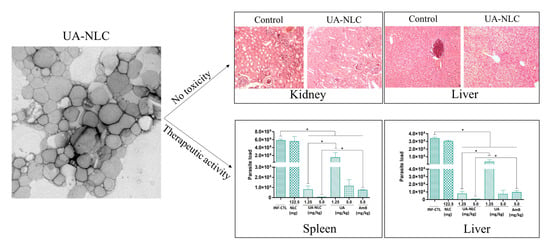
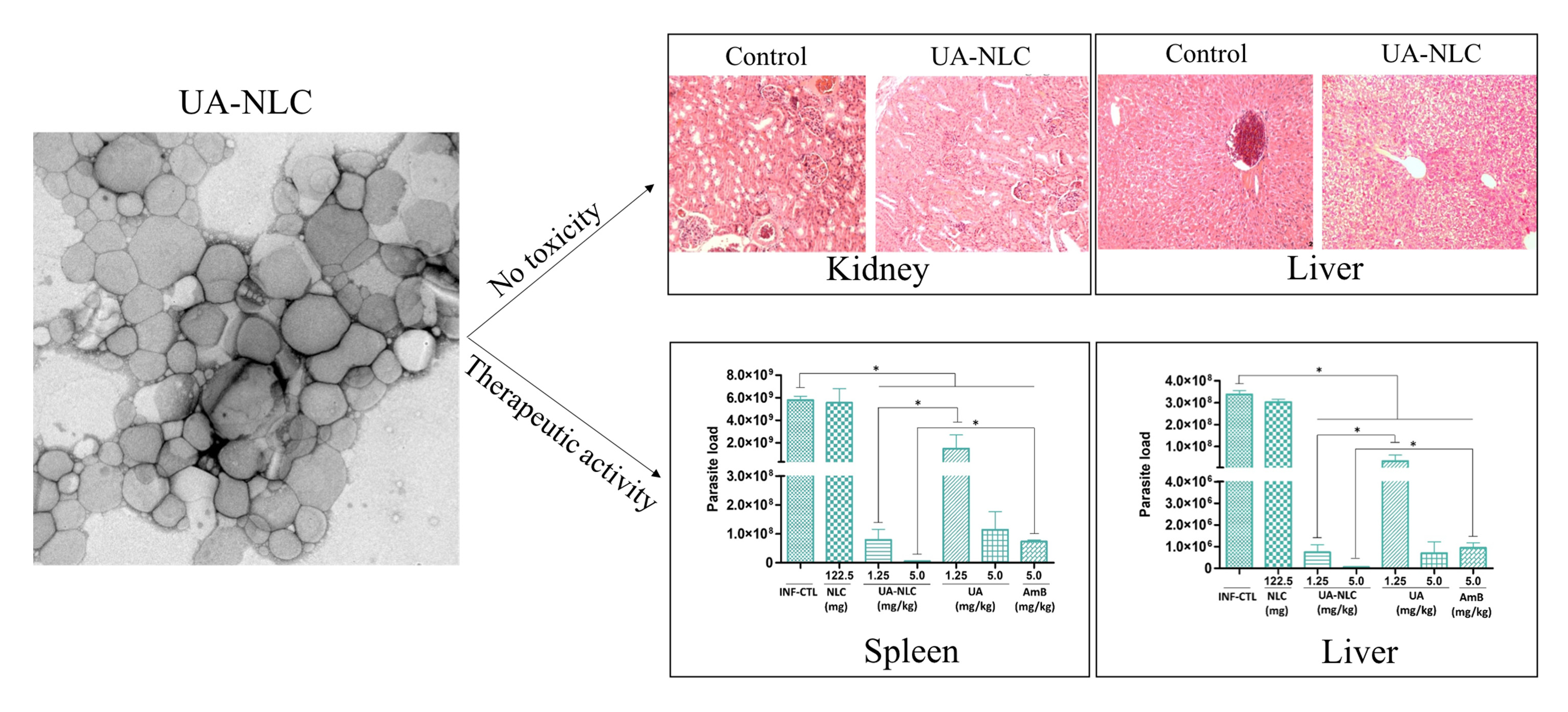
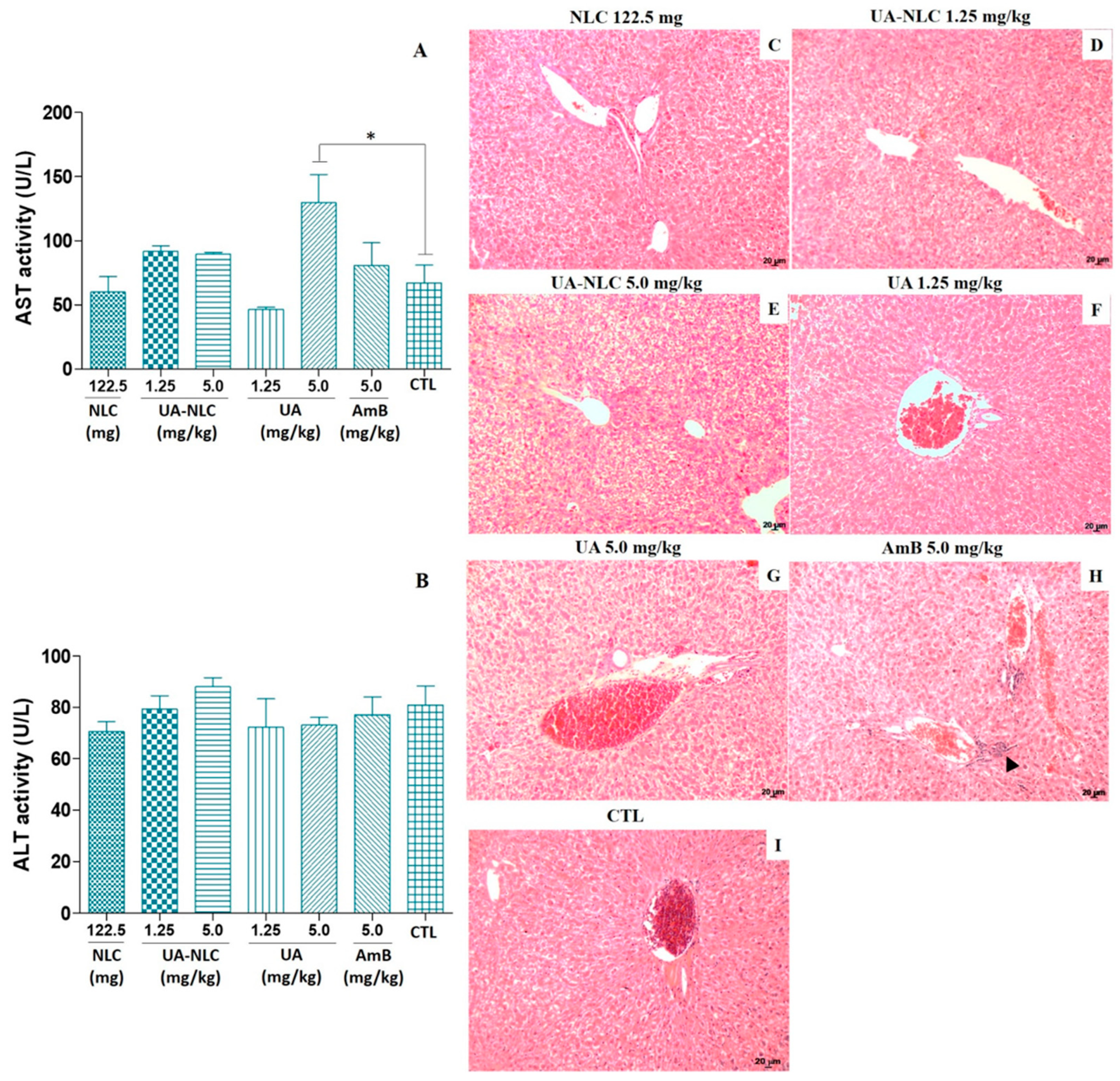
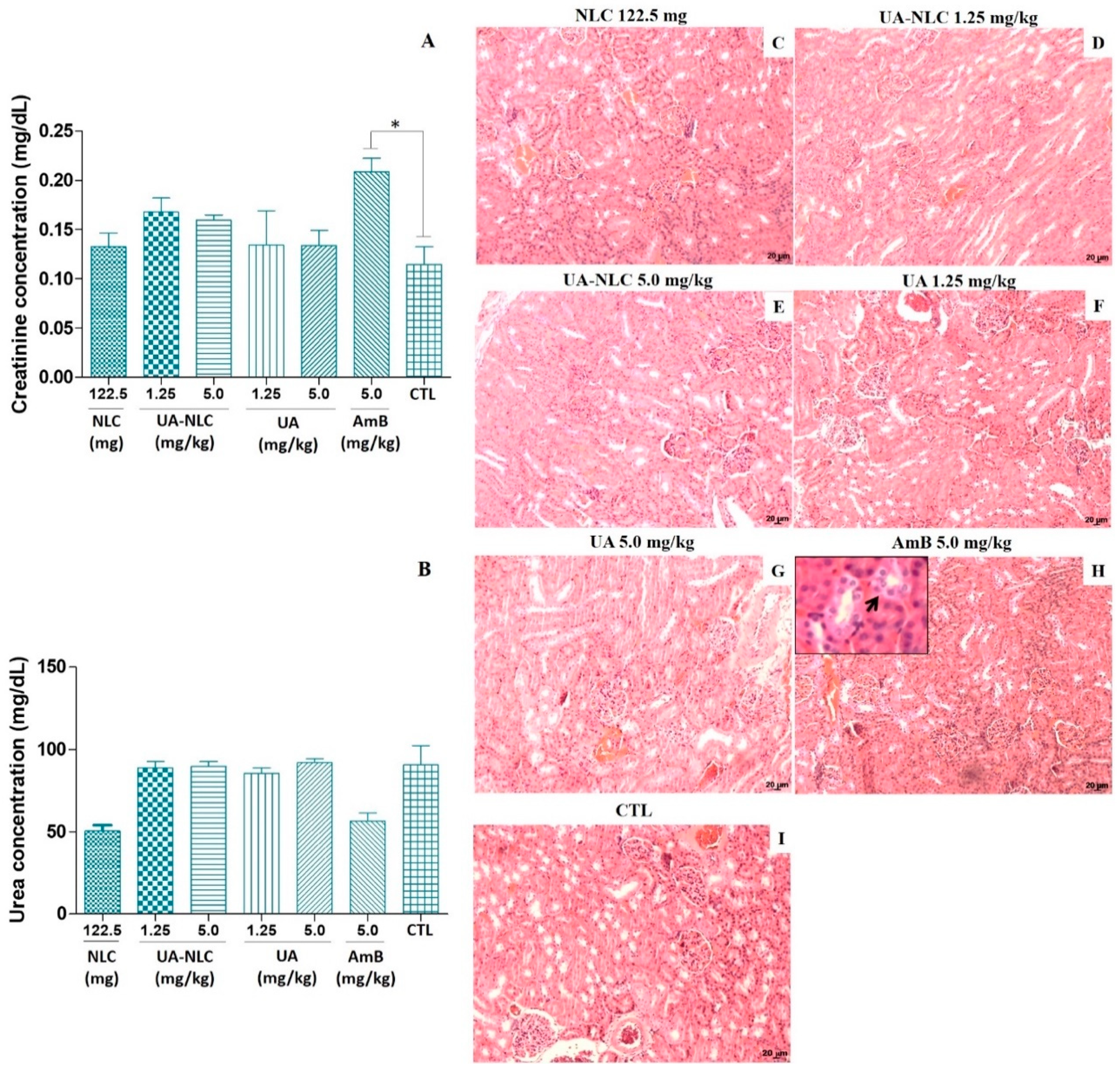
3.2. Biochemical and Histological Changes of Healthy Hamsters Treated with UA-NLC
