
Toxoplasma gondii Proteasome Subunit Alpha Type 1 with Chitosan: A Promising Alternative to Traditional Adjuvant

 ,
, 

Abstract
:
1. Introduction
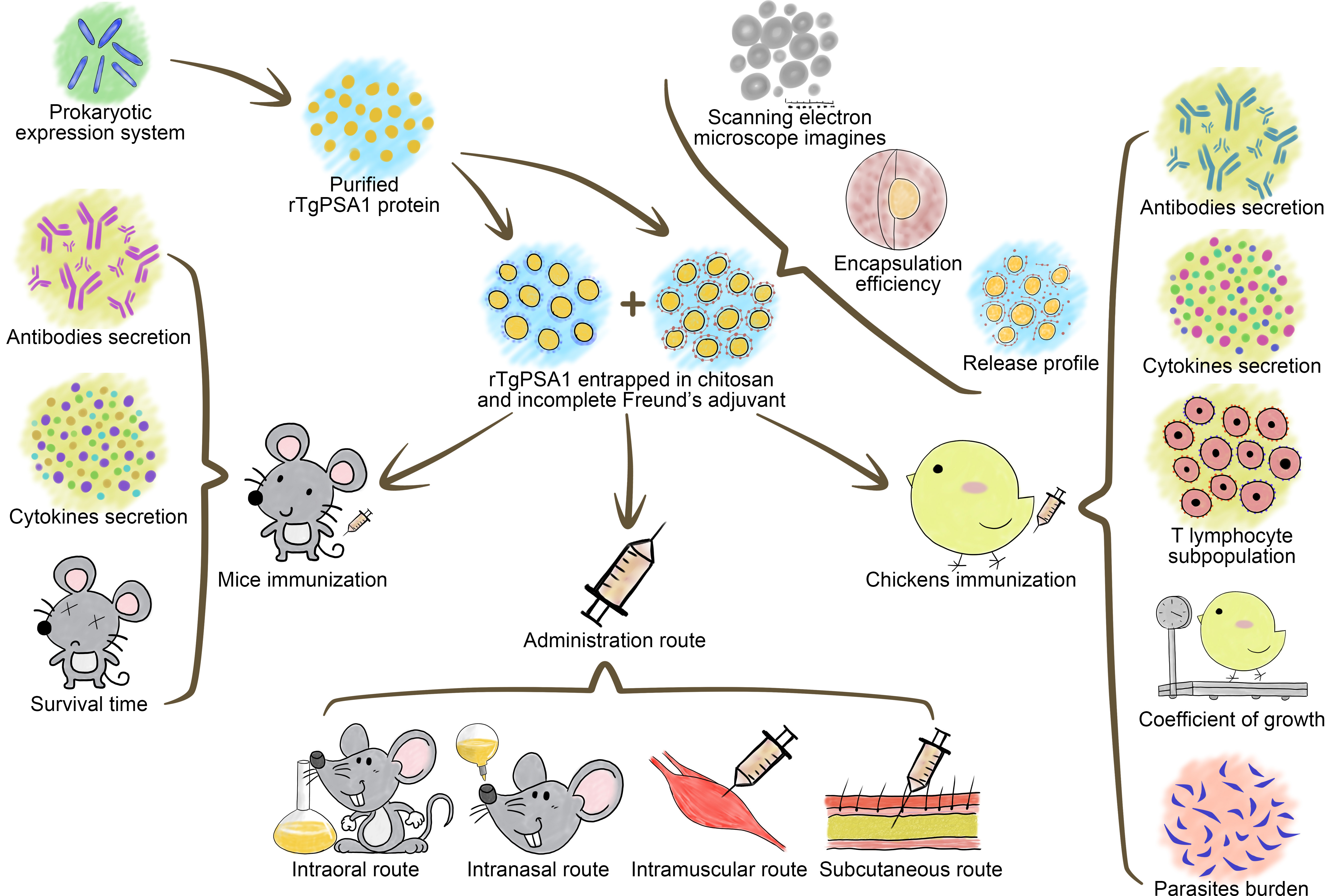
2. Materials and Methods
2.1. Mice, Rats, Poultry, Cell Lines, and T. gondii RH Strain
2.2. Cloning and Molecular Characterization of TgPSA1
2.3. Preparation of Purified Recombinant TgPSA1
2.4. Immunoblot Analysis of Recombinant and Native TgPSA1
2.5. Nanosphere Formulation (rTgPSA1/CS) and Physical Characterization
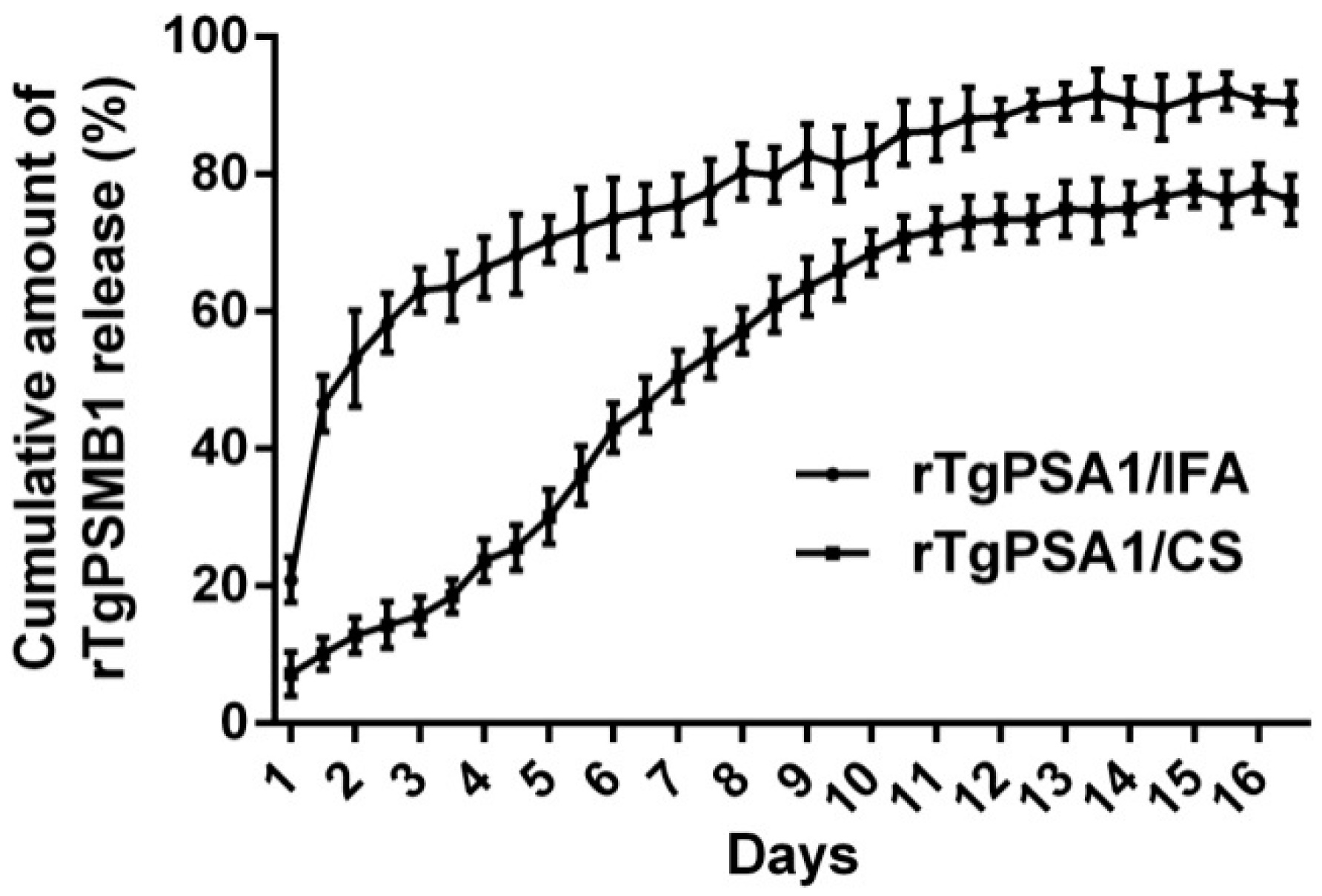
2.6. Release Characteristics In Vitro
2.7. Vaccination and Challenging Schedules in Mice
2.8. Antibody and Cytokine Assays in Mice
2.9. Determination of Optimal Administration Route
2.10. Immunization and Challenging Schedules in Chickens
2.11. Antibody and Cytokine Assays in Chickens
2.12. Detection of T Cell Subsets in Spleen Lymphocytes from Chickens
2.13. Detection of T. gondii by Absolute Quantitative PCR (qPCR)
2.14. Statistical Analysis
3. Results
3.1. Cloning, Expression, and Purification of rTgPSA1
3.2. Immunoblot Analysis of Recombinant and Native TgPSA1
3.3. Physical Characterization and Release Characteristics of rTgPSA1/CS Nanospheres
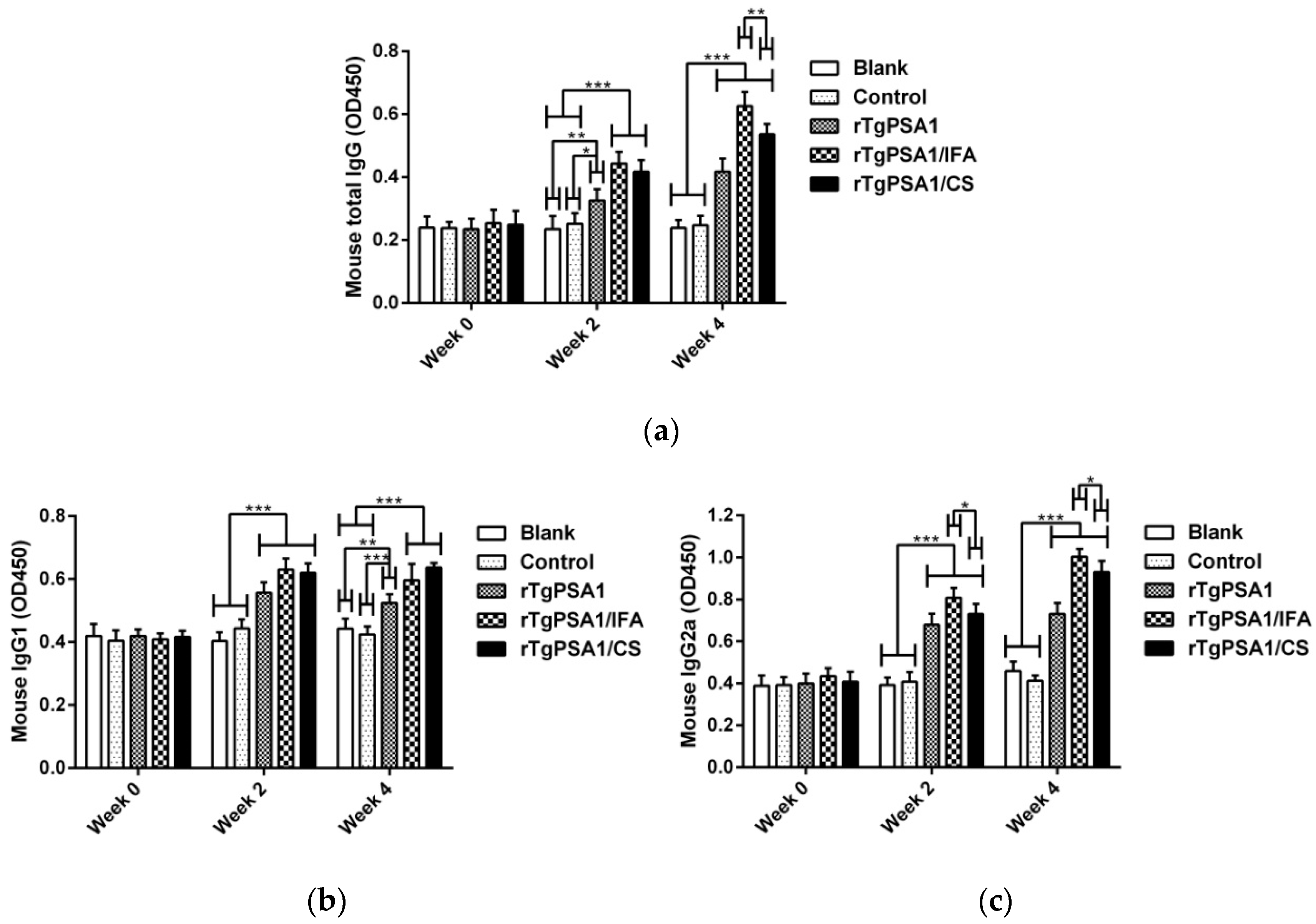
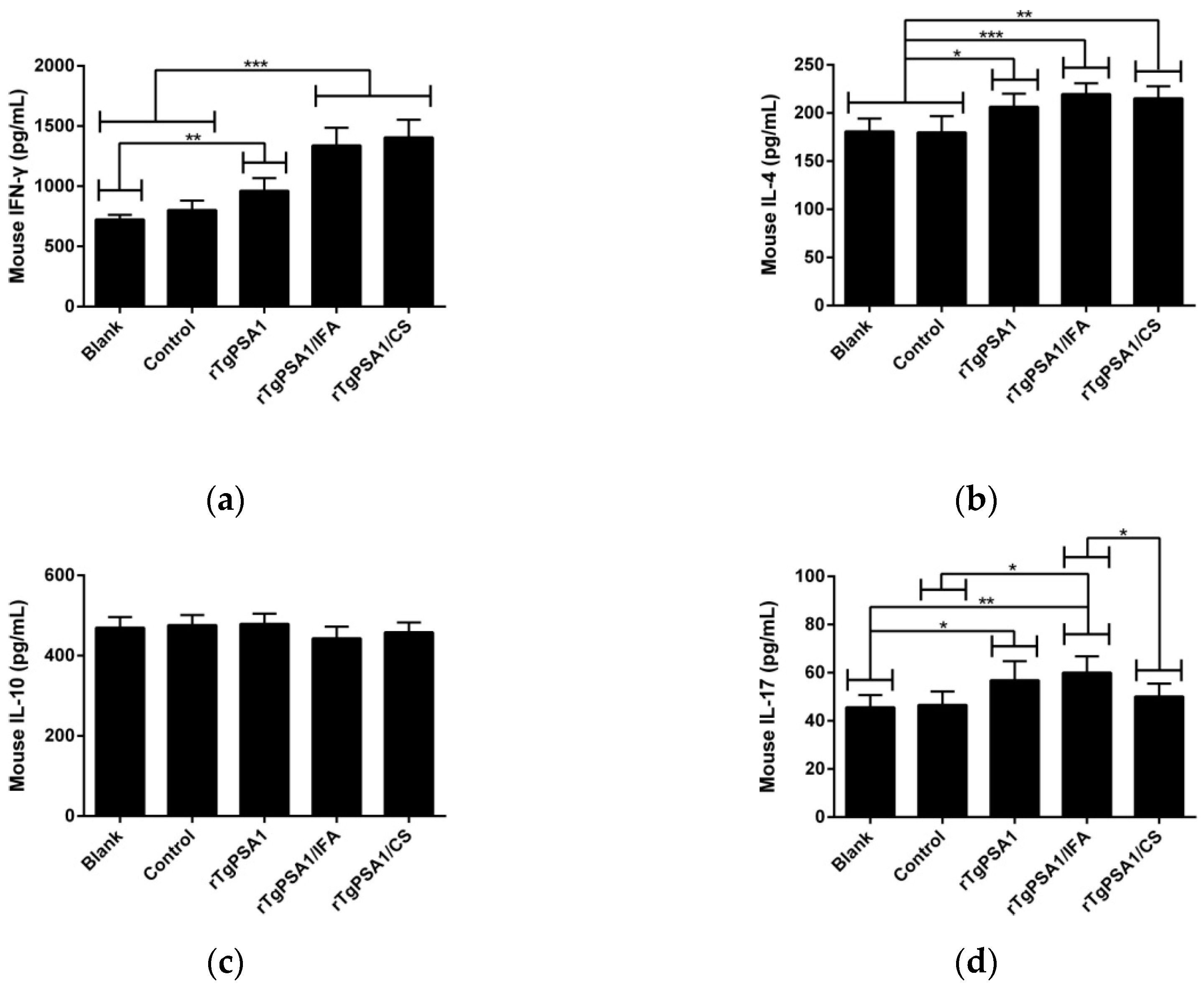
3.4. Antibody and Cytokine Production in Mice
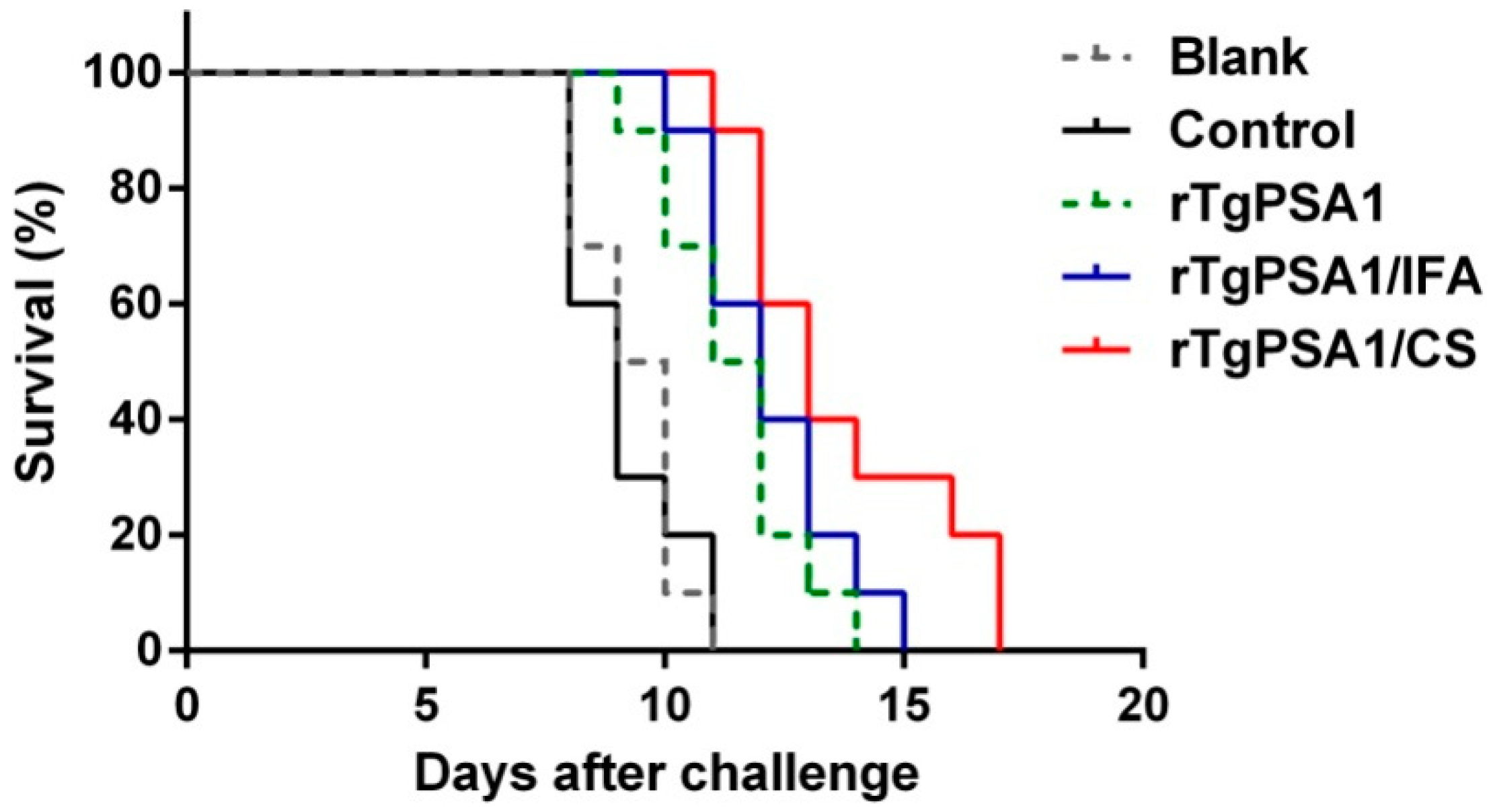
3.5. Immune Protection against Acute Toxoplasmosis
3.6. Comparison of Administration Routes
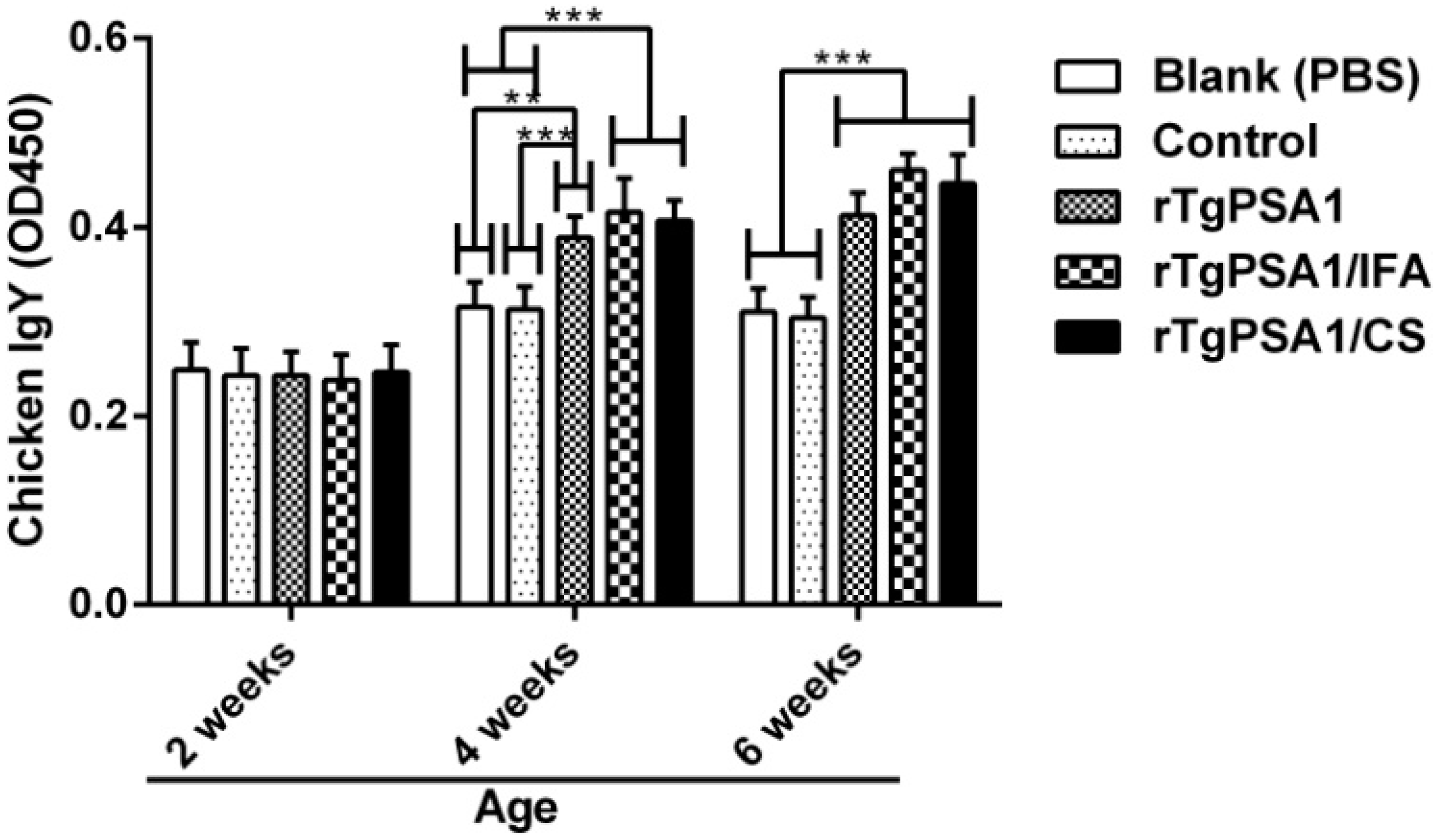
3.7. Antibody and Cytokine Production in Chickens
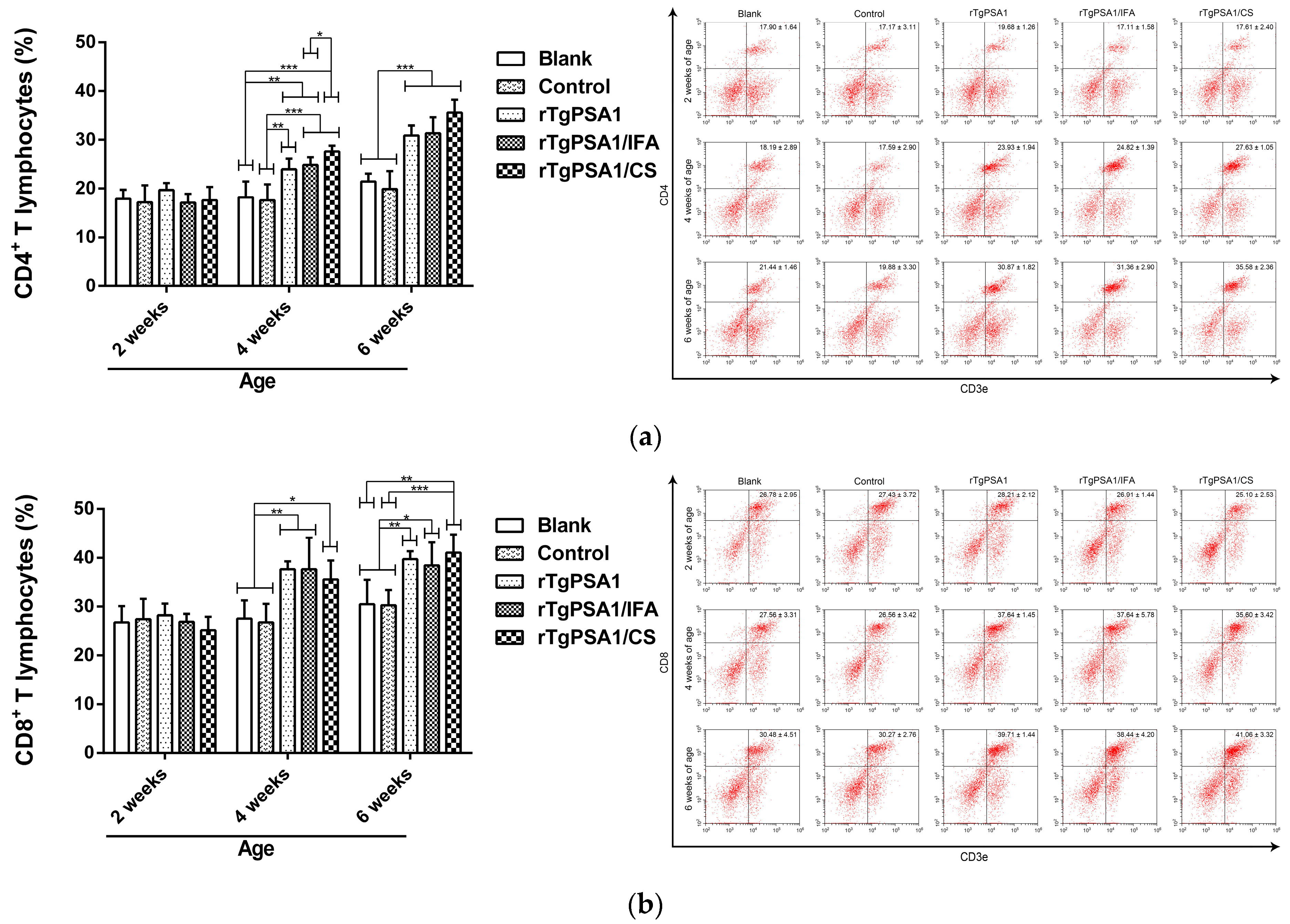
3.8. Analysis of Cellular Immune Response in Spleen Lymphocytes Separated from Chickens
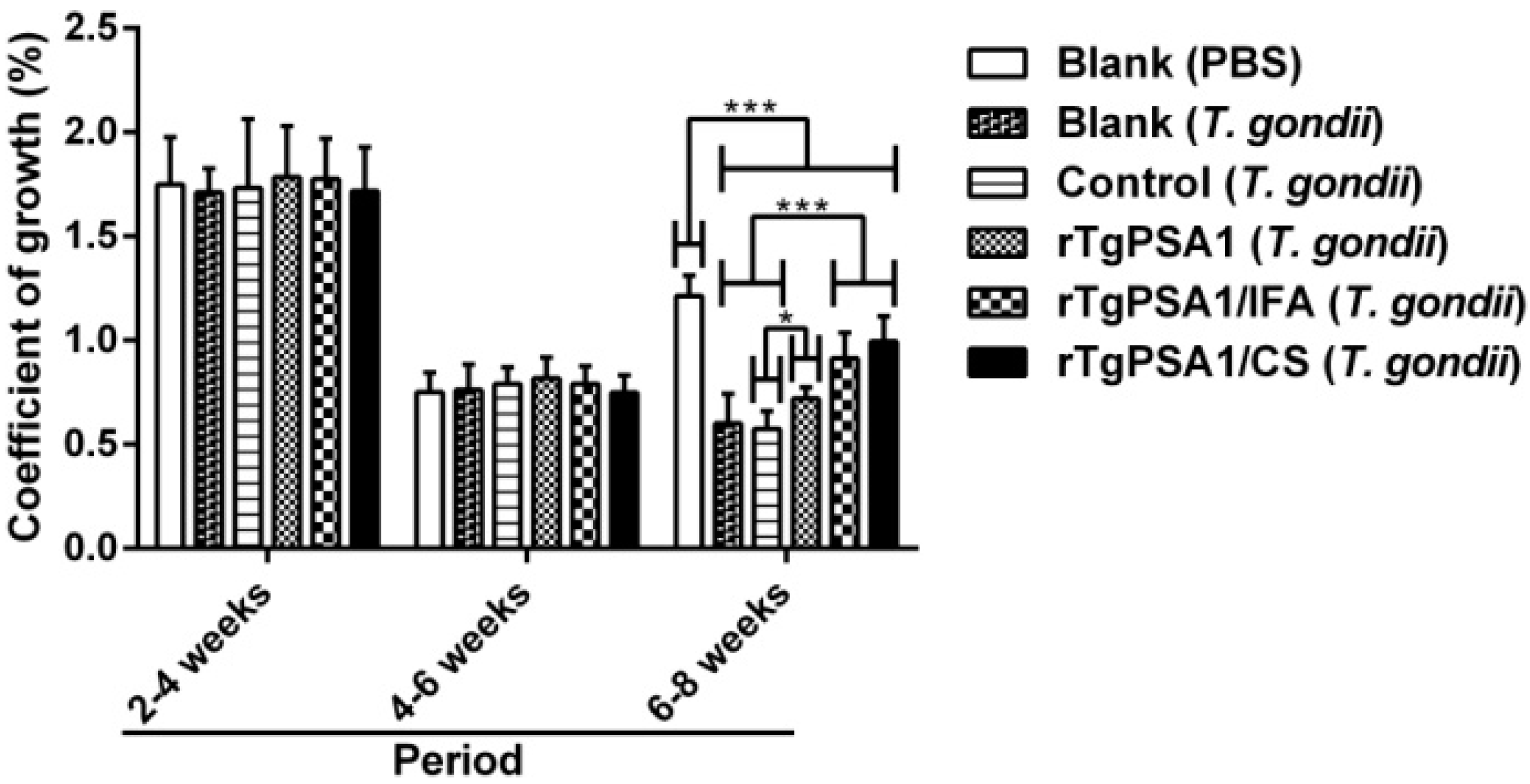
3.9. Growth Coefficient and Parasite Burden of Chickens
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Dubey, J.P.; Jones, J.L. Toxoplasma gondii infection in humans and animals in the United States. Int. J. Parasitol. 2008, 38, 1257–1278. [Google Scholar] [CrossRef]
- Hernandez-Cortazar, I.B.; Acosta-Viana, K.Y.; Guzman-Marin, E.; Ortega-Pacheco, A.; Segura-Correa, J.C.; Jimenez-Coello, M. Presence of Toxoplasma gondii in Drinking Water from an Endemic Region in Southern Mexico. Foodborne Pathog. Dis. 2017, 14, 288–292. [Google Scholar] [CrossRef] [PubMed]
- Arora, N.; Sadovsky, Y.; Dermody, T.S.; Coyne, C.B. Microbial Vertical Transmission during Human Pregnancy. Cell Host Microbe 2017, 21, 561–567. [Google Scholar] [CrossRef]
- Torgerson, P.R.; Mastroiacovo, P. The global burden of congenital toxoplasmosis: A systematic review. Bull. World Health Organ. 2013, 91, 501–508. [Google Scholar] [CrossRef]
- Barakat, A.M.A.; Ahmed, S.O.; Zaki, M.S.; El Fadaly, H.A.; Abd El-Razik, K.A.; El-Hariri, H.M.; Johar, D. New approach to differentiate primary from latent Toxoplasma gondii abortion through immunoglobulin and DNA interpretation. Microb. Pathog. 2018, 125, 66–71. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, C.; Maier, S.; Walker, R.A.; Rehrauer, H.; Joekel, D.E.; Winiger, R.R.; Basso, W.U.; Grigg, M.E.; Hehl, A.B.; Deplazes, P.; et al. An experimental genetically attenuated live vaccine to prevent transmission of Toxoplasma gondii by cats. Sci. Rep. 2019, 9, 1474. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Sun, X.; Yin, H.; Wang, T.; Li, Y.; Zhou, C.; Zhou, H.; He, S.; Cong, H. Chitosan Microsphere Used as an Effective System to Deliver a Linked Antigenic Peptides Vaccine Protect Mice Against Acute and Chronic Toxoplasmosis. Front. Cell Infect. Microbiol. 2018, 8, 163. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Kang, H.J.; Lee, D.H.; Kang, S.M.; Quan, F.S. Virus-like particle vaccines expressing Toxoplasma gondii rhoptry protein 18 and microneme protein 8 provide enhanced protection. Vaccine 2018, 36, 5692–5700. [Google Scholar] [CrossRef]
- Bastos, L.M.; Macedo, A.G., Jr.; Silva, M.V.; Santiago, F.M.; Ramos, E.L.; Santos, F.A.; Pirovani, C.P.; Goulart, L.R.; Mineo, T.W.; Mineo, J.R. Toxoplasma gondii-Derived Synthetic Peptides Containing B- and T-Cell Epitopes from GRA2 Protein Are Able to Enhance Mice Survival in a Model of Experimental Toxoplasmosis. Front. Cell Infect. Microbiol. 2016, 6, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nie, H.; Fang, R.; Xiong, B.Q.; Wang, L.X.; Hu, M.; Zhou, Y.Q.; Zhao, J.L. Immunogenicity and protective efficacy of two recombinant pseudorabies viruses expressing Toxoplasma gondii SAG1 and MIC3 proteins. Vet. Parasitol. 2011, 181, 215–221. [Google Scholar] [CrossRef]
- Buxton, D.; Innes, E.A. A commercial vaccine for ovine toxoplasmosis. Parasitology 1995, 110 (Suppl. 1), S11–S16. [Google Scholar] [CrossRef]
- AMCSF. Risk Profile in Relation to Toxoplasma in the Food Chain. 2012. Available online: http://www.food.gov.uk/sites/default/files/multimedia/pdfs/committee/acmsfrtaxopasm.pdf (accessed on 18 April 2021).
- Foroutan, M.; Zaki, L.; Ghaffarifar, F. Recent progress in microneme-based vaccines development against Toxoplasma gondii. Clin. Exp. Vaccine Res. 2018, 7, 93–103. [Google Scholar] [CrossRef]
- Agallou, M.; Athanasiou, E.; Koutsoni, O.; Dotsika, E.; Karagouni, E. Experimental Validation of Multi-Epitope Peptides Including Promising MHC Class I- and II-Restricted Epitopes of Four Known Leishmania infantum Proteins. Front. Immunol. 2014, 5, 268. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajissa, K.; Zakaria, R.; Suppian, R.; Mohamed, Z. Epitope-based vaccine as a universal vaccination strategy against Toxoplasma gondii infection: A mini-review. J. Adv. Vet. Anim. Res. 2019, 6, 174–182. [Google Scholar] [CrossRef]
- Liu, M.M.; Yuan, Z.G.; Peng, G.H.; Zhou, D.H.; He, X.H.; Yan, C.; Yin, C.C.; He, Y.; Lin, R.Q.; Song, H.Q.; et al. Toxoplasma gondii microneme protein 8 (MIC8) is a potential vaccine candidate against toxoplasmosis. Parasitol. Res. 2010, 106, 1079–1084. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Lu, S.; Tong, Q.; Kong, Q.; Lou, D. The virulence-related rhoptry protein 5 (ROP5) of Toxoplasma Gondii is a novel vaccine candidate against toxoplasmosis in mice. Vaccine 2013, 31, 4578–4584. [Google Scholar] [CrossRef] [PubMed]
- Zheng, B.; Lou, D.; Ding, J.; Zhuo, X.; Ding, H.; Kong, Q.; Lu, S. GRA24-Based DNA Vaccine Prolongs Survival in Mice Challenged With a Virulent Toxoplasma gondii Strain. Front. Immunol. 2019, 10, 418. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pagheh, A.S.; Sarvi, S.; Sharif, M.; Rezaei, F.; Ahmadpour, E.; Dodangeh, S.; Omidian, Z.; Hassannia, H.; Mehrzadi, S.; Daryani, A. Toxoplasma gondii surface antigen 1 (SAG1) as a potential candidate to develop vaccine against toxoplasmosis: A systematic review. Comp. Immunol. Microbiol. Infect. Dis. 2020, 69, 101414. [Google Scholar] [CrossRef] [PubMed]