3.3. Analysis of the Therapeutic Potential of UA-NLC
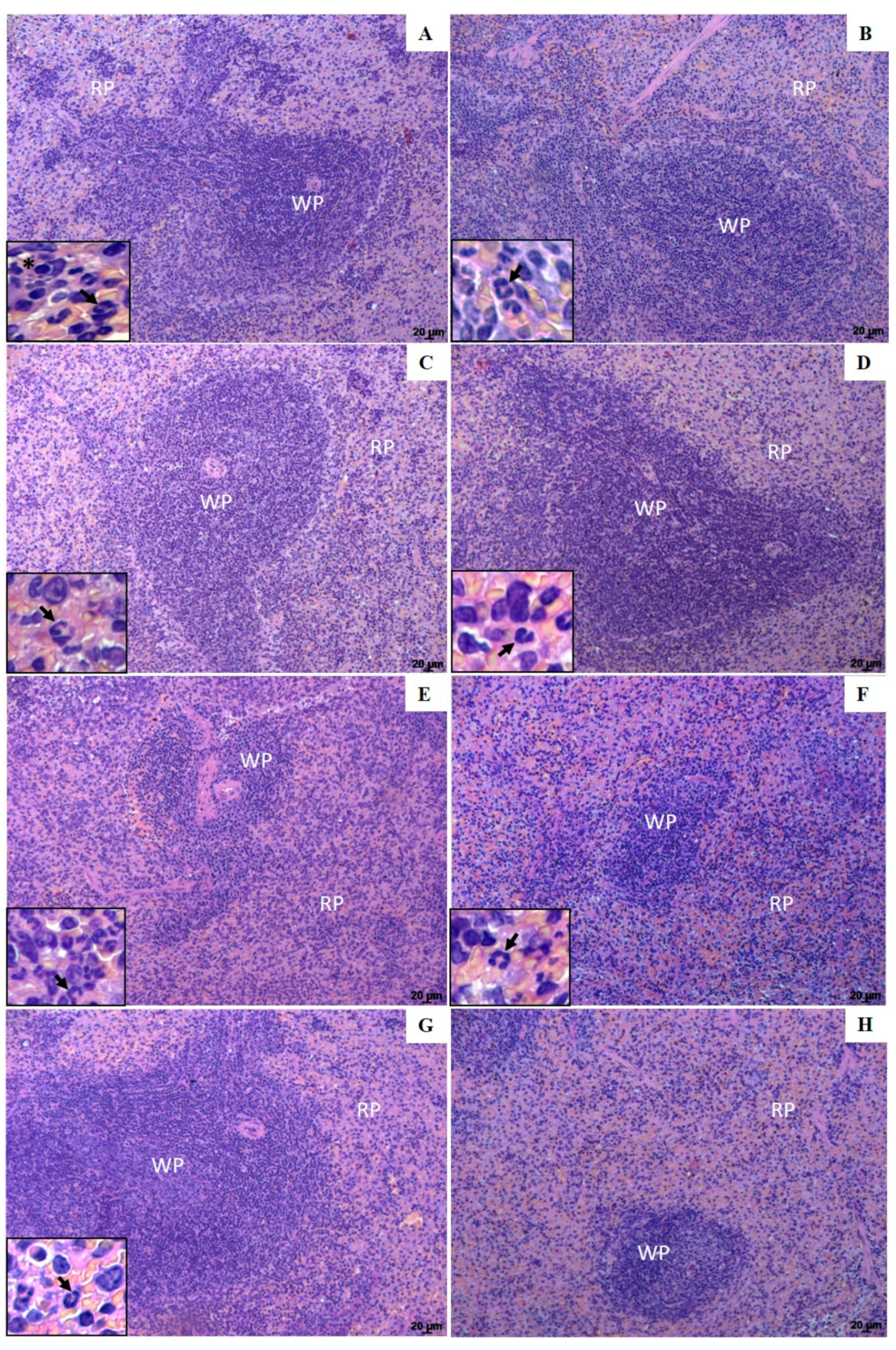
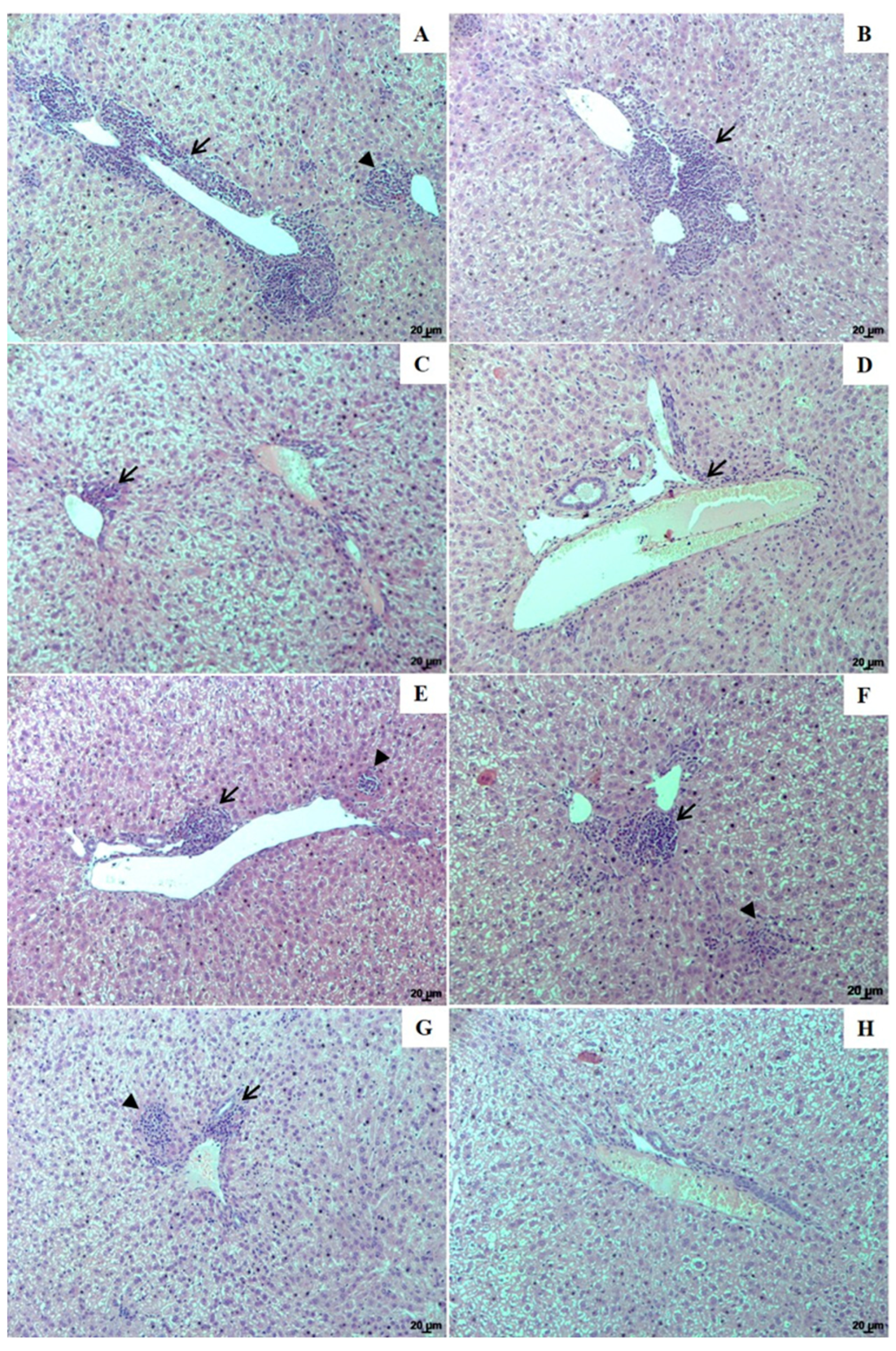
3.4. Histopathological Changes in the Spleen and Liver of Animals Treated with UA-NLC or UA
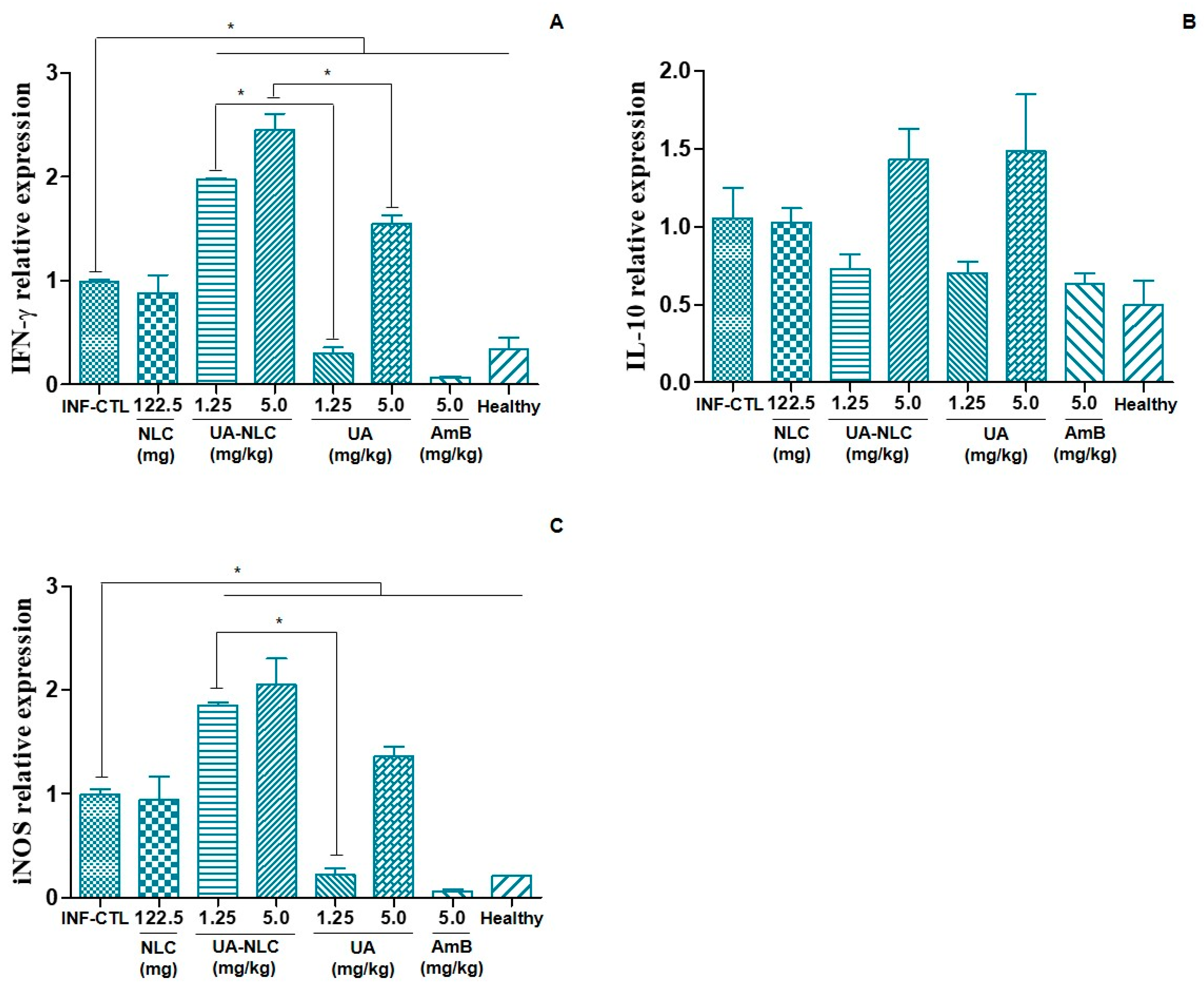
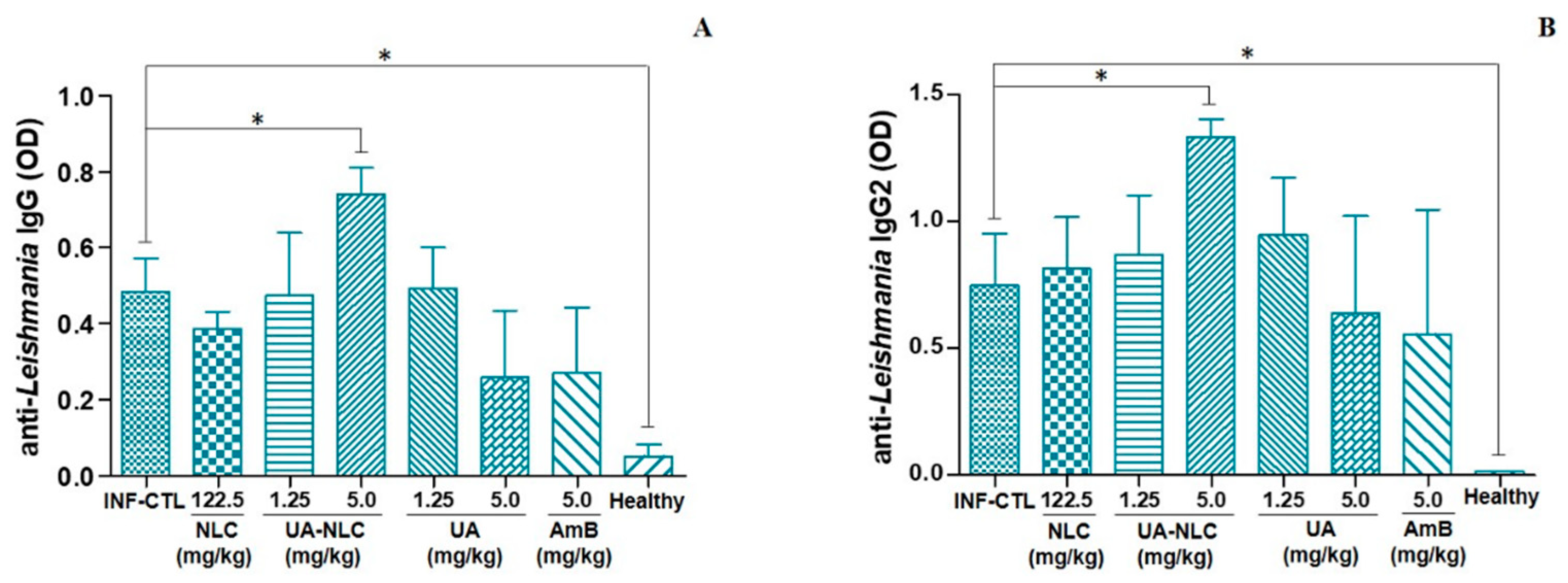
3.5. Analysis of Cellular and Humoral Immune Responses
3.6. Biochemical Analysis of Hamsters Infected and Treated with UA-NLC
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- WHO Expert Committee on the Control of the Leishmaniases & World Health Organization. Control of the leishmaniases: Report of a meeting of the WHO Expert Commitee on the Control of Leishmaniases, Geneva, 22–26 March 2020. Available online: https://apps.who.int/iris/handle/10665/44412 (accessed on 21 May 2021).