- Zhang, N.Z.; Wang, M.; Xu, Y.; Petersen, E.; Zhu, X.Q. Recent advances in developing vaccines against Toxoplasma gondii: An update. Expert Rev. Vaccines 2015, 14, 1609–1621. [Google Scholar] [CrossRef]
- Montazeri, M.; Sharif, M.; Sarvi, S.; Mehrzadi, S.; Ahmadpour, E.; Daryani, A. A Systematic Review of In vitro and In vivo Activities of Anti-Toxoplasma Drugs and Compounds (2006-2016). Front. Microbiol. 2017, 8, 25. [Google Scholar] [CrossRef] [Green Version]
- Fregno, I.; Molinari, M. Proteasomal and lysosomal clearance of faulty secretory proteins: ER-associated degradation (ERAD) and ER-to-lysosome-associated degradation (ERLAD) pathways. Crit Rev. Biochem. Mol. Biol. 2019, 54, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Ciechanover, A. The ubiquitin proteolytic system: From a vague idea, through basic mechanisms, and onto human diseases and drug targeting. Neurology 2006, 66, S7–S19. [Google Scholar] [CrossRef] [PubMed]
- Saeki, Y.; Tanaka, K. Assembly and function of the proteasome. Methods Mol. Biol. 2012, 832, 315–337. [Google Scholar] [CrossRef] [PubMed]
- Pagano, M. Cell cycle regulation by the ubiquitin pathway. FASEB J. 1997, 11, 1067–1075. [Google Scholar] [CrossRef] [PubMed]
- Pahl, H.L.; Baeuerle, P.A. Control of gene expression by proteolysis. Curr. Opin. Cell Biol. 1996, 8, 340–347. [Google Scholar] [CrossRef]
- Piccinini, M.; Mostert, M.; Rinaudo, M.T. Proteasomes as drug targets. Curr. Drug Targets 2003, 4, 657–671. [Google Scholar] [CrossRef] [PubMed]
- Munoz, C.; San Francisco, J.; Gutierrez, B.; Gonzalez, J. Role of the Ubiquitin-Proteasome Systems in the Biology and Virulence of Protozoan Parasites. Biomed. Res. Int. 2015, 2015, 141526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paugam, A.; Bulteau, A.L.; Dupouy-Camet, J.; Creuzet, C.; Friguet, B. Characterization and role of protozoan parasite proteasomes. Trends Parasitol. 2003, 19, 55–59. [Google Scholar] [CrossRef]
- Ward, W.; Alvarado, L.; Rawlings, N.D.; Engel, J.C.; Franklin, C.; McKerrow, J.H. A primitive enzyme for a primitive cell: The protease required for excystation of Giardia. Cell 1997, 89, 437–444. [Google Scholar] [CrossRef] [Green Version]
- Makioka, A.; Kumagai, M.; Kobayashi, S.; Takeuchi, T. Entamoeba invadens: Cysteine protease inhibitors block excystation and metacystic development. Exp. Parasitol. 2005, 109, 27–32. [Google Scholar] [CrossRef]
- Paugam, A.; Creuzet, C.; Dupouy-Camet, J.; Roisin, M.P. Evidence for the existence of a proteasome in Toxoplasma gondii: Intracellular localization and specific peptidase activities. Parasite 2001, 8, 267–273. [Google Scholar] [CrossRef] [Green Version]
- Shaw, M.K.; He, C.Y.; Roos, D.S.; Tilney, L.G. Proteasome inhibitors block intracellular growth and replication of Toxoplasma gondii. Parasitology 2000, 121 Pt 1, 35–47. [Google Scholar] [CrossRef]
- Paugam, A.; Creuzet, C.; Dupouy-Camet, J.; Roisin, P. In vitro effects of gliotoxin, a natural proteasome inhibitor, on the infectivity and proteolytic activity of Toxoplasma gondii. Parasitol. Res. 2002, 88, 785–787. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; O’Hagan, D.T. Recent advances in veterinary vaccine adjuvants. Int. J. Parasitol. 2003, 33, 469–478. [Google Scholar] [CrossRef]