- Alvar, J.; Vélez, I.D.; Bern, C.; Herrero, M.; Desjeux, P.; Cano, J.; Jannin, J.; Boer, M. den Leishmaniasis Worldwide and Global Estimates of Its Incidence. PLoS ONE 2012, 7, e35671. [Google Scholar] [CrossRef] [PubMed]
- Torres-Guerrero, E.; Quintanilla-Cedillo, M.R.; Ruiz-Esmenjaud, J.; Arenas, R. Leishmaniasis: A review. F1000Research 2017, 6, 750. [Google Scholar] [CrossRef] [PubMed]
- Passero, L.F.D.; Cruz, L.A.; Santos-Gomes, G.; Rodrigues, E.; Laurenti, M.D.M.D.; Lago, J.H.G.J.H.G. Conventional Versus Natural Alternative Treatments for Leishmaniasis: A Review. Curr. Top. Med. Chem. 2018, 18, 1275–1286. [Google Scholar] [CrossRef] [PubMed]
- Ghorbani, M.; Farhoudi, R. Leishmaniasis in humans: Drug or vaccine therapy? Drug Des. Devel. Ther. 2018, 12, 25–40. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponte-Sucre, A.; Gamarro, F.; Dujardin, J.-C.; Barrett, M.P.; López-Vélez, R.; García-Hernández, R.; Pountain, A.W.; Mwenechanya, R.; Papadopoulou, B. Drug resistance and treatment failure in leishmaniasis: A 21st century challenge. PLoS Negl. Trop. Dis. 2017, 11, e0006052. [Google Scholar] [CrossRef]
- Passero, L.F.D.; Laurenti, M.D.; Santos-Gomes, G.; Soares Campos, B.L.; Sartorelli, P.; Lago, J.H.G. Plants used in traditional medicine: Extracts and secondary metabolites exhibiting antileishmanial activity. Curr. Clin. Pharmacol. 2014, 9, 187–204. [Google Scholar] [CrossRef]
- Ferreira, D.D.S.; Esperandim, V.R.; Marçal, M.G.; Neres, N.B.D.R.; Cunha, N.L.; Andrade e Silva, M.L.; Cunha, W.R. Natural products and Chagas’ disease: The action of triterpenes acids isolated from Miconia species. Univ. Sci. 2013, 18. [Google Scholar] [CrossRef]
- Choi, W.; Lee, I. Evaluation of Anti-Toxoplasma gondii Effect of Ursolic Acid as a Novel Toxoplasmosis Inhibitor. Pharmaceuticals 2018, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Jesus, J.A.; Fragoso, T.N.; Yamamoto, E.S.; Laurenti, M.D.; Silva, M.S.; Ferreira, A.F.; Lago, J.H.G.; Gomes, G.S.; Passero, L.F.D. Therapeutic effect of ursolic acid in experimental visceral leishmaniasis. Int. J. Parasitol. Drugs Drug Resist. 2017, 7, 1–11. [Google Scholar] [CrossRef]
- Bilbao-Ramos, P.; Serrano, D.R.; Ruiz Saldaña, H.K.; Torrado, J.J.; Bolás-Fernández, F.; Dea-Ayuela, M.A. Evaluating the Potential of Ursolic Acid as Bioproduct for Cutaneous and Visceral Leishmaniasis. Molecules 2020, 25, 1394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamamoto, E.S.; Campos, B.L.S.; Jesus, J.A.; Laurenti, M.D.; Ribeiro, S.P.; Kallás, E.G.; Rafael-Fernandes, M.; Santos-Gomes, G.; Silva, M.S.; Sessa, D.P.; et al. The Effect of Ursolic Acid on Leishmania (Leishmania) amazonensis Is Related to Programed Cell Death and Presents Therapeutic Potential in Experimental Cutaneous Leishmaniasis. PLoS ONE 2015, 10, e0144946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labib, R.M.; Ebada, S.S.; Youssef, F.S.; Ashour, M.L.; Ross, S.A. Ursolic Acid, a Natural Pentacylcic Triterpene from Ochrosia elliptica and Its Role in The Management of Certain Neglected Tropical Diseases. Pharmacogn. Mag. 2016, 12, 319–325. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, E.S.; Campos, B.L.; Laurenti, M.D.; Lago, J.H.G.; Grecco, S.d.S.; Corbett, C.E.; Passero, L.F. Treatment with triterpenic fraction purified from Baccharis uncinella leaves inhibits Leishmania (Leishmania) amazonensis spreading and improves Th1 immune response in infected mice. Parasitol. Res. 2014, 113, 333–339. [Google Scholar] [CrossRef]
- Khalil, N.M.; de Mattos, A.C.; Moraes Moreira Carraro, T.; Ludwig, D.B.; Mainardes, R.M.; Carraro, T.C.M.M.; Ludwig, D.B.; Mainardes, R.M. Nanotechnological strategies for the treatment of neglected diseases. Curr. Pharm. Des. 2013, 19, 7316–7329. [Google Scholar] [CrossRef] [PubMed]
- Ezzati Nazhad Dolatabadi, J.; Valizadeh, H.; Hamishehkar, H. Solid Lipid Nanoparticles as Efficient Drug and Gene Delivery Systems: Recent Breakthroughs. Adv. Pharm. Bull. 2015, 5, 151–159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Monteiro, L.M.; Löbenberg, R.; Fotaki, N.; de Araújo, G.L.B.; Cotrim, P.C.; Bou-Chacra, N. Co-delivery of buparvaquone and polymyxin B in a nanostructured lipid carrier for leishmaniasis treatment. J. Glob. Antimicrob. Resist. 2019, 18, 279–283. [Google Scholar] [CrossRef]
- Eloy, J.d.O.; Oliveira, E.C.V.d.; Marotta-Oliveira, S.S.; Saraiva, J.; Marchetti, J.M. Desenvolvimento e validação de um método analítico por CLAE para quantificação de ácido ursólico em dispersões sólidas. Quim. Nova 2012, 35, 1036–1040. [Google Scholar] [CrossRef] [Green Version]
- Corral, M.J.; Serrano, D.R.; Moreno, I.; Torrado, J.J.; Dominguez, M.; Alunda, J.M. Efficacy of low doses of amphotericin B plus allicin against experimental visceral leishmaniasis. J. Antimicrob. Chemother. 2014, 69, 3268–3274. [Google Scholar] [CrossRef] [Green Version]
- Spada, T.C.; Silva, J.M.R.D.; Francisco, L.S.; Marçal, L.J.; Antonangelo, L.; Zanetta, D.M.T.; Yu, L.; Burdmann, E.A. High intensity resistance training causes muscle damage and increases biomarkers of acute kidney injury in healthy individuals. PLoS ONE 2018, 13, e0205791. [Google Scholar] [CrossRef]
- Laurenti, M.D.; Passero, L.F.D.; Tomokane, T.Y.; Francesquini, F.d.C.; Rocha, M.C.; Gomes, C.M.d.C.; Corbett, C.E.P.; Silveira, F.T. Dynamic of the Cellular Immune Response at the Dermal Site of Leishmania (L.) amazonensis and Leishmania (V.) braziliensis Infection in Sapajus apella Primate. Biomed. Res. Int. 2014, 2014, 1–8. [Google Scholar] [CrossRef]
- Lafuse, W.P.; Story, R.; Mahylis, J.; Gupta, G.; Varikuti, S.; Steinkamp, H.; Oghumu, S.; Satoskar, A.R. Leishmania donovani Infection Induces Anemia in Hamsters by Differentially Altering Erythropoiesis in Bone Marrow and Spleen. PLoS ONE 2013, 8, e59509. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rama Iñiguez, S.; Dea-Ayuela, M.A.; Sanchez-Brunete, J.A.; Torrado, J.J.; Alunda, J.M.; Bolas-Fernández, F. Real-Time Reverse Transcription-PCR Quantification of Cytokine mRNA Expression in Golden Syrian Hamster Infected with Leishmania infantum and Treated with a New Amphotericin B Formulation. Antimicrob. Agents Chemother. 2006, 50, 1195–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gupta, R.; Kushawaha, P.K.; Samant, M.; Jaiswal, A.K.; Baharia, R.K.; Dube, A. Treatment of Leishmania donovani-infected hamsters with miltefosine: Analysis of cytokine mRNA expression by real-time PCR, lymphoproliferation, nitrite production and antibody responses. J. Antimicrob. Chemother. 2012, 67, 440–443. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- de Souza, A.; Marins, D.S.S.; Mathias, S.L.; Monteiro, L.M.; Yukuyama, M.N.; Scarim, C.B.; Löbenberg, R.; Bou-Chacra, N.A. Promising nanotherapy in treating leishmaniasis. Int. J. Pharm. 2018, 547, 421–431. [Google Scholar] [CrossRef] [Green Version]
- Dhiman, N.; Awasthi, R.; Sharma, B.; Kharkwal, H.; Kulkarni, G.T. Lipid Nanoparticles as Carriers for Bioactive Delivery. Front. Chem. 2021, 9. [Google Scholar] [CrossRef]
- Mu, H.; Holm, R. Solid lipid nanocarriers in drug delivery: Characterization and design. Expert Opin. Drug Deliv. 2018, 15, 771–785. [Google Scholar] [CrossRef]
- Hoshyar, N.; Gray, S.; Han, H.; Bao, G. The effect of nanoparticle size on in vivo pharmacokinetics and cellular interaction. Nanomedicine 2016, 11, 673–692. [Google Scholar] [CrossRef] [Green Version]
- Andonova, V.; Peneva, P. Characterization Methods for Solid Lipid Nanoparticles (SLN) and Nanostructured Lipid Carriers (NLC). Curr. Pharm. Des. 2018, 23, 6630–6642. [Google Scholar] [CrossRef]
- Barbosa, R.M.; Casadei, B.R.; Duarte, E.L.; Severino, P.; Barbosa, L.R.S.; Duran, N.; de Paula, E. Electron Paramagnetic Resonance and Small-Angle X-ray Scattering Characterization of Solid Lipid Nanoparticles and Nanostructured Lipid Carriers for Dibucaine Encapsulation. Langmuir 2018, 34, 13296–13304. [Google Scholar] [CrossRef]
- Monteiro, L.M.; Löbenberg, R.; Cotrim, P.C.; Barros de Araujo, G.L.; Bou-Chacra, N. Buparvaquone Nanostructured Lipid Carrier: Development of an Affordable Delivery System for the Treatment of Leishmaniases. Biomed. Res. Int. 2017, 2017, 1–11. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.-N.; Poon, W.; Tavares, A.J.; McGilvray, I.D.; Chan, W.C.W. Nanoparticle–liver interactions: Cellular uptake and hepatobiliary elimination. J. Control. Release 2016, 240, 332–348. [Google Scholar] [CrossRef]
- Longmire, M.; Choyke, P.L.; Kobayashi, H. Clearance properties of nano-sized particles and molecules as imaging agents: Considerations and caveats. Nanomedicine 2008, 3, 703–717. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Moreira, N.d.D.; Vitoriano-Souza, J.; Roatt, B.M.; Vieira, P.M.d.A.; Coura-Vital, W.; Cardoso, J.M.d.O.; Rezende, M.T.; Ker, H.G.; Giunchetti, R.C.; Carneiro, C.M.; et al. Clinical, hematological and biochemical alterations in hamster (Mesocricetus auratus) experimentally infected with Leishmania infantum through different routes of inoculation. Parasit. Vectors 2016, 9, 181. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saravanan, R.; Viswanathan, P.; Pugalendi, K.V. Protective effect of ursolic acid on ethanol-mediated experimental liver damage in rats. Life Sci. 2006, 78, 713–718. [Google Scholar] [CrossRef]