- Freund, J.; Casals, J.; Hosmer, E.P. Sensitization and Antibody Formation after Injection of Tubercle Bacilli and Paraffin Oil. Exp. Biol. Med. 1937, 37, 509–513. [Google Scholar] [CrossRef] [Green Version]
- Stuart-Harris, C.H. Adjuvant influenza vaccines. Bull. World Health Organ. 1969, 41, 617–621. [Google Scholar]
- Freund, J.; Thomson, K.J. A simple, rapid technic of preparing water-in-oil emulsions of penicillin, drugs and biologics. Science 1945, 101, 468–469. [Google Scholar] [CrossRef] [PubMed]
- van der Lubben, I.M.; Verhoef, J.C.; van Aelst, A.C.; Borchard, G.; Junginger, H.E. Chitosan microparticles for oral vaccination: Preparation, characterization and preliminary in vivo uptake studies in murine Peyer’s patches. Biomaterials 2001, 22, 687–694. [Google Scholar] [CrossRef]
- Wang, W.; Meng, Q.; Li, Q.; Liu, J.; Zhou, M.; Jin, Z.; Zhao, K. Chitosan Derivatives and Their Application in Biomedicine. Int. J. Mol. Sci. 2020, 21, 487. [Google Scholar] [CrossRef] [Green Version]
- Illum, L. Chitosan and its use as a pharmaceutical excipient. Pharm. Res. 1998, 15, 1326–1331. [Google Scholar] [CrossRef] [PubMed]
- Wedmore, I.; McManus, J.G.; Pusateri, A.E.; Holcomb, J.B. A special report on the chitosan-based hemostatic dressing: Experience in current combat operations. J. Trauma 2006, 60, 655–658. [Google Scholar] [CrossRef] [Green Version]
- Cheraghipour, K.; Masoori, L.; Ezzatkhah, F.; Salimikia, I.; Amiri, S.; Makenali, A.S.; Taherpour, F.; Mahmoudvand, H. Effect of chitosan on Toxoplasma gondii infection: A systematic review. Parasite Epidemiol. Control. 2020, 11, e00189. [Google Scholar] [CrossRef]
- Wang, J.J.; Zeng, Z.W.; Xiao, R.Z.; Xie, T.A.; Zhou, G.L.; Zhan, X.R.; Wang, S.L. Recent advances of chitosan nanoparticles as drug carriers. Int. J. Nanomed. 2011, 6, 765–774. [Google Scholar] [CrossRef] [Green Version]
- Hagras, N.A.; Allam, A.F.; Farag, H.F.; Osman, M.M.; Shalaby, T.I.; Fawzy Hussein Mogahed, N.M.; Tolba, M.M.; Shehab, A.Y. Successful treatment of acute experimental toxoplasmosis by spiramycin-loaded chitosan nanoparticles. Exp. Parasitol. 2019, 204, 107717. [Google Scholar] [CrossRef]
- El Temsahy, M.M.; El Kerdany, E.D.; Eissa, M.M.; Shalaby, T.I.; Talaat, I.M.; Mogahed, N.M. The effect of chitosan nanospheres on the immunogenicity of Toxoplasma lysate vaccine in mice. J. Parasit. Dis. 2016, 40, 611–626. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yu, Z.Q.; Zhou, T.Y.; Luo, Y.X.; Dong, L.; Li, C.J.; Liu, J.L.; Luo, J.X.; Yan, R.F.; Xu, L.X.; Song, X.K.; et al. Modulation Effects of Toxoplasma gondii Histone H2A1 on Murine Macrophages and Encapsulation with Polymer as a Vaccine Candidate. Vaccines 2020, 8, 731. [Google Scholar] [CrossRef]
- Bivas-Benita, M.; Laloup, M.; Versteyhe, S.; Dewit, J.; De Braekeleer, J.; Jongert, E.; Borchard, G. Generation of Toxoplasma gondii GRA1 protein and DNA vaccine loaded chitosan particles: Preparation, characterization, and preliminary in vivo studies. Int. J. Pharm. 2003, 266, 17–27. [Google Scholar] [CrossRef]
- Islam, N.; Ferro, V. Recent advances in chitosan-based nanoparticulate pulmonary drug delivery. Nanoscale 2016, 8, 14341–14358. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Zhao, G.; Wang, W.; Xie, Q.; Zhang, M.; Yuan, C.; Hassan, I.A.; Liu, X.; Xu, L.; Yan, R.; et al. Pathogenicity of two Toxoplasma gondii strains in chickens of different ages infected via intraperitoneal injection. Avian Pathol. 2014, 43, 91–95. [Google Scholar] [CrossRef] [Green Version]
- Khan, A.; Grigg, M.E. Toxoplasma gondii: Laboratory Maintenance and Growth. Curr. Protoc. Microbiol. 2017, 44, 20C-1. [Google Scholar] [CrossRef] [Green Version]
- Hassan, I.A.; Wang, S.; Xu, L.; Yan, R.; Song, X.; Li, X. Immunoglobulin and cytokine changes induced following immunization with a DNA vaccine encoding Toxoplasma gondii selenium-dependent glutathione reductase protein. Exp. Parasitol. 2014, 146, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Fan, W.; Yan, W.; Xu, Z.; Ni, H. Formation mechanism of monodisperse, low molecular weight chitosan nanoparticles by ionic gelation technique. Colloids Surf. B Biointerfaces 2012, 90, 21–27. [Google Scholar] [CrossRef] [PubMed]
- Skop, N.B.; Calderon, F.; Levison, S.W.; Gandhi, C.D.; Cho, C.H. Heparin crosslinked chitosan microspheres for the delivery of neural stem cells and growth factors for central nervous system repair. Acta Biomater. 2013, 9, 6834–6843. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.L.; Elsheikha, H.M.; Zhu, W.N.; Chen, K.; Li, T.T.; Yue, D.M.; Zhang, X.X.; Huang, S.Y.; Zhu, X.Q. Immunization with Toxoplasma gondii GRA17 Deletion Mutant Induces Partial Protection and Survival in Challenged Mice. Front. Immunol. 2017, 8, 730. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tang, X.M.; Yin, G.W.; Qin, M.; Tao, G.R.; Suo, J.X.; Liu, X.Y.; Suo, X. Transgenic Eimeria tenella as a vaccine vehicle: Expressing TgSAG1 elicits protective immunity against Toxoplasma gondii infections in chickens and mice. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [Green Version]
- Sasai, K.; Aita, M.; Lillehoj, H.S.; Miyamoto, T.; Fukata, T.; Baba, E. Dynamics of lymphocyte subpopulation changes in the cecal tonsils of chickens infected with Salmonella enteritidis. Vet. Microbiol. 2000, 74, 345–351. [Google Scholar] [CrossRef]
- Homan, W.L.; Vercammen, M.; De Braekeleer, J.; Verschueren, H. Identification of a 200-to 300-fold repetitive 529 bp DNA fragment in Toxoplasma gondii, and its use for diagnostic and quantitative PCR. Int. J. Parasitol. 2000, 30, 69–75. [Google Scholar] [CrossRef]
- Skwarczynski, M.; Toth, I. Peptide-based synthetic vaccines. Chem. Sci. 2016, 7, 842–854. [Google Scholar] [CrossRef] [Green Version]
- Pohlmann, A.R.; Fonseca, F.N.; Paese, K.; Detoni, C.B.; Coradini, K.; Beck, R.C.; Guterres, S.S. Poly(-caprolactone) microcapsules and nanocapsules in drug delivery. Expert Opin. Drug Deliv. 2013, 10, 623–638. [Google Scholar] [CrossRef]
- Chuang, S.C.; Ko, J.C.; Chen, C.P.; Du, J.T.; Yang, C.D. Induction of long-lasting protective immunity against Toxoplasma gondii in BALB/c mice by recombinant surface antigen 1 protein encapsulated in poly (lactide-co-glycolide) microparticles. Parasit. Vectors 2013, 6, 34. [Google Scholar] [CrossRef] [Green Version]