- Li, S.; Meng, F.; Liao, X.; Wang, Y.; Sun, Z.; Guo, F.; Li, X.; Meng, M.; Li, Y.; Sun, C. Therapeutic Role of Ursolic Acid on Ameliorating Hepatic Steatosis and Improving Metabolic Disorders in High-Fat Diet-Induced Non-Alcoholic Fatty Liver Disease Rats. PLoS ONE 2014, 9, e86724. [Google Scholar] [CrossRef] [Green Version]
- Martin-Aragón, S.; de las Heras, B.; Sanchez-Reus, M.I.; Benedi, J. Pharmacological modification of endogenous antioxidant enzymes by ursolic acid on tetrachloride-induced liver damagein rats and primary cultures of rat hepatocytes. Exp. Toxicol. Pathol. 2001, 53, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.-H.; Zhou, S.-Y.; Qian, Z.-Z.; Zhang, H.-L.; Qiu, L.-H.; Song, Z.; Zhao, J.; Wang, P.; Hao, X.-S.; Wang, H.-Q. Evaluation of toxicity and single-dose pharmacokinetics of intravenous ursolic acid liposomes in healthy adult volunteers and patients with advanced solid tumors. Expert Opin. Drug Metab. Toxicol. 2013, 9, 117–125. [Google Scholar] [CrossRef]
- Lobo, P.C.B.; Vieira, I.P.; Pichard, C.; Marques, B.S.; Gentil, P.; da Silva, E.L.; Pimentel, G.D. Ursolic acid has no additional effect on muscle strength and mass in active men undergoing a high-protein diet and resistance training: A double-blind and placebo-controlled trial. Clin. Nutr. 2021, 40, 581–589. [Google Scholar] [CrossRef]
- Valenzuela, P.L.; Morales, J.S.; Emanuele, E.; Pareja-Galeano, H.; Lucia, A. Supplements with purported effects on muscle mass and strength. Eur. J. Nutr. 2019, 58, 2983–3008. [Google Scholar] [CrossRef] [PubMed]
- Bang, H.S.; Seo, D.Y.; Chung, Y.M.; Kim, D.H.; Lee, S.-J.; Lee, S.R.; Kwak, H.-B.; Kim, T.N.; Kim, M.; Oh, K.-M.; et al. Ursolic acid supplementation decreases markers of skeletal muscle damage during resistance training in resistance-trained men: A pilot study. Korean J. Physiol. Pharmacol. 2017, 21, 651. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- ZHANG, F.; DAIMARU, E.; OHNISHI, M.; KINOSHITA, M.; TOKUJI, Y. Oleanolic Acid and Ursolic Acid in Commercial Dried Fruits. Food Sci. Technol. Res. 2013, 19, 113–116. [Google Scholar] [CrossRef] [Green Version]
- Berdichevski, R.H.; Luis, L.B.; Crestana, L.; Manfro, R.C. Amphotericin B-related nephrotoxicity in low-risk patients. Braz. J. Infect. Dis. 2006, 10. [Google Scholar] [CrossRef] [PubMed]
- Hamill, R.J. Amphotericin B Formulations: A Comparative Review of Efficacy and Toxicity. Drugs 2013, 73, 919–934. [Google Scholar] [CrossRef]
- Laniado-Laborín, R.; Cabrales-Vargas, M.N. Amphotericin B: Side effects and toxicity. Rev. Iberoam. Micol. 2009, 26, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Deray, G. Amphotericin B nephrotoxicity. J. Antimicrob. Chemother. 2002, 49, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Afzal, I.; Sarwar, H.S.; Sohail, M.F.; Varikuti, S.; Jahan, S.; Akhtar, S.; Yasinzai, M.; Satoskar, A.R.; Shahnaz, G. Mannosylated thiolated paromomycin-loaded PLGA nanoparticles for the oral therapy of visceral leishmaniasis. Nanomedicine 2019, 14, 387–406. [Google Scholar] [CrossRef]
- Esfandiari, F.; Motazedian, M.H.; Asgari, G.; Morowvat, M.H.; Molaei, M.; Heli, H. Paromomycin-loaded mannosylated chitosan nanoparticles: Synthesis, characterization and targeted drug delivery against leishmaniasis. Acta Trop. 2019, 197, 105045. [Google Scholar] [CrossRef]
- Jansook, P.; Fülöp, Z.; Ritthidej, G.C. Amphotericin B loaded solid lipid nanoparticles (SLNs) and nanostructured lipid carrier (NLCs): Physicochemical and solid-solution state characterizations. Drug Dev. Ind. Pharm. 2019, 45, 560–567. [Google Scholar] [CrossRef]
- Fernández-García, R.; Lalatsa, A.; Statts, L.; Bolás-Fernández, F.; Ballesteros, M.P.; Serrano, D.R. Transferosomes as nanocarriers for drugs across the skin: Quality by design from lab to industrial scale. Int. J. Pharm. 2020, 573, 118817. [Google Scholar] [CrossRef]
- Das, S.; Ghosh, S.; De, A.K.; Bera, T. Oral delivery of ursolic acid-loaded nanostructured lipid carrier coated with chitosan oligosaccharides: Development, characterization, in vitro and in vivo assessment for the therapy of leishmaniasis. Int. J. Biol. Macromol. 2017, 102, 996–1008. [Google Scholar] [CrossRef] [PubMed]
- Kar, N.; Chakraborty, S.; De, A.K.; Ghosh, S.; Bera, T. Development and evaluation of a cedrol-loaded nanostructured lipid carrier system for in vitro and in vivo susceptibilities of wild and drug resistant Leishmania donovani amastigotes. Eur. J. Pharm. Sci. 2017, 104, 196–211. [Google Scholar] [CrossRef] [PubMed]
- Kima, P.E.; Soong, L. Interferon Gamma in Leishmaniasis. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [Green Version]
- Namdar Ahmadabad, H.; Shafiei, R.; Hatam, G.R.; Zolfaghari Emameh, R.; Aspatwar, A. Cytokine profile and nitric oxide levels in peritoneal macrophages of BALB/c mice exposed to the fucose-mannose ligand of Leishmania infantum combined with glycyrrhizin. Parasit. Vectors 2020, 13, 363. [Google Scholar] [CrossRef]