- Xu, Y.; Zhang, N.Z.; Wang, M.; Dong, H.; Feng, S.Y.; Guo, H.C.; Zhu, X.Q. A long-lasting protective immunity against chronic toxoplasmosis in mice induced by recombinant rhoptry proteins encapsulated in poly (lactide-co-glycolide) microparticles. Parasitol. Res. 2015, 114, 4195–4203. [Google Scholar] [CrossRef]
- Lee, D.H.; Lee, S.H.; Kim, A.R.; Quan, F.S. Virus-Like Nanoparticle Vaccine Confers Protection against Toxoplasma gondii. PLoS ONE 2016, 11, e0161231. [Google Scholar] [CrossRef] [Green Version]
- Ducournau, C.; Moire, N.; Carpentier, R.; Cantin, P.; Herkt, C.; Lantier, I.; Betbeder, D.; Dimier-Poisson, I. Effective Nanoparticle-Based Nasal Vaccine Against Latent and Congenital Toxoplasmosis in Sheep. Front. Immunol. 2020, 11, 2183. [Google Scholar] [CrossRef] [PubMed]
- Zaharoff, D.A.; Rogers, C.J.; Hance, K.W.; Schlom, J.; Greiner, J.W. Chitosan solution enhances both humoral and cell-mediated immune responses to subcutaneous vaccination. Vaccine 2007, 25, 2085–2094. [Google Scholar] [CrossRef] [Green Version]
- Sinani, G.; Sessevmez, M.; Gok, M.K.; Ozgumus, S.; Alpar, H.O.; Cevher, E. Modified chitosan-based nanoadjuvants enhance immunogenicity of protein antigens after mucosal vaccination. Int. J. Pharm. 2019, 569, 118592. [Google Scholar] [CrossRef]
- Lin, Y.C.; Lou, P.J.; Young, T.H. Chitosan as an adjuvant-like substrate for dendritic cell culture to enhance antitumor effects. Biomaterials 2014, 35, 8867–8875. [Google Scholar] [CrossRef]
- Jiang, L.Q.; Wang, T.Y.; Webster, T.J.; Duan, H.J.; Qiu, J.Y.; Zhao, Z.M.; Yin, X.X.; Zheng, C.L. Intracellular disposition of chitosan nanoparticles in macrophages: Intracellular uptake, exocytosis, and intercellular transport. Int. J. Nanomed. 2017, 12, 6383–6398. [Google Scholar] [CrossRef] [Green Version]
- Wang, F.Z.; Zhang, M.W.; Zhang, D.S.; Huang, Y.; Chen, L.; Jiang, S.M.; Shi, K.; Li, R. Preparation, optimization, and characterization of chitosan-coated solid lipid nanoparticles for ocular drug delivery. J. Biomed. Res. 2018, 32, 411–423. [Google Scholar] [CrossRef]
- He, P.; Davis, S.S.; Illum, L. Chitosan microspheres prepared by spray drying. Int. J. Pharm. 1999, 187, 53–65. [Google Scholar] [CrossRef]
- López, R.G.; Pineda, M.G.; Hurtado, G.; León, R.D.; Fernández, S.; Saade, H.; Bueno, D. Chitosan-coated magnetic nanoparticles prepared in one step by reverse microemulsion precipitation. Int. J. Mol. Sci. 2013, 14, 19636–19650. [Google Scholar] [CrossRef] [Green Version]
- Bachmann, M.F.; Jennings, G.T. Vaccine delivery: A matter of size, geometry, kinetics and molecular patterns. Nat. Rev. Immunol. 2010, 10, 787–796. [Google Scholar] [CrossRef]
- Koppolu, B.P.; Smith, S.G.; Ravindranathan, S.; Jayanthi, S.; Suresh Kumar, T.K.; Zaharoff, D.A. Controlling chitosan-based encapsulation for protein and vaccine delivery. Biomaterials 2014, 35, 4382–4389. [Google Scholar] [CrossRef] [Green Version]
- Correa, D.; Canedo-Solares, I.; Ortiz-Alegria, L.B.; Caballero-Ortega, H.; Rico-Torres, C.P. Congenital and acquired toxoplasmosis: Diversity and role of antibodies in different compartments of the host. Parasite Immunol. 2007, 29, 651–660. [Google Scholar] [CrossRef] [PubMed]
- Sayles, P.C.; Gibson, G.W.; Johnson, L.L. B cells are essential for vaccination-induced resistance to virulent Toxoplasma gondii. Infect. Immun. 2000, 68, 1026–1033. [Google Scholar] [CrossRef] [Green Version]
- Naeem, H.; Sana, M.; Islam, S.; Khan, M.; Riaz, F.; Zafar, Z.; Akbar, H.; Shehzad, W.; Rashid, I. Induction of Th1 type-oriented humoral response through intranasal immunization of mice with SAG1-Toxoplasma gondii polymeric nanospheres. Artif. Cell Nanomed. B 2018, 46, 1025–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, B.; Benson, A.; Kuzmich, L.; DeFranco, A.L.; Yarovinsky, F. Critical coordination of innate immune defense against Toxoplasma gondii by dendritic cells responding via their Toll-like receptors. Proc. Natl. Acad. Sci. USA 2011, 108, 278–283. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pifer, R.; Yarovinsky, F. Innate responses to Toxoplasma gondii in mice and humans. Trends Parasitol. 2011, 27, 388–393. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wagner, A.; Schabussova, I.; Ruttkowski, B.; Peschke, R.; Kur, J.; Kundi, M.; Joachim, A.; Wiedermann, U. Prime-boost vaccination with toxoplasma lysate antigen, but not with a mixture of recombinant protein antigens, leads to reduction of brain cyst formation in BALB/c mice. PLoS ONE 2015, 10, e0126334. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ching, X.T.; Fong, M.Y.; Lau, Y.L. Evaluation of Immunoprotection Conferred by the Subunit Vaccines of GRA2 and GRA5 against Acute Toxoplasmosis in BALB/c Mice. Front. Microbiol. 2016, 7, 609. [Google Scholar] [CrossRef] [PubMed]
- Bessieres, M.H.; Swierczynski, B.; Cassaing, S.; Miedouge, M.; Olle, P.; Seguela, J.P.; Pipy, B. Role of IFN-gamma, TNF-alpha, IL4 and IL10 in the regulation of experimental Toxoplasma gondii infection. J. Eukaryot Microbiol. 1997, 44, 87s. [Google Scholar] [CrossRef]
- Pinzan, C.F.; Sardinha-Silva, A.; Almeida, F.; Lai, L.; Lopes, C.D.; Lourenco, E.V.; Panunto-Castelo, A.; Matthews, S.; Roque-Barreira, M.C. Vaccination with Recombinant Microneme Proteins Confers Protection against Experimental Toxoplasmosis in Mice. PLoS ONE 2015, 10, e0143087. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef]
- Mesquita Júnior, D.; Araújo, J.A.; Catelan, T.T.; Souza, A.W.; Cruvinel Wde, M.; Andrade, L.E.; Silva, N.P. Immune system—Part II: Basis of the immunological response mediated by T and B lymphocytes. Rev. Bras. Reumatol. 2010, 50, 552–580. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kelly, M.N.; Kolls, J.K.; Happel, K.; Schwartzman, J.D.; Schwarzenberger, P.; Combe, C.; Moretto, M.; Khan, I.A. Interleukin-17/interleukin-17 receptor-mediated signaling is important for generation of an optimal polymorphonuclear response against Toxoplasma gondii infection. Infect. Immun. 2005, 73, 617–621. [Google Scholar] [CrossRef] [Green Version]
- Gazzinelli, R.; Xu, Y.; Hieny, S.; Cheever, A.; Sher, A. Simultaneous depletion of CD4+ and CD8+ T lymphocytes is required to reactivate chronic infection with Toxoplasma gondii. J. Immunol. 1992, 149, 175–180. [Google Scholar] [PubMed]
- Zheng, B.; Ding, J.; Chen, X.; Yu, H.; Lou, D.; Tong, Q.; Kong, Q.; Lu, S. Immuno-Efficacy of a T. gondii Secreted Protein with an Altered Thrombospondin Repeat (TgSPATR) As a Novel DNA Vaccine Candidate against Acute Toxoplasmosis in BALB/c Mice. Front. Microbiol. 2017, 8, 216. [Google Scholar] [CrossRef]