- Campos, B.L.S.; Silva, T.N.; Ribeiro, S.P.; Carvalho, K.I.L.; KallÁs, E.G.; Laurenti, M.D.; Passero, L.F.D. Analysis of iron superoxide dismutase-encoding DNA vaccine on the evolution of the Leishmania amazonensis experimental infection. Parasite Immunol. 2015, 37, 407–416. [Google Scholar] [CrossRef]
- Passero, L.F.D.; Dacostabordon, M.L.A.; De Carvalho, A.K.; Martins, L.M.; Corbett, C.E.P.; Laurenti, M.D. Exacerbation of Leishmania (Viannia) shawi infection in BALB/c mice after immunization with soluble antigen from amastigote forms. APMIS 2010, 118, 973–981. [Google Scholar] [CrossRef]
- Ploquin, A.; Szécsi, J.; Mathieu, C.; Guillaume, V.; Barateau, V.; Ong, K.C.; Wong, K.T.; Cosset, F.-L.; Horvat, B.; Salvetti, A. Protection Against Henipavirus Infection by Use of Recombinant Adeno-Associated Virus–Vector Vaccines. J. Infect. Dis. 2013, 207, 469–478. [Google Scholar] [CrossRef] [Green Version]
- Kumari, S.; Samant, M.; Misra, P.; Khare, P.; Sisodia, B.; Shasany, A.K.; Dube, A. Th1-stimulatory polyproteins of soluble Leishmania donovani promastigotes ranging from 89.9 to 97.1kDa offers long-lasting protection against experimental visceral leishmaniasis. Vaccine 2008, 26, 5700–5711. [Google Scholar] [CrossRef]
- Ansari, N.A.; Saluja, S.; Salotra, P. Elevated levels of interferon-γ, interleukin-10, and interleukin-6 during active disease in Indian kala azar. Clin. Immunol. 2006, 119, 339–345. [Google Scholar] [CrossRef]
- Goto, H.; Prianti, M. das G. Immunoactivation and immunopathogeny during active visceral leishmaniasis. Rev. Inst. Med. Trop. Sao Paulo 2009, 51, 241–246. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, W.H.; Lee, I.A. The Mechanism of Action of Ursolic Acid as a Potential Anti-Toxoplasmosis Agent, and Its Immunomodulatory Effects. Pathogens 2019, 8, 61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jamshaid, H.; Din, F.; Khan, G.M. Nanotechnology based solutions for anti-leishmanial impediments: A detailed insight. J. Nanobiotechnology 2021, 19, 106. [Google Scholar] [CrossRef] [PubMed]
- Inselmann, G.; Inselmann, U.; Heidemann, H.T. Amphotericin B and liver function. Eur. J. Intern. Med. 2002, 13, 288–292. [Google Scholar] [CrossRef]
- Yeon Kang, J.; Gao, J.; Hwan Shin, D.; Alvarez, C.; Zhong, W.; Kwon, S.G. Pharmacokinetics and Renal Toxicity of Monomeric Amphotericin B in Rats after a Multiple Dose Regimen. Pharm. Nanotechnol. 2016, 4, 16–23. [Google Scholar] [CrossRef] [Green Version]
- Botero Aguirre, J.P.; Restrepo Hamid, A.M. Amphotericin B deoxycholate versus liposomal amphotericin B: Effects on kidney function. Cochrane Database Syst. Rev. 2015, CD010481. [Google Scholar] [CrossRef] [Green Version]
- Roberts, J.; Bingham, J.; McLaren, A.C.; McLemore, R. Liposomal Formulation Decreases Toxicity of Amphotericin B In Vitro and In Vivo. Clin. Orthop. Relat. Res. 2015, 473, 2262–2269. [Google Scholar] [CrossRef] [Green Version]
- Assis, T.S.M.d.; Rosa, D.C.P.; Teixeira, E.d.M.; Cota, G.; Azeredo-da-Silva, A.L.F.; Werneck, G.; Rabello, A. The direct costs of treating human visceral leishmaniasis in Brazil. Rev. Soc. Bras. Med. Trop. 2017, 50, 478–482. [Google Scholar] [CrossRef] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nanoparticle | PS (nm) | PDI | ZP (mV) | EE (%) |
|---|---|---|---|---|
| NLC | 261.1 ± 3.9 | 0.16 ± 0.013 | −26.12 ± 1.18 | |
| UA-NLC | 266.3 ± 5.4 | 0.18 ± 0.022 | −29.26 ± 1.16 | 59.71 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jesus, J.A.; Sousa, I.M.O.; da Silva, T.N.F.; Ferreira, A.F.; Laurenti, M.D.; Antonangelo, L.; Faria, C.S.; da Costa, P.C.; de Carvalho Ferreira, D.; Passero, L.F.D. Preclinical Assessment of Ursolic Acid Loaded into Nanostructured Lipid Carriers in Experimental Visceral Leishmaniasis. Pharmaceutics 2021, 13, 908. https://doi.org/10.3390/pharmaceutics13060908
Jesus JA, Sousa IMO, da Silva TNF, Ferreira AF, Laurenti MD, Antonangelo L, Faria CS, da Costa PC, de Carvalho Ferreira D, Passero LFD. Preclinical Assessment of Ursolic Acid Loaded into Nanostructured Lipid Carriers in Experimental Visceral Leishmaniasis. Pharmaceutics. 2021; 13(6):908. https://doi.org/10.3390/pharmaceutics13060908
Chicago/Turabian StyleJesus, Jéssica Adriana, Ilza Maria Oliveira Sousa, Thays Nicolli Fragoso da Silva, Aurea Favero Ferreira, Márcia Dalastra Laurenti, Leila Antonangelo, Caroline Silvério Faria, Paulo Cardoso da Costa, Domingos de Carvalho Ferreira, and Luiz Felipe Domingues Passero. 2021. "Preclinical Assessment of Ursolic Acid Loaded into Nanostructured Lipid Carriers in Experimental Visceral Leishmaniasis" Pharmaceutics 13, no. 6: 908. https://doi.org/10.3390/pharmaceutics13060908