- Wang, T.; Yin, H.; Li, Y.; Zhao, L.; Sun, X.; Cong, H. Vaccination with recombinant adenovirus expressing multi-stage antigens of Toxoplasma gondii by the mucosal route induces higher systemic cellular and local mucosal immune responses than with other vaccination routes. Parasite 2017, 24, 12. [Google Scholar] [CrossRef] [Green Version]
- Courret, N.; Darche, S.; Sonigo, P.; Milon, G.; Buzoni-Gatel, D.; Tardieux, I. CD11c- and CD11b-expressing mouse leukocytes transport single Toxoplasma gondii tachyzoites to the brain. Blood 2006, 107, 309–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wohlfert, E.A.; Blader, I.J.; Wilson, E.H. Brains and Brawn: Toxoplasma Infections of the Central Nervous System and Skeletal Muscle. Trends Parasitol. 2017, 33, 519–531. [Google Scholar] [CrossRef] [PubMed]
- Alves, B.F.; Oliveira, S.; Soares, H.S.; Pena, H.F.J.; Conte-Junior, C.A.; Gennari, S.M. Isolation of viable Toxoplasma gondii from organs and Brazilian commercial meat cuts of experimentally infected pigs. Parasitol. Res. 2019, 118, 1331–1335. [Google Scholar] [CrossRef]
- Dubey, J.R.; Bhaiyat, M.I.; de Allie, C.; Macpherson, C.N.; Sharma, R.N.; Sreekumar, C.; Vianna, M.C.; Shen, S.K.; Kwok, O.C.; Miska, K.B.; et al. Isolation, tissue distribution, and molecular characterization of Toxoplasma gondii from chickens in Grenada, West Indies. J. Parasitol. 2005, 91, 557–560. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | Treatment (Each Mouse) | Time for Immunization | Time for Infection | Infection Dose (Each Mouse) |
|---|---|---|---|---|
| Blank | Equal volume of PBS | Week 0 and 2 | Week 4 | 200 tachyzoites |
| Control | 100 μg pET32a vector protein | |||
| rTgPSA1 | 100 μg rTgPSA1 | |||
| rTgPSA1/IFA | rTgPSA1/IFA emulsion containing 100 μg rTgPSA1 | |||
| rTgPSA1/CS | rTgPSA1/CS nanospheres containing 100 μg rTgPSA1 | |||
| Blank | Equal volume of PBS |
| Group | Treatment (Each Chicken) | Time for Immunization | Time for Infection | Infection Dose (Each Chicken) |
|---|---|---|---|---|
| Blank (PBS) | Equal volume of PBS | At 2 and 4 weeks old | At 6 weeks old | Equal volume of PBS (0 tachyzoite) |
| Blank (T. gondii) | Equal volume of PBS | 1 × 107 tachyzoites | ||
| Control | 200 μg pET32a vector protein | |||
| rTgPSA1 | 200 μg rTgPSA1 | |||
| rTgPSA1/IFA | rTgPSA1/IFA emulsion containing 200 μg rTgPSA1 | |||
| rTgPSA1/CS | rTgPSA1/CS nanospheres containing 200 μg rTgPSA1 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Ding, W.; Aleem, M.T.; Su, J.; Liu, J.; Luo, J.; Yan, R.; Xu, L.; Song, X.; Li, X. Toxoplasma gondii Proteasome Subunit Alpha Type 1 with Chitosan: A Promising Alternative to Traditional Adjuvant. Pharmaceutics 2021, 13, 752. https://doi.org/10.3390/pharmaceutics13050752
Yu Z, Ding W, Aleem MT, Su J, Liu J, Luo J, Yan R, Xu L, Song X, Li X. Toxoplasma gondii Proteasome Subunit Alpha Type 1 with Chitosan: A Promising Alternative to Traditional Adjuvant. Pharmaceutics. 2021; 13(5):752. https://doi.org/10.3390/pharmaceutics13050752
Chicago/Turabian StyleYu, Zhengqing, Wenxi Ding, Muhammad Tahir Aleem, Junzhi Su, Junlong Liu, Jianxun Luo, Ruofeng Yan, Lixin Xu, Xiaokai Song, and Xiangrui Li. 2021. "Toxoplasma gondii Proteasome Subunit Alpha Type 1 with Chitosan: A Promising Alternative to Traditional Adjuvant" Pharmaceutics 13, no. 5: 752. https://doi.org/10.3390/pharmaceutics13050752